Abstract
Conventional agricultural practices severely deplete the soil of essential organic matter and nutrients, increasing its vulnerability to disease, drought, and flooding. Permaculture is a form of agroecology adopting a whole ecosystem approach to create a set of principles and design frameworks for enriching soil fertility, but there is little scientific evidence of its efficiency. This study compares two permaculture managed sites with a conventional arable site to investigate the effect of permaculture management on soil fertility. We used phospholipid fatty acid analysis to estimate microbial abundance and diversity and related these to measured soil nutrients and carbon stocks. The potential of permaculture management to mitigate soil greenhouse gas emissions was assessed during a laboratory soil incubation and measurement of greenhouse gases via gas chromatography. Overall, the permaculture managed allotments had three times higher microbial biomass, one and a half times higher nitrogen, and four times higher carbon content than the arable site. Permaculture soils had larger carbon dioxide and nitrous oxide fluxes compared to arable soil, but all sites had a mean negative flux in methane. Permaculture management by use of organic amendments and no-dig practices provides a constant slow release of nutrients and build-up of organic matter and carbon and consequently promotes greater bacterial and fungal biomass within the soil.
1. Introduction
Global growing demand for food has, within the last century, led to the commercialisation and industrialisation of agriculture in what is considered conventional farming today [1,2]. Conventional agriculture systems have little crop diversity and through the practice of tillage, erode topsoil at a rate of 1 mm year−1, depleting it of organic matter, nutrients, resilience against disease, and its ability to retain moisture [3,4]. Consequently, a third of the world’s productive arable land has been lost since the 1950s [2]. Crop yields under conventional management are met by the addition of synthetic fertilisers and pesticides to make up for the depleted soil fertility under heavily mechanised practices [2]. In 2022, the United Nations Food and Agriculture Organisation (FAO) claimed by 2050, 90% of the planet’s topsoil will be at risk [5]. Therefore, there is a need to develop alternative methods of cultivating food that enrich soil fertility, rather than depleting it [6].
Alternative farming practices, such as organic farming, aim to enrich soil fertility through using crop rotation and cover crops and reduce the use of agrochemicals [7,8]. However, organic farms tend to vary in the degree to which they adopt these practices, and many still apply organic inputs but within a monoculture system, rather than developing a whole ecosystem design approach [9]. Permaculture, developed in the 1970s, is a set of principles and design frameworks aimed to incorporate the natural relationships and recycling of an ecosystem within a localised and decentralised food system [10]. The principles emphasise the use of multiple methods within one system to best optimise ecosystem functioning specific to the landscape you are growing within, enhancing above and below ground biodiversity [11,12].
There has been an increase in publications surrounding the topic of permaculture over the past two decades (11,700 publications in the period 2013–19); however, only 1900 of these are peer-reviewed scientific studies [13]. The movement of holistic local food systems, of which permaculture is a part of, has originated from grassroots organisations and community projects [14]. There is much global shared knowledge and understanding of agroecological systems, such as permaculture, regenerative agriculture, and agroforestry, disseminated through training courses, manuals, and books [15]. At the same time, an increase in the number of published scientific research and quantitative analyses of these practices would contribute to expanding and justifying this knowledge, supporting a growth and transition towards sustainable and regenerative food systems improving and supporting healthy soils, healthy food, and a healthy planet [12,16].
Microorganisms contribute to 50% of the earths’ biomass and drive carbon and nitrogen cycling within the soil [17]. De Tombeur et al. [16] investigated the potential influence of permaculture management on soil organic matter distribution, alongside physicochemical properties, compared with conventional arable soil and found enhanced breakdown of particulate organic matter, consequently leading to higher organic carbon and nitrogen contents under permaculture management. Soil fertility is traditionally determined by organic matter and nutrient content, but it has become increasingly assessed by microbial abundance due to enhanced and more-accessible methods of analysis [17,18]. A global meta-analysis of 56 studies and 146 pairwise comparisons revealed 32–84% more microbial biomass, carbon, and nitrogen within organic systems compared to conventional [19]. Arbuscular mycorrhizal fungi (AMF) colonise plant roots, creating a secondary root system called mycelium, and support plant nutrient acquisition, and through symbiotic return, the plant provides organic carbon [20,21,22]. AMF root colonisation was quantified across a seven-year crop rotation under organic and bio-dynamic farming systems, compared with conventional systems, and found 30–60% greater AMF colonised root length within the organic and bio-dynamic managed soils [23]. In addition to greater microbial, carbon, and nutrient content within organically practiced food systems, due to reduced tillage, reduced synthetic fertilisers and use of organic fertilisers have shown to reduce GHGs from agriculture [24,25,26,27,28]. Within the IPCC AR6 2023 Synthesis Report for Policymakers, reducing methane (CH4) and nitrous oxide (N2O) emissions and promoting carbon sequestration within agriculture are suggested within the Agriculture, Forestry, and Other Land Use (AFOLU) mitigation options for scaling up climate actions [29]. Therefore, it is important to investigate how emerging agricultural practices, such as permaculture, can contribute to reducing GHG emissions alongside carbon sequestration and nutrient retention.
