Abstract
Physalis peruviana holds significant economic value, making it crucial to determine optimal cultivation conditions, particularly concerning seed germination under varying water and temperature conditions. Therefore, this study aimed to assess the impacts of heat and water stress on the germination and vigor of P. peruviana seeds. The study was divided into two trials: the first examined the effect of constant temperatures of 10, 15, 20, 25, 30, 35, and 40 °C and alternating temperatures of 20 °C during the dark period and 30 °C during the light period (control) on seed germination and vigor. The second trial investigated germination and vigor under different water and thermal conditions using various osmotic potentials (0; −0.3; −0.6; and −0.9 MPa) and two temperatures (constant 30 °C and alternating 20 °C during the dark period and 30 °C during the light period). Both trials used a 16 h photoperiod. The germination tests revealed optimal (30 °C), moderate (20/30 °C), minimal (20 °C), and inhibited (40 °C) temperatures for the species’ germination. It was found that the first germination count could be conducted on the seventh day after sowing. Low water availability had negative effects on seed germination and vigor, especially at osmotic potentials below −0.45 and −0.61 MPa, combined with temperatures of 30 and 20/30 °C, respectively. Severe water stress, with osmotic potentials equal to or below −0.9 MPa, completely inhibited seed germination and vigor. Reduction in water potential and increased temperature resulted in a significant decrease in the percentage, speed, and quality of P. peruviana germination. These findings indicate that the species does not tolerate extreme temperatures, whether low (less than 15 °C) or high (greater than 35 °C), nor water stress with osmotic potentials lower than −0.45 MPa, much less the combination of these factors.
1. Introduction
Physalis peruviana L. belongs to the Solanaceae family, commonly known as camapu, peruvian groundcherry, goldenberry, or cape gooseberry. This vegetable crop is native to the Andean region and is distinguished by its high added value, with significant potential for food and nutraceutical purposes [1]. The genus comprises over 120 species distributed mainly in the tropical and temperate regions of the Americas [2,3]. These species are easily recognizable due to their fused calyx that encloses and protects the fruits from herbivores and environmental factors [4,5].
In Brazil, P. peruviana is consumed as a high-end fruit and produced on a small scale by small and medium-sized producers [6,7]. However, the species is gaining more prominence in the market, with its fruits being sold fresh or used by food and medicinal industries [7,8]. Therefore, it is considered a versatile crop with significant commercial potential, offering an excellent income alternative [6,7]. Nevertheless, there are no reports of large-scale P. peruviana production in Brazil due to the specific soil and climatic characteristics of each region, as well as a lack of technical information about the crop [1,9].
Although there are studies related to the training system [10], fertilization [11], post-harvest quality of fruits [12], and production costs [13] of the species, research related to crop development remains limited. Therefore, studies are needed, especially in regions where abiotic factors pose significant obstacles to agricultural production. Understanding how P. peruviana responds to different environmental conditions makes it possible to identify more suitable regions for its growth and development.
Due to climatic irregularities, plants are subject to at least some form of abiotic stress throughout their development cycle, consequently affecting their production [14,15,16]. It is believed that physiological changes in P. peruviana occur at both metabolic and morphological levels to adapt to adverse environments [17]. One way to assess the effect of these stressful environmental factors is through seed studies. Physiological and biochemical analyses reveal the initial effects of these stresses during seed germination and seedling emergence [18,19]. Although germination is directly related to the genetic makeup of the species, its expression can be modified by stresses and environmental variations [20]. Therefore, understanding seed physiology is crucial, particularly due to the diverse responses they may exhibit.
The germination of P. peruviana seeds is influenced by several environmental and genetic factors, and it is crucial to consider these influences when choosing the most suitable method to break seed dormancy. The germination of P. peruviana seeds is influenced by various environmental and genetic factors, and it is crucial to consider these influences when choosing the most suitable method to break seed dormancy. However, Nunes et al. [21] found that P. peruviana seeds have low dormancy compared to other wild plants. The authors also observed that dormancy was interrupted by temperature variations, explaining the high rate of germinated seeds in the field in their study. Farook et al. [22] evaluated several Physalis species and found that they exhibit physiological dormancy (PD), which can be released by keeping the seeds in the presence of water. Additionally, gibberellic acid (GA3) can also be used to stimulate the germination of Physalis species seeds.
The recommended sowing depth for P. peruviana is around 0.5 to 1.5 cm deep, with P. peruviana requiring well-drained soil rich in organic matter [21,23]. Regarding the vitality of the seeds, it is essential to ensure that they are mature and healthy at the time of harvest. Mature seeds of P. peruviana are usually orange-yellow in color and have a firm texture. Seed viability can be affected by factors such as inadequate storage, presence of pathogens and seed age. Therefore, correct storage under appropriate temperature and humidity conditions is crucial to maintain seed vitality for longer periods [24,25]. Furthermore, it is important to consider that seed germination and vitality may vary between different varieties of P. peruviana and in different growing environments.
Seed vigor and quality are the most important factors for establishing field crops and, consequently, for achieving productive success. It is through these attributes that producers can achieve uniform stand establishment, which facilitates management and cultural practices [26]. Furthermore, using high-quality seeds adapted to different biotic and abiotic factors proves to be a viable alternative for cultivation in environmentally challenging regions [27]. This fact is particularly relevant for countries with vast expanses, where a variety of climatic conditions are present, such as Brazil.
The evaluation of seeds subjected to factors such as water deficit and temperature fluctuations, or the interaction of these, is crucial and recommended for defining optimal cultivation conditions and ensuring productive success [28,29]. These factors are among the most common and impactful, capable of reducing seedling emergence percentage, leading to uneven stand establishment, and consequently, reduced production [30].
Regarding water deficit, this is one of the most limiting factors in agriculture due to the importance of water in activating various metabolic processes [31]. During germination, water plays a decisive role in enzymatic reactions, solubilization, and transport of metabolites, acting as a reagent in the hydrolytic digestion of seed reserve tissues [32]. Water stress can also lead to the toxicity of certain elements such as sodium and chlorine, which can cause physiological disturbances in plants and even be lethal [33].
Temperature variations can significantly influence physiological and biochemical processes in seeds, directly affecting the initial growth of crops [34]. It is known that the further away from the optimal temperature, the greater the stress on seeds. At critical levels, this can cause damage to seed structure, including altering the inherent metabolic activities of the germination process, which are highly dependent on specific enzymatic systems [35,36].
In light of the above, the economic importance of P. peruviana is evident, highlighting the need to find suitable conditions of water availability and temperature to ensure seed germination. Therefore, studies correlating these two factors are essential to characterize the adaptability mechanisms to these conditions, enabling improved performance of the crop in the field. It is hypothesized that optimal germination conditions, with appropriate levels of water and temperature, will result in greater plant vigor, contributing to more efficient and sustainable field production. Thus, the objective of this study was to evaluate the effects of water and thermal stress on the germination and vigor of P. peruviana seeds.
This study allows us to assess the ability of P. peruviana to tolerate environmental stresses, such as high or low temperatures and water content, providing insights into its resilience under different climatic conditions. The findings from these studies can guide farmers in choosing the most suitable cultivation practices, such as optimal seeding timing and irrigation management, to maximize the production and quality of P. peruviana crops in both open-field and greenhouse cultivation.
2. Materials and Methods
2.1. Experiment Conduct and Plant Material
The experiment was conducted in the Seed Laboratory of the Department of Agronomy at the Federal University of Viçosa (UFV) located in Viçosa, Minas Gerais. The seeds used in the study were obtained from ripe fruits collected from crops in the municipality of Diamantina, Minas Gerais. After harvesting, the fruits were placed in sieves to wash away excess pulp. The extracted seeds were then arranged on trays with paper towels and air-dried under ambient conditions (approximately 25 °C) for 24 h.
