Abstract
Many cultivars of annual ornamental horticulture have great phenotypic qualities but are less tolerant to the effects of current climate change and urbanization. A good example of this in Hungarian ornamental breeding is the Rudbeckia hirta cultivar “Őszifény”. The objective of this study was to determine whether gamma mutation breeding (using 5 Gy, 10 Gy, 30 Gy doses) can be employed to enhance genetic variability and create mutants with novel trait values. Furthermore, it is important to ascertain whether the altered genetic traits are proportionate to the observed changes in phenotype. As an original and innovative aspect of the research, this will assist in establishing appropriate dose rates for the species. Changes induced by gamma radiation have been mapped using morphological, histological, and genetic methods. The radiation dose of 5 Gy resulted in plants with the most favorable characteristics, including significant increases in branching and flower number. Additionally, the phenomenon of hormesis was observed. Beyond these, the dose of 30 Gy also had a significant effect because it resulted in plants with many flowers and bushy habits, making them suitable for use as ornamental potted plants. The 10 Gy dose resulted in a heterogeneous stand but showed the greatest genetic variation. These results may contribute to future breeding work and may pave the way for further urban application of R. hirta in the light of urbanization.
1. Introduction
The demand for ornamental plants is expected to increase in the future due to the growth of global trade [1,2,3]. Nowadays, interactions with urban green infrastructure largely define people’s encounters with nature within cities [4]. The continuous introduction of new species and cultivars remains a focal point in ornamental horticulture [5].
1.1. Possibilities for Mutation Breeding in Ornamental Plants
The genetic variability of ornamental plants cannot be considered as satisfactory, which means that some older cultivars are not able to compete with the current challenges. For instance, they are less suited to perform well in highly stressed urban environments. The maintenance of these cultivars, as well as the introduction of new ones, is driven by market needs, which require rapid techniques that provide high genetic variability, such as mutation breeding.
Mutation breeding is more successful in ornamentals because phenotypic traits, such as changes in flower color, shape, and size and growth habits, are easily detected. In addition, the heterozygous nature of many ornamental plants offers high mutation frequencies. In the last two decades, ion beam radiation has emerged as an effective and unique mutagen in ornamental plant development, as it produces higher mutation frequencies compared to X-ray and gamma radiation [6]. Scientific interest in mutation breeding has declined dramatically over recent decades and has shifted toward the development and application of molecular technologies, and plant breeding has become less important [7]. Nevertheless, these methods are sometimes laborious and complex, especially when trying to improve desired traits without inducing pleiotropic effects. Recently, targeted genome editing (TGE) technology has been used to engineer nucleases, including meganucleases, zinc finger nucleases (ZFN), transcription activator-like effector nucleases (TALEN), and regularly interspaced short palindromic repeat (CRISPR) nucleases to improve traits in economically important crops. TGE has emerged as a new plant breeding tool, representing an alternative approach to classical breeding but with greater mutagenic efficiency [8].
Mutagenesis techniques have been integrated into in vitro breeding and other molecular biology technologies, such as molecular markers and high-throughput mutation screening [9]. Mutagenesis has been used to introduce new genetic variations in ornamental plant breeding for several decades [10,11]. This is why ornamental plants seem to be ideal for mutation breeding, as many traits of economic importance, i.e., flower traits or growth habits, can be easily monitored after mutagenic treatment [12]. Practical mutation breeding has now become a routine technique in many vegetatively propagated ornamental plants and is used by commercial breeding companies [7].
Many ornamental species are heterozygous and often vegetatively propagated, allowing for the detection, selection, and conservation of mutants of the M1 generation. Appropriate mutation induction strategies, such as the use of fractionated doses and repeated irradiation, combined with in vitro breeding techniques, can lead to rapid success in homozygous polyploid species. Despite its random nature, mutation induction remains an attractive method for generating genetic variability in ornamental plants when the desired characteristics are expected within the genetic range of a given species.
1.2. The Most Efficient Irradiation Methods
Different forms of irradiation, including gamma, X-ray, and electron beams, have been investigated for their effectiveness in ornamental horticulture. The potential applications of these, with a focus on their benefits, mechanisms, and potential uses are outlined, with a wide range of references from recent studies.
Gamma irradiation is an excellent method because of its deep penetration and ability to induce mutations uniformly in plant tissues. This type of irradiation has been widely used to create new plant cultivars with desirable traits, such as unique flower colors and shapes and enhanced disease resistance. For example, studies have shown that gamma irradiation can induce desirable mutations in chrysanthemums, resulting in new cultivars with enhanced ornamental value [13].
X-ray radiation, another efficient method, works on similar principles to gamma radiation but uses high-energy photons produced by X-ray machines. The advantage of this method is that they can be produced without using radioactive materials, making them safer and more environmentally friendly. Research has shown that X-ray radiation can effectively induce mutations in ornamental plants such as orchids and lilies, resulting in new, more spectacular cultivars [14]. In addition, studies by Jo et al. [15] have shown that X-ray radiation can improve the ornamental properties of plants while maintaining their overall health and vigor.
Electron beam (e-beam) irradiation is characterized by high energy and low penetration depth, which makes it suitable for surface treatment. Electron beam irradiation has been shown to be effective in inducing beneficial mutations in ornamental species and controlling surface pests without causing significant damage to the plants. Experiments conducted by Kim et al. [16], for example, showed that e-beam irradiation could induce desirable gene changes in petunias, resulting in new flower colors and patterns.
Comparative studies have been conducted to evaluate the effectiveness of different irradiation methods in ornamental nurseries. Kumar et al. [17] compared the effects of gamma, X-ray, and e-beam irradiation on marigolds and found that all three methods were effective in inducing mutations, with gamma irradiation exhibiting the highest mutation frequency. In another study, Maluszynski et al. [18] compared the effectiveness of gamma and X-ray irradiation in inducing mutation in gladiolus and found that both methods were equally effective, although X-ray irradiation offered logistical advantages due to the absence of radioactive waste.