This study investigates soil fertility under two permaculture managed arable soils, one each under urban and rural settings, and compares them with a conventionally managed arable site. Using the main components of organism’s cell membranes, phospholipid fatty acid (PLFA) analysis can be used as an indication of microbial abundance and diversity [30,31], as a primary indication of soil fertility, alongside physicochemical soil properties. Promotion of nitrogen fixation in crop and forage systems through expanded use of legumes has been advocated as a potential GHG mitigation strategy since lower N2O emissions are expected due to the reduced use of synthetic N fertiliser [32]. To our knowledge, there are no previous studies that have investigated the potential for greenhouse gas emission mitigation alongside the microbial community under permaculture managed arable soils.
This study aims to investigate the potential of permaculture management in creating a localised food system that enriches both rural and urban soils and has the potential to mitigate GHG emissions. This study investigates soil fertility under two permaculture managed arable soils, one each under urban and rural settings, compared with a conventionally managed arable site. We hypothesised that permaculture managed soil will be more fertile than conventionally managed arable soil due to
- Greater microbial abundance, with increase in fungal biomass relative to bacterial.
- Higher soil organic matter and total carbon content from organic amendments and microbial activity.
- Greater nutrient retention.
We have also hypothesised that permaculture managed soil will display lower N2O emissions due to the lack of synthetic N fertiliser use, but higher CO2 emissions due to an increase in microbial abundance, while no difference in methane emissions is expected.
2. Materials and Methods
2.1. Study Sites and Sampling Strategy
The permaculture managed soils were sampled from two allotments located within mid Wales (Figure 1); allotment 1 within the RSPB Ynys-Hir nature reserve accommodates a more rural setting than allotment 2 situated within the town of Machynlleth. Both allotments are interbedded by mudstone and sandstone [33]. The conventionally managed arable site situated in the southwest of England on Fenswood Farm, Long Ashton, is underlined by Mercia mudstone and halite [33].
Allotment 1 has been under permaculture management since 2020, and allotment 2 since 2019. Within permaculture, there is a large emphasis on maximising the edge, being the most diverse area within an environment [10]. The allotments used in this study use a design framework with zones to create varying habitats [12]. Perennial beds are located around the sides of the allotments to resemble a woodland edge, with soil that is rarely disturbed, and shorter perennial trees are located on the south side to avoid shading.
Both allotments use a 4-part cropping rotation system (root crops, nightshade, legumes, and brassicas) to recharge and maintain nutrients within the soil [34]. Using this rotation, no same crop is grown in the same soil each year to reduce receiving a constant level of disturbance. The soil samples taken from both allotments were divided equally over beds cultivating legumes and brassicas.
Additionally, green manures combined with winter vegetables (e.g., black oats, green and field beans) were used as cover crops. The straw, oats, and beans harvested were then applied to hot bins for compost breakdown and inoculated with biochar ready for application in spring. When harvesting, the oat and bean roots were left within the soil to provide food for soil biota and avoid disturbance of the mycelium network [10]. Moreover, both allotments used no-dig raised beds to further minimise disturbance to the microbes and reduce compaction [10]. The soil at both allotments was also amended with organic material of biochar, manure, comfrey, and seaweed (site information sourced from John Williamson, allotment owner, personal comms).
Crop rotation was also used at the Fenswood Farm site; first winter wheat, second winter wheat, and spring oats (per season/year), but also including conventional practices such as ploughing, fertilisation, and mechanised harvesting. Soil samples were collected during the cultivation of first winter wheat, sown in late September, and had not received any fertiliser. However, the previously cultivated spring oats received 110 kg N/ha, 139 kg P/ha, 209 kg K/ha, and 40 kg S/ha in one application (as a uniform granular compound fertiliser containing nitrate and ammoniacal-N, P2O5, K2O, and SO3), 6–7 months before soil samples were collected. The soil was also ploughed and drilled with overwinter mustard before spring oats were planted (Andy Hughes, farm manager, personal comms).
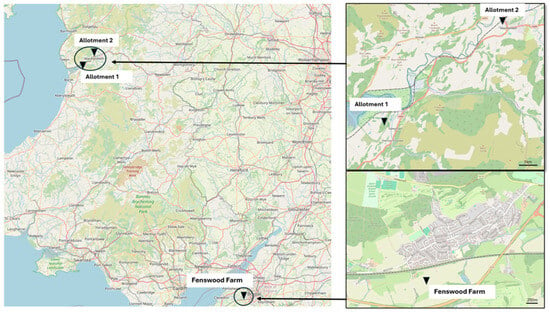
Figure 1.
Location of sample sites. Permaculture management: Allotment 1 (268312, 296,376 BNG) and Allotment 2 (274469, 300849 BNG). Arable conventional management: Fenswood Farm (353396, 169636 BNG). All maps sourced from OpenStreetMap [35].
Figure 1.
Location of sample sites. Permaculture management: Allotment 1 (268312, 296,376 BNG) and Allotment 2 (274469, 300849 BNG). Arable conventional management: Fenswood Farm (353396, 169636 BNG). All maps sourced from OpenStreetMap [35].