To assess the quality and viability of the seeds, a germination test was conducted following the Rules for Seed Analysis [37]. Four replicates of 25 seeds each were used, and placed in germbox containers measuring 11 × 11 × 3.5 cm. Within these containers, the seeds were placed between two layers of germitest paper, previously moistened with distilled water at a ratio of 2.5 times the weight of the dry paper. The seeds were then placed in a B.O.D. (biological oxygen demand) chamber set to an alternating temperature of 20/30 °C, with a 16 h photoperiod. During the 16 h photoperiod, the temperature was maintained at 30 °C, and during the 8-hour dark period, the temperature was kept at 20 °C. Vigor parameters and the percentage of germinated seeds (Gmax) were measured daily between the 5th and 28th day after test installation, with a germination rate of 96% observed. After confirming viability, the seeds were used for the study. The criteria for biological germination in the test and its derivations were seeds that exhibited radicle protrusion of at least 2 mm in length.
The study was divided into two trials: (i) the influence of different temperatures on the germination and vigor of P. peruviana seeds; and (ii) the influence of the combination of stressful factors (temperature and water restriction) on the germination and vigor of P. peruviana seeds.
Both trials were conducted in the laboratory setting. The seeds were placed for germination in germbox containers, following the same characteristics and procedures described previously for testing seed quality and viability. Subsequently, they were placed in B.O.D. set to the temperatures specified in the trials, with 90% humidity and a 16 h photoperiod, for a duration of 28 days. Four replicates of 25 seeds each were used for each treatment, totaling 100 seeds per treatment.
2.2. Influence of Temperature on Seed Germination and Vigor
To assess the effect of temperature on the germination of P. peruviana seeds, constant temperatures of 10, 15, 20, 25, 30, 35, and 40 °C were tested. Additionally, an alternating temperature of 20/30 °C was tested, following the same procedure described in the germination test in Section 2.1. According to the recommendations in the Rules for Seed Analysis [37], the ideal germination temperature for P. alkekengi L. and P. pubescens L. seeds is an alternating temperature of 20/30 °C. Since there is limited information available specifically for P. peruviana, this temperature range was used as the control in the study.
Additionally, the temperature causing a 50% reduction in germination (median temperature), the optimal germination temperature for the species, and the temperature resulting in no germination (0%) were also determined.
2.3. Combination of Temperature and Water Restriction on Seed Germination and Vigor
To assess the effect of simultaneous stresses on the germination of P. peruviana seeds, different combinations of water potentials and temperatures were tested. The trials combined the control temperatures (20/30 °C) and optimal germination temperature (30 °C) identified in the first trial with osmotic potentials of 0; −0.3; −0.6; and −0.9 MPa. It is noteworthy that preliminary tests were conducted earlier, revealing that the minimum temperature (20 °C) resulted in no germination when combined with different osmotic potentials. Similarly, it was found that the osmotic potential of −1.2 MPa prevented seed germination for all tested temperatures.
To simulate the effect of water restriction on the germination of P. peruviana seeds, solutions of polyethylene glycol (PEG 6000) were used as outlined in Table 1, following the procedure also performed by Ma et al. [38]. The concentrations of PEG 6000 used were diluted using a magnetic stirrer. Distilled water was used in the 0 MPa treatment.

Table 1.
Concentration in g L−1 of polyethylene glycol (PEG 6000) estimated at temperatures of 20, 20/30, and 30 °C to achieve different levels of osmotic potential.
Paper towels were replaced every two days to maintain the the solution’s potential and prevent pathogen development.
2.4. Variables Analyzed and Statistical Analysis
In both trials, for the evaluation of seed germination and vigor, the germbox containers were examined daily, with the first count conducted on the fifth day and the final count on the twenty-eighth day after the test setup. Germination assessments took place around 8:00 a.m. The evaluations included counting the number of seeds showing root protrusion and the number of normal seedlings. The number of normal seedlings was counted at the end of the test.
The parameters assessed were: percentage of germinated seeds (Gmax), germination speed index (GSI), time to reach 10% germination (T10), time to reach 50% germination (T50), time to reach 90% germination (T90), germination uniformity (UnifG), mean germination time (MGT), mean germination rate (MGR), and germination synchrony (Sync). The equations used to calculate each characteristic, as described by Silva et al. [39], are presented in Table 2. The calculations were performed using R software version 4.0.5 [40] and the SeedCalc package version 1.0 [39].
Table 2.
Functions contained in the SeedCalc package for calculating indices using daily seed count in the germination/emergence test.
The first count test was conducted concurrently with the germination test, involving the recording of the percentages of normal seedlings observed during the initial count of the germination test, performed from the fifth to the twenty-eighth day after sowing, following the guidelines of the Seed Analysis Rules [37].
The data obtained from all evaluations were subjected to analysis of variance (ANOVA). When the F-test was significant, mean tests at a 5% probability level were used for qualitative factors. The Tukey test was employed for comparing all treatments, while the Dunnett test was used for comparisons with the control treatment. For quantitative factors, the model selection was based on the significance of regression coefficients, using a t-test at a 5% probability level, the coefficient of determination (R2), and biological phenomena. Statistical analyses were conducted using R software version 4.0.5 [40].
3. Results and Discussion
3.1. Experiment I: Effect of Temperatures on Seed Germination and Vigor
Table 3 presents the summaries of the analysis of variance (ANOVA) for the germination variables of P. peruviana. The effect of temperature is observed in all analyzed variables. This table also includes the comparison of results obtained using the alternating temperature of 20/30 °C (control) with the other treatments involving constant temperatures. This approach was necessary due to the nature of the control treatment, as the alternating temperature could not be treated as a quantitative factor; therefore, only the constant temperatures were considered. Thus, the alternating temperature of 20/30 °C was compared using mean tests with the other temperatures, which in turn were analyzed using regression.
Table 3.
Means of percentage of germinated seeds (Gmax), germination speed index (GSI), mean germination time (MGT), time required for 10%, 50%, and 90% germination (T10, T50, and T90), mean germination rate (MGR), germination synchrony (Sync), germination uniformity (UnifG), and number of normal seedlings (NNS) of P. peruviana subjected to different temperatures.
For the Gmax results, it was observed that the constant temperatures between 15 and 35 °C did not differ significantly from the control (20/30 °C). However, the other temperatures differed from the control, with the 10 °C treatment showing a substantial decrease in germination potential. Additionally, at 40 °C, no germination was observed, contrary to the findings of Ozaslan et al. [47] evaluating other species of the genus Physalis. These authors found germination rates greater than 70% for P. angulata and P. philadelphicade.
For the other evaluated variables, the constant temperatures of 10, 15, and 40 °C provided different means compared to the control treatment (20/30 °C). Similarly, to germination, the constant temperature of 40 °C resulted in null values for all other analyzed variables. Overall, the use of constant temperatures between 20 and 35 °C yielded germination variables similar to the control, except for germination synchrony and uniformity. For these variables, all constant temperature treatments differed significantly from the control.
Figure 1 presents the germination variables as a function of temperatures. It is observed that temperature had a quadratic effect on the Gmax of P. peruviana (Figure 1A). According to the fitted regression equation, the temperature of 25.4 °C resulted in the highest Gmax, reaching approximately 97.6%. This aligns with the findings of Carvalho and Nakagawa [48], indicating that the optimal temperature for germination of tropical species is typically between 20 and 30 °C, which is consistent with the results of this study.