Irradiation is a powerful tool in ornamental horticulture, offering solutions for creating new plant cultivars, controlling pests, and improving the aesthetic appearance of plants. Gamma, X-ray, and electron beams each have unique advantages that make them suitable for specific applications. Ongoing research and technological developments are expected to further optimize these methods, making irradiation an integral part of sustainable ornamental horticultural practices.
1.3. Genetic Variability through Mutation Breeding in the Genus Rudbeckia
The most extensive breeding efforts of the genus have been carried out with R. hirta. It became an important annual cultivar after the introduction of improved cultivars by Goldsmith Seeds in the 1960s. There are vegetatively propagated tetraploid cultivars and seed-grown diploid cultivars. The genus has strong self-incompatibility and pseudogamy, a form of apomixis, which appears to be a common reproductive pathway [19].
For some species (R. laciniata, R. nitida, R. maxima, R. triloba), a compact habit is of great interest as it would broaden the possibilities for use in more restricted urban environments. The disease resistance of R. fulgida is also desirable, as problems with Septoria and Ramularia and leaf spot infections are occasionally encountered [20]. A number of examples show successful mutation breeding in the genus Rudbeckia, for instance, to reduce the growth of the tetraploid cultivar R. subtomentosa “Henri Eilers” [21], and, in in vitro rearing, increasing the concentration reduced the survival and regeneration of the callus [22]. However, mutation breeding has also been applied to rooted cuttings of R. hirta. Shukla et al. [23] found that irradiation resulted in reduced plant height and diseased leaves, and differences in pollen shape and size were observed in irradiation-induced abnormal flowers. At lower irradiance (5 Gy), inflorescence color was lighter, whereas at 10 Gy irradiance, striped flowers were observed on shoots.
1.4. The R. hirta Cultivar Used in the Experiments
R. hirta is a popular ornamental species that has been used for decades. The species and cultivars of R. hirta are grown as annuals and are widely used as bedding plants and in container cultivation, as well as cut flowers [19]. R. hirta is an ornamental plant of the Asteraceae family and is also used in landscaping and for soil erosion prevention [24], and it may also have considerable potential for use as a potted plant [25]. The adaptability of R. hirta plays a major role in its distribution [26]. R. hirta includes diploid (2n = 2x = 38) and tetraploid (2n = 4x = 76) species, with a diverse range of flower colors and forms [19]. The following four subspecies of R. hirta have been described: R. hirta var. floridana, R. hirta var. hirta, R. hirta var. pulcherrima, and R. hirta var. angustifolia [27]. R. hirta species have a mature height of 0.5–1.0 m, and tetraploid species typically have larger flowers and greater height [28], despite the fact that it is short-lived and susceptible to certain diseases, including Cercospora leaf spot (Cercospora sp.) and Rhizoctonia (Rhizoctonia sp.) infection [26,29].
In our experiment, we treated “Őszifény”, a popular annual cultivar of R. hirta L., with varying doses of gamma radiation. Our objective is to investigate and evaluate the effects of these applied doses through morphological, histological, and genetic studies that can inform future breeding programs. Rudbeckia species are an important component of built landscapes, green spaces that represent a significant economic activity. Their wild species are common breeding materials because they retain their ease of propagation and market needs [30].
1.5. Directions and Possibilities for Mutation Breeding of R. hirta
The genus Rudbeckia is genetically understudied, with no specific molecular markers currently available. A single study utilized RAPD markers to investigate genetic diversity and crossing potential among R. fulgida var. sullivantii populations with cultivated cultivars [31]. In this study, we applied two commonly used marker techniques to assess the effectiveness of different gamma radiation doses while detecting polymorphism at the DNA level. Start codon targeted (SCoT) markers rely on short conserved regions flanking the ATG start codon, which is conserved in all plant genes. The PCR uses single 18-mer primers [32]. The conserved DNA-derived polymorphism (CDDP) technique is based on short conserved amino acid sequences in proteins. This method utilizes a single 15- to 19-mer primer for PCR and binds at multiple sites within the plant genome due to the presence of these conserved gene sequences [33]. The applied marker techniques do not require prior sequence knowledge, are designed for highly conserved gene sequences, and can distinguish genetic variations in a specific gene that link to a specific trait. SCoT and CDDP markers are similar to RAPD but offer a reproducible approach for detecting polymorphism between different genotypes due to higher annealing temperatures (50 °C). Based on recent study data from the past five years, it is evident that gamma radiation-induced mutation breeding has become one of the most important methods in modern breeding practices. In ornamental plant breeding specifically, this method holds particular significance as it enables the development of new cultivars with novel phenotypic traits and urban tolerance.
2. Materials and Methods
2.1. Experiment Conditions
R. hirta “Őszifény”, which was used in our experiments, is maintained in the gene bank of the Hungarian University of Agricultural and Life Sciences by our research group. This Hungarian cultivar was bred in 1989 and has since become an heirloom cultivar. Despite the abiotic stress tolerance of the cultivar, its use in public areas has been significantly reduced over the past 20 years, the reasons being that it has few branches and thus is less bushy than the newer cultivars, and it only develops 5–15 flowers per plant, in contrast to the newer cultivars (Figure 1).

Figure 1.
R. hirta “Őszifény” characteristics: (a) plant shape, typical inflorescence color, and leaf shape; (b) treated plant form with 5 Gy gamma treatment.
The experiment was carried out from 2021 to 2022. The greenhouse and the field are located in Budapest, right next to each other, at the Budatétény Research Institute of the Hungarian University of Agricultural and Life Sciences, Budapest, Hungary, with GPS coordinates of 47.407508, 19.013425.
It has a continental climate. There are 1830–1950 h of sunshine per year with summer sunshine of around 760 h. The average annual temperature is 11 °C. The frost-free period is between 15 April and 23–25 October. The annual absolute temperature maximum is 32–33 °C, and the absolute minimum in winter is around −17 °C. There is an annual rainfall of 650 mm, of which 450–500 mm falls in the summer half-year. The soil is composed of alluvium from the Danube alluvium and the slope of the stake. The soil is loamy and has a medium humus content [34]. For greenhouse growing, the commercially available Klassmann Potgrond P peat mixture was used.
