Abstract
Bean production is at risk due to climate change, declining water resources, and inadequate crop management. To address these challenges, dynamic models that predict crop growth and development can be used as fundamental tools to generate basic and applied knowledge such as production management and decision support. This study aimed to calibrate and evaluate the SIMPLE model under irrigation conditions for a semi-arid region in north-central Mexico and to simulate thermal time, biomass (Bio), and grain yield (GY) of common beans cv. ‘Pinto Saltillo’ using experimental data from four crop evapotranspiration treatments (ETct) (I50, I75, I100, and I125) applied during the 2020 and 2021 growing seasons. Both experiments were conducted in a randomized complete block design with three replicates. Model calibration was carried out by posing and solving an optimization problem with the differential-evolution algorithm with 2020 experimental data, while the evaluation was performed with 2021 experimental data. For Bio, calibration values had a root-mean-square error and Nash and Sutcliffe’s efficiency of <0.58 t ha−1 and >0.93, respectively, while the corresponding evaluation values were <1.80 t ha−1 and >0.89, respectively. The I50 and I100 ETct had better fit for calibration, while I50 and I75 had better fit in the evaluation. On average, the model fitted for the predicted GY values had estimation errors of 37% and 22% for the calibration and evaluation procedures, respectively. Therefore, an empirical model was proposed to estimate the harvest index (HI), which produced, on average, a relative error of 6.9% for the bean-GY estimation. The SIMPLE model was able to predict bean biomass under irrigated conditions for these semi-arid regions of Mexico. Also, the use of both crop Bio and transpiration simulated by the SIMPLE model to calculate the HI significantly improved GY prediction under ETct. However, the harvest index needs to be validated under other irrigation levels and field experiments in different locations to strengthen the proposed model and design different GY scenarios under water restrictions for irrigation due to climate change.
1. Introduction
Due to its high protein content, the common bean (Phaseolus vulgaris L.) is the most widely produced and consumed legume in the world. Mexico ranks ninth in world production [1]. Annually, its per capita consumption is 8 kg [2]. Zacatecas state, located in north-central Mexico is the main producer of this legume grown in semi-arid and arid agricultural lands [3]. This climate presents droughts, irregular episodes of rainfall, and high evapotranspiration demand during the crop growing season, where the water supply, via irrigation, for common bean cultivation is mandatory. [4]. In Zacatecas, more than 64,000 ha are annually bean-cultivated under irrigation yielding between 1.7 and 2.4 t ha−1 [5], some of the lowest yields worldwide. In addition to adverse climatic conditions, low yields are also accompanied by degraded soils, inadequate crop water management, and aquifer overexploitation. However, yields can be improved by understanding crop management within the soil-plant-atmosphere continuum, where crop growth and development models become relevant. Mathematical models are effective tools for describing and understanding complex systems, but they are also used for crop management, generation of government policies, evaluation, and adaptation to climate change impacts [6]. Dynamic mathematical models are composed of a set of first-order ordinary differential equations or difference equations [7]. These equations have a set of physiological coefficients (e.g., radiation use efficiency), which are difficult to measure directly [8] and estimate [9].
On the other hand, the models used to simulate bean growth and development of the DSSAT family [10] highlight the BEANGRO model [11], GenoGro [12], CROPGRO [13], and the Wageningen family models such as the models WOFOST [14], SODCOM [15], SUCROS [16], and DIACROS [17]. There are other models, such as the Cereal–Legume model, which simulates the growth of beans intercropped with corn, the SSM–Legumes model [18], and the Daisy model [19]. Nevertheless, Yuan et al. [20] and Zhao et al. [21] have developed bean models that require only a few parameters, and other models for irrigation management in beans (SWB model [22]; SALTMED [23]; AquaCrop [24].
Also, most of the models used to simulate crop growth and development require a large number of parameters which are difficult to obtain as a result of the wide diversity of agroecological environments, crops, and varieties. It is here that the importance of models using few parameters is highlighted [25]. The Simple Simulation Model (SSM) developed by Soltani and Sinclair [18] involves five state variables and twenty-seven parameters. The HORTSYST model includes seven state variables and twenty-four parameters [26]. Recently, Zhao et al. [21] proposed the SIMPLE model with two state variables and fourteen parameters for field crops. The SIMPLE model was developed as a generic model based on intercepted radiation and simulates thermal time (TT °C d) and biomass (Bio t ha−1), but also calculates grain yield (GY t ha−1) [21].
The SIMPLE model was adopted for soybean biomass and yield estimations under climate change scenarios of air temperature and atmospheric CO2 [27]. This model has been applied to simulate maize biomass and yield for Vietnam’s autumn-winter and winter-spring growing seasons [28] and to predict flax biomass and yield under four arid and semi-arid scenarios in China. So, the SIMPLE model can be extended to many crop species by adding variable modules such as nutrient dynamics, water stress, temperature stress, or pests [29]. Therefore, the low GY, the economic importance of bean crops, and the lack of growth and development models applied to this species, motivated this research. This study aimed to calibrate and evaluate the SIMPLE model under irrigation conditions for a semi-arid region in north-central Mexico and to simulate the TT, Bio, and GY of common beans cv. ‘Pinto Saltillo’ using experimental data from the 2020 and 2021 growing seasons. So, in the face of the global warming impact on the annual crops, we hypothesized that the SIMPLE model would be useful for simulating bean biomass and yield for the semi-arid agroecological scenarios of north-central Mexico. These kinds of models are particularly important for these growing areas because they only use a few parameters for crop modeling.
2. Materials and Methods
2.1. Description of the Study Area
The experiments were conducted from April to August for the 2020 and 2021 growing seasons at the Zacatecas Experimental Station of the National Institute of Forestry, Agriculture and Livestock Research (INIFAP) located in Calera de Víctor Rosales, Zacatecas, Mexico (22°54′ N; 102°39′ W, elevation 2197 m). The experimental site has a mean annual temperature of 14.6 °C and a mean annual rainfall of 416 mm, 75% of which occurs between July and October [30]. The mean annual pan evaporation is 1609 mm. Before setting up the study, the experimental site was bleached with oatmeal during the winter of 2019–2020 (northern hemisphere). Afterwards, a physicochemical soil analysis was carried out in the soil-water laboratory of INIFAP. The experimental site’s soil is clay loam with a saturation point, field capacity (FC), permanent wilting point (PWP), and average bulk density of 0.48 m3 m−3, 0.29 m3 m−3, 0.14 m3 m−3, and 1.1 g cm−3, respectively. The grain size distribution is 35%, 28%, and 37% for sand, silt, and clay, respectively. The content of N is 3.5 mg kg−1, P is 7.8 mg kg−1 and K is 31 mg kg−1 in the first 40 cm of soil depth. The organic matter content of the soil is low (1.3%) and has a pH of 8.2.
2.2. Genetic Material and Crop Management
‘Pinto Saltillo’ is a common bean variety with indeterminate growth. Seeds were sown on April 16 and April 21 in 2020 and 2021, respectively. The distance between furrows was 0.76 m, with 0.1 m between plants. The planting density was 131, 578 plants ha−1. The planting depth was between 0.06 and 0.07 m. Mineral fertilization consisted of N and P only. The sources of N and P were urea (40 units of N) and mono ammonium phosphate (60 units of P), respectively, which were fractionated during the crop cycle in 20–20, 10–20, and 10–20 units of N and P, respectively. Fertilization doses were applied at 26, 36, and 47 days after sowing (DAS) in 2020 and at 17, 29, and 44 DAS in 2021. Weed, disease, and pest controls were performed as required.
2.3. Irrigation Treatments and Experimental Design
The experiments were conducted in two consecutive growing seasons. They consisted of four irrigation treatments according to the atmospheric demand: 50, 75, 100 (as control), and 125% of crop evapotranspiration (ETc).
Previous to sowing, a gravimetric soil sampling was carried out to determine the residual soil moisture content (θs), to apply the initial irrigation depth (IID) at FC in all irrigation treatments (Equation (1)) [31]. Subsequently, the irrigation schedule consisted of water supply twice a week based on the daily data of the reference evapotranspiration (ETo) obtained from an automated weather station placed 1.5 km from the experiment, and 75% of the effective rainfall (Er > 5 mm) was subtracted from the accumulated ETc (Equation (2)) [32].
where IID, FC, and θs were already defined, Da is the bulk density (g cm−3), and Pd is the profile depth (cm).