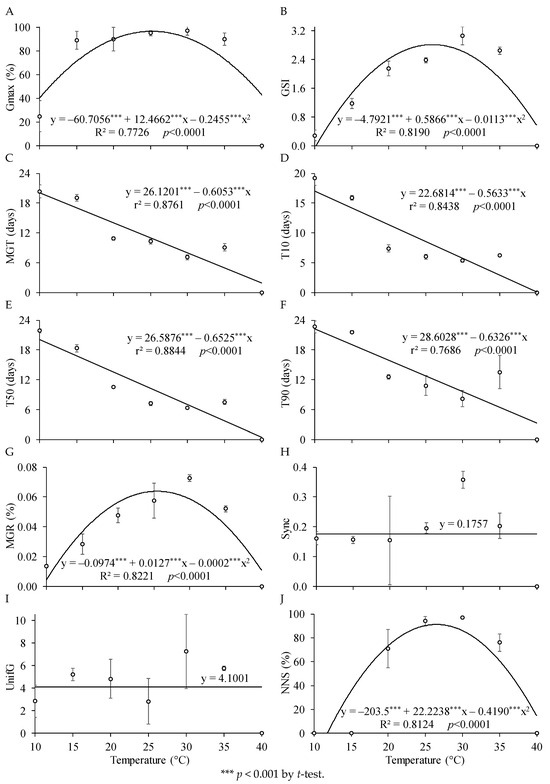
Figure 1.
(A) Percentage of germinated seeds (Gmax), (B) germination speed index (GSI), (C) mean germination time (MGT), time required for germination of (D) 10, (E) 50, and (F) 90% of seeds (T10, T50, and T90), (G) mean germination rate (MGR), (H) germination synchrony (Sync), (I) germination uniformity (UnifG), and (J) number of normal seedlings (NNS) of P. peruviana as a function of different temperatures.
Nunes et al. [21] found a temperature of up to 32 °C suitable for germination of P. peruviana seeds. Ozaslan et al. [47] achieved germination percentages of P. angulata and P. philadelphica seeds close to 100% at a temperature of 35 °C. For some species in the genus, a constant temperature of 25 °C was sufficient to promote high germination of P. ixocarpa L. [49], while an average temperature of 25 °C (alternating between 20 and 30 °C) was adequate to promote high germination of P. angulata L. [50]. These results indicate that each species has a temperature spectrum within which the seed can germinate. For P. peruviana, it is presumed that this temperature requirement is an ecological adaptation, given that this species occurs spontaneously in a wide range of habitats in tropical countries, each exhibiting an optimal germination temperature [50].
For the germination speed index (GSI), it is observed that temperature had a quadratic effect (Figure 1B). According to the regression equation, the temperature of 26.0 °C provided the highest GSI, resulting in a value of 2.82. These results are consistent with those found by Carvalho and Nakagawa [48], who stated that germination will be faster at higher temperatures up to a certain limit. It is noted that when temperatures are higher or lower than those that provided the highest GSI, there may be a reduction in the speed of the germination process, exposing the seeds to adverse factors that can lead to their inviability [51]. Low temperatures can reduce metabolic activity, resulting in a decrease in the number and speed of germinating seeds, as well as an increase in the time required for the process to occur. On the other hand, exposure to higher temperatures can degrade seeds due to membrane integrity loss, thereby becoming a gateway for pathogens [48].
The optimal germination temperature differs from the temperature at which germination speed is highest. In this regard, Cafaro et al. [52] found that the best results for beaver bean (Ricinus communis L.) seed germination were at a temperature of 25 °C, but the highest germination speed was observed at 32 °C. Considering the results of the present study and by Cafaro et al. [52], it is evident that the temperature that enhances GSI is higher than that which maximizes germination.
Additionally, Figure 1 shows that temperatures had a negative linear effect on the variables of mean germination time (MGT) and time required for germination of 10, 50, and 90% of seeds (T10, T50, and T90), meaning that higher temperatures led to shorter germination times. Analyzing these results in isolation, it is understood that the reduction in germination time due to higher temperatures could bring benefits to the cultivation of P. peruviana.
Equations of regression could not be fitted for germination synchrony (Sync) and germination uniformity (UnifG) as functions of seed temperature. For the mean germination rate (MGR) and number of normal seedlings (NNS), quadratic models were fitted. According to the regression equation in Figure 1J, a temperature of 26.5 °C maximized NNS, resulting in a value of 91.2%. Thus, it is observed for P. peruviana that although low temperatures (around 15 °C) and high temperatures (around 40 °C) promoted seed germination, there was subsequently no formation of normal seedlings, rendering these temperatures ineffective for germination. Therefore, 20 and 35 °C were considered the minimum and maximum temperatures for germination of P. peruviana, respectively.
Considering all the variables analyzed so far, the temperature range between 25 °C and 30 °C was considered optimal for the germination of P. peruviana. The ideal germination temperature should vary within this range optimal for seedling emergence and establishment, providing the best combination of germination percentage and speed [53]. When developmental temperatures are kept below or above the ideal range for germination of a particular species, the reorganization of the cellular membrane system can become slower, affecting seed vigor [54]. This occurs because temperatures outside the optimal range cause oxidative damage to the seeds, leading to failure of germination or the production of abnormal seedlings [35]. Some temperature conditions can inhibit seed germination and compromise enzymatic activities related to reserve mobilization, which are crucial for the germination process [31]. Under conditions of low temperatures, for example, seed imbibition may occur, but embryo growth will not occur for most species [55,56], as observed in this study for P. peruviana subjected to temperatures of 10 °C and 15 °C.
On the other hand, high temperatures can denature proteins and alter membranes, leading to progressive seed deterioration. This is mainly due to the influence on enzyme activity and limitations in accessing oxygen [32]. Furthermore, the deleterious effects of high temperatures on plants are related to numerous biochemical and metabolic alterations, such as reduced photosynthetic activity in the chloroplast and oxidative phosphorylation in the mitochondria [57]. This characteristic can be considered a diagnostic trait of the species under study, as this temperature range is capable of enabling the emergence and establishment of seedlings satisfactorily [20,39].
According to the accumulated germination data over the 28-day period after sowing (Figure 2), it was observed that for most tested temperatures, the peak of germination occurred before the fourteenth day after sowing. This phenomenon can be explained by nutrient reserves in the seeds, indicating an intrinsic characteristic of the seeds. For seeds exposed to a temperature of 30 °C, germination began on the 5th day after sowing and, three days later (8 days after sowing), had already stabilized at approximately 90% germination. This characteristic is beneficial and contributes to considering this temperature in the germination test of P. peruviana. A delayed germination process can lead to problems in the later stages of crop development. According to Reed et al. [20], plants from seeds with delayed germination may exhibit physiological and morphological alterations, becoming more sensitive to stress and showing limited competitive ability for resources.
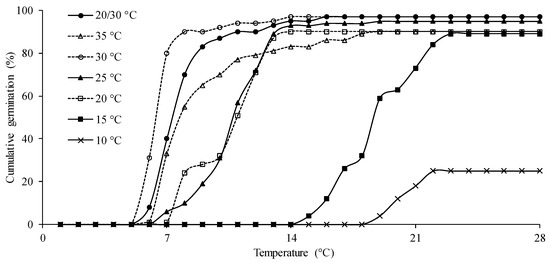
Figure 2.
Cumulative germination of P. peruviana seeds subjected to different temperatures.
Figure 2 also shows that for temperatures of 20/30, 20, 25, and 35 °C, germination began on the sixth day after sowing, with stability in this process around the 14th day. For the lower temperatures of 10 and 15 °C, germination only began on the 19th and 15th days after sowing, respectively. It is noted that at 28 days, for the temperature of 10 °C, there was less than 20% germination of P. peruviana. Despite this germination, as seen in Figure 1J, there was no subsequent formation of normal seedlings, rendering these temperatures ineffective for germination.