The seeds were collected in 2020 and treated with several doses at the Seibersdorf Plant Breeding and Genetics Laboratory station (International Atomic Energy Agency Laboratories (IAEA)) in January 2021. As the irradiation was prepared for us for research purposes, the seed is not commercially available. The seeds were treated once with one of the following doses according to the laboratory’s protocol:
- Control;
- 5 Gy;
- 10 Gy;
- 30 Gy.
Seedlings grown from seed were grown in greenhouse conditions and then in field conditions from May 2021 to December 2021. The 12-week-old seedlings with 5–7 leaves were planted in the open ground after a period of acclimatization.
A random block design was used for the measurements. A total of 30 plants per group were tested in 3 replicates. The M0 generation was used for the measurements, which were grown by irradiating the seeds of mother plants since the primary effect of gamma radiation is best monitored in this generation.
No nutrient supplementation was performed during the rearing period. Irrigation was applied every two days at 3 dL/plant.
Morphological measurements were made on 3 September 2021, as this is when the plants reached full flowering. There was no difference in the onset of flowering between the groups. Full flowering also occurred at the same time. The number of branches and length of the root system were measured manually and with a manual ruler. The plant was laid on the table, the ruler was placed next to it, and the measurement was taken accordingly. The length was measured from the root neck to the root tip (length of roots). Both the fresh and dried mass were measured. The number of flowers was measured as well—for this survey, we counted the number of fully open and opening flowers on the plant. A Kern PCB 6000-1 (Precimer, Budapest, Hungary) analytical laboratory balance was used to measure mass. The plant parts were dried in a VEB MLW Labortechnik Ilmenau (Tech-solute GmbH, Bruchsal, Germany) drying oven for 120 h at 40 °C.
2.2. Histological Measurements
For the histological analyses, the plants were selected randomly from the groups when they were in full bloom, in total, 20 plants from each group. Collecting samples during this period allowed us to examine plant parts that were mature but not showing signs of aging. In the case of leaves and branches, the samples were taken from the middle part of the branch, which had not yet started to senesce. Due to the large number of images taken, the most representative images of the plant groups were selected to show the most important characteristics.
Leaf cross-sections were taken from 5 leaves per plant, with several sampling points per leaf. Each leaf was cleaned with distilled water and then sectioned mechanically with a hand microtome. The fresh sections were examined and photographed to avoid any distortions that may have occurred during fixation and to obtain as accurate a picture of the changes as possible. For the examination of leaf cross-section with light microscopy, the Euromex bScope BS.1153-PLi microscope (Euromex Microscopen B.V., Arnhem, The Netherlands) with a compatible camera (Levenhuk m1400 plus—Levenhuk, Inc., Budapest, Hungary) was used for this study. Due to the nature of the sections, PLi 4/0.1 lenses were used, which provided a magnification of 40×. The eyepiece was of type and size WF120×/20.2.3.
2.3. Genetic Analyses
The morphological characteristics have to be observed during the flowering period when the most important characteristics of ornamental plants—flower shape, color, size, and plant habit—are the most visible. The collection of leaf samples for DNA isolation also was performed at this time when medium-sized and mature leaves (5–10 cm size) were randomly collected. This gave a complete and realistic picture of the condition of the plants. Leaf samples were kept frozen until use. Genomic DNA was isolated from frozen leaf samples using NucleoSpin Plant II, Mini kit (MachereyNagel, GmbH & Co. KG, Düren, Germany) according to the manufacturer’s instructions. Purified DNA was then quantified using Nanodrop 8000 (Thermo Fisher Scientific Inc., Waltham, MA, USA). Based on our preliminary investigations, a total of 14 SCoT and 21 CDDP primers were evaluated. The names and sequences of the primers that yielded the fragments that can be evaluated are listed in the Results section. PCR amplification was performed in a Bioer GeneExplorer (Bioer, Hangzhou, China) in a 10 μL volume. The reaction mixture contained 10 ng of DNA template, 1 μM of primer, and 0.2 μM of each dNTP, 1x reaction puffer, and 0.5 Unit GoTaq G2 DNA polymerase (Promega, Madison, WI, USA). The PCR profile for SCoT and CDDP markers included 2 min of initial denaturation at 94 °C, followed by 35 cycles of 94 °C denaturation for 30 s, 30 s of annealing at 50 °C, 2 min of extension at 72 °C, and 5 min of final extension at 72 °C. Amplified fragments were separated in 1% agarose gel stained with ethidium-bromide. The visualization of PCR products was carried out under UV light and photographed using the Uvitec Essential V6 gel documentation system (Uvitec Cambridge). The PCRs and gel electrophoresis were conducted in two replicates.
2.4. Statistical Analysis
Results were analyzed, compared, and measured, and differences were tested using the IBM SPSS Statistics 26 program with ANOVA. In all cases, the measured data were analyzed at a 95% reliability (significance) level. After the Levene test has been evaluated, if the Sig. > 0.05, the Tukey’s test was used, and if Sig. < 0.05, the Games–Howell post hoc test was used.
In the case of PCR investigations, the resulting DNS bands were converted to a binary format of 0 and 1. The similarity matrix was determined using Past4.03 software [35]. A clustering analysis was performed with the UPGMA method.
3. Results
3.1. Morphological Characterization
3.1.1. Number of Branches
The morphological results clearly demonstrate distinct differences between the doses in terms of the measured parameters (Table 1); in comparison, no statistically significant difference was found for the higher irradiation doses (10 Gy, 30 Gy) compared to the control, and only the lower dose (5 Gy) showed a significant difference. The control group had an average number of 8 branches, with a significant difference at 5 Gy (11.6 branches). The 10 Gy dose had the lowest average number of branches at 5.5. For the 30 Gy dose, the number of branches was 8.4.

Table 1.
Aggregated average data of R. hirta ‘Őszifény’ with gamma radiation treatment (g: gram). Different letters (a, b, c, d or ab) indicate significantly different groups, as follows: number of branches; number of flowers; root fresh mass; root dry mass; branch fresh mass; and branch dry mass (Games–Howell, p < 0.05). p = 0.000.