From here, irrigation treatments were applied based on a climatic water balance proposed by Servin-Palestina et al. [33], using Equation (2):
where Lri is the irrigation depth (mm), ETc is the daily crop evapotranspiration in (mm), Er was already defined, and n is the number of days between irrigation events (twice a week).
To estimate (Equation (3)), the local crop coefficient () for beans was estimated with Equation (4) [33]:
where was already defined and estimated by the Penman–Monteith method [34], and is the phenological-stage fraction of the crop calculated by TT, where zero value means planting and one is the physiological maturity of the crop.
The experiments were conducted in a randomized complete block design based on plot soil slope, where the irrigation treatments were randomized within every three blocks. The experimental unit comprised 12 furrows, each 11 m in length. To avoid horizontal movement of irrigation water among plots, there was a 1.52 and 0.8 m separation among blocks and treatments, respectively.
Irrigation water application per treatment was by self-compensated 6 mil-gauge irrigation tape with emitters spaced at 20 cm and with 0.94 L h−1 flow per emitter. Pre-sowing and establishment of irrigation were the same for all treatments. A volumetric meter was used to determine the amount of water (irrigation depth) applied to each treatment. The cumulative biomass curve was obtained with destructive sampling by collecting plants contained in a 1 m furrow section. From planting to harvest, seven plant samplings were carried out in both experiments. The sampled plants were divided into leaves, stems, and pods. Plant organ samples were oven-dried at 65 °C for 72 h to constant mass. Then, Bio was estimated with the sum of each organ. GY was obtained in triplicate (sub-sampling) for each experimental unit. The plants contained in a 1.5 m linear section of the central rows were harvested for GY determinations at 121 and 122 DAS in 2020 and 2021, respectively. The grain was dried at room temperature and weighed when 12% moisture was reached. Bio data were analyzed with a randomized complete block model and treatment means were grouped by Fisher’s least significant difference test at p ≤ 0.05. All calculations were carried out using the general linear model procedure of Statistical Analysis System software [35].
2.4. Climate Information
Daily climatic data for maximum temperature (Tmax), minimum temperature (Tmin), solar radiation (Rs), rainfall, and ETo for both growing seasons were obtained from the Adcon® automated weather station, located at 22.909° N–102.659° W at a distance of 1.5 km from the experimental site. The Adcon® platform estimates ETo by the Penman–Monteith method, using grass as a reference crop [34].
2.5. Description of the SIMPLE Model
The SIMPLE discrete-time dynamic model, proposed by Zhao et al. [21], simulates water-limited growth, development, and yield of crops using a daily time step, with functions or equations that explain the effect of daily temperature, heat stress, soil water availability, and atmospheric CO2 concentration (Table 1). TT (°C d) and Bio (t ha−1) were the state variables and GY (t ha−1) was an output variable. The SIMPLE model had thirteen parameters (Table 2), nine related to the crop type and four that specify differences among varieties (Figure 1). TT was used for crop growth modeling.

Table 1.
SIMPLE modal auxiliary equations.
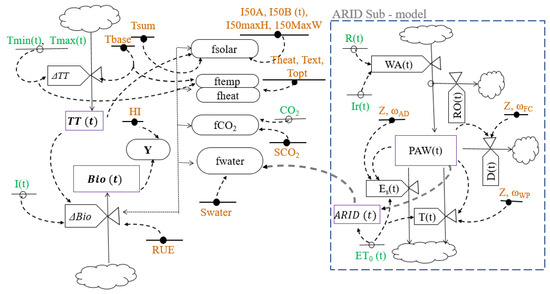
Figure 1.
Relational diagram of the SIMPLE crop growth model. State variables are represented by rectangles, rates of change by valves, parameters with a horizontal line and a circle with black filling, input variables with a circle and a horizontal line, and auxiliary variables with circles. Material flows are represented by normal arrows and information flows with dashed lines.
The SIMPLE model uses cumulative temperature to determine the rates of phenological development [36] as follows:
where TTi (°C d) is the thermal time in the i-th day, ΔTT (°C) is the daily increase in TT, T (°C) is the mean daily temperature, Tb (°C) is the base temperature for phenological crop development and n is the number of simulation days.
Biomass growth is based on radiation use efficiency [37], e.g., a fraction of the daily photosynthetically active radiation is intercepted by the plant and transformed into crop biomass.
where is the cumulative biomass on the i-th day, (t ha−1) is the daily cumulative biomass to physiological maturity, (t ha−1 day−1) is the daily biomass growth rate, n is the number of simulation days and fSolar is the fraction of solar radiation intercepted by crop canopy; RUE (g MJ−1 m−2) is the radiation use efficiency, and f(heat) is the heat stress factor; f(CO2) is the impact of CO2 on RUE, f(Temp) is the temperature impact on biomass growth rate, and f(Water) is the simple water-budget routine to estimated drought stress, factors that may or may not favor biomass accumulation. The f(CO2) effect is expressed through SCO2 (SIMPLE modal auxiliary equations are given in Table 1). This parameter is used for estimating the stress factor due to CO2 contraction.
GY is calculated as the product of total cumulative biomass () and the harvest index (HI) [38] as follows:
The SIMPLE model performs a soil–water balance based on the runoff-curve-number methodology for estimated surface runoff [39]. For this balance, four parameters are used to characterize of the experimental soil plot: (1) available water-holding capacity (0.123 m3 m−3), (2) deep drainage coefficient (0.5), (3) runoff number curve (0.81), and (4) root zone depth (600 mm). In addition, the model uses the standardized agricultural reference index for drought (ARID, Table 1) [40] to relate the effect of soil water content with the cumulative biomass, where Swater is a water stress parameter. The parameters obtained consider that the experiment was carried out on agricultural land with a clay loam texture with little development and a slope > 3. Many of the SIMPLE functions have been used in other crop models, described extensively by Zhao et al. [21].
2.6. Model Calibration
According to the dynamic systems modeling procedure [41,42,43], before calibration, a global sensitivity analysis (GSA) was conducted to identify the most influential parameters in the model [44,45]. The GSA was performed using the Sobol method [46] where ± 20% uncertainty was applied to most of the parameters, avoiding cardinal temperature overlap and using a uniform distribution, except for Tsum and Swater, where a normal function was used (Table 2). The most influential candidate parameters for calibration were I50A, Topt, Swater, Tb, I50maxW, and Tsum.
Table 2.
SIMPLE model parameters with their nominal values and exploration limits used for global sensitivity analysis and calibration in ‘Pinto Saltillo’ beans.
Table 2.
SIMPLE model parameters with their nominal values and exploration limits used for global sensitivity analysis and calibration in ‘Pinto Saltillo’ beans.
| Parameter | Description | Nominal | Threshold * | Units | Cite |
|---|---|---|---|---|---|
| Tsum | Cumulative temperature from sowing to maturity | 1200 | 1047–1356 & | °C d ** | B-G |
| HI | Harvest index | 0.36 | 0.29–0.43 | - | B-G |
| I50A | The cumulative temperature required for leaf area development to intercept 50% of radiation | 450 | 360–540 | °C d | Z |
| I50B | Cumulative temperature till maturity to reach 50% radiation interception due to leaf senescence | 200 | 160–240 | °C d | Z |
| Tb | Baseline temperature for phenology development and growth | 8 | 6.4–9.6 | °C | B-G |
| Topt | The optimal temperature for biomass growth | 30 | 22–30 | °C | B-G |
| RUE | Radiation use efficiency (above ground only and no respiration) | 3.21 | 2.57–3.85 | g MJ−1 m−2 | K |
| I50maxH | Maximum daily reduction in I50B due to heat stress | 90 | 72–108 | °C d | Z |
| I50maxW | Maximum daily reduction in I50B due to drought stress | 20 | 16–24 | °C d | Z |
| Tmax | Threshold temperature to start accelerating heat-stress senescence | 35 | 32.1–42 | °C | O |
| Text | Extreme temperature threshold when RUE becomes 0 due to heat stress | 45 | 42.1–52.5 | °C | Z |
| SCO2 | The relative increase in RUE per ppm of CO2 after 350 ppm | 0.07 | 0.06–0.08 | ppm | Z |
| Swater | Sensitivity of RUE to drought stress | 0.9 | 0.48–1.28 & | - | Z |
* Minimum and maximum limits, & parameters to which normal function was applied, ** degree days. Highlighted parameters are the most influential ones; B-G = Baez-Gonzalez et al. [47], Z = Zhao et al. [21], K = Karimzadeh et al. [48], O = Omae et al. [49].