Also noted in Figure 2 is that most treatments showed germination onset between five and seven days after sowing. Therefore, it is suggested that the seventh day after sowing is the appropriate time to conduct the first germination count to assess the vigor of P. peruviana. Studies with other species suggest the same, as in the case of Cedrela odorata (Spanish cedar) [58], Vicia sativa (common vetch) [59], Combretum leprosum (mofumbo) [60], and Citrullus unatus (watermelon) [61]. In this test, complementary responses to those provided by the germination test are sought, generating more consistent information [62]. This assessment is considered a simple vigor test and assumes that the most vigorous seeds germinate first.
As observed, temperature is one of the main factors affecting germination, both in percentage and speed. This is because it is directly related to the rate of water absorption, influencing the biochemical reactions that are crucial for the germination process [48]. Although temperatures of 20/30 °C and in the range between 25 and 30 °C are considered favorable for P. peruviana seed germination, at 30 °C, as observed, there was rapid stabilization of germinated seeds and germination speed, reducing exposure to biotic and abiotic factors that could compromise their development. Thus, the temperature of 30 °C stood out as the optimal germination temperature for this study.
3.2. Experiment II: Germination and Vigor of Seeds under Different Moisture and Thermal Conditions
Table 4 presents summaries of the analysis of variance for variables related to the germination of P. peruviana seeds subjected to different temperatures and osmotic potentials. Significant interactions were found between the temperature and osmotic potential factors for all evaluated characteristics. Consequently, subsequent analyses were conducted to further investigate the studied factors.
Table 4.
Means of percentage of germinated seeds (Gmax), germination speed index (GSI), mean germination time (MGT), time required for 10%, 50%, and 90% germination (T10, T50, and T90), mean germination rate (MGR), germination uniformity (UnifG), and germination synchrony (Sync) of P. peruviana seeds submitted to different temperatures (T) and osmotic potentials (ѱs).
Table 4 also presents the mean comparisons of variables related to the germination of P. peruviana seeds subjected to different temperatures. The temperatures tested were 20/30 °C, considered the control in the first experiment, and 30 °C, which was the optimal germination temperature for P. peruviana seeds. Higher Gmax was observed at the alternating temperature of 20/30 °C when the osmotic potential was equal to or greater than −0.3 MPa, indicating water restriction. The superior performance under alternating temperatures can be attributed to the findings of Nunes et al. [21], which indicated that temperature variations interrupted the dormancy of P. peruviana. When there was no water restriction imposed (osmotic potential of 0 MPa), the germination of P. peruviana seeds was similar at both studied temperatures.
It was observed that at an osmotic potential of −0.9 MPa, the temperature of 30 °C led to no germination of P. peruviana seeds. This was also observed for other evaluated characteristics. These results indicate that the constant temperature of 30 °C is more sensitive to a water deficit. Therefore, low osmotic potentials can contribute to modifying the optimal temperature, altering the speed and percentage responses of P. peruviana seed germination. These results suggest that in the presence of other limiting factors, germination occurs within a narrower temperature range.
For osmotic potentials of 0, −0.3, and −0.6 MPa, a different behavior of the germination speed index (GSI) concerning germination was observed. With water restriction provided by osmotic potentials of −0.3 and −0.6 MPa, temperatures did not affect the GSI. Without water restriction (osmotic potential of 0 MPa), the GSI was higher in the treatment where seeds were subjected to a temperature of 30 °C. This indicates that the reduction in germination was compensated by an increase in GSI.
For the other characteristics related to germination speed, the same behavior observed for GSI was noted. Temperatures did not cause differences in the values of mean germination time (MGT), times required for germination of 10% (T10) and 90% (T90) of germination, and mean germination rate (MGR) of seeds subjected to osmotic potentials of −0.3 and −0.6 MPa. However, without water restrictions (osmotic potential of 0 MPa), these values were higher in the treatment where seeds were subjected to a temperature of 30 °C.
The enhanced responses of GSI, MGT, T10, T90, and MGR to a temperature of 30 °C in treatments without osmopriming occur because higher temperatures facilitate faster seed imbibition (the first stage of the germination process). This is due to the fact that elevated temperatures reduce water viscosity and increase water kinetic energy, which not only aids in seed imbibition but also benefits the speed of metabolic activities occurring within the seed. However, despite higher temperatures accelerating water absorption by the seed, each of the reactions involved in the germination process has its own thermal requirements, primarily depending on the activity of specific enzymatic systems [48].
Commercial production of P. peruviana will be facilitated if cultural practices can be applied continuously and uniformly. For this to happen, uniformity in plant development is necessary, starting from seed germination. Therefore, parameters of germination quality are important and should be considered in seed analyses. It is observed in Table 4 that temperatures did not affect the values of uniformity (UnifG) and germination synchrony (Sync) of seeds subjected to osmotic potentials of −0.3 and −0.6 MPa. In the non-osmoprimed treatments, temperatures did not affect UnifG, and seeds subjected to 30 °C showed higher Sync values.
It is evident that the germination characteristics of P. peruviana showed better performance when seeds were subjected to a constant temperature of 30 °C without water restriction. These results are consistent with those obtained in Experiment 1 of this study. Under water restriction, with seeds subjected to osmotic potentials of −0.3 and −0.6 MPa, the alternating temperature of 20/30 °C provided a higher germination percentage and did not affect the speed and quality parameters of germination. Additionally, the temperature of 20/30 °C enabled seed germination up to an osmotic potential of −0.9 MPa, whereas the temperature of 30 °C supported germination up to −0.6 MPa.
Figure 3 presents the responses of the studied variables to water restriction provided by osmopriming at two seed temperatures. Regardless of the temperature, quadratic models were able to be fitted for the germination of P. peruviana seeds based on osmotic potentials. It is observed that decreasing osmotic potential led to a reduction in seed germination. The maximum germination values were obtained near an osmotic potential of 0 MPa. Farook et al. [22] observed that Physalis species exhibit physiological dormancy, which can be released by soaking the seeds in water. This phenomenon may have contributed to the findings of the present study. According to the regression equations (Figure 3A), seed germination remained above 80% until osmotic potentials of −0.6140 and −0.4494 MPa for temperatures of 20/30 °C and 30 °C, respectively.
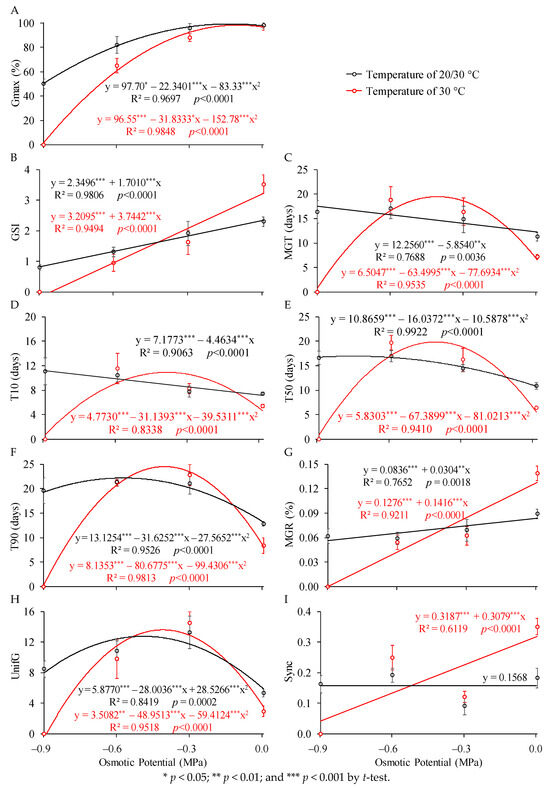
Figure 3.