3.1.2. Number of Flowers
The number of flowers is a crucial characteristic of ornamental plants. Similar to the branching number results (Table 1), the findings for the treated groups reflected a similar pattern. In comparison to the average number of flowers in the control group (11.2), there was no statistically significant difference in the average flower diameter for plants exposed to a dose of 10 Gy (12.8). However, significantly higher results were observed for groups subjected to doses of 5 Gy (17.8) and 30 Gy (20.5), indicating larger flower diameters.
3.1.3. Root Fresh and Dry Mass
The data obtained for both root fresh and dry mass show clear correlations between the variables. Significant differences exist among the applied doses The mean root fresh mass was 14.49 g in the control group, and statistically significant results were observed for all treated groups. Among these groups, the highest value (37.57 g) was recorded in the group exposed to 5 Gy, while the lowest value (17.38 g) was observed in the group exposed to 10 Gy (Table 1). Similar trends can be seen in terms of root dry mass, as well. Here too, the control group yielded the lowest result (1.98 g), whereas all treated groups exhibited higher values. Notably, the highest root dry mass was obtained from plants exposed to a dose of 5 Gy (5.11 g). Interestingly though, no significant difference was observed between results from groups subjected to doses of 10 Gy (2.65 g) and 30 Gy (3.23 g) (Table 1).
3.1.4. Shoot Fresh and Dry Mass
The results for both fresh and dry mass show a similar trend to the data obtained for root mass (Table 1). Nevertheless, there was no significant difference in shoot mass between the control groups and the groups exposed to 10 Gy, either in terms of fresh or shoot dry mass The highest mass was measured in the groups receiving a dose rate of 5 Gy, which significantly outperformed all other groups.
3.2. Histological Analysis
In addition to the morphological measurements, histological surveys were conducted to further validate the findings. The observed alterations in the structure were influenced by gamma radiation, as evidenced by the differences between the control and treated groups. The histological results align with the observations made during morphological assessments. The tissues of the high-size group, which received a 5 Gy dose of radiation, were more developed than those of the control group, with more chloroplasts. The dwarfing characteristics of the 30 Gy group were also seen in the leaves, in which the cells were smaller, with stronger cell walls and tissue architecture.
In the cross-sectional view of the leaves of the control group (Figure 2a), the epidermis appears intact, with a single row of closely spaced cells. This is also the case for the 5 Gy dose group (Figure 2b), in which the cells of the epidermis are also closely aligned, forming a homogeneous unit. The change is observed in the 10 Gy dose group, in which the epidermal cells are more loosely structured. In the 30 Gy dose group, the epidermis cells are small, not homogeneous, and tightly fitting, so that a similarity is observed between the small, compact cells and the morphological, dwarf growth of the plant.
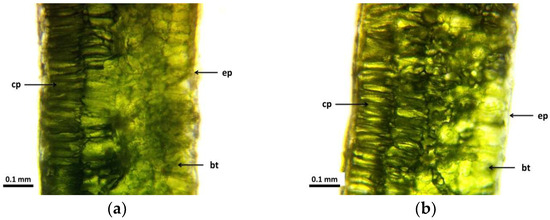
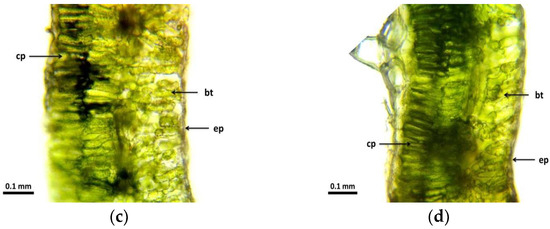
Figure 2.
Cross-sectional images of leaves of R. hirta “Őszifény” exposed to different gamma ray doses: (a) control, (b) 5 Gy, (c) 10 Gy, (d) 30 Gy. The abbreviations shown in the pictures mean the following: ep—epidermis; cp—columnar parenchyma; and bt—basal tissue.
Similar results can be observed when examining the parenchyma and the basal tissue. Here, it can be seen (Figure 2a) that both columnar and spongy parenchyma are homogeneous, without visible vacuoles. The cells are intact and closely spaced. In the group receiving the 5 Gy dose (Figure 2b), the thickness of the cell wall within the columnar parenchyma is significantly increased compared to the control group. The inner cell line also shows increased regularity and density. Parts of the spongy parenchyma show thickened cell walls while retaining strong confinement without the presence of vacuoles. The destruction becomes apparent in plants exposed to the 10 Gy dose (Figure 2c), in which the distortion becomes evident in the inner columnar parenchyma cells, which lack uniformity. Thickened walls are present but not homogeneously distributed among these distorted cells. The spongy parenchyma shows a uniform distortion throughout, with discontinuous or unsealed cell walls and cavities between the cells. In addition, larger vacuoles are also formed at this dose level. At a dose of 30 Gy, tissue uniformity is maintained as follows: both columnar/spongy parenchyma cells retain their integrity while showing thicker cell walls, which contribute to the overall structural strength of the leaf (Figure 2d).
The branch cross-sectional images (Figure 3) show similar results to the leaf cross-sectional images. In the control group (Figure 3a), it can be seen that the epidermis is uniform, with a continuous epidermis with homogeneous cells. The radiation effect can also be detected in the 5 Gy dose intensity samples (Figure 3b). The epidermis is much thicker, and the cells have strong walls and are organized and closed. The cross-sectional image of the branch in the 10 Gy group (Figure 3c) shows that the epidermis was homogeneous, whereas that of the 30 Gy dose group (Figure 3d) was also similar to the cross-sectional view of the leaf, with the epidermis cells tightly closed. The change was also observed in the transport tissue system. In the control group (Figure 3a), secondary thickening was not yet visible. The cambium ring appears dark with some vacuoles. The xylem and phloem are uniform. The effect of gamma radiation was also detectable in samples at a dose intensity of 5 Gy (Figure 3b). The cells of the phloem are well-delimited and have strong cell walls. Secondary thickening was markedly developed. The intestinal tissue of the branch was also markedly homogeneous with a strong cellular structure. No vacuoles are formed. The cross-sectional image of the stem of the 10 Gy group (Figure 3c) shows that vacuoles are beginning to form in the phloem. The cambium is continuous and intact, although there are tissue lesions in the xylem. Secondary thickening has begun to develop, but not to the extent seen at the 5 and 30 Gy dose levels. Vacuoles have also formed in the basal tissue. The cross-sectional view of the branch (Figure 3d) of the 30 Gy dose specimens was also similar to that seen in the leaf cross-section. The branches are well-developed and the cambium is thickened, although lesions are also observed. The phloem is strong, its cells are uniform, and secondary thickening has started to develop, but not yet at the same level of development as in the 5 Gy group. The cells forming the vascular ring are not uniform.