2.6.1. Differential-Evolution Algorithms
Model calibration was performed with a differential evolution (DE) algorithm, which is evolutionary algorithm for solving global optimization problems. The DE algorithm is considered a simple, effective, and efficient heuristic search method inspired by natural evolution [50]. The DE algorithm includes a population of potential solutions and explores the search space using mutation, crossover, and selection operators. This algorithm has only three parameters that must be specified to optimize a problem. The parameters are population size (PS), mutation factor (MF), and crossover probability (CP).
The DE algorithm characteristics applied in the simulation consisted of six estimated parameters (D), a PS of 60 (D · 10), and the number of generations equal to 1000; the minimum values were taken from the mean of 25 runs and the DE/rand/1/bin algorithm strategy. This DE algorithm was programmed in the MATLAB® environment version 2020b. The parameter values of the DE algorithm were PS = 60, MF = 0.6, and CP = 0.9. The exploration threshold of the calibrated parameters is specified in Table 2.
2.6.2. Objective Function
An objective function to optimize a problem is defined as:
where is the biomass predicted by the SIMPLE model at a time and is the measured variable at the time , is the number of samples during the growth period, is the parameter vector established for calibration and is the parameter vector that yields the minimum value of .
2.7. Model Evaluation
After performing the SIMPLE model calibration, the average estimated values and nominal values of the parameters, given in Table 2, were used to perform a new simulation for estimating the TT, Bio, and GY values using the climatic and crop data obtained during the growing season 2021.
2.8. Measures for the Degree of Fit
To measure the calibration and evaluation quality in the biomass simulation by the SIMPLE model, the following measures of agreement [51] between the observed and simulated values were used: (1) the bias (BIAS), (2) the mean absolute error (MAE), (3) the root-mean-square error (RMSE), and (4) the efficiency (EF) proposed by Nash and Sutcliffe (1970), which characterizes the behavior of the simulation model. The perfect model should have an efficiency close to 1 [52].
3. Results
3.1. Climate and Irrigation Schedule
The average multi-year rainfall (2002–2019) from April to August was 276.7 mm. The cumulative precipitation during the experimental period was 352 mm and 387 mm, respectively, for the 2020 and 2021 growing seasons. The maximum temperature occurred in June and May, and the minimum temperature in April for 2020 and 2021, respectively. The average temperature for the growing season 2020 was 4% higher than in 2021 (Figure 2A). However, air temperatures recorded throughout the growing season determined crop development, with flowering starting at 54 and 50 DAS for the 2020 and 2021 growing seasons, respectively. In addition, the average solar radiation was 28.1 MJ m2 day−1 for both experimental years, 3% lower than the historical average (Figure 2B).
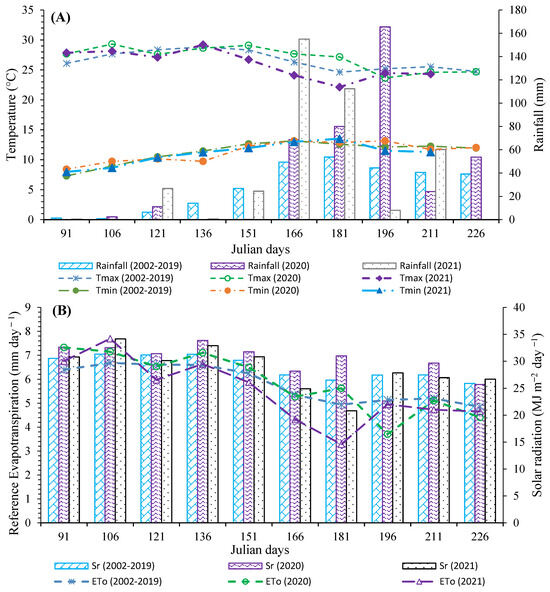
Figure 2.
Climatic variables recorded during the growing season with historical averages (17 years) for the CEZAC INIFAP-Zacatecas station ((A)Temperature and (B) Reference Evapotranspiration). Averages of every fifteen days; Tmax and Tmin are the maximum and minimum temperature, respectively, Sr is the solar radiation, and ETo is the reference evapotranspiration.
Climatic conditions, mainly rainfall and reference evapotranspiration, influenced the irrigation depths applied in each treatment (Table 3). In 2020, low Pp and high ETo were recorded, resulting in a high crop water demand. The I50 (82%) and I125 (66%) irrigation treatments were applied mostly during the reproductive phenological stage in 2020 respectively. The corresponding values for these irrigation treatments at the same phenological stage were 100% and 77%, respectively, in 2021. While the total water applied increased, water use efficiency decreased, and vice versa, in both growing seasons. Water productivity (WP) values were not clear in the 2020 data, but in the 2021 data WP increased as water irrigation inputs decreased (Table 3).
Table 3.
Applied irrigation depth (AID), effective rainfall (ER), total water applied (TWA), water use efficiency (WUE), and water productivity (WP) of bean grain yield (Y) as influenced by irrigation treatments (IT) in Zacatecas, Mexico.
3.2. Biomass Accumulation Curve
During the first 25 DAS, cumulative biomass was very slow and there were no measurable readings among treatments, since emergence occurred at 8 and 7 DAS for the 2020 and 2021 growing seasons, respectively. When irrigation treatments were applied, a dynamic growth proportional to the amount of water applied was observed at the start of flowering at 54 and 50 DAS for 2020 and 2021, respectively. Subsequently, in grain filling, the variation in biomass increase among treatments can be attributed to the presence of rainfall. The most intense rainfall events were recorded in the middle of the reproductive cycle and at the beginning of flowering for the 2020 and 2021 growing seasons, respectively (Table 4 and Table 5). Data generated in 2020 and 2021 were used for the calibration and evaluation procedures, respectively.
Table 4.
Mean values of cumulative biomass (t ha−1 ± standard deviation) of ‘Pinto Saltillo’ bean under irrigation treatments (IT) in Zacatecas, Mexico in the 2020 growing season.
Table 5.
Mean values of cumulative biomass (t ha−1 ± standard deviation) of ‘Pinto Saltillo’ bean under irrigation treatments (IT) in Zacatecas, Mexico, in the 2021 growing season.
3.3. Calibration and Evaluation
Using the differential evolution method, 20 optimizations were carried out. The mean of each parameter per treatment is indicated in Table 6. The largest standard deviation in the parameter set was observed at 1.17 × 10−16. This value is indicative of the fact that the calibration process converged to the global maximum; therefore, the results were considered reliable. On the other hand, for the evaluation of the SIMPLE model, the average of the calibrated parameter values (Table 6) with data from the year 2020 and the nominal values (Table 2) were used, as appropriate. With this, the new parameter vector was generated; subsequently, the biomass was simulated, and then the performance measures were obtained in the evaluation stage for the observed 2021 data.
Table 6.
Vector of parameters resulting from the SIMPLE model calibration using the differential-evolution algorithm.
Using the exploration thresholds in Table 2 for the six parameters in the calibration process, the Tsum, I50A, and I50maxW values were found to be close to the extreme values (Table 6). Tsum shifted to the upper end with a value from 1354 to 1356 °C d. It should be noted that it was tested with other values higher than 1356 °C d (unpublished data). However, overfitting was observed. For instance, the statistics in the calibration are improved for some treatments, but in the evaluation stage the statisticians indicate the low reliability of the SIMPLE model for simulating bean biomass. Topt was inversely proportional to the irrigation treatment, e.g., the treatment with the highest water availability had the lowest Topt value. Swater was the parameter that correlated with water availability within the SIMPLE model. However, no linear trend was observed concerning the treatments. Otherwise, for I125 and I75 irrigation treatments, the Tb was 8.7 °C and 9.6 °C, respectively (Table 6). However, Topt is a cultivar parameter and there should be no variation between irrigation treatments. Also, a relationship between I50A and Swater was observed, with the I50 irrigation treatment having the highest water stress with the lowest I50A value and Swater increasing according to the trend of the parameters reported in the calibration stage. However, Tsum is expected to be the parameter with the greatest variability, since the crop cycle decreases with water stress. The results can be attributed to the method used by the SIMPLE model to estimate crop development.