Sample collection was carried out over two days due to light limitations and time constraints (21–22 November 2022). Samples from Fenswood Farm and allotment 1 were collected on the first day, and samples from allotment 2 were collected on the second day; all stored in the refrigerator overnight and transported in cool boxes to the laboratory the following day. At Fenswood, ten soil samples were collected from 0 to 10 cm depth, 1 m apart, along a straight line transect. At the allotments, 10 soil samples (0–10 cm depth) were collected from both allotments, halved over two beds (5 soil samples from each bed) using a zigzag transect to account for possible variation within the beds [36]. A further ten soil samples were collected at each site in tin foil and paper bags for PLFA analysis requiring no contamination from plastic [31]. Lastly, soil moisture (%) and soil temperature (°C) were measured in situ using probes.
2.2. Soil Geochemical Properties
Prior to any geochemical analysis, each soil sample was manually homogenised and sieved (<2 mm) at field moisture, removing any large roots and stones. Soil moisture was determined gravimetrically by drying at 105 °C for 24 h and until constant weight. Organic matter content was determined by loss on ignition (LOI) after furnacing 5 g of dried soil at 375 °C for 16 h. Following LOI treatment, the LOI ash (3 g) was used to quantify the absolute particle size distribution of the mineral soil fraction using optical laser diffraction by MS3000 Mastersizer (Malvern Panalytical Ltd, Malvern, UK) [37]. Measurements were repeated for each sample until 5 measurements in a row had a relative standard deviation <5%. Finally, the US Department of Agriculture Soil Textural Calculator was used for soil type identification [38], and it was confirmed that all three sites had the same silty/silty loam soil type. Soil pH was also measured in 2:1 water slurries after 10 min settling time using a pH probe.
Homogenised field-moist soils (5 g) were extracted at a ratio of 5:1 with 25 mL 2 M KCl for the determination of exchangeable ammonium (NH4+), nitrate (NO3−), and phosphate (PO43−). The soil slurries were continuously shaken on a reciprocating shaker at 200 rpm for 1 h before being centrifuged at 5000 rpm for 10 min followed by filtration with 0.45 μm 25 mm PES syringe filters. Ammonium was analysed spectrophotometrically on a Gallery Plus Automated Photometric Analyser (Thermo Fisher Scientific, Waltham, USA) using a salicylate-hypochlorite alkaline reaction method measured at 660 nm, nitrate using a vanadium reaction method measured at 540 nm, and phosphate using the molybdenum blue method measured at 880 nm. The limits of detection were 0.005 mg L−1 NH4+-N and 0.010 mg L−1 NO3−-N and PO43−-P. The samples were blank corrected, while the precision as a relative standard deviation (RSD) was <2%.
The dried soils following the gravimetric soil moisture determination were pulverised with a pestle and mortar (<1 mm), and ~10 mg sub-samples were weighed in tin capsules. These samples were subsequently analysed for elemental C and N contents with an elemental analyser (EA) (vario PYRO cube; Elementar Analysensysteme GmbH, Hanau, Germany). The EA was calibrated with sulfanilamide (N: 16.26%, C: 41.81%, S: 18.62%) and the precision RSD was <5% for both C and N.
2.3. Phospholipid Fatty Acid (PLFA) Analysis
PLFAs were extracted and isolated using the method adapted from Buyer and Sasser [39] and Joergensen [31] to quantify microbial biomass in the soils. All glassware was furnaced (450 °C 4 h), and all solvents used were HPLC grade (Rathburn, UK).
PLFAs were extracted using Bligh–Dyer solution prepared by mixing 100 mL buffered water (0.05 M KH2PO4 adjusted to pH 7.2), 125 mL chloroform, and 250 mL methanol (MeOH). Lipids were extracted from soil (1 g) using Bligh–Dyer solvent (3 mL) and sonicated for 15 min. The extracts were centrifuged at 3000 rpm for 5 min, and the supernatant was transferred. This was repeated three times. Subsequently, 2 mL of buffered water and 2 mL of chloroform were added, mixed, and centrifuged at 3000 rpm for 5 min to separate the aqueous and organic phases. The organic layer was transferred to a new vial, and the aqueous layer was washed three times with 1 mL chloroform. The total lipid extract (TLE) was dried at 40 °C under N2.
Before lipid fractionation, activated silica columns were conditioned with 4 mL chloroform; then, TLE was added in chloroform (1 mL). The simple lipids were eluted in 5 mL of chloroform. The glycolipid fraction was eluted afterwards by adding 20 mL of acetone. The phospholipid fraction (PL) was eluted last by adding 5 mL of MeOH, which was taken forward for analysis. The internal standard (10 µL nonadecane, 0.1 mg mL−1) was then added to the PL fraction, then dried under N2 at 40 °C.
The PL fraction was methylated using 3 mL of 5% anhydrous HCl in MeOH, heated at 50 °C for 2 h. After 10 min cooling, 2 mL of saturated NaCl solution and 1 mL of hexane were added and mixed. The organic phase was transferred, and this was repeated three times. The derivatised PL fraction was dried under N2 at 40 °C.
Finally, 50 µL of hexane was added for analysis by gas chromatography—mass spectrometry (GC-MS). The GC was fitted with a VF23-ms column (60 m, 0.32 µm i.d., 0.15 µm film thickness), and the temperature programme was 50 °C (1 min) to 100 °C (10 °C min−1) to 250 °C (4 °C min−1, 15 min hold) with a helium carrier gas flow of 2.0 mL min−1, and the MS under electron ionisation (70 eV) had a full scan range (m/z 15–650) and a scan time of 0.2 s.