(A) Percentage of germinated seeds (Gmax), (B) germination speed index (GSI), (C) mean germination time (MGT), time required for germination of (D) 10, (E) 50, and (F) 90% of seeds (T10, T50, and T90), (G) mean germination rate (MGR), (H) germination uniformity (UnifG), and (I) germination synchrony (Sync) of P. peruviana as a function of different osmotic potentials for different temperatures.
The reduction in osmotic potential leads to a decrease in water uptake by the seeds, which can compromise the sequence of events in the germination process, thereby reducing the germination percentage. Plants subjected to water restriction may exhibit different responses to this stress, which can be defined by the characteristic drought tolerance trait of each species [63]. Cellular responses to stress include changes in cell division and the cell cycle, modifications in the endomembrane system, as well as alterations in cell wall architecture [31].
Silva and Azerêdo [64] also observed greater seed germination of two cactus species (Cereus jamacaru and Pilosocereus pachycladus) at a water potential of −0.2 MPa. Germination decreased from −0.2 MPa to −0.8 MPa, reaching zero at −1.0 and −1.2 MPa. Germination decreased from −0.2 MPa to −0.6 MPa, reaching zero at −0.8 and −1.0 MPa. Oliveira et al. [65] evaluated different osmotic potentials (0; −0.1; −0.2; −0.3; −0.4; −0.5; and −0.6 MPa) and temperatures (25, 30, 35, and 20/30 °C) on the germination of Simira gardneriana seeds. The authors observed that decreasing osmotic potential and increasing temperature led to a reduction in seed germination.
Regarding the germination velocity parameters, it is observed that the decrease in osmotic potential linearly reduced the GSI (Figure 3B) and MGR (Figure 3G). These results are consistent with Felix et al. [66], who evaluated Leucaena leucocephala seeds subjected to water deficit. They found that both the speed and percentage of seed germination decreased when exposed to lower water potentials.
Water is a crucial requirement for initiating germination, as this process is activated only when an adequate amount of water is absorbed, allowing for metabolic activity. Under conditions of reduced water potential, seeds may have enough water to initiate the germination process (Phases I and II) without, however, protruding the radicle and initiating primary root growth (Phase III) [31]. Consequently, germination velocity is reduced, extending the period required to reach the minimum water content needed for embryonic axis emergence.
As previously reported, P. peruviana seeds were more sensitive to osmopriming at a temperature of 30 °C. This is confirmed by the equations fitted for GSI (Figure 3B) and MGR (Figure 3G), where the absolute values of the slope coefficients for the equations related to the 30 °C temperature were greater than those for 20/30 °C.
For MGT (Figure 3C), the osmotic potentials had a linear effect on seeds subjected to a temperature of 20/30 °C and a quadratic effect on those at 20 °C. The quadratic model adjustment was due to the null value of MGT at the osmotic potential of −0.9 MPa, which reduced the time at the expense of a lower germination percentage. For T10 (Figure 3D), osmopriming provided a linear effect at temperature 20/30 °C and a quadratic effect at 30 °C. For T50 and T90 (Figure 3E,F), regardless of the temperature to which the seed was subjected, osmopriming had a quadratic effect on this variable.
For the germination quality parameters, it is observed that the reduction in osmotic potential had a quadratic effect on UnifG (Figure 3H), regardless of seed temperature. For Sync (Figure 3I), in seeds subjected to a temperature of 30 °C, the decrease in osmotic potential led to a linear reduction in its values. In the alternate temperature of 20/30 °C, it was not possible to fit a regression model to the Sync data.
In general, higher temperatures cause enzymatic changes, increasing their requirements and accelerating the respiratory rate of seeds. Thus, for seeds exposed to more negative potentials at 30 °C, there was likely an acceleration of metabolic reactions during the initial stages of germination. Subsequently, this may have led to seed deterioration, preventing them from maintaining hydration conditions to complete the germination process.
For the temperature of 20/30 °C, the thermal fluctuation may have contributed to a more uniform germination process. Cochrane [53] highlights that seeds of many species require daily temperature fluctuations to germinate adequately. Seeds that respond to temperature fluctuations have enzymatic mechanisms that function at different temperatures [67]. According to Ahmad et al. [36], this response corresponds to an adaptation to natural environmental fluctuations.
In Figure 4, the vigor data are presented, determined by germination as a function of days after sowing (DAS) and osmotic potentials. It was observed that as osmotic potentials became more negative, there was a gradual increase in the number of days to the onset of germination. For seeds maintained at a temperature of 20/30 °C (Figure 4A), it is observed from the regression equation that the germination of P. peruviana started at 4, 6, 8, and 11 DAS for osmotic potentials of 0, −0.3, −0.6, and −0.9 MPa, respectively. At the temperature of 30 °C (Figure 4B), P. peruviana germination began at 1, 5, and 10 DAS for osmotic potentials of 0, −0.3, and −0.6 MPa, respectively.
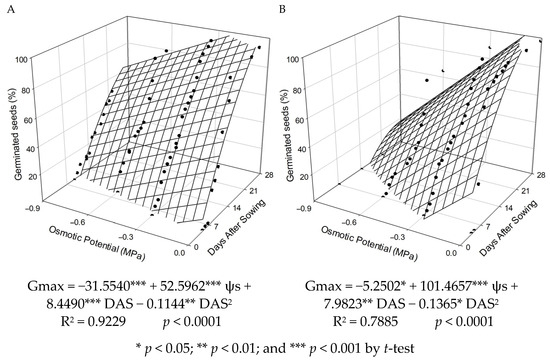
Figure 4.
Percentage of germinated seeds (Gmax) of P. peruviana seeds as a function of different osmotic potentials (ѱs) and days after sowing (DAS) under the following temperature conditions: (A) alternating 20/30 °C and (B) constant 30 °C.
It is observed that seeds exposed to a temperature of 30 °C showed longer times to initiate the germination process when water restriction was increased. It is also noted that the more negative the osmotic potentials used, the longer the time required for the germination process to start. These results suggest that in environments where at least one of these factors acts extremely on the seed germination process, germination is inhibited, even if the other factors are not limiting for the species.
To achieve 80% germination for seeds maintained at a temperature of 20/30 °C, it was necessary, according to the regression equation (Figure 4A), 13, 15, and 18 days after sowing (DAS) for osmotic potentials of 0, −0.3, and −0.6 MPa, respectively. For the temperature of 30 °C (Figure 4B), it required 13 and 22 DAS for osmotic potentials of 0 and −0.3 MPa, respectively. Similar to the present study, germination time was also increased for seeds of white clover (Trifolium repens) in response to water deficit [68]. This was likely due to water restriction promoting longer stays of the seeds in Phase II of the germination process, as reported by Bewley et al. [31].
Some studies suggest that reducing germination time is an advantage in plant establishment [69]. The faster a seed initiates Phase III of germination, where radicle protrusion is visible, the better and faster its establishment in the field and, consequently, exposure to unfavorable conditions [70,71].
In summary, the performance of P. peruviana seeds under water and thermal stress is affected starting from germination. Seedling growth is directly linked to water availability in the substrate and temperature. The results observed at osmotic potentials below −0.3 MPa highlight the detrimental effect of low water availability on the development of P. peruviana seeds. Regarding the poorer outcomes observed at 30 °C in the presence of osmotic solution, this could be attributed to a condition that is mild but not within the ideal range of temperature and water restriction required by the species. These findings are ecologically and commercially important, as they demonstrate the specific water conditions the seeds of this species need to germinate.
