Abstract
Irrigation scheduling, coupled with the use of biofertilizers, constitutes an effective management strategy to overcome soil drought. This study aimed to assess the individual and combined effects of three selected biofertilizers—(R) Bacillus sp. and Bacillus subtilis, (M) native arbuscular mycorrhizal fungi, and (C) compost—on Triticum durum L. The agro-physiological and biochemical traits, along with the soil properties, were evaluated under two different water regimes, 100% crop evapotranspiration (ETc) for the well-watered regime and 30% ETc for the drought-stressed regime under field conditions, using a drip irrigation system. Drought stress (DS) led to significant reductions in the biomass, physiological parameters, and biochemical markers. Furthermore, the application of CM and MR significantly boosted the shoot and root dry weight by 137% and 72%, respectively, under DS compared to the control. Moreover, CM and R resulted in a notable increase of 167% and 130% in the grain and straw yield, respectively, under the same conditions. The normalized difference vegetation index showed improved values in the plants inoculated with MR and CMR, respectively, under DS conditions. Additionally, the soil quality was positively influenced by the application of M and/or R and/or C biofertilizers. These findings provide valuable insights into the intricate soil–plant interactions and the beneficial effects of combined biofertilizers in enhancing wheat’s resilience to drought stress.
1. Introduction
Environmental stress factors disrupt plant growth, development, and productivity, triggering various changes within a given crop [1]. Drought, a detrimental abiotic stress, represents a significant limitation to agricultural production worldwide, requiring efficient management of water resources spanning several agricultural and human activities [2]. Water deficit can hamper plant growth at the morphological, physiological, and biochemical scales [3], impeding shoot growth, stomatal closure, and impairing the photosynthetic system [4]. Furthermore, it causes the overproduction of reactive oxygen species, leading to membrane damage, a decreased photosynthetic rate, compromised membrane stability, and negatively altered water relations [5,6].
Durum wheat (Triticum durum L.) is a substantial crop species that plays a crucial role as both food and feed [7,8,9]. Thus, improving durum wheat production is globally recognized as a critical component of promoting food security. Wheat exhibits adaptability to different environmental conditions, facilitating widespread cultivation and long-term storage [10]. In addition, it holds economic significance as a valuable cereal crop in Morocco, a country with a net import reliance on wheat [11]. Hence, Moroccan durum wheat imports are expected to reach 5.40 million tons (Mt) according to the FAO this marketing year, compared with 4.80 Mt in 2022/23 and an average of around 4 Mt over the three previous marketing years. Purchases of durum wheat are also expected to rise compared with 2022/23, from 1.02 Mt to 1.12 Mt [11].
Drip irrigation (DI) stands as a prevalent method to irrigate annual row crops like durum wheat [12]. This sustainable modern technique delivers water gradually, targeting specific portions of the plant’s root zone [13]. Beyond its basic function, DI yields a range of benefits: optimized water management, improved crop establishment, enhanced weed control, increased yields, and improved crop quality [14]. Characterized by a minimal flow rate, DI allows meticulous regulation of the water distribution, ensuring precise irrigation directly at the root level, thus reducing losses through infiltration or evaporation [15,16]. Farmers increasingly need to learn how to leverage sustainable technologies to enhance their businesses. To address these challenges, a variety of technologies and agricultural products have emerged. Efficient techniques for managing soil, such as the use of arbuscular mycorrhizal fungi (AMF), organic amendments, and plant growth-promoting rhizobacteria (PGPR), can significantly improve the soil fertility, crop tolerance, water availability, and root surface area, offering powerful tools for drought mitigation in both irrigated and non-irrigated regions [17]. AMF, in particular, have been recognized for their ability to enhance plant tolerance to abiotic stress [18]. They facilitate increased water and nutrient uptake by the host plant in exchange for essential fatty acids and carbohydrates [19,20]. Moreover, AMF application can stimulate antioxidant enzyme activity and osmolyte accumulation under various abiotic stress conditions, including water deficits. Current research suggests that combining AMF with organic amendments can further boost soil fertility [21]. Organic compost serves as a beneficial alternative to excessive chemical fertilizer use in field crops [22]. It enhances the soil water-holding capacity, strengthens the soil structure, and improves nutrient retention [23]. Additionally, compost application enhances plant and soil health, reduces nutrient losses via leaching or volatilization, prevents soil erosion, increases soil water storage and the carbon content, and promotes long-term soil fertility [24]. Green composts are rich in nutrients, especially nitrogen (N), phosphorus (P), potassium (K), and a range of micronutrients, including magnesium, copper, and iron [25]. Furthermore, the action of advantageous soil microbes in enhancing the water status through interactions with plant roots and soil is a promising approach for sustainable agriculture [26]. Many PGPR enhance plant drought tolerance through mechanisms such as higher photosynthetic efficiency [27], increased production of antioxidant enzymes, and non-enzymatic antioxidants [28]. One critical trait of these bacteria is the enzyme 1-aminocyclopropane-1-carboxylate (ACC) deaminase [29], which supports plant growth under stress by preventing ethylene accumulation, which otherwise inhibits growth [30,31].
In the context of climate change, there remains a knowledge gap when it comes to applying biofertilizers in line with advanced irrigation management and scheduling technologies, such as drip irrigation based on crop evapotranspiration in agricultural fields. Hence, this study was carried out to assess the efficacy of indigenous biofertilizers, either alone or combined, in increasing durum wheat plants’ tolerance to drought conditions. The conducted field experiments evaluated the agro-physiological and biochemical responses of durum wheat plants grown under limited water availability. We hypothesized that: (1) the single and/or combined application of biofertilizers can improve growth, physiology, yield, and tolerance to water stress in the field; (2) biofertilizers can provide an alternative to the excessive use of chemical fertilizers (NPK) under water-stress conditions; and (3) biofertilizers can positively improve soil physicochemical properties and glomalin content compared with NPK application under both water regimes.
2. Materials and Methods
2.1. Study Location
The field experiments were conducted near Marrakesh, Morocco (31°34′46.2″ N, 8°16′13.1″ W), in a semi-arid region with an average annual rainfall of 347 mm, mainly from February to June. Summers are dry and warm, with temperatures reaching up to 38 °C, while winters are humid and mild, with temperatures not dropping below 3.5 °C, according to data from a nearby meteorological station. The fields had no prior treatments with chemical or organic fertilizers. The physicochemical properties of the agricultural soil are detailed as follows: pH, 8.36; electrical conductivity (EC) (mS/cm), 0.38; organic matter (%), 1.78; available phosphorus (mg/kg), 98.2 and total nitrogen (%), 0.76; with a sandy clay loam texture, 44%, 32%, and 24%; cation exchange capacity, 4.48 meq/100 g; bulk density, 1.39 g/cm3, respectively, and the physico-chemical characteristics of the water are as follows: pH, 7.74; EC, 0.73 (mS/cm); TKN, 12.21 (mg/L); and COD, 36.91 (mg/L).
2.2. Experimental Design
The trial was performed during the spring wheat growing seasons, from February to May, in 2022–2023. Each plot was isolated by a 60 cm wide concrete wall to prevent water infiltration. The size of each plot was 1.2 m2 (1.5 m wide × 0.8 m long). A factorial design with two main factors, factor 1: drought conditions testing two water regimes (100% ETc for the well-watered (WW) and 30% ETc for the drought-stressed (DS) and factor 2: treatments with the different biofertilizers applied alone or in combination, with negative (without NPK) and positive (with NPK) controls, was set. Eighteen treatments were applied in a randomized complete block design (RCBD), including two water regimes for WW and DS. Each treatment was repeated three times. The study treatments included uninoculated negative control plants (Ct−), uninoculated positive control plants with chemical fertilizers N, P, and K (Ct+), plants treated with the AMF consortium (M), plants treated by bacterial strains (R), plants amended with compost (C), and their double and triple combinations (MR), (CR), (CM), and (CMR) (Figure 1). Three-week-old plants were subjected to drought stress (DS) treatments. The agro-physiological and biochemical characteristics were subsequently evaluated upon reaching maturity at 12 weeks of age. The approach for calculating the crop water requirements utilized in this study was developed by the Food and Agriculture Organization of the United Nations (FAO). The following equation was used to determine the crop evapotranspiration:
where ETc is the crop evapotranspiration; Kc is the crop coefficient, which integrates the crop characteristics and the average effects of soil evaporation and varies during the growing period, where the initial stage (Kc ini = 0.7), the mid-season stage (Kc mid = 1.15) and at the end of the late season stage (Kc end = 0.3) are determine according to Allen et al. [32]; and ETo is the reference evapotranspiration (mm) computed from meteorological data obtained from a local station using the Penman–Monteith method [32].
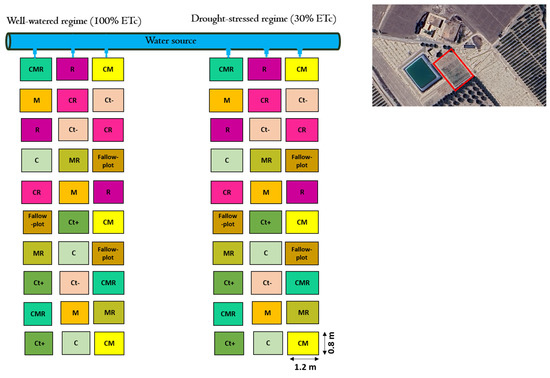
Figure 1.
Experimental design of the field trial with all the applied biofertilizers under two water regimes (100% ETc: well-watered and 30% ETc: drought stress) at the site location.
2.3. Drip Irrigation System
The field trial utilized a drip irrigation system (DIS) comprising a control unit with a pressure tank, plate filter, and control valves, alongside distribution lines. The main water line consisted of polyethylene pipes. The drip lines were connected to capillary tubes with individual opening valves per plot, spaced 20 cm apart, and had a flow rate of 8 L H−1 under the WW regime and 4 L H−1 under the DS regime. Adjacent drip lines were spaced 20 cm apart, and each experimental plot featured two laterals. The irrigation water was sourced from a depth of 200 m in the water table.
2.4. Isolation of Microorganisms and Plant Inoculation
Two bacterial strains, Bacillus subtilis and Bacillus sp., were sourced from the Tafilalet palm grove, located 500 km southeast of Marrakesh, Morocco, a semi-arid region. To isolate these strains, soil was mixed with sterile 0.9% NaCl solution and agitated for 30 min. Serial dilutions were prepared and administered in aliquots (105 and 106) on phosphate-enriched growth medium (NBRIY) without yeast extract from the National Institute for Botanical Research. The bacteria were incubated for 48 h at 28 °C. Colonies were subsequently purified by restreaking on Tryptone Soy Agar (TSA) to isolate individual colonies. Each strain was then cultured in liquid Tryptone Soy Broth (TSB) with agitation at 30 °C for 48 h until reaching an optical density of approximately 1 at 600 nm (equivalent to 1 × 109 CFU/mL). A bacterial solution containing equal quantities of both strains (2 mL) was applied to the soil. A second injection of the same volume was performed 15 days later to increase the bacterial population in the soil and ensure the infection of newly developed roots. The resilience of these PGPR strains to water deprivation was evaluated using polyethylene glycol [33].
This study also included a native AMF consortium called Aoufous, which originates from the same location (Tafilalet palm grove). It consists of 15 distinct species [21]. Zea mays L. acted as the host plant for the AMF consortium’s three-month cultivation in pots. The mycorrhizal inoculum, which included mycorrhizal spores, hyphae, and maize root pieces, was taken from the cultivated crop’s rhizosphere. For the wheat inoculation, 100 g of the inoculum was applied to each plot’s root system.
The compost used in this field study was made from green waste (quack grass), as reported by Meddich et al. [34]. The compost’s physicochemical characteristics were as follows: available phosphorus 700 ppm, nitrogen 1.5%, potassium 0.5%, and organic carbon 20%. The compost was applied at a rate of 480 g/plot (4 t/ha) to the relevant plot.
2.5. Plant Management
The durum wheat cultivar (Triticum durum L. var. carioca) was grown in this experiment. It is widely planted because of its good resistance to frost and lodging and is highly consumed in Morocco. A compound fertilizer (NPK) according to the recommendation of the Moroccan Ministry of Agriculture and Fisheries was applied at a rate of (140 kg N/ha as ammonium nitrate + 80 kg P2O5/ha as superphosphate + 50 kg K2O/ha as potassium sulfate) before sowing [35]. The durum wheat was sown at a seeding rate of 150 kg ha−1, with a density of 300 plants m−2, and covered with a 2 cm thick layer of soil.
2.6. Morphological Growth Traits and Mycorrhization Rate
Measurements of the shoot height, root length, spikelet height, seed count, and dry biomass of shoots and spikelets—obtained after the samples were dried at 80 °C for 48 h—were used to assess the growth of the wheat plants. The root samples were cleansed with 10% KOH at 90 °C for 30 min in order to measure the mycorrhization. After that, they were rinsed with distilled water. Then, using the procedure described by Phillips and Hayman [36], the roots were acidified for ten minutes using 2% hydrochloric acid and stained with Trypan blue for twenty minutes at 90 °C [37], and then the microscopic evaluation of mycorrhizal root colonization rates was carried out. Equations (1) and (2) were utilized to assess the mycorrhization frequency (MF) and intensity (MI), respectively.
With the following infection rates, and the number of fragments represented by the notations 5, 4, 3, 2, and 1, respectively, we obtain (n5, n4, n3, n2, and n1): Class 1: trace; Class 0: no mycorrhization; Class 5: greater than 91%; Class 4: between 51% and 90%; Class 3: between 11% and 50%; Class 2: less than 10%.
2.7. Photosynthetic Efficiency, Gas Exchanges, and Carotenoid Pigment Quantification
A fluorometer (OPTISCIENCE, OS30p, Hudson, NY, USA) was used to compute the PSII (Fv/Fm) of the chlorophyll fluorescence. After Harley et al. [38], the stomatal conductance (gs) was measured using a porometer (CI-340, Handheld Photosynthesis System, Washington, DC, USA). The concentrations of carotenoids and total chlorophyll were measured using Hartmut Lichtenthaler’s method [39]. To extract carotenoid, 100 mg of leaf tissue was homogenized in 80% acetone that had been cooled beforehand, and then centrifuged at 10,000× g for 10 min. After the supernatants were gathered, a UV/visible spectrophotometer was used to detect the absorbance at 480, 645, and 663 nm.
2.8. Total Soluble Sugars, Protein Profile, and Free Proline
The content of total soluble sugars (TSS) was measured using the procedure outlined by Dubois et al. (1956) [40]. For the TSS extraction, a 0.1 g aliquot of previously cooled leaf powder was mixed with 80% ethanol (v/v). Then, 0.2 mL of phenol, 1 mL of sulfuric acid concentration, and 0.2 mL of supernatant were combined following centrifugation. To calculate the TSS, the absorbance at 485 nm was measured. The frozen aerial piece (0.1 g) was combined with 4 mL of 1 M phosphate buffer (pH 7), 2.5% insoluble polyvinylpolypyrrolidone (PVPP), and 0.1 mM EDTA for the protein content assay. The supernatant was used to quantify the total soluble proteins [41] and antioxidant enzyme activity [42,43] following centrifugation at 4 °C. Fresh samples (0.1 g) were powdered in 4 mL of 40% ethanol and stored at 4 °C for the entire night in order to measure the proline concentration. One milliliter of a mixture comprising 20% ethanol, 1% ninhydrin, and 60% acetic acid was combined with the ethanol extract. After 20 min of heating the combination to 90 °C, the optical density (OD) measured at 520 nm [44].
2.9. Antioxidant Enzymes Determination
The peroxidase activity was measured using the procedure described by Polle et al. Here, 2 mL of phosphate buffer (0.1 M, pH 7), 1 mL of guaiacol (20 mM), 0.3% H2O2 (10 mM), and 0.1 mL of enzyme extract made up the reaction mixture. After adding 0.5 mL of 0.3% H2O2 (10 mM), the reaction started. At 470 nm POX activity was measured as described by Polle et al. [43] while, the activity of polyphenol oxidase (PPO) was determined using the method of Hori et al. [45]. For this, 2 mL of catechol (10 mM) and 0.1 mL of enzyme extract were combined in phosphate buffer (pH 7). Then, 1 mg of protein per enzyme unit, or PPO activity, was defined as the quantity of enzyme that causes a rise in absorbance at 420 nm of 0.001 min−1.