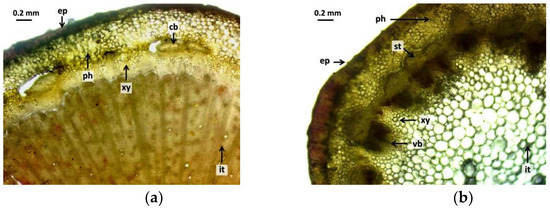
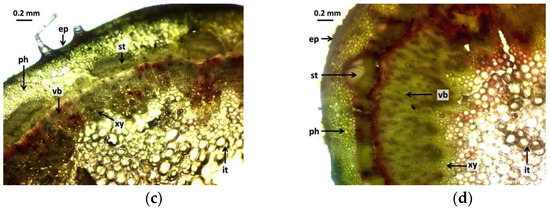
Figure 3.
Cross-sectional images of branches of R. hirta “Őszifény” exposed to different gamma ray doses: (a) control, (b) 5 Gy, (c) 10 Gy, (d) 30 Gy. The abbreviations shown in the pictures mean the following: ep—epidermis; xy—xylem; cb—cambium; ph—phloem, st—secondary thickening; vb—vascular bundles; and it—internal tissue.
3.3. Genetic Analyses
Assessment of molecular changes using SCoT and CDDP markers and genetic differences among the mutants generated by various irradiation doses were examined using two marker systems, SCoT and CDDP. Only markers that gave clear and reproducible fragments were evaluated (Figure 4). Across the marker analyses, both the appearance of new amplicons and the loss of normal fragments were also observed.
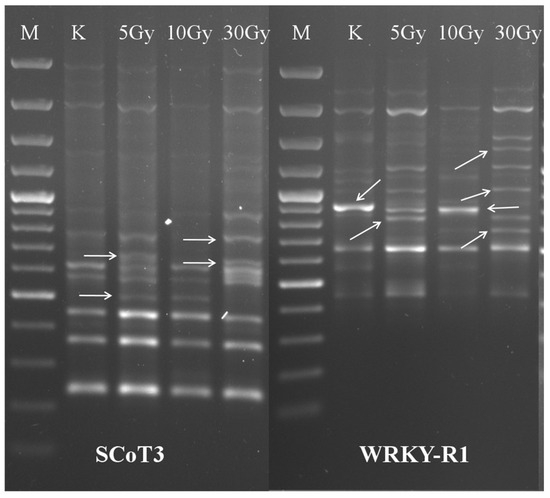
Figure 4.
Examples of genetic polymorphism detection with SCoT (SCoT3) and CDDP (WRKY-R1) markers in response to different irradiation. Polymorphic fragments are marked with arrows. M: DNA molecular weight marker (ThermoScientific GeneRuler 100 bp Lader Plus/3000 bp, 2000 bp, 1500 bp, 1200 bp, 1031 bp, 900 bp, 800 bp, 700 bp, 600 bp, 500 bp, 400 bp, 300 bp, 200 bp,100 bp); K: untreated control; 5 Gy, 10 Gy, 30 Gy irradiated plants.
Of the 14 SCoT markers tested, 9 gave evaluable patterns. SCoT primers amplified a total of 85 reproducible amplicons. The number of amplified fragments ranged from 6 for SCoT1 to 15 for SCoT12, with a mean of 9.4 bands per primer (Table 2). The average polymorphism for all primers was 42.7%.
Table 2.
List of SCoT primers that give an assessable pattern, number of amplified PCR products, polymorphic bands, and polymorphism percentage.
Of the investigated 21 CDDP, 10 gave an assessable pattern. The total number of amplified PCR fragments was 98. The number of amplicons ranged from 5 for ABP1-2 to 16 for WRKY-R1, with a mean of 9.8 bands per primer (Table 3). The average polymorphism was 45.1%, which was similar to the SCoT markers.
Table 3.
List of CDDP primers that give an assessable pattern, number of amplified PCR products, polymorphic bands, and polymorphism percentage.
By analyzing the combined marker data, we constructed a distance matrix (Table 4) and conducted a cluster analysis (Figure 5).
Table 4.
Genetic similarity matrix based on Jaccard coefficients.
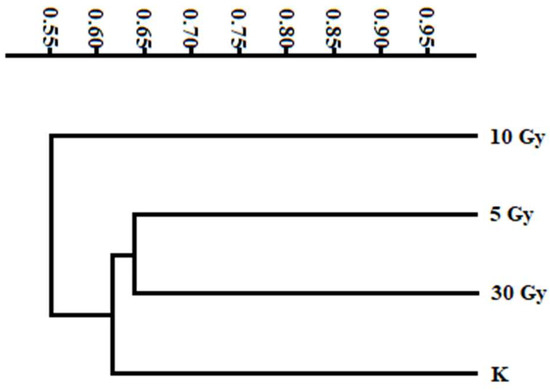
Figure 5.
Dendrogram derived from combined marker data of SCoT, CDDP, and RAPD techniques.
The highest dissimilarity value was observed in the 10 Gy mutant, while the lowest was found in the 30 Gy mutant. In the dendrogram created based on genetic similarity coefficients, mutants were grouped into two main clusters as follows: control (K) and 5 Gy and 30 Gy clustered together, whereas the 10 Gy mutant formed a separate group. This suggests that most changes emerged as a result of exposure to a radiation dose of 10 Gy.