4. Conclusions
P. peruviana exhibits good germination performance and vigor under thermal and water stress conditions, indicating its potential for successful cultivation in environments where abiotic factors influence crop development.
The most suitable temperature for P. peruviana germination is 30 °C, with 20/30 °C being a viable median condition. Temperatures of 20 °C and 40 °C represent the thermal limits, with the latter preventing germination.
An osmotic potential threshold for drought tolerance is −0.9 MPa at 30 °C, while lower osmotic potentials than −0.9 MPa are critical at 20/30 °C. Osmotic potentials up to −0.45 MPa at 30 °C and −0.61 MPa at 20/30 °C maintain germination rates above 80%.
Adverse abiotic stresses, particularly the interaction between reduced osmotic potentials and elevated temperatures, significantly reduce the speed and quality of P. peruviana germination.
Germination tests should begin on the seventh day after sowing, with the final count on the twenty-eighth day to ensure an accurate assessment of germination performance.
Understanding the optimal and limiting thermal and water conditions for P. peruviana germination is crucial for optimizing cultivation practices. Effective management of temperature and water conditions can significantly enhance plant productivity and quality, contributing to more efficient and resilient agriculture.
This investigation underscores the importance of understanding P. peruviana seeds’ responses to environmental variables, providing a solid foundation for management strategies that minimize the negative impacts of abiotic stresses and maximize cultivation success.
Author Contributions
Conceptualization, E.M.d.F., L.J.d.S. and F.F.d.C.; methodology, E.M.d.F. and L.J.d.S.; validation, E.M.d.F., C.N.G., L.J.d.S. and F.F.d.C.; formal analysis, E.M.d.F. and F.F.d.C.; investigation, E.M.d.F.; resources, L.J.d.S. and F.F.d.C.; data curation, E.M.d.F.; writing—original draft preparation, E.M.d.F., C.N.G. and F.F.d.C.; writing—review and editing, F.F.d.C.; visualization, E.M.d.F., C.N.G., L.J.d.S. and F.F.d.C.; supervision, C.N.G. and F.F.d.C.; project administration, F.F.d.C.; funding acquisition, F.F.d.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Coordination for the Improvement of Higher Education Personnel, Brazil (CAPES), Finance Code 001 and the National Council for Scientific and Technological Development, Brazil (CNPq), Process 308769/2022-8.
Data Availability Statement
The original contributions presented in the study are included in the article, further inquiries can be directed to the corresponding author.
Acknowledgments
We thank the Department of Agriculture Engineering (DEA) and the Graduate Program in Agricultural Engineering (PPGEA) of the Federal University of Viçosa (UFV) for supporting the researchers.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Diniz, F.O.; Chamma, L.; Novembre, A.D.L.C. Germination of Physalis peruviana L. seeds under varying conditions of temperature, light, and substrate. Rev. Ciênc. Agron. 2020, 51, e20166493. [Google Scholar] [CrossRef]
- Silva, D.F.; Pio, R.; Soares, J.D.R.; Nogueira, P.V.; Peche, P.M.; Villa, F. The production of Physalis spp. seedlings grown under different-colored shade nets. Acta Sci. Agron. 2016, 38, 257–263. [Google Scholar] [CrossRef]
- Feng, S.; Zheng, K.; Jiao, K.; Cai, Y.; Chen, C.; Mao, Y.; Wang, L.; Zhan, X.; Ying, Q.; Wang, H. Complete chloroplast genomes of four Physalis species (Solanaceae): Lights into genome structure, comparative analysis, and phylogenetic relationships. BMC Plant Biol. 2020, 20, 242. [Google Scholar] [CrossRef] [PubMed]
- Ponce, O.V.; Martínez, J.S.; Tavares, M.D.P.Z.; Mares, L.E.V. Traditional management of a small-scale crop of Physalis angulata in Western Mexico. Genet. Resour. Crop Evol. 2016, 63, 1383–1395. [Google Scholar] [CrossRef]
- Melo, E.S.; Assis, F.A.; Carvalho, F.J.; Assis, G.A.; Rodrigues, F.A. Does diatomaceous earth improve agronomic characteristics and induce resistance to arthropod pest in physalis? Rev. Ciênc. Agrovet. 2022, 21, 504–515. [Google Scholar] [CrossRef]
- Silva, L.S.; Villa, F.; Silva, D.F.; Silva, E.C.; Ritter, G.; Eberling, T. Micropropagation of Physalis species with economic potential. Ceres 2021, 68, 521–529. [Google Scholar] [CrossRef]
- Barroso, N.S.; Fonseca, J.S.T.; Ramos, C.A.S.; Nascimento, M.N.; Soares, T.L.; Pelacani, C.R. Impact of the maturity stage on harvest point of fruits and physiological quality of Physalis peruviana L. seeds. Rev. Bras. Frutic. 2022, 44, e-848. [Google Scholar] [CrossRef]
- Guiné, R.P.F.; Gonçalves, F.J.A.; Oliveira, S.F.; Correia, P.M.R. Evaluation of phenolic compounds, antioxidant activity and bioaccessibility in Physalis peruviana L. Int. J. Fruit Sci. 2020, 20, 470–490. [Google Scholar] [CrossRef]
- Freitas, E.M.; Vital, T.N.B.; Guimarães, G.F.C.; Silveira, F.A.; Gomes, C.N.; Cunha, F.F. Determination of the permanent wilting point of Physalis peruviana L. Horticulturae 2023, 9, 873. [Google Scholar] [CrossRef]
- Lima, J.E.; Cruz, M.C.M.; Alves, D.A.; Santos, N.C.; Guimarães, A.G. Pruning, training system, and climate conditions for the perennial cultivation of physalis. Pesq. Agropec. Bras. 2021, 56, e01850. [Google Scholar] [CrossRef]
- Souza, H.B.F.; Silva, M.S.; Rodrigues, M.S.; Nóbrega, R.S.A.; Conceição, A.L.S. Physalis peruviana L. cultivated in dystrocohesive yellow latosol is responsive to organic fertilization. Rev. Cienc. Agrar. 2021, 64, 1635. [Google Scholar]
- Hayati, P.; Hosseinifarahi, M.; Abdi, G.; Radi, M.; Taghipour, L. Melatonin treatment improves nutritional value and antioxidant enzyme activity of Physalis peruviana fruit during storage. Food Meas. 2023, 17, 2782–2791. [Google Scholar] [CrossRef]
- Bendlin, L.; Senff, C.O.; Franco, C.K.; Souza, A.; Veiga, C.P.; Duclós, L.C. Agribusiness management of Physalis peruviana L. fruit in Brazil. Bulg. J. Agric. Sci. 2016, 22, 691–704. [Google Scholar]