2.10. Stress Indicators (Malondialdehyde and Hydrogen Peroxide) Determination
We used Dhindsa and Matowe’s method [46] for measuring malondialdehyde (MDA). Here, 0.05 g of frozen leaf powder subsamples were combined with 1 mL of 90% acetone and 10% trichloroacetic acid (TCA) using a homogenizer. After centrifugation, 0.5 mL of 0.6% thiobarbituric acid (TBA) and 0.5 mL of 0.1% phosphoric acid were mixed with 0.25 mL of supernatant. The mixture was then incubated at 100 °C for 30 min in an ice bath to cease the process. After adding 0.75 mL of 1-butanol, the intensity of the absorption at 450, 532, and 600 nm was measured to estimate the MDA level.
Utilizing the technique described by Velikova et al. [47], the hydrogen peroxide (H2O2) concentration of the leaves was ascertained. After mixing 0.1 g of frozen leaf powder with 2 mL of 10% (w/v) TCA, the mixture was centrifuged at 15,000× g. One milliliter of potassium iodide (1 M) and five milliliters of potassium phosphate buffer (10 mM, pH 7) were added to the supernatant in order to quantify the H2O2. Following an incubation period of three minutes, the absorbance at 390 nm was measured and a standard curve for H2O2 was produced.
2.11. Soil Quality
Soil samples were collected from the root zone of wheat plants during the harvesting period and examined. A conductivity meter (HI-9033, Hanna Instruments, Padova, Italy) was used to measure the electrical conductivity (EC), and a pH meter (HI 9025, Woonsocket, RI, USA) was used to analyze the available phosphorus (AP). Each soil sample was sieved to a size of 2 mm, and the mixture was stirred for 30 min before the pH was measured. The methods outlined in [48] were used to evaluate the total organic matter (TOM) and total organic carbon (TOC), which requires the OM to be oxidized with dichromate of potassium when sulfuric acid is present. The NTK (total nitrogen content) was measured using the Kjeldahl technique [49].
Cornejo et al.’s method [50] was used to assess the total glomalin-related soil protein (T-GRSP). To extract the T-GRSP from 1 g of soil, we employed 4 mL of 50 mM sodium citrate buffer (pH 8.0). The extract underwent a 1 h autoclaving process at 121 °C and a 1 h centrifugation at 10,000× g. The Bradford assay was used to determine the T-GRSP content.
2.12. Data Collection
The soil moisture and temperature were measured at 20 cm intervals using a time domain reflectometry (TDR/The SDI-12 Sensor Reader) instrument (Marcon Ln., Ste. 175, Meridian, ID, USA). After sowing, measurements were taken every week.
2.13. Normalized Difference Vegetation Index (NDVI)
The GreenSeeker-NDVI was recorded every 15 days. The GS-NDVI of the entire plot was measured once using a hand-held sensor (HCS100, Trimble Inc., Sunnyvale, CA, USA) at 60 to 70 cm above the plant canopy between midday and 1 a.m. The recorded data were stored in the instrument’s internal memory and could be downloaded at a later date in *.csv format.
2.14. Statistical Data Analysis
The results illustrate the mean of three replications per treatment ± standard error. Statistical analysis was carried out using COSTAT software (version 6.3). The data underwent an analysis of variance, followed by Tukey’s honest significant difference (p ≤ 0.05) as a post hoc test. Using SPSS 23.0 software (multivariate analysis of variance (MANOVA), IBM, Armonk, NY, USA), the significant differences between the tested factors (AMF; M, Compost; C, PGPR; R, NPK; Ct+ and drought; DS) and their interactions, with a 95% confidence level, were determined using Tukey’s honest significant difference test (p ≤ 0.05). Principal component analysis was performed on all the growth, physiological, biochemical, and soil physicochemical features (PCA). The PCA and the heatmap analyses were carried out using R Studio (version 2023.9.1.494).
3. Results
3.1. Growth Measurement and Symbiotic Development
3.1.1. Mycorrhizal Colonization
The findings showed a significant reduction in both the frequency (MF) and the intensity (MI) of AMF infection in wheat plant roots under drought stress conditions (p < 0.05) (Figure 2). Plants that were neither inoculated nor amended (Ct−) exhibited a root MF of 40% under well-watered conditions (WW) and 36% under drought stress (DS). However, it should be noted that durum wheat plants treated with a conventional NPK chemical fertilizer (Ct+) showed a greater reduction in root colonization, regardless of the water regime applied. In addition, the roots of plants treated with NPK (Ct+) showed an MF of 47% and 29% under WW and DS, respectively. In contrast, plants treated with M, either alone or in combination (MR, CM, and CMR), revealed the highest MF, around 86%, 80%, 63%, and 66%, respectively under WW and 80, 73, 53, and 63%, respectively, under DS (Figure 2A). This suggests that the inoculation with AMF resulted in increased root colonization in durum wheat plants under both stressful and non-stressful conditions. Moreover, the MI was 12% under WW and 8% under DS for control plants (Ct−), while for plants treated with the M, it was 51% under WW and 21% compared to the Ct− and Ct+ under DS (Figure 2B). The interaction of M×C×R had a significant effect (p < 0.001) on the MI and MF (Table S1).
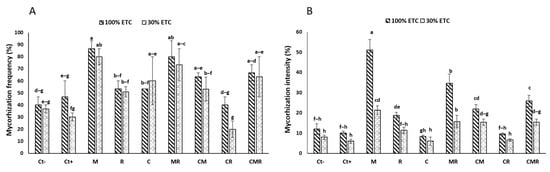
Figure 2.
The impact of two water regimes on the mycorrhization frequency (A) and mycorrhizal intensity (B) (100% ETc: well-watered and 30% ETc: drought stress). PGPR: consortium R; M: AMF consortium; Ct+: positive control plants; Ct−: negative control plants. The presented data are the means ± SD. Tukey’s HSD indicates that bars with identical lettering in every graphic do not differ substantially at p < 0.05.
3.1.2. Growth Assessment
Water stress negatively affected all the growth parameters, including the shoot height, root length, spikelet height, number of seeds, and dry biomass of spikelet (Table 1) (p < 0.05), shoot and root dry weight (SDW and RDW) (Figure 3A,B). Reducing irrigation to 30% ETc showed a very significant reduction in the control (Ct−) compared with CM and CMR-treated plants by 136% and 118%, respectively. Similarly, under 30% ETc, the RDW was significantly enhanced in plants inoculated by MR (71%), and CMR (57%) compared to the Ct−, but the highest value was found in plants treated with Ct+. The double application of AMF + PGPR (MR) and AMF + compost (CM) provided better results and mitigated the effects of DS on growth parameters. As for the grain and straw yield (Figure 3C,D), they showed a positive impact of the application of biofertilizers. The highest grain yield was observed in durum wheat plants treated with the double combination of M and CM compared with two controls under 30% ETc. The interaction of M×DS×C had a significant effect (p < 0.001) on the SDW, grain yield, and straw yield (Table S1).

Table 1.
Water regimes, 100% ETc and 30% ETc affected the growth characteristics of the negative and positive control (Ct− and Ct+) durum wheat plants, as well as those inoculated with native arbuscular mycorrhizal fungi (M), bacterial consortium (R), and/or compost (C). The data reported are means ± SE. Values with the same letter are not significantly different at p < 0.05 Tukey’s test.
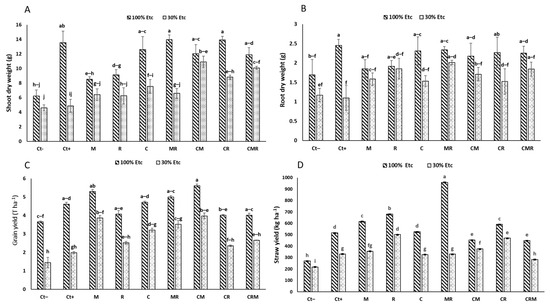
Figure 3.
The effect of the water regime applications on the growth markers in wheat plants that have been infected with natural arbuscular mycorrhizal fungus (M) and/or a bacterial consortium (R) and treated with compost (C). (A) Shoot dry weight, (B) root dry weight, (C) grain yield, and (D) straw yield are shown in the panels. The information shows the three biological replicates’ mean ± SE. Means with identical letters do not differ substantially at p < 0.05.
Concerning the growth parameters, durum wheat plants treated with biofertilizers applied separately showed no significant difference from those treated with NPK (Ct+), regardless of the water regime applied.
3.2. Effect of Field-Drought Stress and Microbial Biofertilizers on the Efficiency of the Photosynthetic Machinery
Plants under 30% ETc drought exposure showed substantial decreases in the physiological indices, including stomatal conductance (gs) and chlorophyll fluorescence (Fv/Fm), total chlorophyll (Chl T) and carotenoids (p < 0.05) (Figure 4). The use of M and/or R considerably enhanced the gs under 100% ETc, especially in durum wheat plants amended with C alone by 21% and Ct+ by 10% compared to the Ct− plants (Figure 4A). On the other hand, the foliar Fv/Fm ratio of plants treated with R was significantly improved (11%) under conditions of severe water limitation compared with Ct− (Figure 4B). Under stress conditions, durum wheat plants doubly inoculated with M and R (MR) and/or amended with compost (CMR) showed an improvement in the chlorophyll pigment content (T_Chl) compared to the controls (Figure 4C). This increase in the chlorophyll pigments remained insignificant in wheat plants treated with Ct+ (Figure 4C). However, the carotenoid levels showed a remarkable reduction in plants inoculated with R and M (MR) and amended with C (CMR) compared with controls (Ct− and Ct+) (Figure 4D). The interaction of M×R×DS×C had a significant effect (p < 0.001) on the Fv/Fm, Chl T, and carotenoids (Table S1).
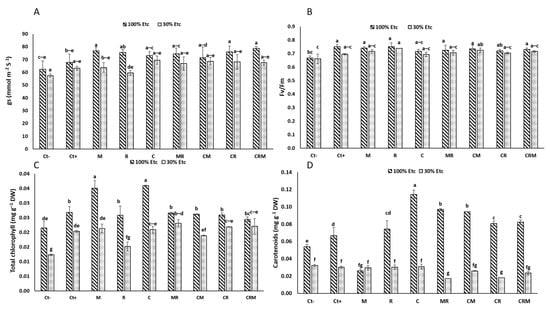
Figure 4.
The impact of the water regimes on the physiological characteristics of durum wheat plants that have been infected with native arbuscular mycorrhizal fungi (M) and/or a bacterial consortium (R) and treated with compost (C). The following are displayed in panels: (A) carotenoids; (B) total chlorophyll; (C) chlorophyll fluorescence; and (D) stomatal conductance. The information shows the three biological replicates’ mean ± SE. Means with identical letters do not differ substantially at p < 0.05.
3.3. Total Soluble Sugar, Protein Content, and Proline Quantification in Durum Wheat Plants
In comparison with normal irrigated plants, DS induced a remarkable accumulation of total soluble sugars (TSS), proteins, and proline in durum wheat leaves (Table 2). Under the same conditions, durum wheat plants treated with MR accumulated higher levels of protein by 19% compared to the control plants (Ct−). In contrast, NPK-treated plants showed a reduction in the protein contents by 11% compared to Ct−. Under DS, untreated wheat plants accumulated significant concentrations of TSS and proline content compared with other treatments, including Ct+. The interaction of DS had a significant effect (p < 0.001) on the TSS, proteins, and proline content (Table S1).
Table 2.
The impact of the water regimes, 100% ETc and 30% ETc on the osmolyte content and antioxidant enzyme activity (POX: peroxidase and PPO: polyphenol oxidase) of negative control (Ct−), positive control (Ct+), durum wheat plants inoculated with native arbuscular mycorrhizal fungi (M), bacterial consortium (R), and/or compost (C) was investigated. The presented data represent means ± standard error. Values with the same letter are not significantly different at p < 0.05 Tukey’s test.
3.4. Enzymatic Activities in Durum Wheat Plants
We investigated the changes in the activity of antioxidant enzymes, including polyphenol oxidase (PPO) and peroxidase (POX), in response to M and/or C and/or R in field-grown durum wheat plants. The activities of POX and PPO were notably elevated under field-drought conditions, as indicated in Table 2. However, the PPO activity was increased when durum wheat plants were treated with single (M), double (MR), triple (CMR) association, and Ct+ under 30% ETc by 23%, 22%, 17%, and 8%, respectively, compared to the Ct−. Moreover, the POX activity registered higher values in plants treated with the R, CR, and MR treatments compared to the controls, especially Ct−. Under DS, the interaction between M and C had a significant effect on the POX (p < 0.05) and PPO (p < 0.005) (Table S1).
3.5. Stress Markers in Durum Wheat Plants
The H2O2 and MDA concentrations in the durum wheat leaves were increased by exposure to drought stress (Figure 5A,B). Control plants subject to drought deficit had the highest values of H2O2 and MDA, 20% and 11%, respectively, in the leaves. In contrast, the lowest H2O2 and MDA values were recorded in plants treated with the double combination MR by roughly 50% and 52%, respectively, in the leaves compared with DS control plants, particularly Ct− (Figure 5A,B). Under DS, the interaction of R had a significant effect on the MDA (p < 0.05, Table S1).
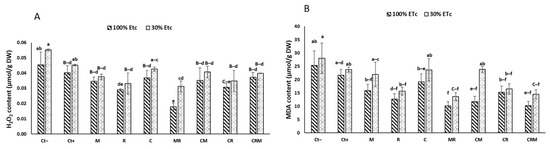
Figure 5.
The effects on the (A) H2O2 and (B) MDA activity of two distinct water regimes (100% ETc and 30% ETc) in wheat plants treated with compost (C), native arbuscular mycorrhizal fungus (M), and/or a bacterial consortium (R). The information shows the three biological replicates’ mean ± SE. At p < 0.05, there is no significant difference between means that have the same letters.
3.6. Soil Parameters
After harvest, the quality of the agricultural soil was evaluated, focusing on parameters like the pH, electrical conductivity (EC), total organic matter (TOM), total organic carbon (TOC) and AP, total nitrogen (N) contents, and total glomalin-related soil protein (T-GRSP) concentration. The data presented in Table 3 reveal that the presence of organic matter, the N, P, and glomalin production in the amended soil increased under well-watered (WW) conditions. Conversely, under drought stress (DS) conditions, the soil pH after harvest improved with R inoculation, and the EC was enhanced by approximately 15% in the C treatment compared to the negative control. Furthermore, the application of biofertilizers significantly elevated the TOM and TOC, phosphorus, and glomalin content compared to the control. Specifically, under DS conditions, the highest TOM and TOC values (100% and 80%) were observed in soils treated with R and MR, respectively. While the highest phosphorus values were recorded in soils treated with the CMR by 1374% and CM by 293% compared with Ct−, they were improved by 280% and 5.6%, respectively, compared with Ct+ under drought stress. Additionally, under DS conditions, the most substantial increases in the total GRSP (T-GRSP) were observed in soils treated with CR (118%) and M (112%) compared to the controls, including Ct−. However, a low T-GRSP content was observed in soil treated with Ct+. Under the same conditions, treatments with M and C improved the nitrogen (NTK) concentration by 0.19% and 0.16%, respectively, compared to the stressed Ct−. Under DS, the interaction between M and R had a significant effect on the T-GRSP (p < 0.001) and TOM (p < 0.005, Table S1).
Table 3.
Impact of the water regimes, 100% ETc and 30% ETc on the physicochemical parameters of soil after harvest of negative and positive control (Ct−) and (Ct+) durum wheat plants and plants inoculated with native arbuscular mycorrhizal fungi (M) and/or bacterial consortium (R) and/or compost (C). Data presented are means ± SE. Values with the same letter are not significantly different at p < 0.05 (Tukey’s test).
3.7. Collected Data from GreenSeeker and TDR Tools
Figure 6A,B show two main parameters. Figure 6A shows the normalized difference vegetation index (NDVI), where the variation in this parameter is shown as a function of the different treatments applied (Ct−, Ct+, M, C, R, MR, CR, CM, and CMR) and varies between 0 and 0.8. Under DS conditions, the NDVI decreased compared to wheat plants subject to normal irrigation. The most frequent improvement was recorded in plants treated with the double combination between MR and the triple combination CMR with a good distribution based on the median compared to the other treatments, including Ct− and Ct+. Figure 6B mentions the variation in the soil moisture and temperature as a function of the phenological stages, rainfall, and irrigation events in durum wheat growing. From seed germination to wheat plant maturity, we observed an increase in the soil moisture, which reached 32% v/v at the heading stage and is inversely proportional to temperature with a 25 °C at the maturity stage, as well as an increase in the soil moisture, especially at the development stage of durum wheat plants, with every applied irrigation.