4. Discussion
The morphological results show that all doses applied have an effect on the species studied, as described by Ibrahim et al. [6]. They suggest that mutation breeding generally has well-detectable phenotypic effects and that among the irradiation doses, the use of gamma irradiation is appropriate. This caused a high degree of genetic, and consequently phenotypic, variation compared to the results of Kumar et al. [17]. Plants radiated with 5 Gy showed significantly increased branching, number of flowers, and fresh and dry weights of the shoot and root, compared to the control. This may also be indicative of the hormesis phenomenon, whereby low-dose exposure can produce individuals of much larger size and biomass. Nevertheless, this type may also be applicable in breeding because although the breeding objectives described by Daughtrey and Benson [20] are aimed at compact plants, these large plants may be well suited for use in the urban backgrounds of associations or as structural plants, as described by Palmer et al. [19] in their work. However, because of its well-developed root system, this variety can also be used for soil erosion control, as recommended by Mircea et al. [24]. This is also supported by the results obtained using histological parameters, as in the groups receiving a 5 Gy radiation dose; strong tissue architecture, uniform, strong cells and transport tissues, and a transport tissue system in a more advanced state of development compared to the control group, which may also indicate a stronger resistance, were observed. For both the 5 Gy and 30 Gy doses, the results of Datta et al. [13] demonstrated that gamma radiation can induce uniform, homogeneous mutations in tissues.
The number of flowers also contributes to this result, which is significant compared to the control group—R. hirta is also ornamental in habit and flowers. In parallel with the large size, the dwarf variety may also be important for the trade. Individuals subjected to 30 Gy exhibited significantly smaller plants, yet produced the highest number of flowers, which is consistent with the breeding objectives described by Daughtrey and Benson [20]. Although Shukla et al. [23] claim in their work that radiation intensity affected flower color, this was not the case in the results of this experiment. In the case of the groups, the flower color did not change, although Daughtrey and Benson [20] describe the appearance of a new flower color as a desirable trait associated with mutation. The microscopic results are similar to the morphological results, as the small dwarf plants showed a small but dense tissue arrangement, as explained by Oates et al. [21], who suggest that the mutation is capable of reducing height and biomass in the genus Rudbeckia. Plants exposed to 30 Gy of radiation showed increased stiffness not only within the leaves but also throughout the plant. The variety could be further bred to become an occasional ornamental pot plant, as recommended by Beckwith [25]. In fact, the group receiving a 10 Gy dose was heterogeneous in its characteristics. This is clearly demonstrated by the genetic results obtained, which show that the 10 Gy dose is not closely related to the genetic traits of either the 5 Gy or the 30 Gy dose group, The large difference from the control group is evident in this case, and it can be stated that this mutant is not suitable for further research. Large variations in herd size and flower number were also detected, and this is reflected in the morphological results. However, the low number of branches and the small number of flowers are not suitable for use as an ornamental plant. This result is also closely correlated with the histological results, in which the basal tissue cells are heterogeneous; some cells have a stronger cell wall, and others seem to have weaker ones. However, negative changes were observed in the plants exposed to the 10 Gy dose compared to the leaves of the control group; these included leaf distortion and blind spots within their structure.
It was obvious that all dose levels affected root mass, in contrast to the lowest development observed in the control group, and mutation breeding can be used to monitor species variability, as shown by Ibrahim et al. [6]. Mutation techniques may also be suitable for breeding, according to Suprasanna et al. [9]. Doses of 10 Gy and 30 Gy showed less pronounced increases in root mass but still produced statistically significant effects. The most marked difference compared to the control was observed at the 5 Gy irradiation—a result consistent with the trends observed for branching and flower number.
Interestingly, the 10 Gray irradiation doses caused the greatest changes at the genetic level. Greater genetic variability does not necessarily manifest at the phenotypic level, too. Mutations can occur in both coding and non-coding regions. Changes in coding regions do not always result in amino acid substitutions and are, therefore, not phenotypically expressed. Unfortunately, changes in genome constitution induced by physical radiation are uncontrollable. This may be one of the reasons why the 10 Gy irradiations caused the largest genetic changes, which were not accompanied by similar phenotypic changes. Whole genome sequencing may be necessary to detect this contradiction.
Also, several studies have demonstrated that higher doses of irradiation do not always lead to greater genetic changes in plant genomes [36,37,38]. Our results affirm that utilizing combined data from SCoT and CDDP techniques is effective for detecting and assessing genetic diversity among mutants. Molecular markers associated with desirable agronomic traits can be employed for genetic mapping purposes, as well as marker-assisted selection, which is an approach discussed by Du et al. [39].
5. Conclusions
Due to increasing urbanization and the effects of climate change, many ornamental plant species that were often used in the past are becoming heritage species and may need to be bred for new traits, and gamma mutation offers a way to achieve this. This study reveals that gamma radiation significantly affects R. hirta “Őszifény”, as evidenced by morphological, histological, and genetic analyses. From the morphological and histological results obtained, it was obvious that the 10 Gy radiation dose resulted in the highest mutation rate. However, this dose had detrimental effects on ornamental plant breeding and production, manifesting as reduced branching and flower numbers, and yielded fresh and dried mass data statistically similar to the control group. Additionally, leaf tissue deformation further suggests that a radiation dose of 10 Gy is unsuitable for this cultivar. On the other hand, a dose rate of 5 Gy led to vigorous plants with increased branching, flower numbers, and mass data. The application of a 5 Gy radiation dose also revealed a hormesis phenomenon, which is an intriguing finding that warrants further investigation. If proven true in future studies, it could serve as an excellent tool for breeding R. hirta. Equally promising were the results observed at a radiation dose of 30 Gy, which produced significantly higher morphologic results compared to the control group—indicating its suitability for potential future breeding efforts in this species.
The high root mass observed along with increased flower numbers and branching traits are valuable characteristics for urban plants due to their contribution to proper drainage during sudden rainfall events while also enhancing drought tolerance, which is a crucial factor in arid regions. Moreover, the abundant flowers add aesthetic value to these mutant cultivars.