- Chaudhry, S.; Sidhu, G.P.S. Climate change regulated abiotic stress mechanisms in plants: A comprehensive review. Plant Cell Rep. 2022, 41, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Krishna, R.; Karkute, S.G.; Ansari, W.A.; Jaiswal, D.K.; Verma, J.P.; Singh, M. Transgenic tomatoes for abiotic stress tolerance: Status and way ahead. Biotech 2019, 9, 13205. [Google Scholar] [CrossRef] [PubMed]
- Sandrini, M.; Nerva, L.; Sillo, F.; Balestrini, R.; Chitarra, W.; Zampieri, E. Abiotic stress and belowground microbiome: The potential of omics approaches. Int. J. Mol. Sci. 2022, 23, 1091. [Google Scholar] [CrossRef] [PubMed]
- Velandia, D.M.; Barrera, E.C. Effect of salt stress on growth and metabolite profiles of cape gooseberry (Physalis peruviana L.) along three growth stages. Molecules 2021, 26, 2756. [Google Scholar] [CrossRef] [PubMed]
- Yadav, S.P.; Bharadwaj, R.; Nayak, H.; Mahto, R.; Singh, R.K.; Prasad, S.K. Impact of salt stress on growth, productivity and physicochemical properties of plants: A Review. Int. J. Chem. Stud. 2019, 7, 1793–1798. [Google Scholar]
- Johnson, R.; Puthur, J.T. Seed priming as a cost effective technique for developing plants with cross tolerance to salinity stress. Plant Physiol. Biochem. 2021, 162, 247–2571. [Google Scholar] [CrossRef]
- Reed, R.C.; Bradford, K.J.; Khanday, I. Seed germination and vigor: Ensuring crop sustainability in a changing climate. Heredity 2022, 128, 450–459. [Google Scholar] [CrossRef]
- Nunes, A.L.; Sossmeier, S.; Gotz, A.P.; Bispo, N.B. Germination eco-physiology and emergence of Physalis peruviana seedlings. J. Agr. Sci. Technol. 2018, 8, 352–359. [Google Scholar] [CrossRef]
- Farooq, S.; Onen, H.; Ozaslan, C.; El-Shehawi, A.M.; Elseehy, M.M. Characteristics and methods to release seed dormancy of two ground cherry (Physalis) species. J. Appl. Res. Med. Aromat. Plants 2021, 25, 100337. [Google Scholar] [CrossRef]
- Freitas, E.M.; Silva, G.H.; Guimarães, G.F.C.; Vital, T.N.B.; Vieira, J.H.; Silveira, F.A.; Gomes, C.N.; Cunha, F.F. Evapotranspiration and crop coefficient of Physalis peruviana cultivated with recycled paper as mulch. Sci. Hortic. 2023, 320, 112212. [Google Scholar] [CrossRef]
- Souza, C.L.M.; Souza, M.O.; Oliveira, R.S.; Pelacani, C.R. Physalis peruviana seed storage. Rev. Bras. Cienc. Agrar. 2016, 20, 263–268. [Google Scholar] [CrossRef]
- Tenorio, M.L.O.; Dekker, M.; van Boekel, M.A.J.S.; Verkerk, R. Evaluating the effect of storage conditions on the shelf life of cape gooseberry (Physalis peruviana L.). LWT 2017, 80, 523–530. [Google Scholar] [CrossRef]
- Costa, C.J.; Meneghello, G.E.; Jorge, M.H.A.; Costa, E. The importance of physiological quality of seeds for agriculture. Colloq. Agrar. 2021, 17, 102–119. [Google Scholar] [CrossRef]
- Lamichhane, J.R.; Debaeke, P.; Steinberg, C.; You, M.P.; Barbetti, M.J.; Aubertot, J.N. Abiotic and biotic factors affecting crop seed germination and seedling emergence: A conceptual framework. Plant Soil 2018, 432, 1–28. [Google Scholar] [CrossRef]
- Xue, X.; Du, S.; Jiao, F.; Xi, M.; Wang, A.; Xu, H.; Jiao, Q.; Zhang, X.; Jiang, H.; Chen, J.; et al. The regulatory network behind maize seed germination: Effects of temperature, water, phytohormones, and nutrients. Crop J. 2021, 9, 718–724. [Google Scholar] [CrossRef]
- Staniak, M.; Krok, E.S.; Kocira, A. Responses of soybean to selected abiotic stresses—Photoperiod, temperature and water. Agriculture 2023, 13, 146. [Google Scholar] [CrossRef]
- Araújo, E.D.; Assis, M.O.; Guimarães, C.M.; Araújo, E.F.; Borges, A.C.; Cunha, F.F. Superabsorbent polymers and sanitary sewage change water availability during the cowpea emergence phase. Nativa 2024, 12, 37–48. [Google Scholar] [CrossRef]
- Bewley, J.D.; Bradford, K.; Hilhorst, H.; Nonogaki, H. Seeds: Physiology of Development, Germination and Dormancy, 3rd ed.; Springer: New York, NY, USA, 2013; 405p. [Google Scholar]
- Prazeres, C.S.; Coelho, C.M.M. Osmolyte accumulation and antioxidant metabolism during germination of vigorous maize seeds subjected to water deficit. Acta Sci. Agron. 2020, 42, e42476. [Google Scholar] [CrossRef]
- Shahid, M.A.; Sarkhosh, A.; Khan, N.; Balal, R.M.; Ali, S.; Rossi, L.; Gómez, C.; Mattson, N.; Nasim, W.; Sanchez, F.G. Insights into the physiological and biochemical impacts of salt stress on plant growth and development. Agronomy 2020, 10, 938. [Google Scholar] [CrossRef]
- Kamatchi, K.A.M.; Anitha, K.; Kumar, K.A.; Senthil, A.; Kalarani, M.K.; Djanaguiraman, M. Impacts of combined drought and high-temperature stress on growth, physiology, and yield of crops. Plant Physiol. Rep. 2024, 29, 28–36. [Google Scholar] [CrossRef]
- Bouteau, H.M. The seed and the metabolism regulation. Biology 2022, 11, 168. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, M.; Waraich, E.A.; Skalicky, M.; Hussain, S.; Zulfiqar, U.; Anjum, M.Z.; Habib, R.M.; Brestic, M.; Ratnasekera, D.; Tamayo, L.L.; et al. Adaptation strategies to improve the resistance of oilseed crops to heat stress under a changing climate: An overview. Front. Plant Sci. 2021, 12, 767150. [Google Scholar] [CrossRef] [PubMed]
- Brasil, Ministério da Agricultura e Reforma Agrária. Regras para Análise de Sementes, 1st ed.; SNDA/DNDV/CLAV: Brasília, Brazil, 2009; 399p. [Google Scholar]
- Ma, L.; Wei, J.; Han, G.; Sun, X.; Yang, X. Seed osmopriming with polyethylene glycol (PEG) enhances seed germination and seedling physiological traits of Coronilla varia L. under water stress. PLoS ONE 2024, 19, e0303145. [Google Scholar] [CrossRef] [PubMed]
- Silva, L.J.; Medeiros, A.D.; Oliveira, A.M.S. SeedCalc, a new automated R software tool for germination and seedling length data processing. J. Seed Sci. 2019, 41, 250–257. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing, 4.0.5 version; R Development Core Team: Vienna, Austria, 2021; Available online: www.r-project.org/ (accessed on 27 March 2024).