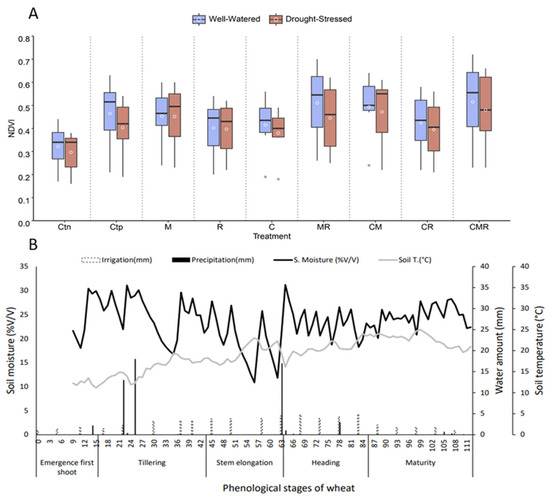
Figure 6.
(A) Variation of the normalized difference vegetation index depending on the different applied treatments; Ctn: negative control, Ctp: positive control, compost (C) and inoculated with native arbuscular mycorrhizal fungi (M), and/or a bacterial consortium (R) under the two water regimes applied (100% ETc: well-watered and 30% ETc: drought stress). (B) The relationship between the soil moisture, soil temperature, and irrigation regarding wheat plants’ phenology. Boxplots show the median (horizontal line), quartiles (boxes), and mean (white circle).
3.8. Principal Component Analysis
Principal component analysis (PCA) was used to understand the relationships between the different parameters tested as a function of the water regimes and biofertilizers. The findings indicated that the first (PC1) and second (PC2) components explained 58.1% of the variability, with PC1 accounting for 45.5% and PC2 for 12.6% (Figure 7). The PCA results showed that the majority of the examined parameters were either positively or negatively correlated. Furthermore, PC1 had the highest divergence, as opposed to PC2. The PCA of individuals showed that the Ms, CMs, MRs, Cs, CRs, Rs, and CMRs treatments were positively correlated along the PC1 axis under the drought-stressed regime and they were negatively correlated with the well-watered treatments except Ctn for PC1. This goes hand in hand with the strong contribution revealed in the PCA variables between the biochemical parameters (POX, PPO, and Prot), AP in the soil and their positive correlation with drought-stressed treatments, especially the dual and triple combinations (CM, MR, and CMR). Moreover, the PCA also showed a clear positive correlation under the well-watered regime between the growth parameters (RDW, SN, SDW, SpDW, and Gyield), physiology (Fv/Fm, gs), and soil characteristics (Glm). The stress markers (H2O2, and MDA), proline content and sugar content were positively correlated with the stressed controls (Ctps and Ctns) and well-watered negative control (Ctn) for PC1.
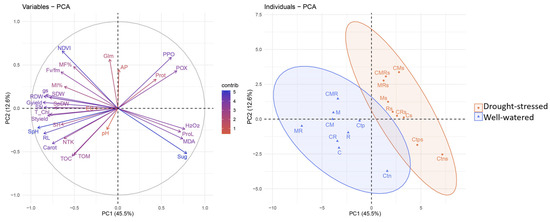
Figure 7.
Principal component analysis (PCA) between individuals and variables.
3.9. Cluster Analysis and Dendrograms in a Heat Map Matrix
This analysis makes it simpler to comprehend the data by presenting the parameters with higher (blue color) and lower (red color) values and arranging them. For example, in the half right of the heatmap, the growth parameters (SN, SH, RL, SH, SpH, SpDW, Styield, RDW, Gyield, and SDW), physiological parameters (gs, Fv/Fm, T-Chl, and Carot), and soil physicochemical properties (AP, TOM, TOC, Glm, pH, EC, and NTK) were significantly increased and positively correlated by the single or combined application of the biofertilizers, in particular C, M, and the combination of C with microorganisms (CR) under drought stress conditions (Figure 8). On the other hand, a stronger positive correlation was observed concerning the biochemical traits (MDA, H2O2, PPO, POX, Sug, Prot, and ProL) in durum wheat plants treated with C and/or M and/or R under well-watered conditions.
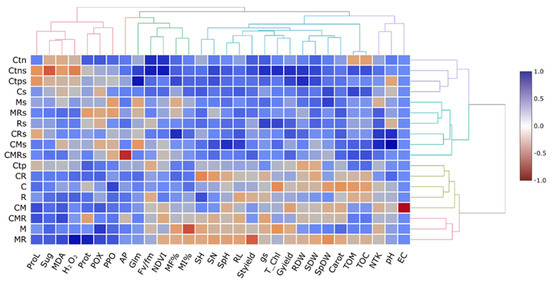
Figure 8.
Heatmap analyses of wheat grown under two water regimes (100 and 30% ETc) and submitted to different applied treatments; Ctn: negative control, Ctp: positive control, compost (C) and inoculated with native arbuscular mycorrhizal fungi (M), and/or a bacterial consortium (R). Treatments with (s) means under 30% ETc. proL: proline; Sug: sugar; MDA: malondialdehyde; H2O2: hydrogen peroxide; prot: protein; POX: peroxidase activity; PPO: polyphenol oxidase activity; AP: available phosphorus; Glm: glomalin; Fv/Fm: chlorophyll fluorescence; NDVI: normalized difference vegetation index; MF%: mycorrhizal frequency; MI%: mycorrhizal intensity; SH: shoot height; SN: spikelet number; SpH: spikelet height; RL: roots length; Styield: straw yield; gs: stomatal conductance; T_chl: total chlorophyll; Gyield: grain yield; RDW: root dry weight; SDW: shoot dry weight; SpDW: spikelet dry weight; Carot: carotenoids; TOM: organic matter; TOC: total organic carbon; NTK: nitrogen total Kjeldahl; pH: hydrogen potential, and EC: electrical conductivity.
4. Discussion
Water scarcity is a significant challenge to agriculture, impacting crop development and productivity owing to an imbalance between the quantity of reactive oxygen species and their antioxidants [51]. Plants must adapt to environmental pressures to survive. In arid areas, drip irrigation (DI) stands out as the most widely prevalent water-saving technology for increasing yields in agricultural systems [52]. A study by Yang et al. [53] on spring wheat in the field revealed that DI highlighted the role of soil moisture in the soil–plant–atmosphere continuum, enhanced crop activity by increasing the field temperature, especially the soil temperature (+1.10 °C), and improved the yield (+10.3%). Biofertilizers, such as AMF, PGPR, and compost, represent alternative strategies. Inoculating and/or amending wheat plants with these natural agents has proven to be beneficial, offering protection against oxidative stressors, especially drought. Further investigation is needed to ensure that the cost-effectiveness and widespread adoption of these technologies can surpass current approaches. Likewise, microbial biofertilizers, including AMF and PGPR, either individually or in combination, are gaining prominence in harsh environmental conditions. They significantly reduce the negative effects of drought stress on growth, photosynthetic apparatus, nutrient uptake, physiological traits, and oxidative stress. These biofertilizers also demonstrate high osmotic stress tolerance under water deficit conditions.
To ensure production under water stress conditions and to assist in retaining and preserving soil fertility, mycorrhiza may be used in sustainable agriculture. They possess the ability to modulate physiological and biochemical responses to drought stress, as well as a substantial ability to counteract drought-induced oxidative damage through improved antioxidant defenses [20]. Several studies have demonstrated that AMF may alleviate the effects of drought stress in maize, oat, and durum wheat plants [33,54,55]. Our findings revealed that drought stress substantially inhibited root colonization when incorporated with the M consortium. This observation aligns with prior research [56,57,58]. It is plausible that the mycorrhizae used in our study exhibited “pre-adaptation” to low water content, which may account for the limited unfavorable reactions of M to water stress, considering the duration and severity of the drought [59]. DS can inhibit wheat growth substantially. It lowers cereal and pseudo-cereal production, biomass, and growth, according to several studies [60,61]. DS influences the leaf area, chlorophyll content, photosynthetic effectiveness, plant stature, and the opening of stomatal cells. According to Iqbal et al. [61], the crop yield, food quality, and nutritional value are all negatively affected by drought. However, its tolerance in plants requires modifications at several levels, including at the entire plant, tissue, physiological, and molecular levels [62,63]. The current study demonstrated that the application of biofertilizers in the rhizosphere of wheat plants produced favorable changes in the shoot and spikelet height, leaf and seed number, root length, shoot, root, and spikelet dry weight. This improvement might be attributed to increased cell division and growth production of hormones [64]. The beneficial effects of mycorrhizal and PGPR associations, as well as compost amendment, on wheat growth under water deficit conditions may be attributed to enhanced nutrient uptake, particularly of immobile nutrients such as P and N present in the substrate [65]. Previous research showed that when plants were stressed by drought, date palm and other AMF-inoculated plants acquired more N and P in their leaves than non-mycorrhizal plants and that the agronomic, physiological, and chemical characteristics were greatly enhanced with the application of the triple combination of PGPR + AMF + compost [33,66]. The most significant results were detected in plants treated with the triple combination CMR for the growth traits of wheat plants (mainly leaves number, spikelet and shoot height, root length, and spikelet dry weight) under 30% ETc, which is in line with several studies that have reported the major role of microbial and non-microbial biofertilizers, such as R, M, and C, in enhancing plant tolerance to abiotic stresses [67]. Previous findings [68,69] showed that the concurrent inoculation of various microbial strains, along with organic amendment, can boost the performance of plants. Radzikowska-Kujawska et al. [70] demonstrated that inoculation with PGPR considerably increased lettuce growth and resistance to abiotic stress due to its phosphate solubilizing capabilities. For instance, compared to the control, the C, R + C, and M + C treatments resulted in a considerable increase in the shoot biomass of lettuce plants of 160%, 120%, and 156%, respectively [71]. Moreover, M may stimulate plant development by altering plant photosynthetic and antioxidant features [72]. Furthermore, mycorrhized plants have been reported to show enhanced development under both well-watered and water-stressed conditions [73]. This was reportedly due to the improvement in P nutrition [74] and the absorption of water owing to the efficiency of the extraradical mycelium in gathering water from the soil even in water-deficient conditions [75].
Photosynthetic pigments serve as vital markers of plant performance under environmental stress [76]. Drought stress impacts wheat plant physiology and growth features, leading to reductions in the leaf area, stomatal conductance, and over-excitation of the photosystems. This leads to photo-oxidative damage in photosystem II, which contains chlorophylls [77]. Moreover, plants treated with biofertilizers exhibited improved physiological traits, particularly photosynthetic machinery, and mitigated the harmful effects of stress [78]. This enhancement may be attributed to the activation of enzymes involved in pigment production [79], ultimately leading to improved photosynthetic performance. Studies have shown a considerable correlation between high photosynthetic efficiency and effective water circulation in tissues [80]. Additionally, abscisic acid may induce a hormonal signal that improves gas exchange in lettuce plants during water stress [81].
Indeed, the obtained results showed that the Aoufous consortium, either provided alone or combined with PGPR and/or compost, led to a significant increase in the gs, Fv/Fm, total chlorophyll, and carotenoids, potentially improving the rate of carbon acquisition by wheat leaf tissue. These findings align with C3 crops cultivated under well-irrigated and water-stressed conditions [56]. Previous research under water constraint revealed that plants treated with M had greater stomatal conductivity than control plants [82,83]. Moreover, they demonstrated that stressed plants could exhibit significantly decreased stomatal conductance. M, either applied alone or in combination with R, resulted in much greater gs increases than uninoculated stressed control plants. According to Meddich et al. [84], plants infected with the Aoufous consortia had greater stomatal conductance, which correlated with increased Ψ leaf and osmotic adjustment. In this context, it is important to remember that M affects the hormonal balance in plants, which controls stomatal activity during water stress [85,86]. According to Ouledali et al. [87], plants treated with M had lower ABA levels than control plants under stress circumstances, which might contribute to greater transpiration and root water absorption. Consistent with previous studies, the administration of M increased the water content and stomatal opening, resulting in better photosystem II (PSII) efficiency (Fv/Fm) under water stress conditions [82,88]. Our findings showed that both the AMF and the two PGPR consortia utilized in this work boosted the Fv/Fm in water-stressed plants, implying that inoculated plants maintained normal light energy usage in photochemical processes despite severe water stress. Previous research has demonstrated that AMF inoculation can increase the maximal quantum yield of PSII under water stress [89,90]. It has been confirmed that the inoculation of plants with M can enhance the drought tolerance in control plants by mitigating damage to the photosystem reaction centers to some degree [91]. Non-inoculated plants subjected to drought stress showed a decrease in the gs, which could be interpreted as a rapid mechanism against dehydration. However, the application of microbial biofertilizers led to increased water uptake, gs, and Fv/Fm, while also reducing the degradation of chlorophyll and carotenoids under drought stress conditions. This improvement in the physiological traits suggests a better photosynthetic apparatus performance, leading to increased CO2 assimilation for photosynthesis [21].
Most plants respond to abiotic stress by accumulating metabolites [92]. According to Saxena et al. [93], plants collect suitable solutes such as soluble sugars, proline, glycine betaine, organic acids, potassium, calcium, and so on to promote water absorption by reducing the cytoplasmic osmotic potential. In our investigation, wheat plants showed elevated levels of soluble sugars in the presence of drought stress, which were further boosted by the application of PGPR and AMF. This response may serve as a defensive mechanism to uphold osmotic equilibrium and minimize free radical damage [94]. Additionally, research by Pasbani et al. [95] demonstrated that the co-inoculation of AMF and PGPR produces synergistic and cumulative effects. It has been reported that the combined action of AMF and PGPR enhanced the production of organic acids, soluble sugars, and antioxidant enzymes while reducing the Na+ levels in plants experiencing water stress [96]. One of the ways that plants respond to abiotic stress is by accumulating certain molecules (proteins, sugars) that can regulate osmoregulation [97]. This facilitates the absorption of water by soils with very low moisture levels [98]. Our research shows that DS significantly increased the sugars and proteins in wheat plants treated with biofertilizers, either alone or in combination, compared to well-irrigated plants. These results concur with those of other studies [99,100]. Furthermore, the improved photosynthetic efficiency was associated with the increased accumulation of total soluble sugars (TSS) brought about by treatments comprising compost (C) and compost plus mycorrhizal fungus (CM). Increased levels of organic acids and increased activities of sucrose phosphate synthase, acid invertase, and alpha and beta amylases followed this improvement. These circumstances show an increased need for carbon, which arbuscular mycorrhizal fungi (AMF) help to supply [101]. The protein contents of wheat plants were positively influenced by the combined presence of M and R, consistent with the findings of Ouhaddou et al. [102]. This could also be explained by the over-expression of genes encoding aquaporins, which contribute positively to plant water uptake [103]. In the present study, water deficit significantly increased the proline contents. However, a pronounced increase was observed in the M and CM treatments, which accords with the study of Begum et al. [104], who showed that the inoculation of AMF improved the proline content in Nicotiana tabacum under drought stress. The considerable quantity of proline obtained might be attributed to the ability of M and/or C to promote its production by increasing the activity of the pyrroline-5-carboxylate synthase (P5CS) gene [105]. Such a rise in proline under DS helps wheat plants endure drought, particularly by safeguarding the photosynthetic machinery by sustaining overall cellular integrity (proteins, DNA, and lipids) [106] and/or maintaining a water balance in the plant [107].