Genetic testing has provided novel insights into mutational breeding in R. hirta. This study represents the first instance in which molecular genetic markers have supported such efforts. Our molecular assays demonstrated that start codon targeted (SCoT) and conserved DNA-derived polymorphism (CDDP) techniques are suitable for detecting genetic changes caused by induced mutations. Although the markers employed do not cover the entire genome and not all changes are phenotypically expressed, they prove valuable in determining appropriate radiation doses. Furthermore, if these polymorphic fragments can be linked to new traits resulting from mutations, they hold the potential for future marker-based selection processes. The gamma mutation technique could be a fundamental tool for increasing genetic variability in R. hirta “Őszifény”, thus creating new phenotypic individuals and further breeding. The use of low doses is likely to result in hormesis, producing large individuals, while the use of 30 Gy may produce individuals suitable for pot culture. Genetically, these are the closest to the parent strain. The genetic distance of 10 Gy from them also shows unfavorable phenotypic attributes and is not recommended for further breeding.
In summary, this study concluded that 5 Gy and 30 Gy may be suitable for increasing genetic variability in R. hirta. In contrast, the heterogeneous population produced by 10 Gy is not suitable for this purpose. Since R. hirta is an important ornamental plant for urban use and breeding new varieties is important to maintain this status, 5 Gy may be suitable for breeding large-scale varieties that are suitable as background or structural plants, while 30 Gy may be suitable for breeding varieties that are suitable for pot production.
Author Contributions
Conceptualization, S.K., K.H., D.H.-F., Z.K., A.S. and L.O.; methodology, S.K. and K.H.; investigation, S.K., K.H., D.H.-F., Z.K., A.S. and L.O.; data curation, S.K., K.H. and D.H.-F.; writing—original draft preparation, S.K., K.H., D.H.-F., Z.K. and A.S.; writing—review and editing, S.K., K.H., D.H.-F. and M.A.W.; visualization, S.K., K.H., Z.K. and A.S.; supervision, D.H.-F., M.A.W. and L.O. All authors have read and agreed to the published version of the manuscript.
Funding
The research was supported by the project “The feasibility of the circular economy during national defense activities” of the 2021 Thematic Excellence Programme of the National Research, Development and Innovation Office under grant no.: TKP2021-NVA-22, led by the Centre for Circular Economy Analysis.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors on request.
Acknowledgments
We are grateful to our colleagues Zsanett Istvánfi, Zsolt Lénárt, Attila Janik, and Györgyné Gondos for giving us the opportunity to carry out this work.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Kowarik, I. Urban ornamentals escaped from cultivation. In Crop Ferality and Volunteerism; Gressel, J., Ed.; CRC Press: Boca Raton, FL, USA, 2005; pp. 97–121. [Google Scholar] [CrossRef]
- Wani, M.A.; Nazki, I.T.; Din, A.; Iqbal, S.; Wani, S.A.; Khan, F.U.; Neelofar, B. Floriculture sustainability initiative: The dawn of new era. Sustain. Agric Res. 2018, 27, 91–127. [Google Scholar]
- Farinati, S.; Betto, A.; Palumbo, F.; Scariolo, F.; Vannozzi, A.; Barcaccia, G. The new green challenge in urban planning: The right genetics in the right place. Horticulturae 2022, 8, 761. [Google Scholar] [CrossRef]
- Hoyle, H.; Jorgensen, A.; Warren, P.; Dunnett, N.; Evans, K. “Not in their front yard” The opportunities and challenges of introducing perennial urban meadows: A local authority stakeholder perspective. Urban For. Urban Green. 2017, 25, 139–149. [Google Scholar] [CrossRef]
- Wani, M.A.; Din, A.; Nazki, I.T.; Rehman, T.U.; Al-Khayri, J.M.; Jain, S.M.; Lone, R.A.; Bhat, Z.A.; Mushtaq, M. Navigating the future: Exploring technological advancements and emerging trends in the sustainable ornamental industry. Front. Environ. Sci. 2023, 11, 1188643. [Google Scholar] [CrossRef]
- Ibrahim, R.; Ahmad, Z.; Salleh, S.; Hassan, A.A.; Ariffin, S. Mutation Breeding in Ornamentals. In Ornamental Crops; Handbook of Plant Breeding, 11; Van Huylenbroeck, J., Ed.; Springer: Cham, Switzerland, 2018. [Google Scholar] [CrossRef]
- Schum, A. Mutation breeding in ornamentals: An efficient breedingmethod? Acta Hort. 2003, 612, 47–60. [Google Scholar] [CrossRef]
- Subburaj, S.; Tu, L.; Jin, Y.T.; Bae, S.; Seo, P.J.; Jung, Y.J.; Lee, G.J. Targeted genome editing, an alternative tool for trait improvement in horticultural crops. Hortic. Environ. Biotechnol. 2016, 57, 531–543. [Google Scholar] [CrossRef]
- Suprasanna, P.; Ganapathi, T.R.; Ghag, S.B.; Jain, S.M. Genetic modifications of horticultural plants by induced mutations and transgenic approach. IX Int. Symp. Vitr. Cult. Hortic. Breed. 2016, 1187, 219–232. [Google Scholar] [CrossRef]
- Jain, S.M. Mutation-assisted breeding for improving ornamental plants. Acta Hort. 2016, 714, 85–98. [Google Scholar]
- Jain, S.M. Mutagenesis in crop improvement under the climate change. Rom. Biotechnol. Lett. 2010, 15, 88–106. [Google Scholar]
- Schum, A.; Preil, W. Induced mutations in ornamental plants. In Somaclonal Variation and Induced Mutations in Crop Improvement; Springer: Dordrecht, The Netherlands, 1998; pp. 333–366. [Google Scholar]