In the calibration process, the cumulative biomass simulated did not follow a linear trend concerning the irrigation treatment. Values fluctuated between 3.47 t ha−1 and 10.2 t ha−1 for the I50 and I100 irrigation treatments, respectively. The I75 irrigation treatment had the maximum RMSE value (0.59 t ha−1) of biomass. In the evaluation process, the predicted bean biomass adequately fitted the observed data for the four irrigation treatments with RMSE < 1.80 t ha−1 and EF > 0.89 values, with average values of 1.48 t ha−1 and 0.92 for RMSE and EF, respectively. In addition, the I50 irrigation treatment had the best fit in calibration and evaluation processes according to the statistics used to measure model performance (Table 7).
Table 7.
Evaluation statistics of the SIMPLE model for simulating the biomass of beans subjected to different moisture levels.
The simulated biomass for each irrigation treatment responded to a sigmoidal trend curve in both the calibration and evaluation process (Figure 3). Bean plants had slow growth before flowering (51 DAS) until pod formation (59 DAS), then an exponential growth was observed between 60 and 80 DAS and, finally, crop growth had an asymptotic pattern from grain filling (85 DAS) until maturity (120 DAS). In the evaluation process, the simulated biomass had a positive linear trend with respect to the irrigation treatments, e.g., the more available soil moisture, the greater the biomass accumulation. In addition, biomass values were higher than those values obtained during calibration for all irrigation treatments. This condition can be attributed to the fact that during the 2020 growing season (calibration data), rainfall was < 35.4 mm and reference evapotranspiration was > 62.6 mm higher than in 2021 (evaluation data). Also, as expected, the Bias, MAE, RMSE, and EF statistics showed better fit in calibration than in evaluation. In the model evaluation, I100 had the lowest values, with 1.8 t ha−1 and 0.89 for RMSE and EF, respectively. However, the overall efficiency for all irrigation treatments was 0.92, which is acceptable for simulating crop biomass.
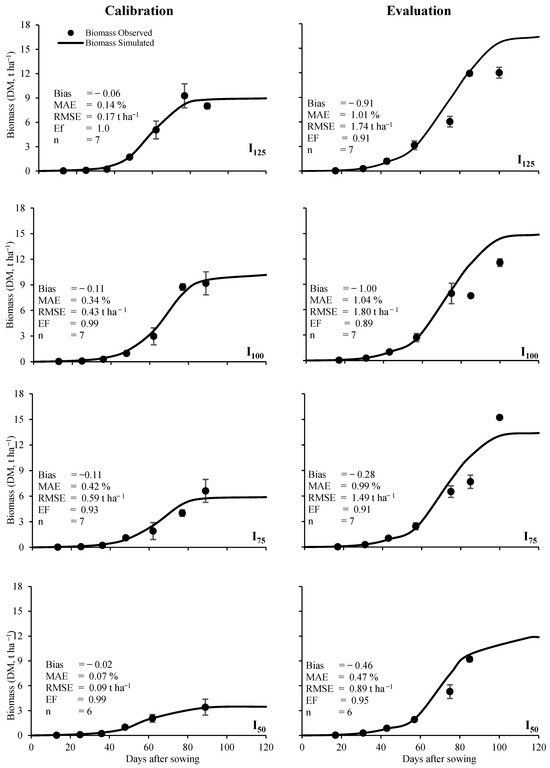
Figure 3.
Observed vs. simulated biomass (DM = dry matter) for the four irrigation treatments. In calibration (2020 data) and evaluation (2021 data), MAE is the mean absolute error, RMSE is the root-mean-square error, and EF is the efficiency.
The overall data performance for the calibration and evaluation processes, with the relationship between observed and simulated biomass, is given in Figure 4. The evidence confirmed that the SIMPLE model calibration had a better fit between observed and simulated dry biomass compared with the evaluation procedure (Figure 4A and Figure 4B, respectively). In the calibration process, the values furthest from the 1:1 line corresponded to the data obtained at 77 and 89 DAS from the I125 and I75 irrigation treatment, respectively (Figure 4A). That is, the general statistics evidenced that the model simulated bean biomass adequately under all irrigation treatments tested here. In the evaluation process, values less than 4 t ha−1 overlapped on the 1:1 line (Figure 4B). That is, it presented a good fit for the vegetative stage, while in the reproductive stage the estimated values were underestimated. Nevertheless, in the evaluation stage the SIMPLE model’s statistics pointed to a good performance in predicting bean biomass under all irrigation treatments studied here.
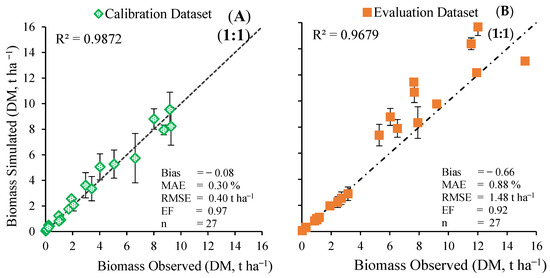
Figure 4.
Calibration (A) and evaluation (B) processes of observed and simulated biomass. At each point, the vertical bars indicate the standard deviation, MAE is the mean absolute error, RMSE is the root-mean-square error, and EF is the efficiency.
The grain yield estimation (Y; t ha−1) of the ‘Pinto Saltillo’ bean by the SIMPLE model using the HI parameter of 0.36 was unsatisfactory (unreported data). The average errors for all irrigation treatments in the calibration and evaluation procedures were 37.1% and 21.6% for the 2020 and 2021 data, respectively. They ranged between 12.7% and 61.8%, corresponding to the I100 and I50 irrigation treatments for the growing seasons of 2020 and 2021, respectively. For this reason, a multiple regression model involving biomass and transpiration was proposed to estimate the harvest index (HI) and reduce estimation errors.
3.4. Harvest Index
The regression model developed for estimating the harvest index (HI) considered two steps: (1) the selection of the regression model and (2) the calibration process.
In the first step, a regression model was generated using the observed Bio and Y, using data obtained at 100 DAS in 2021. Triplicate data from each irrigation treatment and HI were calculated by relating Y to Bio. Also, the sum of transpiration (STj), resulting from the last 100 days of the SIMPLE model simulation, was used. Subsequently, relative values were obtained by relating Bioj, Yj, and HIj to their corresponding maximum values. The first-order multiple regression model (Equation (12)) was performed in the statistical analysis system (SAS, 2011). The model was significant (p < 0.05) and explained 50% (R2) of the dataset variability, with a coefficient of variation of 18.3% and 0.13 RMSE.
where HIj_max is the maximum harvest index observed (0.29), Bioj_max is the maximum cumulative biomass observed (16.7 t ha−1), and STj_max is the sum of simulated transpiration (419 mm).
In the second step, the simulated cumulative biomass (BioSim) and the sum of simulated transpiration (STSim) at the end of the growing season were used for each of the treatments for 2020 and 2021. BioSim and STSim were the results of simulation for all treatments with the SIMPLE model, using the calibrated parameters in Table 7.
Consecutively, the values of , , , and were optimized until reaching a minimum RMSE between observed and simulated yield. For the optimization analysis, the lsqnonlin.m function of the MATLAB® optimization tool was used. remained fixed; however, there was a better fit when using the base harvest index of 0.36 obtained for the ‘Pinto Saltillo’ variety under climatic conditions similar to those in this study. The model for estimating the resulting harvest index was the following:
To determine the model’s performance, corrected relative error (CRE) was used to avoid negative values. CRE measures the quality of the estimate, where values >10% are considered ‘excellent’ and 10 > REC < 20 are ‘good’ for estimating grain yield.
where represents the observed yield and the value predicted by the model as a function of BioSim and .
The proposed regression model simulated the harvest index reliably for most treatments because a CRE of less than 10% was observed in almost all treatments, except for the I50 and I100 irrigation treatments in the 2020 and 2021 growing seasons, respectively (Table 8).
Table 8.
Simulated and observed grain yield of ‘Pinto Saltillo’ bean under irrigation treatments (IT) in Zacatecas, Mexico.
4. Discussion
4.1. Climate and Irrigation Schedule
Temperature is one of the environmental factors that influence crop growth and development [53]. Barrios et al. [54] pointed out that beans can grow with average temperatures ranging between 15 and 27 °C, with an optimal temperature of 25 °C. In contrast, Beebe et al. [55] indicated that the optimal temperature for this crop is between 17.5 and 23.1 °C. The air temperature during the two growing seasons was in good agreement with those observed in other studies (Figure 2).