Data were acquired and analysed using Xcalibur (version 4.1). The individual fatty acid methyl esters between chain lengths C14 and C20 were identified using characteristic mass spectra and compared to available in-house standards or reported in the literature. The concentration (µg−1) of each fatty acid was determined in comparison to the internal standard by integration. Assignments of PLFAs were adapted from Joergensen [31]. Firmicute-derived PLFAs were i14:0, i15:0, i16:0, i17:0, i18, a15:0, a16:0, a17:0, a18:0, and a19:0, where i indicates iso-branching, and a indicates anteiso-branching. Actinobacteria-derived PLFAs were 10Me16:0, 10Me17:0, and 10Me18:0, where 10Me indicates methyl substitution on C10 [40]. The sum of firmicutes and actinobacteria was used to represent Gram-positive bacteria. The sum of cy17:0, cy19:0, 16:1ω7, 16:1ω9, 17:1ω8, and 18:1ω7 was used to represent Gram-negative bacteria, where cy indicates cyclopropane fatty acids, and ω indicates the position of the double bond. The sum of 16:1ω5c, 18:1ω9c, 18:2ω6c (c indicates cis), and 18:3ω6,9,12 was used to represent AMF, Zygomycota, Ascomycota, Basidiomycota, and unspecified fungi, the sum of which was total fungal PLFAs [41]. Other fatty acids 14:0, 15:0, 16:0, 17:0, 18:0, 20:0, and 20:4ω6,9,12,15 were classed as unspecified microbial PLFAs, as they occur in both bacteria and fungi [42].
2.4. Greenhouse Gas Incubation
After homogenisation, 10 g of field-moist soil was weighed into 100 mL acid-washed serum bottles. The bottles were stoppered with butyl rubber septa and were incubated in the dark at 20 °C for 24 h. Gas samples (5 mL) were collected from the bottle headspace, via syringe and needle through the septa, at times 0, 1, 3, 6, 12, and 24 h, and the gas samples were transferred into pre-evacuated 3.5 mL borosilicate exetainer vials (Labco, Ceredigion, UK). After each sampling, the sampled headspace gas was replaced with atmospheric air to maintain atmospheric pressure throughout the incubation period. At the end of the incubation, the exetainer vials were loaded on a PAL3 autosampler mounted on top of an Agilent 7890A gas chromatograph (Agilent Technologies Ltd., Santa Clara CA, USA) equipped with μECD and FID detectors, and 1 mL of gas was sub-sampled and analysed for N2O, CO2, and CH4, respectively. Greenhouse gas flux rates were determined by linear regression between 0 and 24 h when the linear regression r2 was >0.9 and the fluxes were above the minimum detectable concentration difference (MDCD) for each gas. The instrument precision, at atmospheric concentrations for the 3 gases, was determined from repeated analyses of 8 lab air samples, and the relative standard deviation was <5% for all of them, while the minimum detectable concentration difference (MDCD) was 9 ppb N2O, 72 ppb CH4, and 31 ppm CO2, respectively.
2.5. Statistical Analyses
Before statistical analysis, raw data of all parameters were tested for normal distribution using the Shapiro–Wilk test, and data not normally distributed were log-transformed to allow use of parametric tests. One-way ANOVA (analysis of variance) was used to test whether soil properties statistically varied between permaculture and conventional management (p < 0.05), and Tukey post hoc tests (HSD) were used to assess statistical pairwise difference between sites (p < 0.01). Multiple and simple linear regression models were used to assess potential causality between soil nutrients, organic matter, microbial abundance, and soil gas fluxes (p < 0.01). Durbin–Watson test tested for spatial autocorrelation within the regression models, identifying no autocorrelation within the model if p-value was high and the test statistic was between 1.5 and 2.5. Pearson correlation tests assessed relationships between soil organic matter content, soil nutrients, soil greenhouse gas fluxes, and microbial abundance. All statistical analyses and graphs were produced using R Studio software (version RStudio 2023.12.1 +402).
3. Results
3.1. Soil Geochemical Properties
The soil geochemical properties are shown in Table 1. The moisture content of the allotment soils was significantly higher than the arable soil in Fenswood Farm, and their pH was lower compared to the more alkaline Fenswood soil (ANOVA, both p < 0.001). The allotment soils had significantly higher nitrogen, phosphorus, and carbon contents than the conventional arable soil, with the more rural allotment 1 (C/N = 12.4) displaying higher nitrogen content than the urban allotment 2 (C/N = 17.6), which was richer in carbon content.

Table 1.
Soil geochemical properties in the two permaculture managed soils and the conventional arable soils. Data are means ± standard error (SE). Similar capital letters indicate no significant difference in the mean according to ANOVA (p < 0.05). The USDA Soil Textural Calculator was used to classify soil type.