Plants synthesize antioxidant enzymes as a means of counteracting the effects of reactive oxygen species (ROS) in response to oxidative stress [104,108,109]. Our study found that DS significantly enhanced the antioxidant activity of POX and PPO in plants that received each of the three biofertilizers concurrently. Wheat plants exposed to severe drought stress exhibited a significant increase in the PPO and POX activities due to microbial action and/or compost association as compared to the control group. When the soil was inoculated with MR, the POX activity was higher, while plants inoculated with M alone or in combination with R showed higher PPO activity. Antioxidant enzymes and drought stress have a favorable association, according to research by Benaffari et al. [21]. Stressed plants have been found to have greater concentrations of antioxidant enzymes than non-stressed plants, and it has been suggested that combining M and PGPR will increase the production of compounds for antioxidant enzymes and ROS scavenging, as documented by previous findings [20,110,111]. Plants synthesize antioxidant molecules such as reduced glutathione (GSH), carotenoids, and phenolic substances. Superoxide dismutase, catalase, ascorbate peroxidase, glutathione reductase, and glutathione peroxidase are some of the most important antioxidant enzymes [112]. The elevated activity of these antioxidant enzymes enhances the osmotic adjustment capacity, thereby preserving the water status in Triticum aestivum L. and Cucumis sativus L. [113]. This is achieved through the reduction of lipid peroxidation and membrane damage, as well as the restoration of cell turgor. However, the mechanism reduces the damage caused by stress markers, such as MDA and H2O2 [114,115]. In our investigation, the use of biofertilizers dramatically lessened these adverse effects as compared to the controls. Ouhaddou et al. [115] showed that in conditions of severe water stress (33% FC), lettuce plants treated with PGPR and/or AMF and/or compost, applied individually or in combination, showed a significant reduction in the values of H2O2 and MDA compared to the non-biostimulated plants and increased levels of antioxidant enzymes. In addition, using microbial biofertilizers enabled wheat plants to overcome and alleviate this oxidative stress through substantial levels of antioxidant defensive mechanisms, water availability, osmoregulation, and scavenger genes provided by microorganisms [116,117,118]. On the other hand, inoculating microorganisms and amending soil with compost improves soil properties such as the total organic matter (TOM), total organic carbon (TOC), available phosphorus (AP), total nitrogen content (N), and glomalin content resulting in better wheat growth. Our results demonstrated that the initial analysis of the agricultural soil revealed low TOM and micronutrients, and the application of the recommended biofertilizers enhanced the soil’s physicochemical parameters. Treating wheat plants with compost and/or microorganisms leads to greater harvests owing to the improved soil structure. Ren et al. [119] suggest that bacterial biofertilizers, alone or in combination, modify the soil characteristics in the rhizosphere via stabilizing the pH and electric conductivity (EC). The study by Sun et al. [120] found that compost can help preserve microbial populations from drought stress. Gaiotti et al. [121] reported that adding microbes and compost significantly enhanced the soil quality, particularly the TOM and mineral nutrients. In our study, when the treatments were combined with compost and/or PGPR, P uptake occurred more efficiently than when AMF alone was used. The substantial solubilization of AP by applying microorganisms may account for the high quantities of AP seen in soil treated with the triple combination (CRM) under water stress conditions. This could be due to the enhanced ability of plants to absorb compost-derived nutrients through AMF and/or to the greater P availability in compost, as well as the ability of PGPR to solubilize phosphorus and increase its bioavailability in the root zone, which is in line with the study by Etesami et al. [122]. The ability of plants to absorb P from organic matter is affected by AMF directly or indirectly, while they can also increase the number of bacteria that are phosphate solubilizers and/or mineralize P in the enzyme phosphatase [123]. It is well known that the availability of nutrients, especially inorganic N, is essential for the rate at which AMF colonize plants and for their function inside them. In line with our findings, a research study by Liu et al. [124] showed that compost supplied nitrogen to the soil and AMF inoculation had a significant role in transporting nitrogen to the plant through the hyphae. Through the mineralization process, the soil’s richness in organic matter might provide a sufficient supply of important elements, such as nitrogen [125]. Moreover, AMF may enhance the soil’s chemical and nutritional qualities by a variety of methods, such as the aggregates, soil structure, and P solubilization by the release of glomalin. Important studies demonstrated the connection between the glomalin concentration of soil and its physicochemical characteristics. However, the production of glomalin is a critical factor in the contribution of AMF to soil aggregation. Glomalin is a glycoprotein substance that improves the soil structure by holding soil particles into aggregates and stabilizing them, and it provides benefits by improving the soil aeration and drainage as well as generating higher microbial activity [124,126]. Glomalin can also be useful for sequestering a variety of hazardous substances/ions [127,128]. Previous studies suggested that organic amendment and AMF may modify the soil structure by inducing the formation of aggregates and improving the soil water availability under stressful conditions [54,129,130]. Furthermore, Trichoderma, which can produce extracellular enzymes, secondary metabolites, and hormones, can increase soil aggregation by increasing root colonization and the amount of AMF spores [131,132]. As indicated in the study by [133], PGPR strains promoted the establishment and expansion of the soil microecological habitat of the pine tree seedlings.
The NDVI is considered a valuable tool for assessing green biomass. Researchers have shown that the applied water regime significantly affected the parameters of the NDVI, with values ranging from 0.03 to 0.71 observed during the early and late stages of wheat plant growth [134]. Similarly, the NDVI at the booting stage ranged from 0.67 to 0.81 in wheat crops [135], while the application of biofertilizers increased the NDVI of buckwheat [136]. From an agronomic perspective, surface irrigation introduces greater variability in water availability for plants. For the soil moisture, it plays a crucial role in several sectors, including agriculture [137]. A study by Bhatt and Kukal [138] showed that accelerated evaporation and reduced soil moisture retention were correlated with elevated soil temperature, a trend in line with our findings. Notably, our observations indicated a lower soil temperature during the growth stages. Variations in the soil moisture levels corresponding to irrigation events could be related to the root system’s ability to penetrate the deep soil layers, as mentioned in a study of winter wheat under different irrigation regimes in North China [139].
5. Conclusions
Overall, our findings suggest that bio-inoculation with compost, rather than separate application, may improve durum wheat cultivation without compromising the yield or soil nutrients in drought situations. Even more intriguingly, the combination of CMR has demonstrated beneficial effects on the plant biomass and colonization of the mycorrhizal/microbial environment. By investigating the agro-physiological and biochemical mechanisms governing microbiome balancing induced by drought, we gained insight into how stress detection and microbiome regulation function in durum wheat. The beneficial effects of these biological agents on the plant biomass, physiology, and tolerance are likely linked to nutrient availability in the soil, the “transportome” in symbiotic relationships, and physio-metabolic mechanisms promoting drought tolerance in durum wheat. Practically, by enhancing the soil fertility, triggering plant growth, and improving wheat’s physiological and biochemical characteristics, these mechanisms consistently improve the soil physicochemical properties, reduce oxidative stress marker accumulation, and enhance enzyme defense. These insights also have effects on the methods of finding microorganisms that are efficient at colonizing the endosphere for long-term protection, which may be used to harness microbial populations to confer drought resistance upon field crops. In the context of rising climatic unpredictability, greater wheat biomass and yield on soil with decreasing moisture content may have major consequences for more sustainable farming practices.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/agronomy14061316/s1, Table S1: Result of the multivariate analysis of variance (MANOVA) test of the independent variables, including drought stress (DS), compost (C) and arbuscular mycorrhizal fungi (M) and plant growth promoting rhizobacteria (R) inoculation and interaction among them.
Author Contributions
Conceptualization, A.M., F.E. and A.B.; methodology, A.M., C.I. and R.O.; validation, R.L. and A.M.; formal analysis, A.B., C.I., A.A., E.A.A., R.U., E.A.B. and F.-E.S.; data curation, L.E.-C. and F.E.; investigation, A.B., A.A. and E.A.B.; writing—original draft preparation, C.I. and F.-E.S.; writing—review and editing, A.M., A.B., A.A., E.A.A., R.L., R.U., R.O., F.E., E.A.B. and L.E.-C.; visualization, R.L.; supervision, A.M.; funding acquisition, E.A.A. and R.U. All authors have read and agreed to the published version of the manuscript.
Funding
The authors wish to thank the researcher supporting project number RSP2024R45 at King Saud University Riyadh, Saudi Arabia, for the financial support.
Data Availability Statement
Data are contained within the article.
Acknowledgments
The authors wish to thank the researchers supporting project number RSP2024R45 at King Saud University Riyadh, Saudi Arabia, for the financial support. The authors gratefully acknowledge the FOSC project (Sus-Agri-CC) from the European Union’s Horizon 2020 research and innovation program under grant agreement N 862555. This project has also benefited from the support of the company “Concamine” (Berrechid, Morocco), which has made available plots of land and its equipment.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Chieb, M.; Gachomo, E.W. The Role of Plant Growth Promoting Rhizobacteria in Plant Drought Stress Responses. BMC Plant Biol. 2023, 23, 407. [Google Scholar] [CrossRef] [PubMed]
- Hirwa, H.; Peng, Y.; Zhang, Q.; Qiao, Y.; Leng, P.; Tian, C.; Yang, G.; Muhirwa, F.; Diop, S.; Kayiranga, A.; et al. Virtual Water Transfers in Africa: Assessing Topical Condition of Water Scarcity, Water Savings, and Policy Implications. Sci. Total Environ. 2022, 835, 155343. [Google Scholar] [CrossRef] [PubMed]
- Balbaa, M.G.; Osman, H.T.; Kandil, E.E.; Javed, T.; Lamlom, S.F.; Ali, H.M.; Kalaji, H.M.; Wróbel, J.; Telesiñski, A.; Brysiewicz, A.; et al. Determination of Morpho-Physiological and Yield Traits of Maize Inbred Lines (Zea mays L.) under Optimal and Drought Stress Conditions. Front. Plant Sci. 2022, 13, 959203. [Google Scholar] [CrossRef]
- Abd El Mageed, T.A.; Semida, W.; Hemida, K.A.; Gyushi, M.A.H.; Rady, M.M.; Abdelkhalik, A.; Merah, O.; Brestic, M.; Mohamed, H.I.; El Sabagh, A.; et al. Glutathione-Mediated Changes in Productivity, Photosynthetic Efficiency, Osmolytes, and Antioxidant Capacity of Common Beans (Phaseolus vulgaris) Grown under Water Deficit. PeerJ 2023, 11, 15343. [Google Scholar] [CrossRef] [PubMed]
- Vijayaraghavareddy, P.; Lekshmy, S.V.; Struik, P.C.; Makarla, U.; Yin, X.; Sreeman, S. Production and Scavenging of Reactive Oxygen Species Confer to Differential Sensitivity of Rice and Wheat to Drought Stress. Crop. Environ. 2022, 1, 15–23. [Google Scholar] [CrossRef]
- Rady, M.M.; Boriek, S.H.K.; El-Mageed, T.A.A.; El-Yazal, M.A.S.; Ali, E.F.; Hassan, F.A.S.; Abdelkhalik, A. Exogenous Gibberellic Acid or Dilute Bee Honey Boosts Drought Stress Tolerance in Vicia faba by Rebalancing Osmoprotectants, Antioxidants, Nutrients, and Phytohormones. Plants 2021, 10, 748. [Google Scholar] [CrossRef]
- Snapp, S.; Morrone, V. Perennial Wheat; Montana State University Extension: Bozeman, MT, USA, 2014; pp. 1–3. [Google Scholar]
- Abdollahi, M.R.; Zaefarian, F.; Hunt, H.; Anwar, M.N.; Thomas, D.G.; Ravindran, V. Wheat Particle Size, Insoluble Fibre Sources and Whole Wheat Feeding Influence Gizzard Musculature and Nutrient Utilisation to Different Extents in Broiler Chickens. J. Anim. Physiol. Anim. Nutr. 2019, 103, 146–161. [Google Scholar] [CrossRef]
- Lachuga, Y.; Meskhi, B.; Pakhomov, V.; Semenikhina, Y.; Kambulov, S.; Rudoy, D.; Maltseva, T. Experience in the Cultivation of a New Perennial Cereal Crop Trititrigia in the Conditions of South of the Rostov Region. Agriculture 2023, 13, 605. [Google Scholar] [CrossRef]
- Tadesse, W.; Zegeye, H.; Debele, T.; Kassa, D.; Shiferaw, W.; Solomon, T.; Negash, T.; Geleta, N.; Bishaw, Z.; Assefa, S. Wheat production and breeding in Ethiopia: Retrospect and prospects. Crop Breed. Genet. Genom. 2022, 4, 3. [Google Scholar]
- Faostat Global Cereal Production Forecast Unchanged from Last Month, Utilization and Trade up, and Stocks down But Still Foreseen to Reach an All-Time High. 2023. Available online: https://uga.ua/en/news/global-cereal-production-forecast-unchanged-from-last-month-utilization-and-trade-up-and-stocks-down-but-still-foreseen-to-reach-an-all-time-high/ (accessed on 14 February 2024).
- Yang, D.; Li, S.; Kang, S.; Du, T.; Guo, P.; Mao, X.; Tong, L.; Hao, X.; Ding, R.; Niu, J. Effect of Drip Irrigation on Wheat Evapotranspiration, Soil Evaporation and Transpiration in Northwest China. Agric. Water Manag. 2020, 232, 106001. [Google Scholar] [CrossRef]
- Kavianand, G.; Nivas, V.M.; Kiruthika, R.; Lalitha, S. Smart Drip Irrigation System for Sustainable Agriculture. In Proceedings of the IEEE International Conference on Technological Innovations in ICT for Agriculture and Rural Development, TIAR 2016, Chennai, India, 15–16 July 2016; pp. 19–22. [Google Scholar] [CrossRef]
- Azad, N.; Behmanesh, J.; Rezaverdinejad, V.; Khodaverdiloo, H.; Thompson, S.E.; Mallants, D.; Ramos, T.B.; He, H. CNN Deep Learning Performance in Estimating Nitrate Uptake by Maize and Root Zone Losses under Surface Drip Irrigation. J. Hydrol. 2023, 625, 130148. [Google Scholar] [CrossRef]
- Illés, Á.; Szabó, A.; Mousavi, S.M.N.; Bojtor, C.; Vad, A.; Harsányi, E.; Sinka, L. The Influence of Precision Dripping Irrigation System on the Phenology and Yield Indices of Sweet Maize Hybrids. Water 2022, 14, 2480. [Google Scholar] [CrossRef]
- FAO. Chapitre 6: Drip Irrigation. 2012. Available online: https://www.fao.org/4/s8684e/s8684e07.htm (accessed on 14 February 2024).