- Datta, S.K.; Chakrabarty, D.; Mandal, A.K. Gamma irradiation: A potential tool for quality improvement and management of ornamental plants. Plant Stress 2005, 1, 32–35. [Google Scholar]
- Kiong, A.L.P.; Ling Pick, A.K.; Hussein, S. Physiological responses of Orthosiphon stamineus plantlets to gamma irradiation. Am. Eurasian J. Sustain. Agric. 2008, 2, 135–139. [Google Scholar]
- Jo, Y.D.; Lee, H.S.; Kim, J.B. Effect of X-ray irradiation on mutation induction and genetic diversity in rice. Breed. Sci. 2018, 68, 454–460. [Google Scholar]
- Kim, J.H.; Park, S.H.; Kim, J.S. Mutation breeding in ornamental plants using gamma irradiation. Hortic. Sci. Technol. 2017, 35, 1–10. [Google Scholar]
- Kumar, G.; Singh, H.; Sirohi, U. Comparative study of gamma, X-ray, and electron beam irradiation on mutation induction in marigold (Tagetes spp.). J. Hort. Res. 2017, 25, 212–218. [Google Scholar]
- Maluszynski, M.; Kasha, K.J.; Szarejko, I.; Maluszynska, J. Mutation techniques. In Advances in Plant Breeding Strategies; Springer: Berlin, Germany, 2009; pp. 159–194. [Google Scholar]
- Palmer, I.E.; Bir, R.E.; Lynch, N.P.; Ranney, T.G. Crossability, cytogenetics, and reproductive pathways in Rudbeckia subgenus Rudbeckia. HortScience 2009, 44, 44–48. [Google Scholar] [CrossRef]
- Daughtrey, M.L.; Benson, D.M. Principles of plant health management for ornamental plants. Annu. Rev. Phytopathol. 2005, 43, 141–169. [Google Scholar] [CrossRef]
- Oates, K.M.; Touchell, D.H.; Ranney, T.G. Induced variation in tetraploid Rudbeckia subtomentosa ‘Henry Eilers’ regenerated from gamma-irradiated callus. HortScience 2013, 48, 831–834. [Google Scholar] [CrossRef]
- Oates, K. Genetic Improvement of Rudbeckia spp. and Evaluation of a New Triploid Campsis Cultivar. Master’s Thesis, NC State University, Raleigh, NC, USA, 2011. [Google Scholar]
- Shukla, R.; Nath, P.; Jugran, H.M.; Gupta, M.N. Studies on gamma irradiation of Rudbeckia laciniata cultivar ‘Golden Glow’. J. Nucl. Agric. Biol. 1986, 15, 207–210. [Google Scholar]
- Mircea, C.; Cioancă, O.; Draghia, L.; Hăncianu, M. Morphological characteristics and polyphenol variations in Rudbeckia hirta L. Rom. Biotechnol. Lett. 2015, 20, 10688. [Google Scholar]
- Beckwith, D.D. Characterization of Juvenility and Photoperiodic Responses of Rudbeckia hirta Originating from Different Latitudes. Doctoral Dissertation, Virginia Tech, Blacksburg, Virginia, 1991. [Google Scholar]
- Harkess, R.L.; Lyons, R.E. Rudbeckia hirta L.: A versatile North American wildflower. HortScience 1994, 29, 134–227. [Google Scholar] [CrossRef]
- USDA Allain, L. Black-Eyed Susan. Available online: https://plants.usda.gov/DocumentLibrary/plantguide/pdf/pg_ruhi2.pdf (accessed on 1 July 2019).
- Palmer, I.E.; Touchell, D.H.; Ranney, T.G. In-vitro polyploid induction of Rudbeckia spp. South. Nurserymen Assn. Res. Conf. 2008, 53, 186–189. [Google Scholar]
- Fulcher, A.; Dunwell, W.C.; Wolfe, D. Rudbeckia taxa evaluation. South. Nurserymen Assoc. Res. Conf. 2003, 48, 510–512. [Google Scholar]
- Jourdan, P. Genetic Resources of Herbaceous Ornamentals in North America. In North American Crop Wild Relatives; Springer: Cham, Switzerland, 2019; Volume 2, pp. 607–643. [Google Scholar]
- Scott, L.; Molano-Flores, B.; Koontz, J.A. Comparisons of genetic variation and outcrossing potential between the sensitive species Rudbeckia fulgida var. sullivantii (Asteraceae) and its cultivar. Trans. Ill State Acad. Sci. 2007, 100, 129–144. [Google Scholar]
- Collard, B.C.Y.; Mackill, D.J. Start Codon Targeted (SCoT) polymorphism: A simple, novel DNA marker technique for generating gene-targeted markers in plants. Plant Mol. Biol. Rep. 2009, 27, 86–93. [Google Scholar] [CrossRef]
- Collard, B.C.Y.; Mackill, D.J. Conserved DNA-derived polymorphism (CDDP): A simple and novel method for generating DNA markers in plants. Plant Mol. Biol. Rep. 2009, 27, 558–562. [Google Scholar] [CrossRef]
- Boronkay, G. Evaluation of the items of rose garden Budatétény for hip decorativeness using mathematical models. Kertgazdaság Hortic. 2018, 50, 37–46. [Google Scholar]
- Hammer, O.; Harper, D.A.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Due, M.S.; Susilowati, A.; Yunus, A. The effect of gamma rays irradiation on diversity of Musa paradisiaca var. sapientum as revealed by ISSR molecular marker. Biodivers 2019, 20, 1416–1422. [Google Scholar]
- Wang, P.; Zhang, Y.; Zhao, L.; Mo, B.; Luo, T. Effect of gamma rays on Sophora davidii and detection of DNA polymorphism through ISSR marker. BioMed. Res. Inter. 2017, 2017, 8576404. [Google Scholar]
- Yarar, G.; Kocak, M.; Denli, N.; Cavagnaro, P.F.; Yildiz, M. Determination of the effective radiation dose for mutation breeding in purple carrot (Daucus carota L.) and possible variations formed. Mol. Biol. Rep. 2022, 49, 5219–5228. [Google Scholar] [CrossRef]
- Du, Y.; Luo, S.; Zhao, J.; Feng, Z.; Chen, X.; Ren, W.; Liu, X.; Wang, Z.; Yu, L.; Li, W.; et al. Genome and transcriptome-based characterization of high energy carbon-ion beam irradiation induced delayed flower senescence mutant in Lotus japonicus. BMC Plant Biol. 2021, 21, 510. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).