3.2. Microbial Abundance
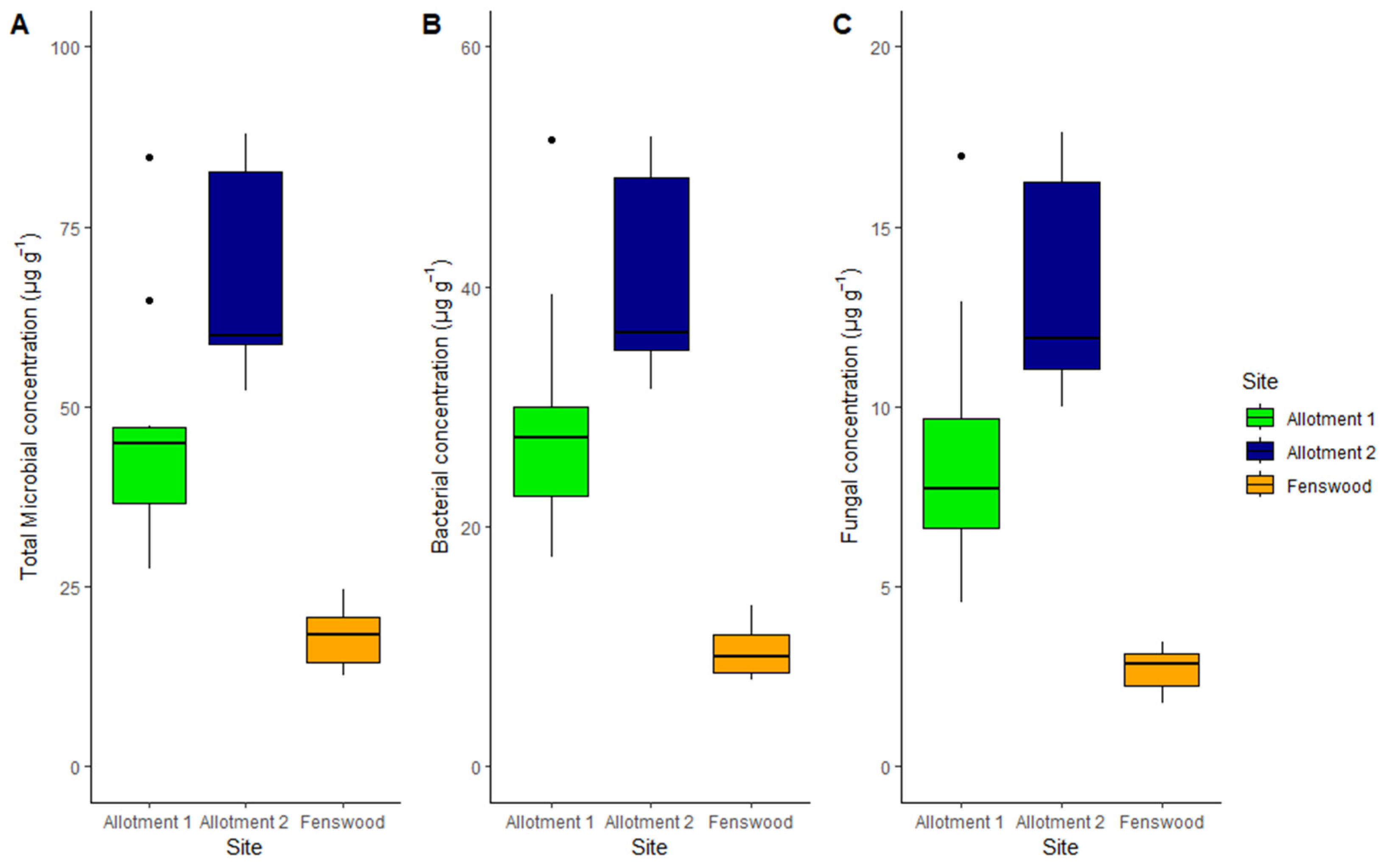
Total microbial abundance varies significantly under permaculture and conventional management (ANOVA, p < 0.001) (see Figure 2). Both allotments 1 and 2 have a much greater total microbial abundance than the conventionally managed soil (HSD, 28.68 µg g−1 and 50.24 µg g−1 difference, respectively, p <0.001). However, the non-statistically significant difference in microbial abundance between the allotments reiterates the effect of permaculture management on soil microbial biomass.
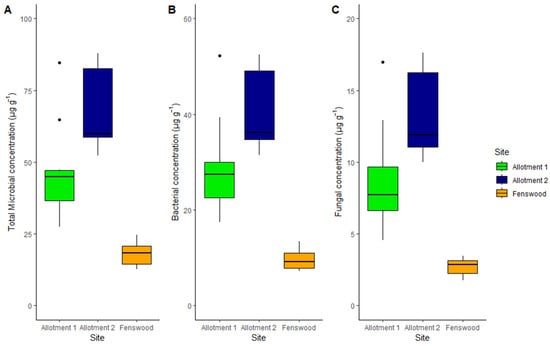
Figure 2.
Median and interquartile range of total microbial (A), bacterial (B), and fungal (C) concentration by site. Fungal concentration varies between all three sites (ABC), but bacterial and total microbial concentrations do not statistically vary between the permaculture allotments (AAB).