- ISTA. The germination test. In International Rules for Seed Testing; ISTA: Zurich, Switzerland, 2015. [Google Scholar] [CrossRef]
- Maguire, J.D. Speed of germination-aid selection and evaluation for seedling emergence and vigor. Crop Sci. 1962, 2, 176–177. [Google Scholar] [CrossRef]
- Farooq, M.; Basra, S.M.A.; Ahmad, N.; Hafeez, K. Thermal hardening: A new seed vigor enhancement tool in rice. J. Integr. Plant Biol. 2005, 47, 187–193. [Google Scholar] [CrossRef]
- Labouriau, L.G. Uma nova linha de pesquisa na fisiologia da germinação das sementes. In Congresso Nacional de Botânica; Sociedade Botânica do Brasil: Porto Alegre, Brazil, 1983. [Google Scholar]
- Primack, R.B. Variation in the phenology of natural populations of montane shrubs in New Zealand. J. Ecol. 1980, 68, 849–862. [Google Scholar] [CrossRef]
- Demilly, D.; Ducournau, S.; Wagner, M.H.; Dürr, C. Digital imaging of seed germination. Plant Im. Anal. 2014, 1, 147–164. [Google Scholar] [CrossRef]
- Ozaslan, C.; Farooq, S.; Onen, H.; Ozcan, S.; Bukun, B.; Gunal, H. Germination biology of two invasive physalis species and implications for their management in arid and semi-arid regions. Sci. Rep. 2017, 7, 16960. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, N.M.; Nakagawa, J. Sementes: Ciência, Tecnologia e Produção, 5th ed.; FUNEP: Jaboticabal, Brazil, 2012; 590p. [Google Scholar]
- Barroso, N.S.; Fonseca, J.S.T.; Nascimento, M.N.; Soares, T.L.; Pelacani, C.R. Physiological quality of Physalis ixocarpa Brot. ex Hornem seeds in relation to maturation stage and growing season. Pesqui. Agropecu. Trop. 2023, 53, e74090. [Google Scholar] [CrossRef]
- Santiago, W.R.; Gama, J.S.N.; Benedito, C.P.; Sousa, E.M.; Torres, S.B. Ecophysiological aspects of the germination of Physalis angulata L. seeds. Caatinga 2023, 36, 980–987. [Google Scholar] [CrossRef]
- Silva, M.S.A.; Yamashita, P.O.M.; Rossi, A.A.B.; Carvalho, M.A.C.; Concenço, G.; Sá, M.E. Influence of light and temperature on seed germination of Macroptilium lathyroides. South Am. J. Basic Educ. Tech. Technol. 2020, 7, 311–324. [Google Scholar]
- Cafaro, V.; Alexopoulou, E.; Cosentino, S.L.; Patanè, C. Germination response of different castor bean genotypes to temperature for early and late sowing adaptation in the Mediterranean regions. Agriculture 2023, 13, 1569. [Google Scholar] [CrossRef]
- Cochrane, J.A. Thermal requirements underpinning germination allude to risk of species decline from climate warming. Plants 2020, 9, 796. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Shen, Y.; Shi, F. Effect of temperature, light, and storage time on the seed germination of Pinus bungeana Zucc. Ex Endl.: The role of seed-covering layers and abscisic acid changes. Forests 2020, 11, 300. [Google Scholar] [CrossRef]
- Visscher, A.M.; Lorenzo, E.C.; Toorop, P.E.; Silva, L.J.; Yeo, M.; Pritchard, H.W. Pseudophoenix ekmanii (Arecaceae) seeds at suboptimal temperature show reduced imbibition rates and enhanced expression of genes related to germination inhibition. Plant Biol. 2022, 22, 1041–1051. [Google Scholar] [CrossRef]
- Li, S.; Yang, W.; Guo, J.; Li, X.; Lin, J.; Zhu, X. Changes in photosynthesis and respiratory metabolism of maize seedlings growing under low temperature stress may be regulated by arbuscular mycorrhizal fungi. Plant Physiol. Biochem. 2020, 154, 1–10. [Google Scholar] [CrossRef]
- Catiempo, R.L.; Photchanachai, S.; Bayogan, E.R.V.; Aree, C.W. Impact of hydropriming on germination and seedling establishment of sunflower seeds at elevated temperature. Plant Soil Environ. 2021, 67, 491–498. [Google Scholar] [CrossRef]
- Maldonado, S.S.; Salanueva, C.A.O.; Mattana, E.; Ulian, T.; Way, M.; Lorenzo, E.C.; Aranda, P.D.D.; Saade, R.L.; Valdéz, O.T.; Arevalo, N.I.R.; et al. Thermal time and cardinal temperatures for germination of Cedrela odorata L. Forests 2019, 10, 841. [Google Scholar] [CrossRef]
- Zhang, Y.; Tao, Q.; Zhang, R.; Ma, Y.; Xing, J.; Zhou, S.; Liu, Y.; Meng, F.; Sun, J. A seed vigour test based on radicle emergence during germination at 5 °C for four forage species. Seed Sci. Technol. 2023, 51, 361–369. [Google Scholar] [CrossRef]
- Leal, C.C.P.; Torres, S.B.; Dantas, N.B.D.L.; Aquino, G.S.M.; Alves, T.R.C. Water stress on germination and vigor of “mofumbo” (Combretum leprosum Mart.) seeds at different temperatures. Rev. Ciênc. Agron. 2020, 51, e20186357. [Google Scholar] [CrossRef]
- Cheng, W.; Zhang, A.; Zhu, J.; Li, Y.; Wang, P. Study of salicylic acid influence on seedling growth and nitrogen metabolism in watermelon (Citrullus lanatus L.). J. Food Eng. Technol. 2018, 7, 47–53. [Google Scholar] [CrossRef]
- Ortiz, T.A.; Gomes, G.R.; Takahashi, L.S.A. Standardization of the accelerated aging test methodology for snap bean seed to differentiate batches by physiological quality. Int. J. Veg. Sci. 2024, 30, 1–7. [Google Scholar] [CrossRef]
- Feng, W.; Lindner, H.; Robbins, N.E.; Dinneny, J.R. Growing out of stress: The role of cell-and organ-scale growth control in plant water-stress responses. Plant Cell 2016, 28, 1769–1782. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.H.C.S.; Azerêdo, G.A.D. Germination of cactus seeds under saline stress. Caatinga 2022, 35, 79–86. [Google Scholar] [CrossRef]
- Oliveira, F.N.; Oliveira, J.R.; Torres, S.B.; Freitas, R.M.O.; Nogueira, N.W. Germination and initial development of Simira gardneriana seedling under water stress and at different temperatures. Rev. Bras. Eng. Agríc. Ambient. 2017, 21, 333–338. [Google Scholar] [CrossRef]
- Felix, F.C.; Araújo, F.S.; Silva, M.D.; Ferrari, C.S.; Pacheco, M.V. Water and thermal stress on the germination Leucaena leucocephala (Lam.) de wit seeds. Rev. Bras. Cienc. Agrar. 2018, 13, e5515. [Google Scholar] [CrossRef]
- Oliveira, A.K.M.; Ribeiro, J.W.F.; Pereira, K.C.L.; Silva, C.A.A. Effects of temperature on the germination of Diptychandra aurantiaca (Fabaceae) seeds. Acta Sci. Agron. 2013, 35, 203–208. [Google Scholar] [CrossRef]
- Ling, Y.; Zhao, Y.; Cheng, B.; Tan, M.; Zhang, Y.; Li, Z. Seed priming with chitosan improves germination characteristics associated with alterations in antioxidant defense and dehydration-responsive pathway in white clover under water stress. Plants 2022, 11, 2015. [Google Scholar] [CrossRef] [PubMed]
- Bano, H.; Bhat, J.I.A.; Lone, F.A.; Noor, F.; Bhat, M.A.; Nazir, N. Effect of phytohormones and other dormancy breaking chemicals on seed germination of Inula racemosa. Int. J. Curr. Microbiol. Appl. Sci. 2019, 8, 866–876. [Google Scholar] [CrossRef]
- Ataíde, G.M.; Borges, E.E.L.; Gonçalves, J.F.C.; Guimarães, V.M.; Flores, A.V. Physiological changes during hydration of Dalbergia nigra ((Vell.) Fr. All. ex Benth.) seeds. Ciênc. Florest. 2016, 26, 615–625. [Google Scholar] [CrossRef]
- Gomes, J.P.; Oliveira, L.M.; França, C.S.S.; Dacoregio, H.M.; Bortoluzzi, R.L.C. Morphological characterization of seedlings during the germination seeds of Psidium cattleianum and Acca sellowiana (myrtaceae). Ciênc. Florest. 2015, 25, 1035–1041. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).