- Tahiri, A.; Meddich, A.; Raklami, A.; Alahmad, A.; Bechtaoui, N.; Anli, M.; Göttfert, M.; Heulin, T.; Achouak, W.; Oufdou, K. Assessing the Potential Role of Compost, PGPR, and AMF in Improving Tomato Plant Growth, Yield, Fruit Quality, and Water Stress Tolerance. J. Soil Sci. Plant Nutr. 2021, 22, 743–764. [Google Scholar] [CrossRef]
- Latef, A.A.H.A.; Chaoxing, H. Arbuscular Mycorrhizal Influence on Growth, Photosynthetic Pigments, Osmotic Adjustment and Oxidative Stress in Tomato Plants Subjected to Low Temperature Stress. Acta Physiol. Plant 2011, 33, 1217–1225. [Google Scholar] [CrossRef]
- Boutaj, H.; Meddich, A.; Wahbi, S.; Moukhli, A.; El Alaoui-Talibi, Z.; Douira, A.; Filali-Maltouf, A.; El Modafar, C. Effect of Arbuscular Mycorrhizal Fungi on Verticillium Wilt Development of Olive Trees Caused by Verticillium dahliae. Res. J. Biotechnol. 2019, 14, 79–88. [Google Scholar]
- Zou, Y.N.; Wu, Q.S.; Kuča, K. Unravelling the Role of Arbuscular Mycorrhizal Fungi in Mitigating the Oxidative Burst of Plants under Drought Stress. Plant Biol. 2021, 23, 50–57. [Google Scholar] [CrossRef]
- Benaffari, W.; Boutasknit, A.; Anli, M.; Ait-El-mokhtar, M.; Ait-Rahou, Y.; Ben-Laouane, R.; Ahmed, H.; Mitsui, T.; Baslam, M.; Meddich, A. The Native Arbuscular Mycorrhizal Fungi and Vermicompost-Based Organic Amendments Enhance Soil Fertility, Growth Performance, and the Drought Stress Tolerance of Quinoa. Plants 2022, 11, 393. [Google Scholar] [CrossRef]
- Raklami, A.; Meddich, A.; Pajuelo, E.; Marschner, B.; Heinze, S.; Oufdou, K. Combined Application of Marble Waste and Beneficial Microorganisms: Toward a Cost Effective Approach for Restoration of Heavy Metals Contaminated Sites. Envirion. Sci. Pollut. Res. 2022, 29, 45683–45697. [Google Scholar] [CrossRef]
- Singh, V.K.; Malhi, G.S.; Kaur, M.; Singh, G.; Jatav, H.S. Use of Organic Soil Amendments for Improving Soil Ecosystem Health and Crop Productivity. In Ecosystem Services: Types, Management and Benefits; Nova Science Publishers, Inc.: Hauppauge, NY, USA, 2022. [Google Scholar]
- Ayangbenro, A.S.; Chukwuneme, C.F.; Ayilara, M.S.; Kutu, F.R.; Khantsi, M.; Adeleke, B.S.; Glick, B.R.; Babalola, O.O. Harnessing the Rhizosphere Soil Microbiome of Organically Amended Soil for Plant Productivity. Agronomy 2022, 12, 3179. [Google Scholar] [CrossRef]
- Meddich, A.; Oufdou, K.; Boutasknit, A.; Raklami, A.; Tahiri, A.; Ben-Laouane, R.; Ait-El-Mokhtar, M.; Anli, M.; Mitsui, T.; Wahbi, S.; et al. Use of Organic and Biological Fertilizers as Strategies to Improve Crop Biomass, Yields and Physicochemical Parameters of Soil. In Nutrient Dynamics for Sustainable Crop Production; Springer: Singapore, 2019; pp. 247–288. ISBN 9789811386602. [Google Scholar]
- Sawicka, B.; Pszczotkowski, P.; Barbas, P.; Skiba, D.; Bienia, B. Biotechnology: Role in Ecological Sustainability and Research; John Wiley & Sons: Hoboken, NJ, USA, 2022. [Google Scholar]
- Ikan, C.; Ben-Laouane, R.; Ouhaddou, R.; Ghoulam, C.; Meddich, A. Co-Inoculation of Arbuscular Mycorrhizal Fungi and Plant Growth-Promoting Rhizobacteria Can Mitigate the Effects of Drought in Wheat Plants (Triticum durum). Plant Biosyst. 2023, 157, 907–919. [Google Scholar]
- Arora, S.; Jha, P.N. Drought-Tolerant Enterobacter Bugandensis WRS7 Induces Systemic Tolerance in Triticum aestivum L. (Wheat) under Drought Conditions. J. Plant Growth Regul. 2023, 42, 7715–7730. [Google Scholar] [CrossRef]
- Mushtaq, Z. PGPR: Present Role, Mechanism of Action and Future Prospects along Bottlenecks in Commercialization. EQA-Int. J. Environ. Qual. 2020, 41, 9–15. [Google Scholar] [CrossRef]
- Chandwani, S.; Amaresan, N. Role of ACC Deaminase Producing Bacteria for Abiotic Stress Management and Sustainable Agriculture Production. Envirion. Sci. Pollut. Res. 2022, 29, 22843–22859. [Google Scholar] [CrossRef]
- Camaille, M.; Fabre, N.; Clément, C.; Barka, E.A. Advances in Wheat Physiology in Response to Drought and the Role of Plant Growth Promoting Rhizobacteria to Trigger Drought Tolerance. Microorganism 2021, 9, 687. [Google Scholar] [CrossRef]
- Allen, R.; Pereira, L.; Raes, D.; Smith, M. Crop Evapotranspiration-Guidelines for Computing Crop Water Requirements-FAO Irrigation and Drainage Paper 56; FAO: Rome, Italy, 1998. [Google Scholar]
- Anli, M.; Baslam, M.; Tahiri, A.; Raklami, A.; Symanczik, S.; Boutasknit, A.; Ait-El-Mokhtar, M.; Ben-Laouane, R.; Toubali, S.; Ait Rahou, Y.; et al. Biofertilizers as Strategies to Improve Photosynthetic Apparatus, Growth, and Drought Stress Tolerance in the Date Palm. Front. Plant Sci. 2020, 11, 516818. [Google Scholar] [CrossRef]
- Meddich, A.; Elouaqoudi, F.; Khadra, A.; Bourzik, W. Valorisation Des Déchets d’origine Végétale et Industrielle Par Compostage. Rev. Compos. Matériaux Avancées 2016, 26, 451–469. [Google Scholar] [CrossRef]
- Rouzbeh, R.; Daneshian, J.; Farahani, H.A. Super Nitro plus Influence on Yield and Yield Components of Two Wheat Cultivars under NPK Fertilizer Application. J. Plant Breed. Crop Sci. 2009, 1, 293–297. [Google Scholar]
- Phillips, J.M.; Hayman, D.S. Improved Procedures for Clearing Roots and Staining Parasitic and Vesicular-Arbuscular Mycorrhizal Fungi for Rapid Assessment of Infection. Trans. Br. Mycol. Soc. 1970, 55, 158-IN18. [Google Scholar] [CrossRef]
- Trouvelot, A. Measure du taux de mycorrhization d’un systeme radiculaire. Recherche de methods d’estimation ayant une signification fonctionnelle. In Physiological and Genetical Aspects of Mycorrhizae, Proceedings of the 1st European Symposium on Mycorrhizae, Dijon, France, 1–5 July 1985; INRA: Paris, France, 1986; pp. 217–221. [Google Scholar]
- Harley, P.C.; Loreto, F.; Di Marco, G.; Sharkey, T.D. Theoretical Considerations When Estimating the Mesophyll Conductance to CO2 Flux by Analysis of the Response of Photosynthesis to CO2. Plant Physiol. 1992, 98, 1429–1436. [Google Scholar] [CrossRef]
- Lichtenthaler, H. Chlorophylls and Carotenoids: Pigments of Photosynthetic Biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric Method for Determination of Sugars and Related Substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Bradford, M. A Rapid and Sensitive Method for the Quantitation Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding MARION. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Tejera García, A.; Olivera, M.; Iribarne, C.; Lluch, C. Partial Purification and Characterization of a Non-Specific Acid Phosphatase in Leaves and Root Nodules of Phaseolus vulgaris. Plant Physiol. Biochem. 2004, 42, 585–591. [Google Scholar] [CrossRef] [PubMed]
- Polle, A.; Otter, T.; Seifert, F. Apoplastic Peroxidases and Lignification in Needles of Norway Spruce (Picea abies L.). Plant Physiol. 1994, 106, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Carillo, P. Nitrogen Metabolism in Durum Wheat under Salinity: Accumulation of Proline and Glycine Betaine. Funct. Plant Biol. 2008, 35, 412–426. [Google Scholar] [CrossRef] [PubMed]
- Hori, K.; Wada, A.; Shibuta, T. Changes in Phenoloxidase Activities of the Galls on Leaves of Ulmus Davidana Formed by Tetraneura fusiformis (Homoptera: Eriosomatidae). Appl. Entomol. Zool. 1997, 32, 365–371. [Google Scholar] [CrossRef]
- Dhindsa, R.S.; Matowe, W. Drought Tolerance in Two Mosses: Correlated with Enzymatic Defence Against Lipid Peroxidation. J. Exp. Bot. 1981, 32, 79–91. [Google Scholar] [CrossRef]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative Stress and Some Antioxidant Systems in Acid Rain-Treated Bean Plants Protective Role of Exogenous Polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Aubert, G. Méthodes D’analyses des Sols, 2nd ed.; Methods of Soil Analysis; Centre Régional de Documentation Pédagogique: Marseille, France, 1978; p. 191. [Google Scholar]
- Bremner, J.M. Determination of Nitrogen in Soil by the Kjeldahl Method. J. Agric. Sci. 1960, 55, 11–33. [Google Scholar] [CrossRef]