For all three sites, bacterial abundance contributes more to the overall total microbial abundance (59%, mean of all soil samples) than fungal biomass (18%, mean of all soil samples) (see Figure 2B,C). The pairwise differences in allotment 1 and allotment 2 against Fenswood for bacterial biomass are 19.38 µg g−1 and 31.05 µg g−1, respectively, (p < 0.001) and 6.01 µg g−1 and 10.57 µg g−1, respectively, (p < 0.01) for fungal biomass. Therefore, both bacterial and fungal abundance are significantly greater under permaculture managed soils compared to conventionally managed soils. However, fungal abundance does statistically differ between the permaculture practiced allotment soils (p < 0.01), implying fungi varies more than bacteria between the sites.
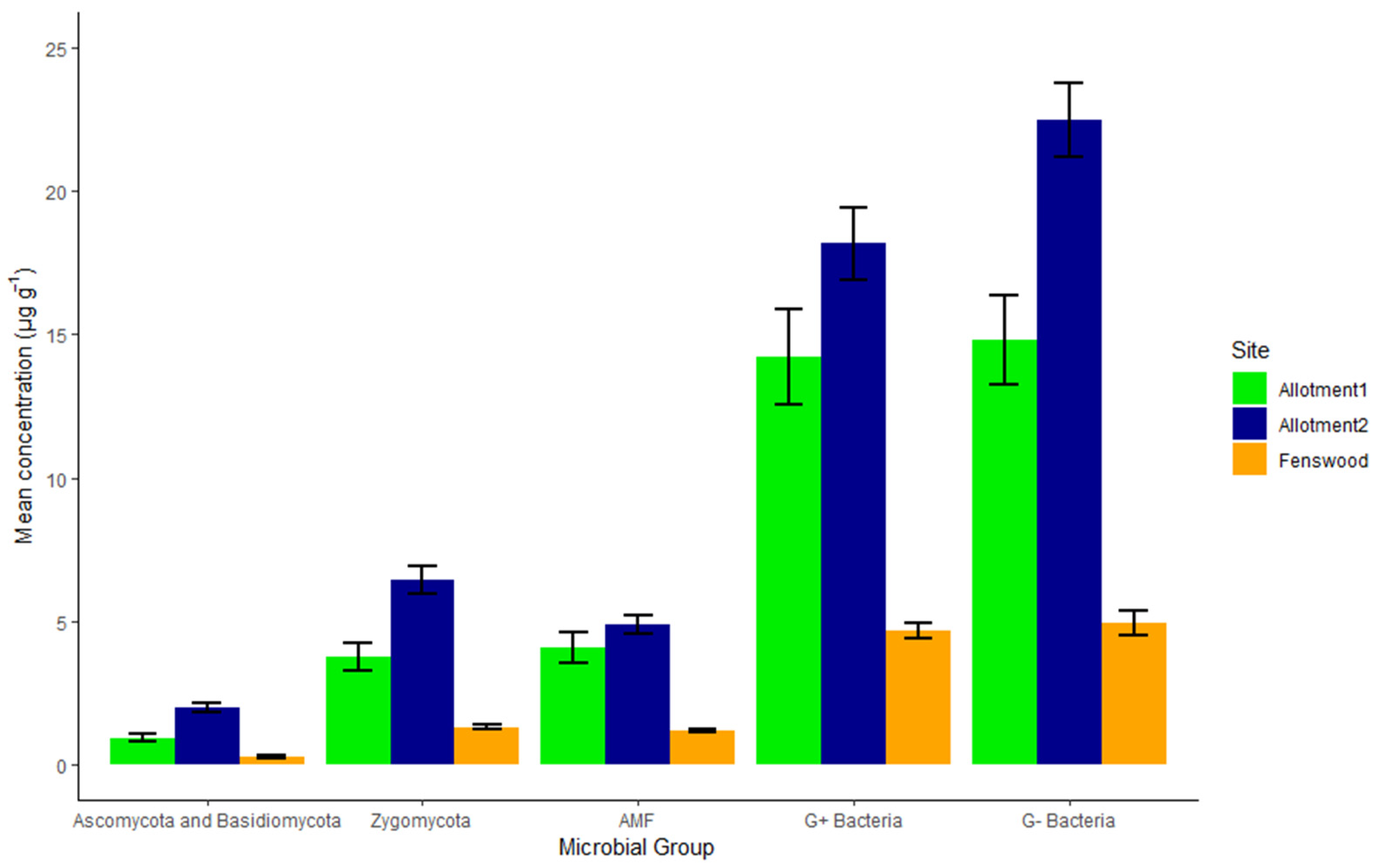
As shown in Figure 3, a clear pattern is seen for the distribution of all PLFA microbial groups between sites: allotment 2 has the greatest concentration and Fenswood has the lowest. AMF and G− bacteria exhibit no significant difference between the permaculture sites, indicating a similar abundance of these fungi and bacteria within both permaculture managed allotments. Allotments 1 and 2 have 2.9 µg g−1 and 3.69 µg g−1 (HSD, p < 0.001) higher G− bacteria concentration than Fenswood, respectively.
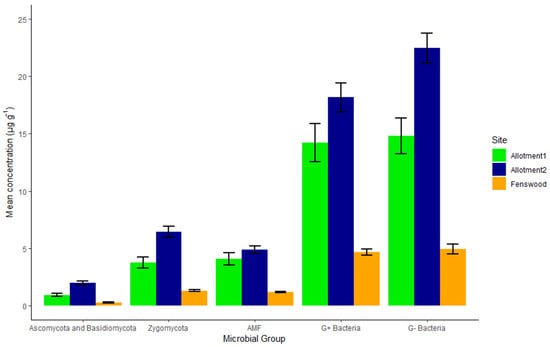
Figure 3.