Late rainfed-bean varieties planted in northern Mexico require more than 240 mm of rainfall during the production cycle with yields of 0.6 t ha−1, but in rainy years (323 mm of rainfall) high yields could be achieved (1.4 t ha−1) [56]. High rainfall was recorded atypically in both growing seasons studied during the vegetative stage, with 96% and 87% for 2020 and 2021, respectively (Figure 2). Lynch and van Beem [57] reported that the bean crop can experience water stress due to erratic rainfall, with the reproductive stage being the most sensitive [58]; this did not occur during the experimental period.
Beebe et al. [55] state that the bean crop requires from 363 to 450 mm of irrigation depth throughout the growing season. Rai et al. [59] applied 375 mm in the largest irrigated treatment under semi-arid conditions in Wyoming, USA. In the experiments described here, the irrigation depths applied were lower than that indicated by Rai et al. [59]. In this study, considering the ER plus the applied irrigation depths, the total water applied for I50 was 535 mm and 520 mm, respectively, for the 2020 and 2021 growing seasons (Table 3). These rainfalls may have temporally masked the effect of the I50 and I75 irrigation treatments.
4.2. Cumulative Biomass Curve
In the 2020 growing season, the irrigation treatments accumulated less total biomass compared with their counterparts in the 2021 growing season (Table 3 and Table 4). This may be attributed to lower water availability at the flowering stage in the 2020 growing season due to the value used, which could underestimate the water requirements of bean crops [58]. Water stress at the flowering stage reduces leaf number and stem length, and therefore, this is reflected in grain mass per plant [58]. Simulated biomass increased proportionally to the irrigation treatments in both growing seasons (Table 8). This was clearer in 2020 than in 2021. In the latter year, 53% of the rainfall occurred at the pre-flowering stage. This could then, temporally, mask the irrigation treatment effect. The opposite occurred in the former year, where 70% of rainfall occurred at the grain-filling stage in favor of beans plants under I100 and I125. Therefore, based on Equations (7) and (8), bean plants experiencing I100 and I125 irrigation treatments suggest a greater canopy for intercepting solar radiation and, therefore, more photo assimilates were available and distributed to growing organs (sink), mainly to the grains (Table 8) [60]. Therefore, the SIMPLE model may be suggested to simulate bean Bio and Y for different scenarios in the semi-arid agricultural lands of north-central Mexico.
4.3. Calibrated Parameters
The value of standard deviation close to zero and the value of the objective function > 0.6 are reliable indicators for determining the effectiveness of search algorithms, according to Trejo-Zúñiga et al. [61]. In addition, the DE algorithm offers better approximations of the global optimum compared to other algorithms [61,62]. On the other hand, Tsum is the parameter that indicates the cumulative temperature from sowing to physiological maturity. The Tsum values reported for beans ranged between 950 and 2700 °C d [21,47,59,63,64]. However, the physiological maturity of the bean crop is mainly for beans with determined growth habits, variety, and photoperiod. Some studies indicate that it is mainly irrigation management which modifies the development cycle, although the SIMPLE model did not adequately represent this effect. I50A is the cumulative temperature requirement for leaf area development to intercept 50% of radiation; in beans, values of 450 °C d have been reported [21]. Tb is the temperature at which crop development stops or starts. For beans, Tb values of 5 °C [59], 8 °C [47], and 12 °C [64] have been reported. Topt is the optimal temperature at which growth reaches its maximum expression; thus, values of 25 °C [65], 27 °C [21], and 30 °C [47] have been reported. Furthermore, beans can withstand extreme temperatures of between 5 and 40 °C [66]. The Tsum, Topt and Tb values reported as a result of calibration in this study are within the ranges mentioned by the authors cited above (Table 6).
The Swater is the parameter of RUE sensitivity to the ARID index [21]. That is, it incorporates the water availability effect on the crop, where an ARID equal to zero indicates that the crop is at its maximum water demand (no stress) [40]. However, the average value for the treatment with the highest irrigation depth applied (I125 in 2020) between vegetative growth and flowering initiation stages was 0.39 (moderate stress). This was indicative that Kc underestimated the ETc compared with the 0.09 (no stress) observed between flowering initiation and physiological maturity, which was coincident with an atypically rainy season for both growing seasons. In addition, the linear behavior of Swater values and the applied irrigation depth was not observed. That is, Swater alone did not reflect the water effect in the model because there is an interaction between parameters. Besides, as pointed out by Bulatewicz et al. [67], here it was not possible to explore a wider range of irrigation depths due to the establishment of the rainy season which equalized water storage in the soil, thereby masking the effect of the irrigation treatments. Teweldebrhan et al. [68] argue that there is a multiplicative interaction between the parameters of a model; that is, the effect of one parameter is reflected in the output variables, as long as another parameter intervenes.
4.4. Calibration and Evaluation
In individual performance (Figure 3), the biomass curve obtained was similar to that established by other authors [69,70,71]. The calibration and evaluation statistics were within an acceptable range for all irrigation treatments [72,73]. Also, the observed RMSE was slightly higher than those recorded for beans subjected to different moisture levels evaluated with the SALTMED model, specifically for water balance [74]. In general, the statistics observed in this study (Figure 4) were considered acceptable in the crop biomass simulation, considering that the estimated values of the parameters are only approximations of the real values and probably have a fairly substantial error [9], taking into consideration the fact that the bean crop has a significant genotype x environment interaction [54] and that the SIMPLE model does not consider genetic parameters.
In the calibration stage, the observed biomass values were lower than those reported in the evaluation; these results are attributable to the climatic conditions of 2021, which favored bean development. In this respect, Emam et al. [75] point out that common bean biomass decreased, as did the water supply. However, the biomass values found in this study agreed with other studies. For example, Morales-Rosales et al. [76] and Dewedar et al. [74] reported values of 13.0 t ha−1 and 14.6 t ha−1, respectively, for irrigated beans, while mean values between 4.0 and 8.0 t ha−1 for different bean varieties were reported by Acosta-Gallegos and Rosales-Serna [77].
4.5. Harvest Index
The estimate of the performance with the SIMPLE model raises a linear relationship with the estimated biomass. That is, it calculates the yield as a percentage of the cumulative simulated biomass at the end of the cycle without considering any other environmental or management factor. Nevertheless, here, HI was not linearly proportionally to the cumulative biomass at the maturity stage. Attributes such as crop management and genotypic adaptation may play a part in the components of yield productivity under water limitation [78]. In addition, several methods have been proposed to calculate HI, which consider cumulative biomass after anthesis, and whose behaviors were nonlinear [79]. Bean HI is related to the availability of photoassimilates which are exported mainly to sink organs (grains) [76]. These are, in turn, related directly to the intercepted solar radiation by the crop and the soil water availability to satisfy bean crop evapotranspiration demand.
The yield data generated in both experiments agreed with those of Ranjan et al. [80]. These authors, evaluating moisture and tillage levels in ‘Pinto Saltillo’ beans, obtained grain yields between 3.0 and 5.8 t ha−1. Thus, the model proposed in this study to estimate grain yield (Y) gave excellent results compared with those reported with the CROPGRO Dry-bean model. Dallacort et al. [81] state that this model underestimated Y by between 19% and 29% for beans under water-deficit conditions. Monpremier et al. [82] also used CROPGRO Dry-bean to simulate Y of different bean varieties with relative errors of 32.1% and 15.6% in the calibration and evaluation procedures, respectively. The SSM-Legumes model produced an RMSE of 0.41 t ha−1 in the grain-yield simulation [83]. In another study, relative differences < 4.9% between observed and simulated bean yields under different irrigated conditions were reported by using the SALMED model [74]. Similarly, with the AquaCrop model, an RMSE > 0.28 t ha−1 was found [84].
5. Conclusions
The SIMPLE model acceptably simulates biomass dynamics in the common bean crop, since when calibrating the I50A, Topt, Swater, Tb, I50maxW, and Tsum parameters a good fit between predictions and measurements was obtained. Despite having only a few parameters, the SIMPLE model had the potential to simulate bean biomass under different moisture conditions, and therefore, it can be a feasible tool for planning productive activities for bean cultivation. However, when changes in soil moisture are very small, the model cannot express these in the biomass simulation.
In the evaluation process under conditions other than rainfall and evapotranspiration, the SIMPLE model adequately simulates crop biomass. The results indicate that the SIMPLE model is a reliable and robust tool for simulating biomass. It can be used to calculate the daily water demand of common beans as a function of growing degree days. Therefore, irrigation can be programmed in real-time for arid and semi-arid conditions in northern Mexico.