- Cornejo, P.; Meier, S.; Borie, G.; Rillig, M.C.; Borie, F. Glomalin-Related Soil Protein in a Mediterranean Ecosystem Affected by a Copper Smelter and Its Contribution to Cu and Zn Sequestration. Sci. Total Environ. 2008, 406, 154–160. [Google Scholar] [CrossRef]
- Kour, D.; Yadav, A.N. Bacterial Mitigation of Drought Stress in Plants: Current Perspectives and Future Challenges. Curr. Microbiol. 2022, 79, 248. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Lu, X.; Gu, S.; Guo, X. Improving Nutrient and Water Use Efficiencies Using Water-Drip Irrigation and Fertilization Technology in Northeast China. Agric. Water Manag. 2020, 241, 106352. [Google Scholar] [CrossRef]
- Yang, D.; Li, S.; Wu, M.; Yang, H.; Zhang, W.; Chen, J.; Wang, C.; Huang, S.; Zhang, R.; Zhang, Y. Drip Irrigation Improves Spring Wheat Water Productivity by Reducing Leaf Area While Increasing Yield. Eur. J. Agron. 2023, 143, 126710. [Google Scholar] [CrossRef]
- Ben-Laouane, R.; Baslam, M.; Ait-El-mokhtar, M.; Anli, M.; Boutasknit, A.; Ait-Rahou, Y.; Toubali, S.; Mitsui, T.; Oufdou, K.; Wahbi, S.; et al. Potential of Native Arbuscular Mycorrhizal Fungi, Rhizobia, and/or Green Compost as Alfalfa (Medicago sativa) Enhancers under Salinity. Microorganisms 2020, 8, 1695. [Google Scholar] [CrossRef]
- Boutasknit, A.; Baslam, M.; Ait-El-mokhtar, M.; Anli, M.; Ben-Laouane, R.; Douira, A.; El Modafar, C.; Mitsui, T.; Wahbi, S.; Meddich, A. Arbuscular Mycorrhizal Fungi Mediate Drought Tolerance and Recovery in Two Contrasting Carob (Ceratonia siliqua L.) Ecotypes by Regulating Stomatal, Water Relations, and (in)Organic Adjustments. Plants 2020, 9, 80. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Hu, Y.; Zhang, K.; Tian, C.; Guo, J. Arbuscular Mycorrhizal Fungi Improve Plant Growth of Ricinus communis by Altering Photosynthetic Properties and Increasing Pigments under Drought and Salt Stress. Ind. Crops Prod. 2018, 117, 13–19. [Google Scholar] [CrossRef]
- Zhang, F.; Zou, Y.N.; Wu, Q.S.; Kuča, K. Arbuscular Mycorrhizas Modulate Root Polyamine Metabolism to Enhance Drought Tolerance of Trifoliate Orange. Environ. Exp. Bot. 2020, 171, 103926. [Google Scholar] [CrossRef]
- He, J.D.; Zou, Y.N.; Wu, Q.S.; Kuča, K. Mycorrhizas Enhance Drought Tolerance of Trifoliate Orange by Enhancing Activities and Gene Expression of Antioxidant Enzymes. Sci. Hortic. 2020, 262, 108745. [Google Scholar] [CrossRef]
- Quiroga, G.; Erice, G.; Aroca, R.; Chaumont, F.; Ruiz-lozano, J.M. Enhanced Drought Stress Tolerance by the Arbuscular Mycorrhizal Symbiosis in a Drought-Sensitive Maize Cultivar Is Related to a Broader and Differential Regulation of Host Plant Aquaporins than in a Drought-Tolerant Cultivar. Front. Plant Sci. 2017, 8, 268043. [Google Scholar] [CrossRef]
- Shah, Z.H.; Rehman, H.M.; Akhtar, T.; Daur, I.; Nawaz, M.A.; Ahmad, M.Q.; Rana, I.A.; Atif, R.M. Redox and Ionic Homeostasis Regulations against Oxidative, Salinity and Drought Stress in Wheat (A Systems Biology Approach). Front. Genet. 2017, 8, 141. [Google Scholar] [CrossRef]
- Iqbal, M.S.; Singh, A.K.; Ansari, M.I. Effect of Drought Stress on Crop Production. In New Frontiers in Stress Management for Durable Agriculture; Springer: Singapore, 2020; pp. 35–47. [Google Scholar]
- Farooq, M.; Aziz, T.; Wahid, A.; Lee, D.J.; Siddique, K.H. Chilling Tolerance in Maize: Agronomic and Physiological Approaches. Crop. Pasture Sci. 2009, 60, 501–516. [Google Scholar] [CrossRef]
- Sakran, R.; Ghazy, M.; Rehan, M.; Plants, A.A. Molecular Genetic Diversity and Combining Ability for Some Physiological and Agronomic Traits in Rice under Well-Watered and Water-Deficit Conditions. Plants 2022, 11, 702. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Singh, G.; Khanna, V. Growth of Lentil (Lens Culinaris Medikus) as Influenced by Phosphorus, Rhizobium and Plant Growth Promoting Rhizobacteria. Indian J. Agric. Res. 2016, 50, 567–572. [Google Scholar] [CrossRef]
- Darakeh, S.A.S.S.; Weisany, W.; Tahir, N.A.R.; Schenk, P.M. Physiological and Biochemical Responses of Black Cumin to Vermicompost and Plant Biostimulants: Arbuscular Mycorrhizal and Plant Growth-Promoting Rhizobacteria. Ind. Crops Prod. 2022, 188, 115557. [Google Scholar] [CrossRef]
- Niaz, H.; Mushtaq, Z.; Jaffar, M.; Zhang, J.; Asghar, H. Mycorrhizae-Rhizobacterial Interaction to Enhance the Growth, Development, Nutrient Content, and Physiological Attributes of Okra in Compost Amended Soil. Res. Sq. 2023. [Google Scholar] [CrossRef]
- Bhardwaj, D.; Ansari, M.W.; Sahoo, R.K.; Tuteja, N. Biofertilizers Function as Key Player in Sustainable Agriculture by Improving Soil Fertility, Plant Tolerance and Crop Productivity. Microb. Cell Fact. 2014, 13, 66. [Google Scholar] [CrossRef] [PubMed]
- Azemi-Ardakani, M.; Dehestani-Ardakani, M.; Zarei, A.; Soltani-Gerdfaramarzi, S. Influence of Different Soil Amendments on Drought Stress Tolerance of Maclura Pomifera. Plant Physiol. Rep. 2020, 25, 405–417. [Google Scholar] [CrossRef]
- Furlan, A.; Bianucci, E.; Sequeira, M.; Álvarez, L.; Peralta, J.M.; Valente, C.; Guarnieri, V.; Castro, S. Combined Application of Microbial and Non-Microbial Biostimulants to Improve Growth of Peanut Plants Exposed to Abiotic Stresses. In Microbial Probiotics for Agricultural Systems; Springer: Cham, Switzerland, 2019; pp. 239–256. [Google Scholar] [CrossRef]
- Radzikowska-Kujawska, D.; John, P.; Piechota, T.; Nowicki, M.; Kowalczewski, P.Ł. Response of Winter Wheat (Triticum aestivum L.) to Selected Biostimulants under Drought Conditions. Agriculture 2023, 13, 121. [Google Scholar] [CrossRef]
- Raklami, A.; Tahiri, A.; Bechtaoui, N.; Abdelhay, E.G.; Pajuelo, E.; Baslam, M.; Meddich, A.; Oufdou, K. Restoring the Plant Productivity of Heavy Metal-Contaminated Soil Using Phosphate Sludge, Marble Waste, and Beneficial Microorganisms. J. Envirion. Sci. 2021, 99, 210–221. [Google Scholar] [CrossRef]
- Yang, Y.; Han, X.; Liang, Y.; Ghosh, A.; Chen, J.; Tang, M. The Combined Effects of Arbuscular Mycorrhizal Fungi (AMF) and Lead (Pb) Stress on Pb Accumulation, Plant Growth Parameters, Photosynthesis, and Antioxidant Enzymes in Robinia Pseudoacacia L. PLoS ONE 2015, 10, e0145726. [Google Scholar] [CrossRef]
- Liu, C.; Zhang, F.; Zhang, D.; Srivastava, A.K.; Wu, Q. Mycorrhiza Stimulates Root-Hair Growth and IAA Synthesis and Transport in Trifoliate Orange under Drought Stress. Sci. Rep. 2018, 8, 1978. [Google Scholar] [CrossRef]
- Le Pioufle, O.; Ganoudi, M.; Calonne-salmon, M.; Dhaou, F. Ben Rhizophagus irregularis MUCL 41833 Improves Phosphorus Uptake and Water Use Efficiency in Maize Plants During Recovery from Drought Stress. Front. Plant. Sci. 2019, 10, 459006. [Google Scholar] [CrossRef]
- Bahadur, A.; Batool, A.; Nasir, F.; Jiang, S.; Mingsen, Q.; Zhang, Q. Mechanistic Insights into Arbuscular Mycorrhizal Fungi-Mediated Drought Stress Tolerance in Plants. Int. J. Mol. Sci. 2019, 20, 4199. [Google Scholar] [CrossRef]
- Bashir, S.S.; Hussain, A.; Hussain, S.J.; Ali, O.; Nabi, S.Z.; Dar, N.A.; Baloch, F.S. Plant Drought Stress Tolerance: Understanding Its Physiological, Biochemical and Molecular Mechanisms. Biotechnol. Biotechnol. Equip. 2022, 35, 1912–1925. [Google Scholar] [CrossRef]
- Baccari, S.; Elloumi, O.; Chaari-Rkhis, A.; Fenollosa, E.; Morales, M.; Drira, N.; Ben Abdallah, F.; Fki, L.; Munné-Bosch, S. Linking Leaf Water Potential, Photosynthesis and Chlorophyll Loss with Mechanisms of Photo- and Antioxidant Protection in Juvenile Olive Trees Subjected to Severe Drought. Front. Plant Sci. 2020, 11, 614144. [Google Scholar] [CrossRef] [PubMed]
- Raheleh, K.; Ghassemi, S.; Asghari, B. Bio-Fertilizer Improves Physio-Biochemical Characteristics and Grain Yield of Safflower (Carthamus tinctorius L.) under Drought Stress. Russ. Agric. Sci. 2019, 45, 458–463. [Google Scholar] [CrossRef]
- Baslam, M.; Mitsui, T.; Hodges, M.; Priesack, E.; Herritt, M.T.; Aranjuelo, I.; Sanz-Sáez, Á. Photosynthesis in a Changing Global Climate: Scaling Up and Scaling Down in Crops. Front. Plant Sci. 2020, 11, 00882. [Google Scholar] [CrossRef] [PubMed]