Mean concentration and standard error (n = 10) of identified PLFA microbial groups within the soil by site. The permaculture managed allotments have a greater mean concentration for all PLFA microbial groups compared to conventionally managed Fenswood soils. G- bacteria and AMF do not statistically vary between the allotments themselves (AAB), in comparison to Zygomycota, Ascomycota, and Basidiomycota, and G+ Bacteria varying between all three sites (ABC) (HSD, p < 0.05).
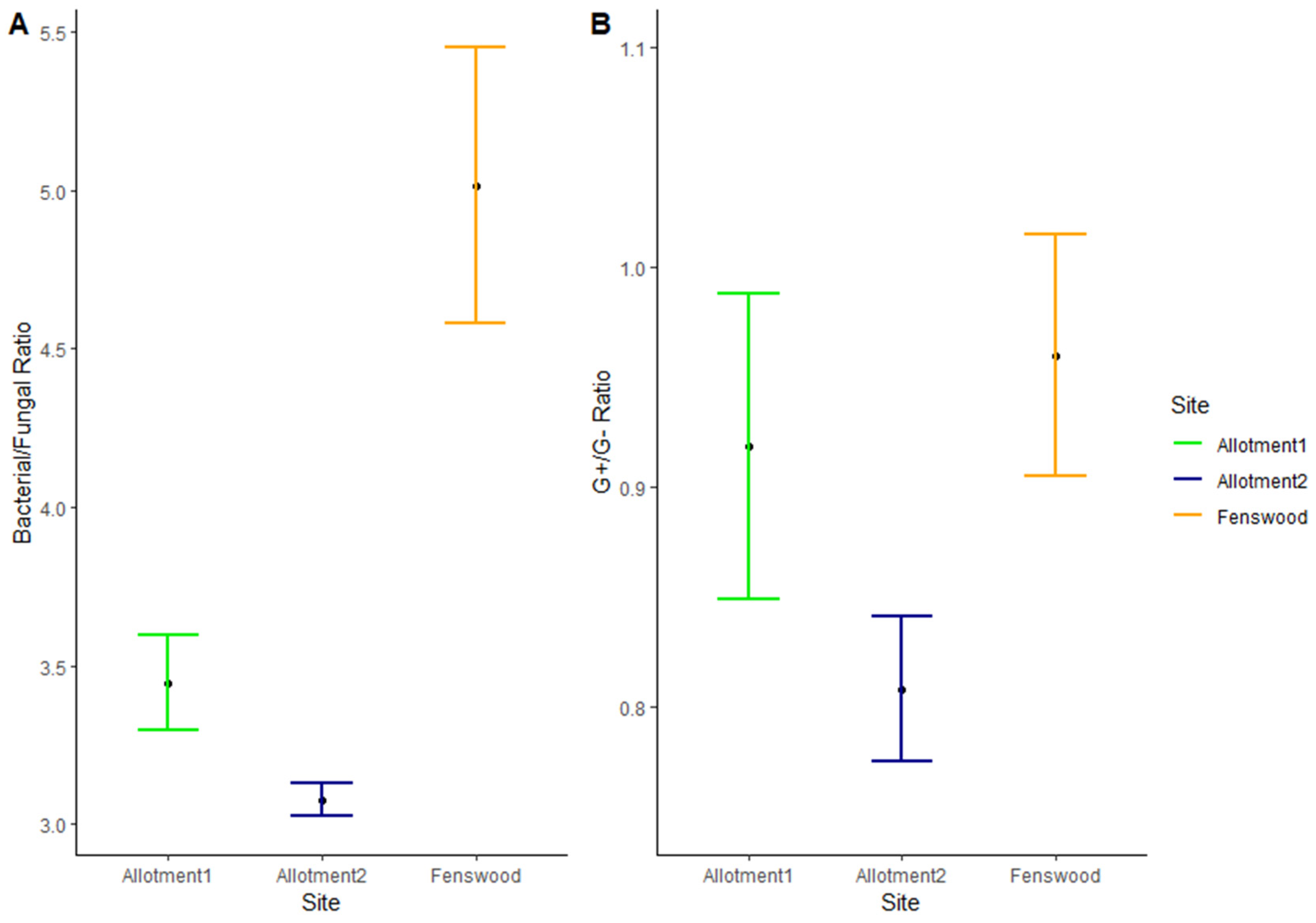
Furthermore, the bacterial/fungal (B/F) ratio significantly varies between sites (ANOVA, p < 0.01), with both permaculture managed allotments 1 and 2 having a much lower B/F mean ratio compared to Fenswood (HSD, 1.57 and 1.94, respectively, p < 0.05) (see Figure 4A). This suggests a larger abundance of both bacteria and fungi within the permaculture managed soils, whilst Fenswood has a much lower abundance of fungi compared to its bacterial biomass.