The cumulative bean biomass concerning the applied irrigation depth responded to a linear model. While the cumulative biomass curve showed a sigmoid behavior, the grain yield had an expected behavior. Nevertheless, in the proposed multiple linear regression model, a good bean-yield estimation was possible. However, the harvest index needs to be validated under other irrigation levels and further field irrigation experiments in different locations are needed to strengthen the SIMPLE model to simulate accurate bean yield under climate change scenarios.
Author Contributions
Conceptualization, M.S.-P. and I.L.-C.; methodology, M.S.-P., I.L.-C., A.R.-G. and R.S.-M.; software, A.R.-G. and I.L.-C.; validation, A.R.-G. and I.L.-C.; formal analysis, J.A.Z. and J.Á.C.-R.; investigation, J.A.Z. and J.Á.C.-R.; resources, I.L.-C. and R.S.-M.; data curation, J.Á.C.-R. and A.R.-G.; writing—original draft preparation, M.S.-P., I.L.-C., A.R.-G. and R.S.-M.; writing—review and editing, M.S.-P., A.R.-G. and J.A.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was partially supported by Universidad Autónoma Chapingo (UACh), project “Simulation of bean growth, development and yield using dynamic mathematical models” ref. no. 20115-C-86.
Data Availability Statement
The datasets generated and/or analyzed during the current study are not publicly available because this information belongs to our sponsors (Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecurias, Universidad Autónoma Chapingo, and The Consejo Nacional de Humanidades, Ciencias y Tecnologías of Mexico), but they are available from the main author upon reasonable request.
Acknowledgments
The first author thanks The Consejo Nacional de Ciencia y Tecnología (CONACYT) of Mexico and the Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP) for supporting his Ph.D. program at Universidad Autónoma Chapingo grand ref. no.: 714120. We also thank the editor and reviewers for the comments and suggestions to enhance the final manuscript presentation.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- FAOSTAT. FAO Statistics, Food and Agriculture Organization of the United Nations. 2022. Available online: http://www.fao.org/faostat/en/ (accessed on 20 January 2022).
- INEGI (Instituto Nacional De Estadistica y Geografia). Censo Agropecuario 2007. 2012. Available online: http://www3.inegi.org.mx (accessed on 15 May 2022).
- García, E. Modificaciones al Sistema de Clasificación Climática de Köppen (Quinta ed.). Instituto de Geografía-Universidad Nacional Autónoma de México (UNAM): México, DF, México, 2004. Available online: http://www.publicaciones.igg.unam.mx/index.php/ig/catalog/view/83/82/251-1 (accessed on 22 June 2022).
- Schwartz, H.F.; Brick, M.A. Dry Bean Pest Management and Production, 3rd ed.; Cooperative Extension Resource Center, Colorado State University: Fort Collins, CO, USA, 2015. [Google Scholar]
- SIAP. Servicio de Información y Estadística Agroalimentaria y Pesquera Información Agrícola. Avances Mensuales por Estado. 2015. Available online: http://www.siap.sagarpa.gob.mx (accessed on 15 May 2022).
- Seidel, S.J.; Palosuo, T.; Thorburn, P.; Wallach, D. Towards improved calibration of crop models—Where are we now and where should we go? Eur. J. Agron. 2018, 94, 25–35. [Google Scholar] [CrossRef]
- Chalabi, Z.; Zhou, W. Optimal control methods for agricultural systems. Acta Hortic. 1996, 406, 221–228. [Google Scholar] [CrossRef]
- Bhar, A.; Kumar, R.; Qi, Z.; Malone, R. Coordinate descent based agricultural model calibration and optimized input management. Comput. Electron. Agric. 2020, 172, 105353. [Google Scholar] [CrossRef]
- Wallach, D. Crop model calibration: A statistical perspective. Agron. J. 2011, 103, 1144–1151. [Google Scholar] [CrossRef]
- Jones, J.W.; Hoogenboom, G.; Porter, C.H.; Boote, K.J.; Batchelor, W.D.; Hunt, L.A.; Wilkens, P.W.; Singh, U.; Gijsman, A.J.; Ritchie, J.T. The DSSAT cropping system model. Eur. J. Agron. 2003, 18, 235–265. [Google Scholar] [CrossRef]
- Hoogenboom, G.; White, J.W.; Jones, J.W.; Boote, K.J. Beangro V1. 01 Dry Bean Crop Growth Simulation Model: Users Guide; Department of Agricultural Engineering, University of Florida: Gainesville, FL, USA, 1991. [Google Scholar]
- White, J.W.; Hoogenboom, G. Simulating effects of genes for physiological traits in a process-oriented crop model. Agron. J. 1996, 88, 416–422. [Google Scholar] [CrossRef]
- Hoogenboom, G.; White, J.W.; Jones, J.W.; Boote, K.J. BEANGRO: A process-oriented dry bean model with a versatile user interface. Agron. J. 1994, 86, 182–190. [Google Scholar] [CrossRef]
- van Keulen, E.L.; Wolf, J. Modelling of Ugricultural Production: Weather, Soil and Crops; Simulation Monographs, Pudoc: Wageningen, The Netherlands, 1986. [Google Scholar]
- O’Callaghan, J.R.; Maende, C.; Wyseure, G.C.L. Modelling the intercropping of maize and beans in Kenya. Comput. Electron. Agric. 1994, 11, 351–365. [Google Scholar] [CrossRef]
- Van Keulen, H.; Penning de Vries, E.W.T.; Drees, E.M. A summary model for crop growth. In Simulation of Plant Growth and Crop Production; de Vries, E.W.T.P., Laar, H.H., Eds.; Simulation Monograph, Pudoc: Wageningen, The Netherlands, 1982; pp. 87–99. [Google Scholar]
- Verdoodt, A.; Van Ranst, E.; Ye, L. Daily simulation of potential dry matter production of annual field crops in tropical environments. Agron. J. 2004, 96, 1739–1753. [Google Scholar] [CrossRef]
- Soltani, A.; Sinclair, T.R. Modeling Physiology of Crop Development, Growthand Yield; CABI: Wallingford, UK, 2012. [Google Scholar]
- Seidel, S.J.; Rachmilevitch, S.; Schütze, N.; Lazarovitch, N. Modelling the impact of drought and heat stress on common bean with two different photosynthesis model approaches. Environ. Model. Softw. 2016, 81, 111–121. [Google Scholar] [CrossRef]
- Yuan, M.; Ruark, M.D.; Bland, W.L. A simple model for snap bean (Phaseolus vulgaris L.) development, growth and yield in response to nitrogen. Field Crops Res. 2017, 211, 125–136. [Google Scholar] [CrossRef]
- Zhao, C.; Liu, B.; Xiao, L.; Hoogenboom, G.; Boote, K.J.; Kassie, B.T.; Pavan, W.; Shelia, V.; Kim, K.S.; Hernandez-Ochoa, I.M.; et al. A SIMPLE crop model. Eur. J. Agron. 2019, 104, 97–106. [Google Scholar] [CrossRef]
- Annandale, J.G.; Benadé, N.; Jovanovic, N.Z.; Steyn, J.M.; Du Sautoy, N. Facilitating Irrigation Scheduling by Means of the Soil Water Balance Model; Water Research Commission Report No. 753/1/99; Water SA Publishes: Pretoria, South Africa, 1999. [Google Scholar]
- Ragab, R. A holistic generic integrated approach for irrigation, crop and field management: The SALTMED model. Environ. Model. Softw. 2002, 17, 345–361. [Google Scholar] [CrossRef]
- Steduto, P.; Hsiao, T.C.; Raes, D.; Fereres, E. AquaCrop—The FAO Crop Model to Simulate Yield Response to Water: I. Concepts Underlying Principles. Agron. J. 2009, 101, 426–437. [Google Scholar] [CrossRef]
- Wang, J.; Li, X.; Lu, L.; Fang, F. Parameter sensitivity analysis of crop growth models based on the extended Fourier Amplitude Sensitivity Test method. Environ. Model. Softw. 2013, 48, 171–182. [Google Scholar] [CrossRef]
- Martínez-Ruiz, A.; López-Cruz, I.L.; Ruiz-García, A.; Pineda-Pineda, J.; Prado-Hernández, J.V. HortSyst: A dynamic model to predict growth, nitrogen uptake, and transpiration of greenhouse tomatoes. Chil. J. Agric. Res. 2019, 79, 89–102. [Google Scholar] [CrossRef]
- Pham, Q.V.; Nguyen, T.T.; Vo, T.T.; Le, P.H.; Nguyen, X.T.; Duong, N.V.; Le, C.T. Applying the SIMPLE crop model to assess soybean (Glicine max.(L.) Merr.) biomass and yield in tropical climate variation. Agronomy 2023, 13, 1180. [Google Scholar] [CrossRef]
- Suliansyah, I.; Arlius, F.; Chaniago, I.; Xuan NT, T.; Duong VT, H.; Dung, N.P.; Van Quang, P. Simulation of Maize Biomass and Yield in An Giang, Vietnam, under Climate Variabilities. Trends Sci. 2024, 21, 7490. [Google Scholar] [CrossRef]
- Suliansyah, I.; Arlius, F.; Chaniago, I.; Xuan NT, T.; Van Quang, P. Literature Review Crop Modeling and Introduction a Simple Crop Model. J. Appl. Agric. Sci. Technol. 2023, 7, 197–216. [Google Scholar] [CrossRef]
- Medina-García, G.; Rumayor-Rodríguez, A.F.; Cabañas-Cruz, B.; Luna-Flores, M.; Ruiz-Corral, J.A.; Gallegos-Valdez, C.; Madero-Tamargo, J.; Gutiérrez-Sánchez, J.R.; Rubio-Díaz, S.; Bravo-Lozano, A.G. Potencial Productivo de Especies Agrícolas en el Estado de Zacatecas; Libro Técnico Núm. 2; Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias. Centro de Investigación Regional Norte Centro: Zacatecas, Mexico, 2003; 157p.