- Garcia, A.; Gaju, O.; Bowerman, A.F.; Buck, S.A.; Evans, J.R.; Furbank, R.T.; Gilliham, M.; Millar, A.H.; Pogson, B.J.; Reynolds, M.P.; et al. Enhancing crop yields through improvements in the efficiency of photosynthesis and respiration. New Phytol. 2023, 237, 60–77. [Google Scholar] [CrossRef]
- Ruiz-Lozano, J.M.; Aroca, R.; Zamarreño, Á.M.; Molina, S.; Andreo-Jiménez, B.; Porcel, R.; García-Mina, J.M.; Ruyter-Spira, C.; López-Ráez, J.A. Arbuscular Mycorrhizal Symbiosis Induces Strigolactone Biosynthesis under Drought and Improves Drought Tolerance in Lettuce and Tomato. Plant Cell Environ. 2016, 39, 441–452. [Google Scholar] [CrossRef] [PubMed]
- Augé, R.M.; Toler, H.D.; Saxton, A.M. Arbuscular Mycorrhizal Symbiosis Alters Stomatal Conductance of Host Plants More under Drought than under Amply Watered Conditions: A Meta-Analysis. Mycorrhiza 2015, 25, 13–24. [Google Scholar] [CrossRef]
- Boutasknit, A.; Baslam, M.; Ait-El-Mokhtar, M.; Anli, M.; Ben-Laouane, R.; Ait-Rahou, Y.; Mitsui, T.; Douira, A.; El Modafar, C.; Wahbi, S.; et al. Assemblage of Indigenous Arbuscular Mycorrhizal Fungi and Green Waste Compost Enhance Drought Stress Tolerance in Carob (Ceratonia siliqua L.) Trees. Sci. Rep. 2021, 11, 22835. [Google Scholar] [CrossRef]
- Meddich, A.; Jaiti, F.; Bourzik, W.; El Asli, A.; Hafidi, M. Use of Mycorrhizal Fungi as a Strategy for Improving the Drought Tolerance in Date Palm (Phoenix dactylifera). Sci. Hortic. 2015, 192, 468–474. [Google Scholar] [CrossRef]
- Miransari, M.; Abrishamchi, A.; Khoshbakht, K.; Niknam, V. Plant Hormones as Signals in Arbuscular Mycorrhizal Symbiosis. Crit. Rev. Biotechnol. 2012, 34, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Chitarra, W.; Pagliarani, C.; Maserti, B.; Lumini, E.; Siciliano, I.; Cascone, P.; Schubert, A.; Gambino, G.; Balestrini, R.; Guerrieri, E. Insights on the Impact of Arbuscular Mycorrhizal Symbiosis on Tomato Tolerance to Water Stress. Plant Physiol. 2016, 171, 1009–1023. [Google Scholar] [CrossRef]
- Ouledali, S.; Ennajeh, M.; Ferrandino, A.; Khemira, H.; Schubert, A.; Secchi, F. Influence of Arbuscular Mycorrhizal Fungi Inoculation on the Control of Stomata Functioning by Abscisic Acid (ABA) in Drought-Stressed Olive Plants. S. Afr. J. Bot. 2019, 121, 152–158. [Google Scholar] [CrossRef]
- Sheng, M.; Tang, M.; Chen, H.; Yang, B.; Zhang, F.; Huang, Y. Influence of Arbuscular Mycorrhizae on Photosynthesis and Water Status of Maize Plants under Salt Stress. Mycorrhiza 2008, 18, 287–296. [Google Scholar] [CrossRef]
- Sánchez-Blanco, M.J.; Ferrández, T.; Morales, M.A.; Morte, A.; Alarcón, J.J. Variations in Water Status, Gas Exchange, and Growth in Rosmarinus Officinalis Plants Infected with Glomus Deserticola under Drought Conditions. J. Plant Physiol. 2004, 161, 675–682. [Google Scholar] [CrossRef] [PubMed]
- Yooyongwech, S.; Samphumphuang, T.; Tisarum, R.; Theerawitaya, C.; Cha-Um, S. Arbuscular Mycorrhizal Fungi (AMF) Improved Water Deficit Tolerance in Two Different Sweet Potato Genotypes Involves Osmotic Adjustments via Soluble Sugar and Free Proline. Sci. Hortic. 2016, 198, 107–117. [Google Scholar] [CrossRef]
- Rahbarian, R.; Khavari-Nejad, R.; Ganjeali, A.; Bagheri, A.; Najafi, F. Drought Stress Effects on Photosynthesis, Chlorophyll Fluorescence and Water Relations in Tolerant and Susceptible Chickpea (Cicer arietinum L.) Genotypes. Acta Biol. Cracoviensia Ser. Bot. 2011, 53, 47–56. [Google Scholar] [CrossRef]
- Meddich, A.; Ait Rahou, Y.; Boutasknit, A.; Ait-El-Mokhtar, M.; Fakhech, A.; Lahbouki, S.; Benaffari, W.; Ben-Laouane, R.; Wahbi, S. Role of Mycorrhizal Fungi in Improving the Tolerance of Melon (Cucumus melo) under Two Water Deficit Partial Root Drying and Regulated Deficit Irrigation. Plant Biosyst. 2022, 156, 469–479. [Google Scholar] [CrossRef]
- Saxena, R.; Kumar, M.; Tomar, R.S. Plant Responses and Resilience towards Drought and Salinity Stress. Plant Arch. 2019, 19, 50–58. [Google Scholar]
- Ahanger, M.A.; Hashem, A.; Abd-Allah, E.F.; Ahmad, P. Arbuscular Mycorrhiza in Crop Improvement under Environmental Stress. Emerg. Technol. Manag. Crop Stress Toler. 2014, 2, 69–95. [Google Scholar] [CrossRef]
- Pasbani, B.; Salimi, A.; Aliasgharzad, N.; Hajiboland, R. Scientia Horticulturae Colonization with Arbuscular Mycorrhizal Fungi Mitigates Cold Stress through Improvement of Antioxidant Defense and Accumulation of Protecting Molecules in Eggplants. Sci. Hortic. 2020, 272, 109575. [Google Scholar] [CrossRef]
- Begum, N.; Wang, L.; Ahmad, H.; Akhtar, K.; Roy, R.; Khan, M.I.; Zhao, T. Co-Inoculation of Arbuscular Mycorrhizal Fungi and the Plant Growth-Promoting Rhizobacteria Improve Growth and Photosynthesis in Tobacco Under Drought Stress by Up-Regulating Antioxidant and Mineral Nutrition Metabolism. Microb. Ecol. 2022, 83, 971–988. [Google Scholar] [CrossRef] [PubMed]
- Khan, R.; Ma, X.; Shah, S.; Wu, X.; Shaheen, A.; Xiao, L.; Wu, Y.; Wang, S. Drought-Hardening Improves Drought Tolerance in Nicotiana Tabacum at Physiological, Biochemical, and Molecular Levels. BMC Plant Biol. 2020, 20, 486. [Google Scholar] [CrossRef] [PubMed]
- Abobatta, W.F. Plant Responses and Tolerance to Combined Salt and Drought Stress. In Salt and Drought Stress Tolerance in Plants: Signaling Networks and Adaptive Mechanisms; Springer: Cham, Switzerland, 2020; pp. 17–52. [Google Scholar] [CrossRef]
- Ali, N.; Akmal, M. Wheat Growth, Yield, and Quality under Water Deficit and Reduced Nitrogen Supply. A Review. Gesunde Pflanz. 2022, 74, 371–383. [Google Scholar] [CrossRef]
- Sattar, A.; Sher, A.; Ijaz, M.; Ul-Allah, S.; Rizwan, M.S.; Hussain, M.; Jabran, K.; Cheema, M.A. Terminal Drought and Heat Stress Alter Physiological and Biochemical Attributes in Flag Leaf of Bread Wheat. PLoS ONE 2020, 15, e0232974. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Song, F.; Liu, S.; Liu, F. Role of Arbuscular Mycorrhiza in Alleviating Salinity Stress in Wheat (Triticum aestivum L.) Grown under Ambient and Elevated CO2. J. Agron. Crop. Sci. 2016, 202, 486–496. [Google Scholar] [CrossRef]
- Ouhaddou, R.; Ben-Laouane, R.; Lahlali, R.; Anli, M.; Ikan, C.; Boutasknit, A.; Slimani, A.; Oufdou, K.; Baslam, M.; Ait Barka, E.; et al. Application of Indigenous Rhizospheric Microorganisms and Local Compost as Enhancers of Lettuce Growth, Development, and Salt Stress Tolerance. Microorganisms 2022, 10, 1625. [Google Scholar] [CrossRef]
- Santander, C.; Aroca, R.; Cartes, P.; Vidal, G.; Cornejo, P. Aquaporins and Cation Transporters Are Differentially Regulated by Two Arbuscular Mycorrhizal Fungi Strains in Lettuce Cultivars Growing under Salinity Conditions. Plant Physiol. Biochem. 2021, 158, 396–409. [Google Scholar] [CrossRef]
- Begum, N.; Ahanger, M.; Botany, L. AMF Inoculation and Phosphorus Supplementation Alleviates Drought Induced Growth and Photosynthetic Decline in Nicotiana Tabacum by Up-Regulating antioxidant metabolism and osmolyte accumulation. Environ. Exp. Bot. 2020, 176, 104088. [Google Scholar] [CrossRef]
- Wang, H.; Tang, X.; Wang, H.; Shao, H.-B. Proline Accumulation and Metabolism-Related Genes Expression Profiles in Kosteletzkya Virginica Seedlings under Salt Stress. Front. Plant Sci. 2015, 6, 792. [Google Scholar] [CrossRef] [PubMed]
- Kaur, G.; Asthir, B. Proline: A Key Player in Plant Abiotic Stress Tolerance. Biol. Plant 2015, 59, 609–619. [Google Scholar] [CrossRef]
- Ullah, A.; Farooq, M. The Challenge of Drought Stress for Grain Legumes and Options for Improvement. Arch. Agron. Soil Sci. 2022, 68, 1601–1618. [Google Scholar] [CrossRef]
- Ojuederie, O.; Olanrewaju, O.; Babalola, O. Plant Growth Promoting Rhizobacterial Mitigation of Drought Stress in Crop Plants: Implications for Sustainable Agriculture. Agronomy 2019, 9, 712. [Google Scholar] [CrossRef]
- Gupta, A.; Bano, A.; Rai, S.; Mishra, R.; Singh, M.; Stress, S.S.-P. Mechanistic Insights of Plant-Microbe Interaction towards Drought and Salinity Stress in Plants for Enhancing the Agriculture Productivity. Plant Stress 2022, 4, 100073. [Google Scholar] [CrossRef]
- Ali, S.; Moon, Y.; Hamayun, M.; Khan, M.A.; Bibi, K.; Lee, I.; Ali, S.; Moon, Y.; Hamayun, M.; Aaqil, M. Pragmatic Role of Microbial Plant Biostimulants in Abiotic Stress Relief in Crop Plants. J. Plant Interact. 2022, 17, 705–718. [Google Scholar] [CrossRef]
- Das, D.; Bisht, K.; Chauhan, A.; Gautam, S.; Jaiswal, J.P.; Salvi, P.; Lohani, P. Morpho-Physiological and Biochemical Responses in Wheat Foliar Sprayed with Zinc-Chitosan-Salicylic Acid Nanoparticles during Drought Stress. Plant Nano Biol. 2023, 4, 100034. [Google Scholar] [CrossRef]
- Gil-Ortiz, R.; Naranjo, M.Á.; Atares, S.; Vicente, O. Antioxidant Responses of Water-Stressed Cherry Tomato Plants to Natural Biostimulants. Agronomy 2023, 13, 2314. [Google Scholar] [CrossRef]
- Habib, N.; Ali, Q.; Ali, S.; Javed, M.T.; Haider, M.Z.; Perveen, R.; Shahid, M.R.; Rizwan, M.; Abdel-daim, M.M.; Elkelish, A. Use of Nitric Oxide and Hydrogen Peroxide for Better yield of wheat (Triticum aestivum L.) under water deficit conditions: Growth, osmoregulation, and antioxidative defense mechanism. Plants 2020, 9, 285. [Google Scholar] [CrossRef]
- Demirci-Çekiç, S.; Özkan, G.; Avan, A.N.; Uzunboy, S.; Çapanoğlu, E.; Apak, R. Biomarkers of Oxidative Stress and Antioxidant Defense. J. Pharm. Biomed. Anal. 2022, 209, 114477. [Google Scholar] [CrossRef] [PubMed]
- Ouhaddou, R.; Ech-chatir, L.; Anli, M.; Ben-Laouane, R.; Boutasknit, A.; Meddich, A. Secondary Metabolites, Osmolytes and Antioxidant Activity as the Main Attributes Enhanced by Biostimulants for Growth and Resilience of Lettuce to Drought Stress. Gesunde Pflanz. 2023, 75, 1737–1753. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Parvin, K.; Bardhan, K.; Nahar, K.; Anee, T.I. Biostimulants for the Regulation of Reactive Oxygen Species Metabolism in Plants under Abiotic Stress. Cells 2021, 10, 2537. [Google Scholar] [CrossRef] [PubMed]
- Singh, J.S. Impact of PGPR Inoculation on Growth and Antioxidant Status of Wheat under Saline Conditions. Plant Biol. 2012, 14, 605–611. [Google Scholar] [CrossRef] [PubMed]
- Islam, F.; Yasmeen, T.; Ali, Q.; Ali, S.; Saleem, M.; Hussain, S.; Rizvi, H. Ecotoxicology and Environmental Safety In Fl Uence of Pseudomonas Aeruginosa as PGPR on Oxidative Stress Tolerance in Wheat under Zn Stress. Ecotoxicol. Environ. Saf. 2014, 104, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Ren, C.; Kong, C.; Yan, K.; Xie, Z. Transcriptome Analysis Reveals the Impact of Arbuscular Mycorrhizal Symbiosis on Sesbania Cannabina Expose to High Salinity. Sci. Rep. 2019, 9, 2780. [Google Scholar] [CrossRef]
- Sun, D.; Li, K.; Bi, Q.; Zhu, J.; Zhang, Q.; Jin, C.; Lingli, L.; Xianyong, L. Effects of Organic Amendment on Soil Aggregation and Microbial Community Composition during Drying-Rewetting Alternation. Sci. Total Environ. 2017, 574, 735–743. [Google Scholar] [CrossRef] [PubMed]
- Gaiotti, F.; Marcuzzo, P.; Belfiore, N.; Lovat, L.; Fornasier, F.; Tomasi, D. Influence of Compost Addition on Soil Properties, Root Growth and Vine Performances of Vitis vinifera Cv Cabernet Sauvignon. Sci. Hortic. 2017, 225, 88–95. [Google Scholar] [CrossRef]
- Etesami, H.; Jeong, B.R.; Glick, B.R. Contribution of Arbuscular Mycorrhizal Fungi, Phosphate–Solubilizing Bacteria, and Silicon to P Uptake by Plant. Front. Plant Sci. 2021, 12, 699618. [Google Scholar] [CrossRef]
- Iftikhar, A.; Farooq, R.; Akhtar, M.; Khalid, H.; Hussain, N.; Ali, Q. Ecological and Sustainable Implications of Phosphorous—Solubilizing Microorganisms in Soil. Discov. Appl. Sci. 2024, 6, 33. [Google Scholar] [CrossRef]
- Liu, J.; Liu, X.; Zhang, Q.; Li, S.; Sun, Y.; Lu, W.; Ma, C. Response of Alfalfa Growth to Arbuscular Mycorrhizal Fungi and Phosphate-Solubilizing Bacteria under Different Phosphorus Application Levels. AMB Express 2020, 10, 200. [Google Scholar] [CrossRef] [PubMed]
- Raza, A.; Charagh, S.; Abbas, S.; Hassan, M.U.; Saeed, F.; Haider, S.; Sharif, R.; Anand, A.; Corpas, F.J.; Jin, W.; et al. Assessment of Proline Function in Higher Plants under Extreme Temperatures. Plant Biol. 2023, 25, 379–395. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Wang, Q.; Wang, H.; Nie, S.; Liang, Z. Effects of Soil Salinity on the Content, Composition, and Ion Binding Capacity of Glomalin-Related Soil Protein (GRSP). Sci. Total Environ. 2017, 581–582, 657–665. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Xiong, J.; Fang, L.; Han, F.; Zhao, X.; Fan, Q.; Tan, W. Sequestration of Heavy Metals in Soil Aggregates Induced by Glomalin-Related Soil Protein: A Five-Year Phytoremediation Field Study. J. Hazard. Mater. 2022, 437, 129445. [Google Scholar] [CrossRef]
- González-Chávez, M.C.; Carrillo-González, R.; Wright, S.F.; Nichols, K.A. The Role of Glomalin, a Protein Produced by Arbuscular Mycorrhizal Fungi, in Sequestering Potentially Toxic Elements. Environ. Pollut. 2004, 130, 317–323. [Google Scholar] [CrossRef] [PubMed]
- Kohler, J.; Caravaca, F.; Roldán, A. An AM Fungus and a PGPR Intensify the Adverse Effects of Salinity on the Stability of Rhizosphere Soil Aggregates of Lactuca Sativa. Soil Biol. Biochem. 2010, 42, 429–434. [Google Scholar] [CrossRef]
- Morris, E.K.; Morris, D.J.P.; Vogt, S.; Gleber, S.C.; Bigalke, M.; Wilcke, W.; Rillig, M.C. Visualizing the Dynamics of Soil Aggregation as Affected by Arbuscular Mycorrhizal Fungi. ISME J. 2019, 13, 1639–1646. [Google Scholar] [CrossRef] [PubMed]
- Hussein, K.A.; Joo, J.H. Plant Growth-Promoting Rhizobacteria Improved Salinity Tolerance of Lactuca sativa and Raphanus sativus. J. Microbiol. Biotechnol. 2018, 28, 938–945. [Google Scholar] [CrossRef]
- Parihar, P.; Singh, S.; Singh, R.; Singh, V.P.; Prasad, S.M. Effect of Salinity Stress on Plants and Its Tolerance Strategies: A Review. Environ. Pollut. 2015, 22, 4056–4075. [Google Scholar] [CrossRef]
- Song, Q.; Song, X.; Deng, X.; Luo, J.; Wang, J.; Min, K.; Song, R. Effects of Plant Growth Promoting Rhizobacteria Microbial on the Growth, Rhizosphere Soil Properties, and Bacterial Community of Pinus sylvestris Var. Mongolica Seedlings. Scand. J. For. Res. 2021, 36, 249–262. [Google Scholar] [CrossRef]
- Cabrera-Bosquet, L.; Molero, G.; Stellacci, A.; Bort, J.; Nogués, S.; Araus, J. NDVI as a Potential Tool for Predicting Biomass, Plant Nitrogen Content and Growth in Wheat Genotypes Subjected to Different Water and Nitrogen Conditions. Cereal Res. Commun. 2011, 39, 147–159. [Google Scholar] [CrossRef]
- Ahmad, N.; Javed, A.; Gohar, S.; Ahmed, J.; Sher, A.; Abdullah, M.; Asghar, S.; Javed, K.; Iqbal, J.; Kumar, S.; et al. Estimation of Drought Effects on Different Bread Wheat Genotypes Using Morpho-Physiological Traits. Biochem. Syst. Ecol. 2022, 104, 104483. [Google Scholar] [CrossRef]
- Krupa, M.; Witkowicz, R. Biostimulants as a Response to the Negative Impact of Agricultural Chemicals on Vegetation Indices and Yield of Common Buckwheat (Fagopyrum esculentum Moench). Agriculture 2023, 13, 825. [Google Scholar] [CrossRef]
- Chatterjee, S.; Desai, A.R.; Zhu, J.; Townsend, P.A.; Huang, J. Soil Moisture as an Essential Component for Delineating and Forecasting Agricultural Rather than Meteorological Drought. Remote Sens. Environ. 2022, 269, 112833. [Google Scholar] [CrossRef]
- Bhatt, R.; Kukal, S.S. Soil Moisture Dynamics during Intervening Period in Rice-Wheat Sequence as Affected by Different Tillage Methods at Ludhiana, Punjab, India. Soil Environ. 2015, 34, 82–88. [Google Scholar]
- Li, Q.; Dong, B.; Qiao, Y.; Liu, M.; Zhang, J. Root Growth, Available Soil Water, and Water-Use Efficiency of Winter Wheat under Different Irrigation Regimes Applied at Different Growth Stages in North China. Agric. Water Manag. 2010, 97, 1676–1682. [Google Scholar] [CrossRef]
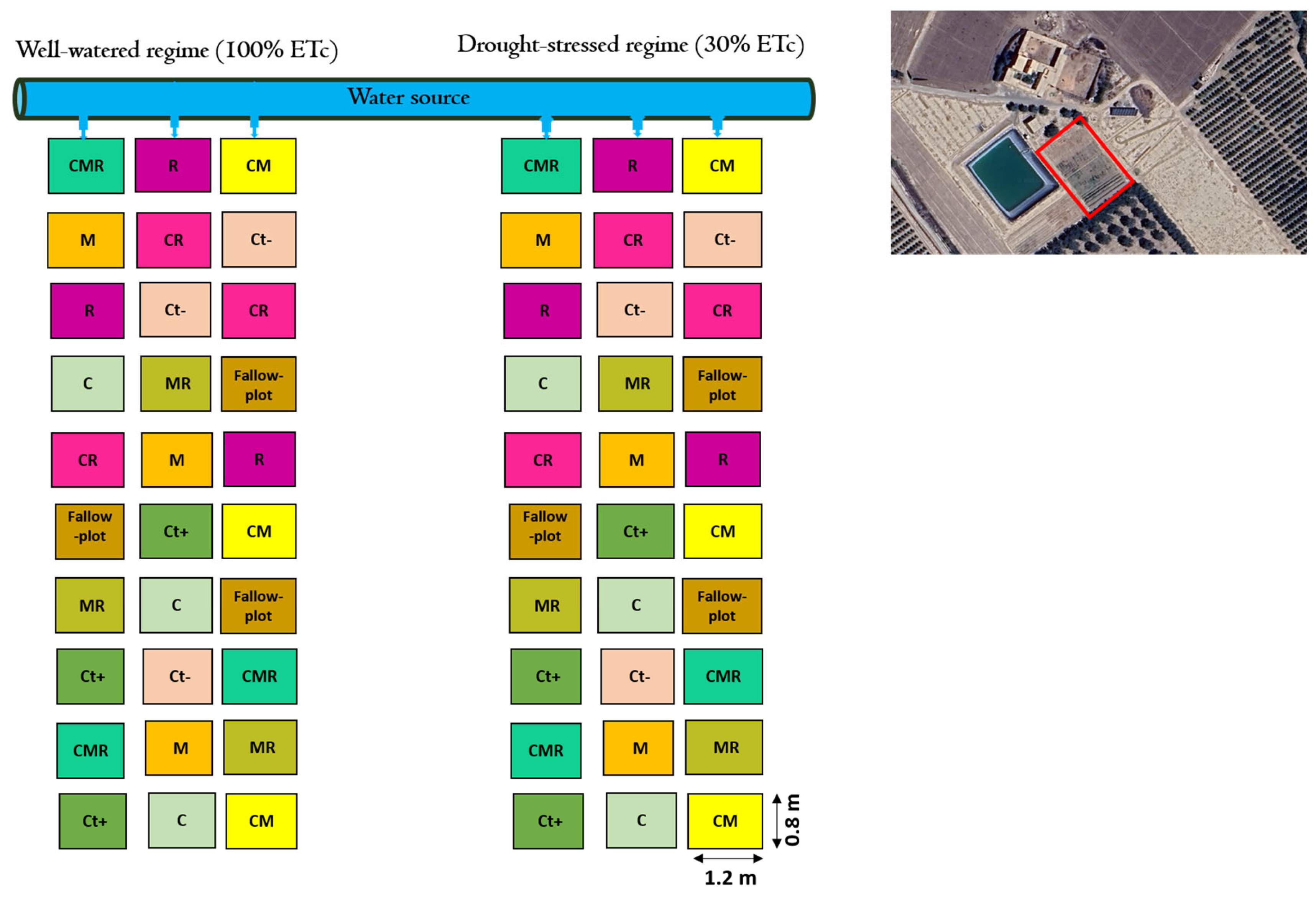
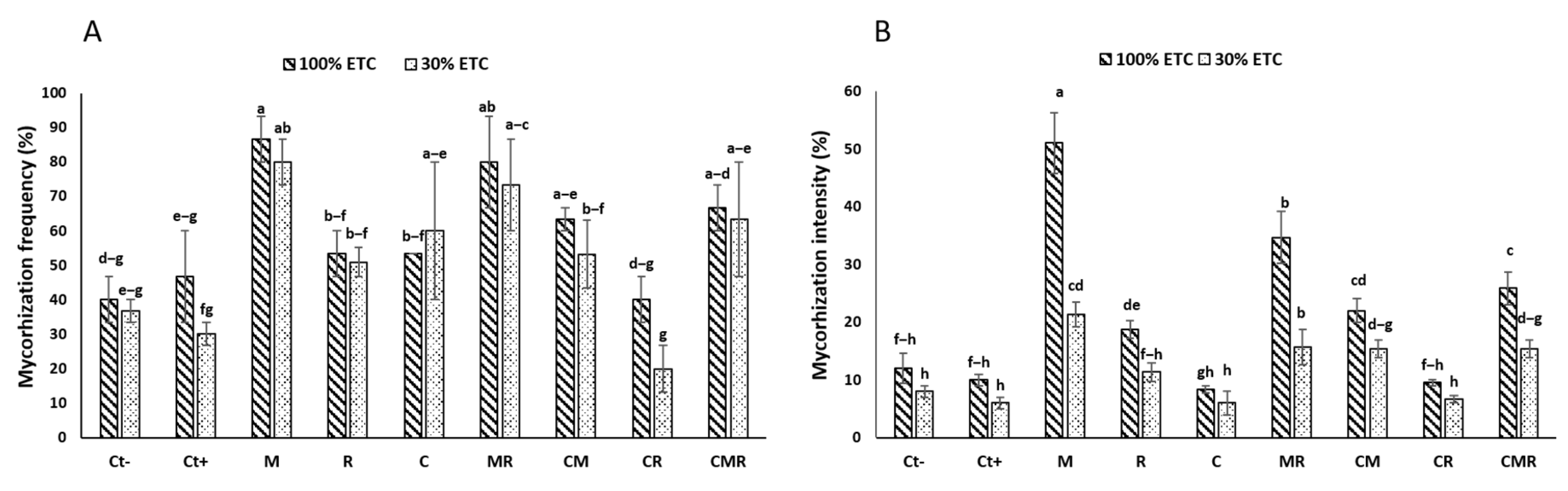
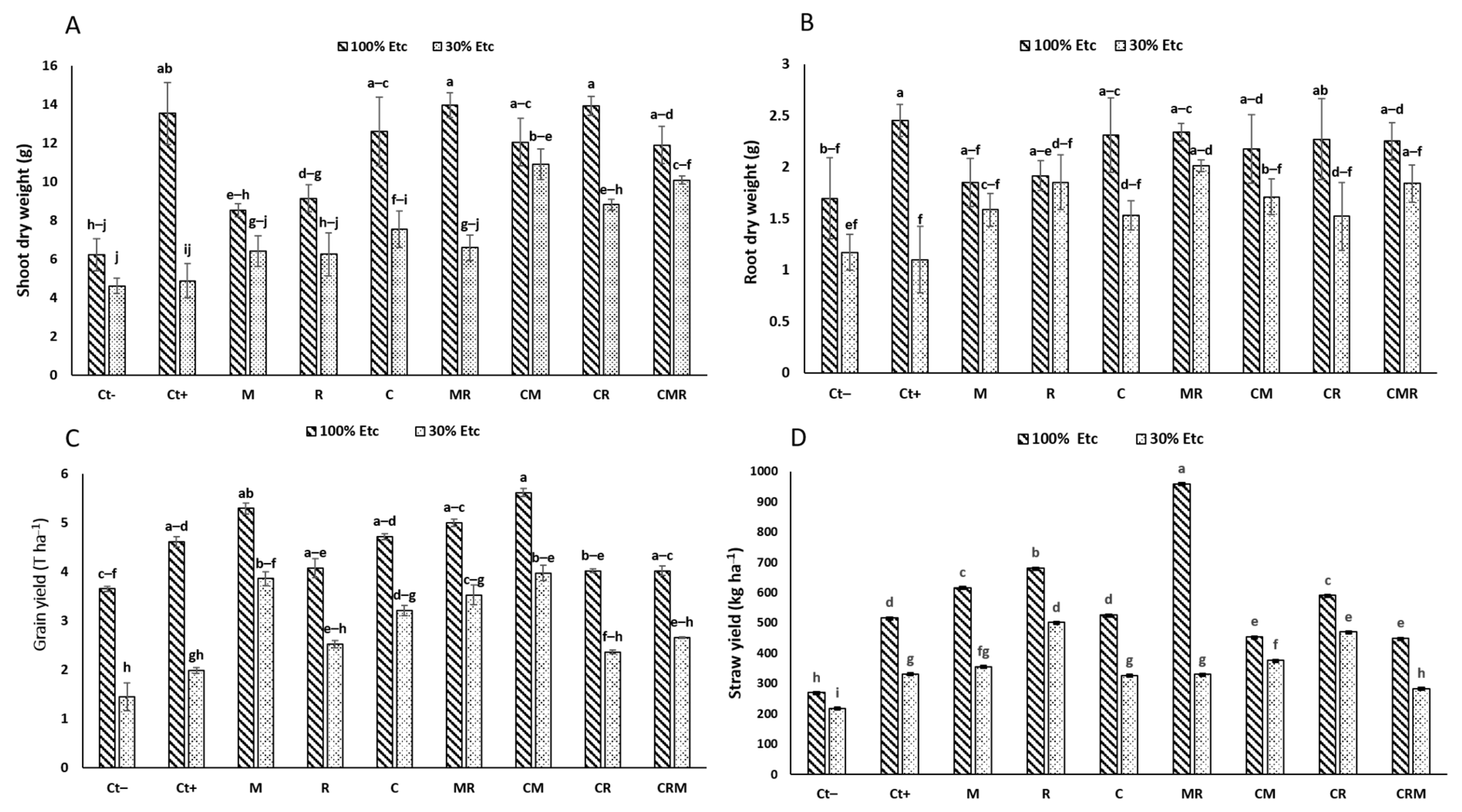
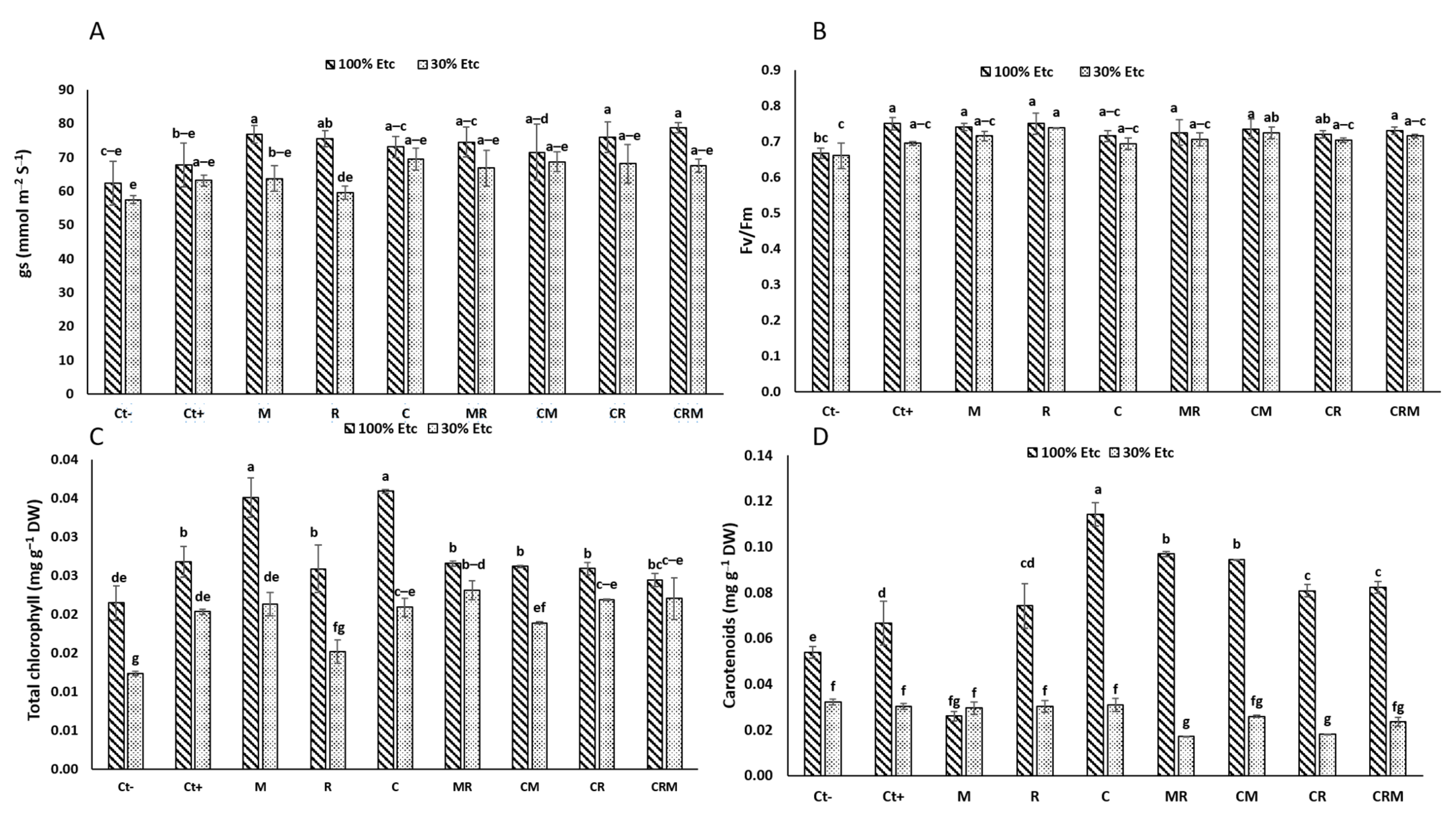
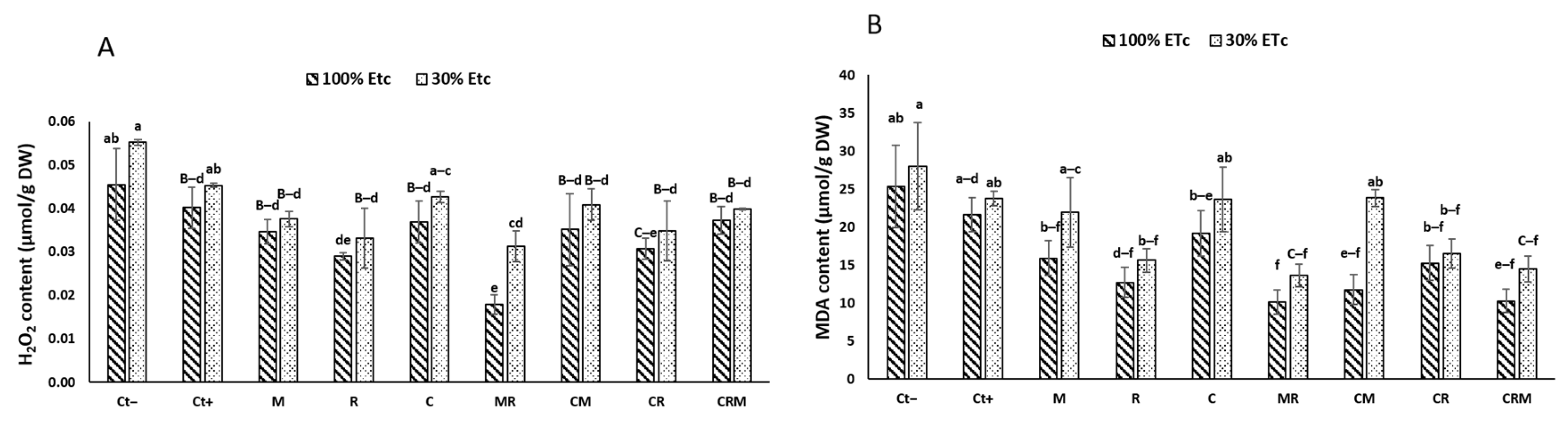
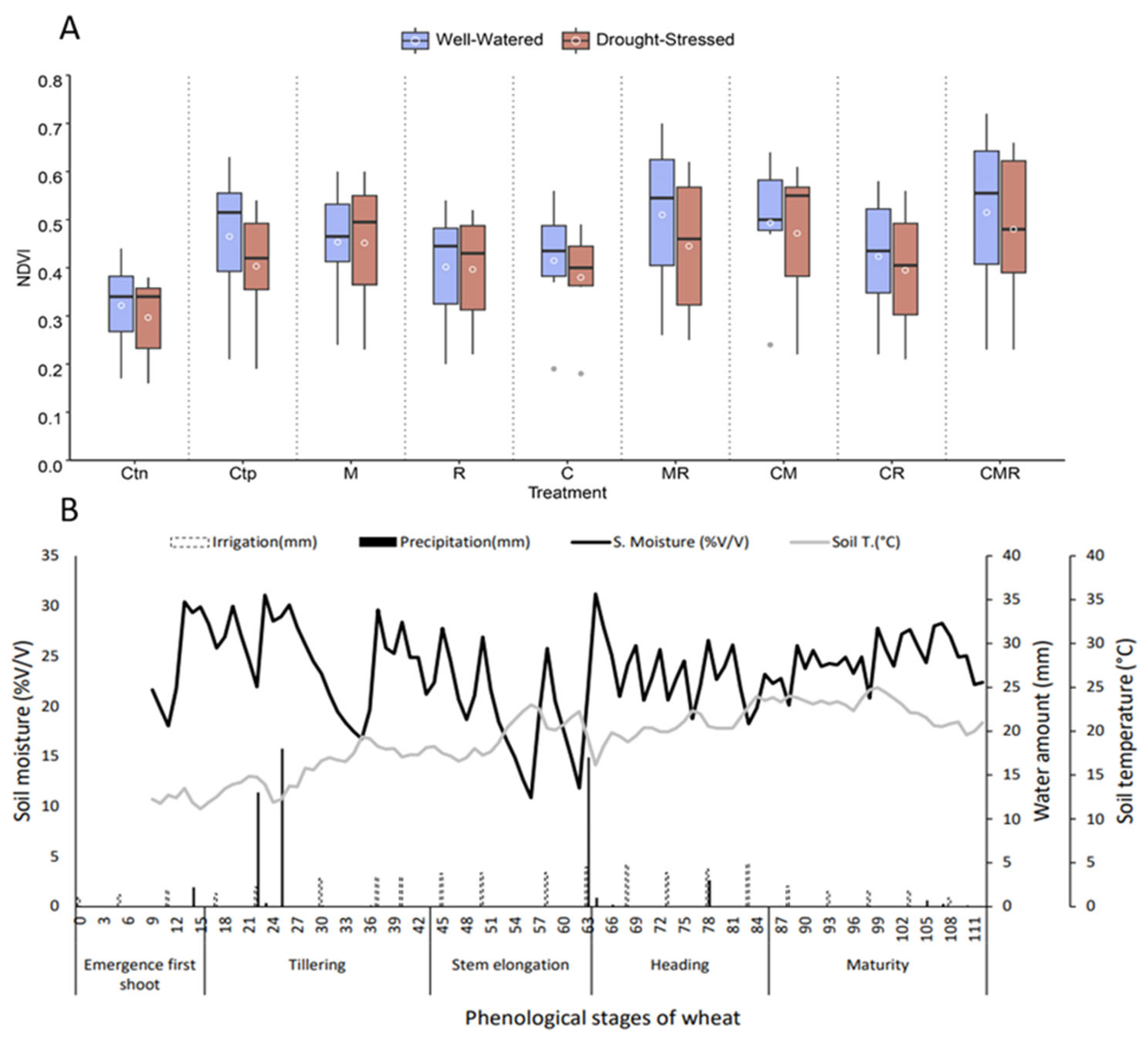
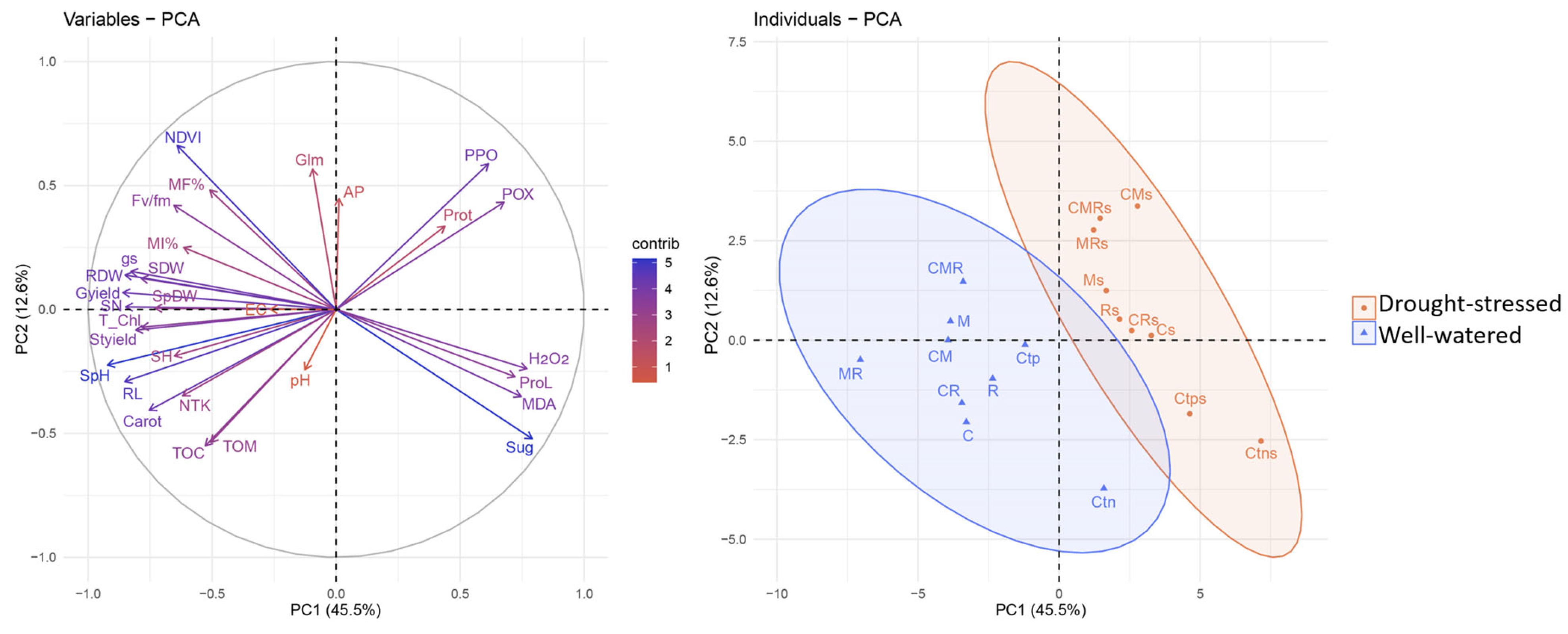
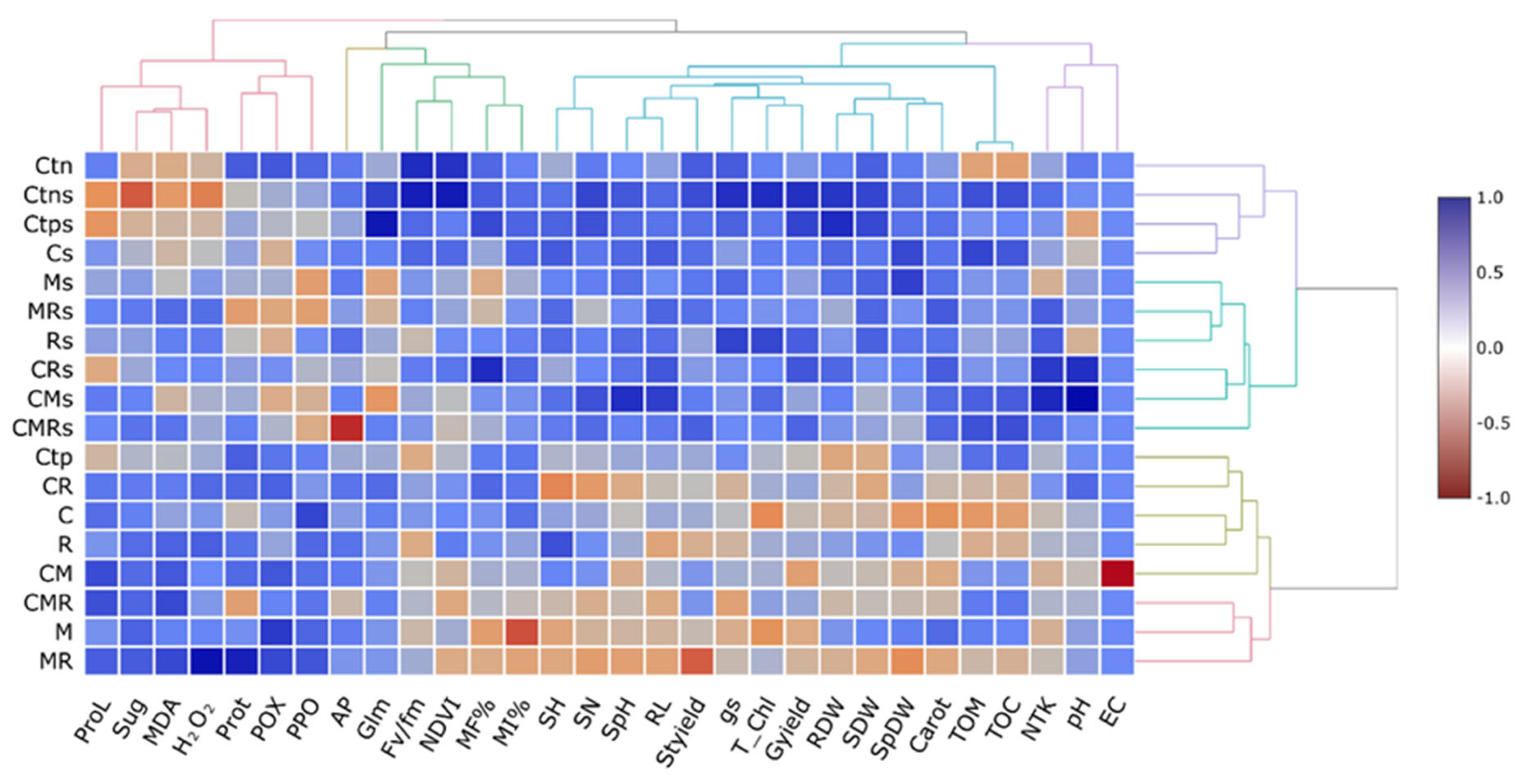
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).