- Palacios, V.E. Por Qué, Cuándo, Cuánto y Cómo Regar? Para Lograr Mejores Cosechas; DF Editorial Trillas: México City, Mexico, 2002; 214p. [Google Scholar]
- Serna-Pérez, A.; Zegbe, J.A.; Mena-Covarrubias, J. Rendimiento y calidad de chile seco Mirasol cultivado bajo riego parcial de la raíz. Rev. Chapingo Ser. Hortic. 2011, 17, 19–24. Available online: http://www.scielo.org.mx/pdf/rcsh/v17nspe1/v17nspe1a4.pdf (accessed on 10 April 2022). [CrossRef]
- Servín-Palestina, M.; Tijerina Chávez, L.; Medina García, G.; Palacios Velez, O.; Flores Magdaleno, H. Sistema para programar y calendarizar el riego de los cultivos en tiempo real. Rev. Mex. De Cienc. Agrícolas 2017, 8, 423–430. [Google Scholar] [CrossRef][Green Version]
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Evapotranspiración del Cultivo: Guía Para la Determinación de Los Requerimientos de Agua de Los Cultivos. Estudio FAO Riego y Drenaje; FAO: Roma, Italy, 2006; pp. 56–323. Available online: http://www.fao.org/docrep/009/x0490s/x0490s00.htm (accessed on 1 April 2024).
- SAS Institute Inc. SAS 9.3 System Options: Reference, 2nd ed.; SAS Institute Inc.: Cary, NC, USA, 2011. [Google Scholar]
- Ritchie, J.T.; Godwin, D.C.; Otter-Nacke, S. CERES-Wheat: A User-Oriented Wheat Yield Model. Preliminary Documentation; AGRISTARS Publication No. YM-U3-04442-JSC-18892; Michigan State University: East Lansing, MI, USA, 1985; Available online: https://cir.nii.ac.jp/crid/1573387449661175552 (accessed on 1 April 2024).
- Monteith, J.L. Light distribution and photosynthesis in field crops. Ann. Bot. 1965, 29, 17–37. [Google Scholar] [CrossRef]
- Amir, J.; Sinclair, T.R. A model of water limitation on spring wheat growth and yield. Field Crops Res. 1991, 28, 59–69. [Google Scholar] [CrossRef]
- SCS. Soil Conservation Service, National Engineering Handbook, Section 4: Hydrology; US Department of Agriculture: Washington, DC, USA, 1972.
- Woli, P.; Jones, J.; Ingram, K.; Fraisse, C. Agricultural reference index for drought (ARID). Agron. J. 2012, 104, 287–300. [Google Scholar] [CrossRef]
- Ljung, L.; Glad, T. Modeling of Dynamic Systems; PTR Prentice Hall: Englewood Cliffs, NJ, USA, 1994. [Google Scholar]
- Voinov, A.; Gaddis, E.J.B. Lessons for successful participatory watershed modeling: A perspective from modeling practitioners. Ecol. Model. 2008, 216, 197–207. [Google Scholar] [CrossRef]
- Bala, B.K.; Arshad, F.M.; Noh, K.M. System Dynamics. Modelling and Simulation; Springer: Singapore, 2017; 278p. [Google Scholar] [CrossRef]
- Saltelli, A.; Tarantola, S.; Campolongo, F.; Ratto, M. Sensitivity Analysis in Practice. A Guide to Assesing Scientific Models; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2004; 219p. [Google Scholar] [CrossRef]
- Saltelli, A.; Ratto, M.; Tarantola, S.; Campolongo, F. Sensitivity analysis practices: Strategies for model-based inference. Reliab. Eng. Syst. Saf. 2006, 91, 1109–1125. [Google Scholar] [CrossRef]
- Saltelli, A.; Ratto, M.; Andres, T.; Campolongo, F.; Cariboni, J.; Gatelli, D.; Tarantola, S. Global Sensitivity Analysis: The Primer; John Wiley & Sons: Hoboken, NJ, USA, 2008. [Google Scholar] [CrossRef]
- Baez-Gonzalez, A.D.; Fajardo-Díaz, R.; Garcia-Romero, G.; Osuna-Ceja, E.; Kiniry, J.R.; Meki, M.N. High Sowing Densities in Rainfed Common Beans (Phaseolus vulgaris L.) in Mexican Semi-Arid Highlands under Future Climate Change. Agronomy 2020, 10, 442. [Google Scholar] [CrossRef]
- Karimzadeh Soureshjani, H.; Nezami, A.; Kafi, M.; Tadayon, M. Responses of two common bean (Phaseolus vulgaris L.) genotypes to deficit irrigation. Agric. Water Manag. 2019, 213, 270–279. [Google Scholar] [CrossRef]
- Omae, H.; Kumar, A.; Shono, M. Adaptation to high temperature and water deficit in the common bean (Phaseolus vulgaris L.) during the reproductive period. J. Bot. 2012, 2012, 803413. [Google Scholar] [CrossRef]
- Price, K.V.; Storn, R.M.; Lampinen, J.A. Differential Evolution. A Practical Approach to Global Optimization; Springer: Berlin/Heidelberg, Germany, 2005; 538p. [Google Scholar]
- Wallach, D.; Makowski, D.; Jones, J.W.; Brun, F. Working with Dynamic Crop Models: Methods, Tools and Examples for Agriculture and Environment, 3rd ed.; Academic Press: London, UK, 2019. [Google Scholar]
- Krause, P.; Boyle, D.P.; Bäse, F. Comparison of different efficiency criteria for hydrological model assessment. Adv. Geosci. 2005, 5, 89–97. [Google Scholar] [CrossRef]
- Luo, Q. Temperature thresholds and crop production: A review. Clim. Chang. 2011, 109, 583–598. [Google Scholar] [CrossRef]
- Barrios Gómez, E.J.; López Castañeda, C.; Kohashi Shibata, J.; Acosta Gallegos, J.A.; Miranda Colín, S.; Mayek Pérez, N. Avances en el mejoramiento genético del frijol en México por tolerancia a temperatura alta ya sequía. Rev. Fitotec. Mex. 2011, 34, 247–255. Available online: http://www.scielo.org.mx/pdf/rfm/v34n4/v34n4a6.pdf (accessed on 1 April 2024).
- Beebe, S.; Ramirez, J.; Jarvis, A.; Rao, I.M.; Mosquera, G.; Bueno, J.M.; Blair, M.W. Genetic Improvement of Common Beans and the Challenges of Climate Change Crop Adaptation to Climate Change; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2011; pp. 356–369. [Google Scholar]
- Martínez-Gamiño, M.A.; Osuna-Ceja, E.S.; Padilla-Ramírez, J.S.; Acosta-Gallegos, J.A.; Loredo-Osti, C. Tecnología Para la Producción de Frijol en el Norte Centro de México; Libro Técnico No. 4; Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias Publication Series: San Luis Potosí, Mexico, 2008.
- Lynch, J.; van Beem, J.J. Growth and architecture of seedling roots of common bean genotypes. Crop Sci. 1993, 33, 1253–1257. [Google Scholar] [CrossRef]
- Boutraa, T.; Sanders, F.E. Influence of water stress on grain yield and vegetative growth of two cultivars of bean (Phaseolus vulgaris L.). J. Agron. Crop Sci. 2001, 187, 251–257. [Google Scholar] [CrossRef]
- Rai, A.; Sharma, V.; Heitholt, J. Dry bean [Phaseolus vulgaris L.] growth and yield response to variable irrigation in the arid to semi-arid climate. Sustainability 2020, 12, 3851. [Google Scholar] [CrossRef]
- Munier-Jolain, N.G.; Munier-Jolain, N.M.; Roche, R.; Ney, B.; Duthion, C. Seed growth rate in grain legumes I. Effect of photoassimilate availability on seed growth rate. J. Exp. Bot. 1998, 49, 1963–1969. [Google Scholar] [CrossRef]
- Trejo-Zúñiga, E.C.; López-Cruz, I.L.; Ruíz-García, A. Parameter estimation for crop growth model using evolutionary and bio-inspired algorithms. Agrociencia 2013, 47, 671–682. Available online: http://www.scielo.org.mx/pdf/agro/v47n7/v47n7a4.pdf (accessed on 1 April 2024).
- Civicioglu, P.; Besdok, E. A conceptual comparison of the Cuckoo-search, particle swarm optimization, differential evolution and artificial bee colony algorithms. Artif. Intell. Rev. 2013, 39, 315–346. [Google Scholar] [CrossRef]
- Ghassemi-Golezani, K.; Mardfar, R.A. Effects of limited irrigation on growth and grain yield of common bean. J. Plant Sci. 2008, 3, 230–235. [Google Scholar] [CrossRef]
- Ghassemi-Golezani, K.; Abriz, S.F.; Hassanpour-Bourkheili, S. Development of mung-bean seed vigour under different irrigations and plant densities. Int. J. Plant Anim. Environ. Sci. 2014, 4, 208–211. [Google Scholar]
- Masaya, P.; White, J.W. Adaptation to photoperiod and temperature. In Common Beans: Research for Crop Improvement; Schoonhoven, A.V., Voysest, O., Eds.; Centre for Agricultural Bioscience International and Centro Internacional de Agricultura Tropical (Cali, Colombia): Oxford, UK, 1991; pp. 445–500. [Google Scholar]
- Fernandez, F.; Gepts, P.; Lopez, M. Etapas de desarrollo en la planta de frijol. In Frijol: Investigación y producción. Programa de las Naciones Unidas (PNUD); López Genes, M., Fernández, O., Fernando, O., van Schoonhoven, A., Eds.; Centro Internacional de Agricultura Tropical (CIAT): Cali, Colombia, 1985; pp. 61–78. [Google Scholar]
- Bulatewicz, T.; Jin, W.; Staggenborg, S.; Lauwo, S.; Miller, M.; Das, S.; Andresen, D.; Peterson, J.; Steward, D.R.; Welch, S.M. Calibration of a crop model to irrigated water use using a genetic algorithm. Hydrol. Earth Syst. Sci. 2009, 13, 1467–1483. [Google Scholar] [CrossRef]
- Teweldebrhan, A.T.; Burkhart, J.F.; Schuler, T.V. Parameter uncertainty analysis for an operational hydrological model using residual-based and limits of acceptability approaches. Hydrol. Earth Syst. Sci. 2018, 22, 5021–5039. [Google Scholar] [CrossRef]
- White, J.W.; Izquierdo, J. Physiology of yield potential and stress tolerance. In Common Beans. Research for Crop Improvement; van Schoonhoven, A., Voysest, O., Eds.; CAB International: Wallingford, UK, 1991; pp. 287–382. [Google Scholar]
- Silva, V.M.; Andrade, M.J.; Moraes, A.R.; Vieira, N.M.; Alves, J., Jr. Dry matter accumulation by common bean cv. BRS MG Talismã. Bean Improv. Coop. 2005, 48, 154–155. [Google Scholar]
- Lépiz-Ildefonso, R.; Chavarín-Espinoza, I.E.; López-Alcocer JD, J.; Rodríguez-Guzmán, E. Dry matter accumulation during developmental stages of common bean cultivars. Rev. Fitotec. Mex. 2018, 41, 275–282. [Google Scholar] [CrossRef][Green Version]
- Soltani, A.; Alimagham, S.M.; Nehbandani, A.; Torabi, B.; Zeinali, E.; Dadrasi, A.; Zand, E.; Ghassemi, S.; Pourshirazi, S.; Alasti, O.; et al. SSM-iCrop2: A simple model for diverse crop species over large areas. Agric. Syst. 2020, 182, 102855. [Google Scholar] [CrossRef]
- Mathobo, R.; Marais, D.; Steyn, J.M. Calibration and validation of the SWB model for dry beans (Phaseolus vulgaris L.) at different drought stress levels. Agric. Water Manag. 2018, 202, 113–121. [Google Scholar] [CrossRef]
- Dewedar, O.; Plauborg, F.; Marwa, M.; El-shafie, A.; Ragab, R. Improving water saving, yield, and water productivity of bean under deficit drip irrigation: Field and modelling study using the SALTMED model. Irrig. Drain. 2020, 70, 224–242. [Google Scholar] [CrossRef]
- Emam, Y.; Shekoofa, A.; Salehi, F.; Jalali, A.H. Water stress effects on two common bean cultivars with contrasting growth habits. Am. Eurasian J. Agric. Environ. Sci. 2010, 9, 495–499. [Google Scholar]
- Morales-Rosales, E.J.; Escalante-Estrada JA, S.; López-Sandoval, J.A. Crecimiento, índice de cosecha y rendimiento de frijol (Phaseolus vulgaris L.) en unicultivo y asociado con girasol (Helianthus annuus L.). Univ. Cienc. 2008, 24, 1–10. [Google Scholar]
- Acosta-Gallegos, J.A.; Rosales-Serna, R. Biomasa y sus componentes en variedades indeterminadas de frijol. Memoria. In 35a Reunion Anual de PCCMCA; Programa Cooperativo Centroamericano para Mejoramiento de Cultivos y Animales: San Perdo Sula, Honduras, 1989. [Google Scholar]
- Debaeke, P.; Aboudrare, A. Adaptation of crop management to water-limited environments. Eur. J. Agron. 2004, 21, 433–446. [Google Scholar] [CrossRef]
- Kemanian, A.R.; Stöckle, C.O.; Huggins, D.R.; Viega, L.M. A simple method to estimate harvest index in grain crops. Field Crops Res. 2007, 103, 208–216. [Google Scholar] [CrossRef]
- Ranjan, R.; Chandel, A.K.; Khot, L.R.; Bahlol, H.Y.; Zhou, J.; Boydston, R.A.; Miklas, P.N. Irrigated pinto bean crop stress and yield assessment using ground based low altitude remote sensing technology. Inf. Process. Agric. 2019, 6, 502–514. [Google Scholar] [CrossRef]
- Dallacort, R.; de Freitas, P.S.L.; Faria, R.T.; Goncalves, A.C.A.; Rezende, R.; Guimaraes, R.M.L. Simulation of bean crop growth, evapotranspiration and yield in Paraná State by the CROPGRO-Drybean model. Acta Sci. Agron. 2011, 33, 429–436. [Google Scholar] [CrossRef]
- Mompremier, R.; Her, Y.; Hoogenboom, G.; Migliaccio, K.; Muñoz-Carpena, R.; Brym, Z.; Colbert, R.W.; Jeune, W. Modeling the response of dry bean yield to irrigation water availability controlled by watershed hydrology. Agric. Water Manag. 2021, 243, 106429. [Google Scholar] [CrossRef]
- Marrou, H.; Sinclair, T.R.; Metral, R. Assessment of irrigation scenarios to improve performances of Lingot bean (Phaseolus vulgaris) in southwest France. Eur. J. Agron. 2014, 59, 22–28. [Google Scholar] [CrossRef]
- Espadafor, M.; Couto, L.; Resende, M.; Henderson, D.W.; Garcia-Vila, M.; Fereres, E. Simulation of the responses of dry beans (Phaseolus vulgaris L.) to irrigation. Trans. ASABE 2017, 60, 1983–1994. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).