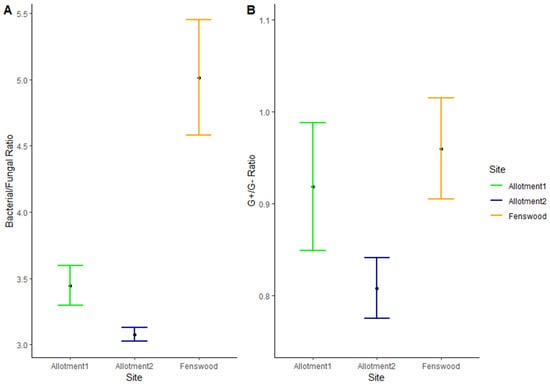
Figure 4.
Mean and standard error (n = 10) of (A) bacterial/fungal ratio and (B) G+/G− bacteria ratio. B/F ratio does not statistically vary between allotments (HSD = AAB). G+/G− ratio does not statistically vary between all three sites (HSD = AAA).
A slightly different pattern is shown in Figure 4B for the G+/G− bacteria ratio per site, as there is no statistically significant pairwise difference between the sites, which may be caused by the large standard errors. Despite this, the ratios show allotment 2 has a high abundance of both G+ and G− bacterial PLFA indicated by its much lower G+/G− ratio, and overall, the permaculture managed allotments have a greater abundance of G− bacteria than the conventionally managed Fenswood site.
3.3. Greenhouse Gas Fluxes
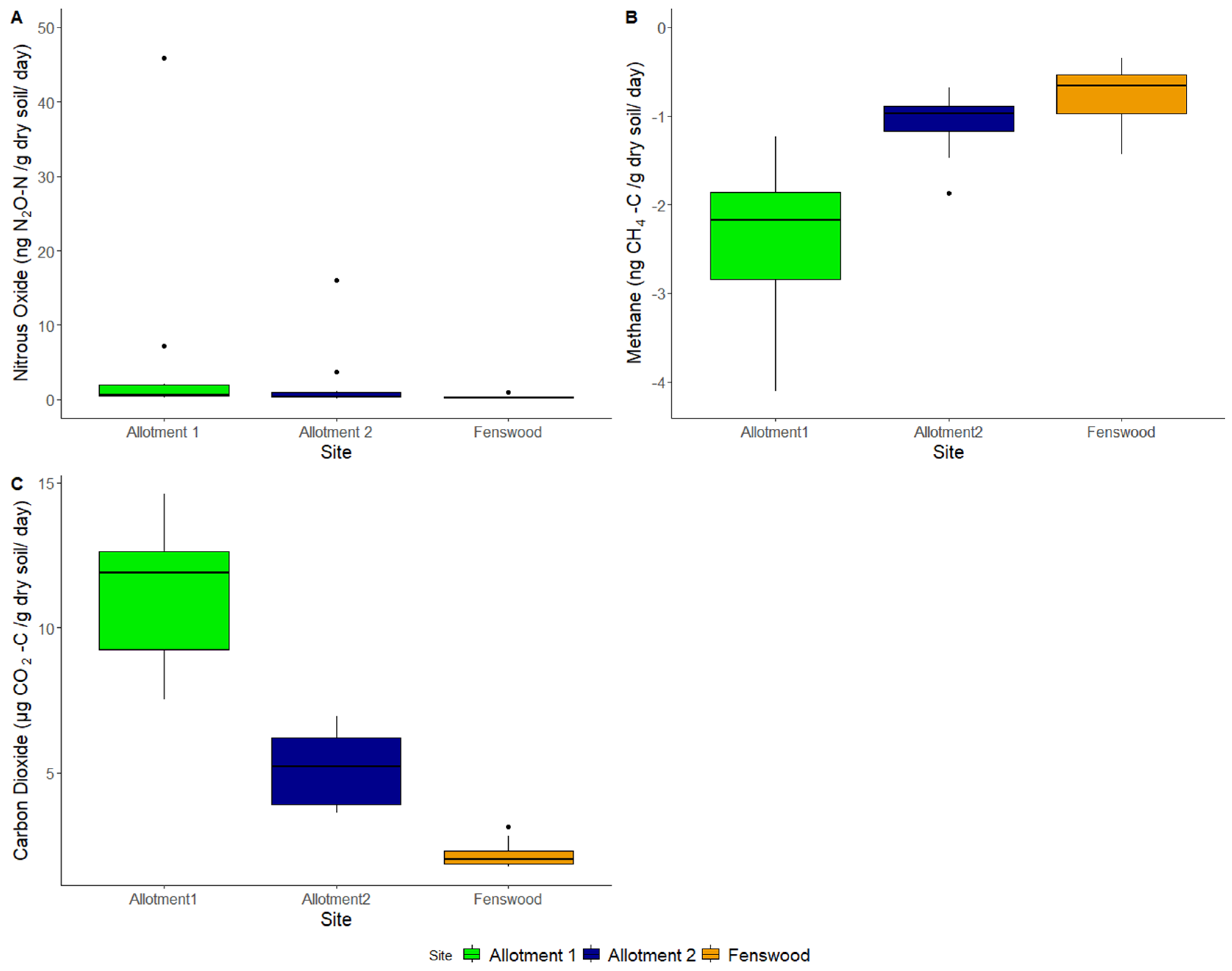
CO2 significantly varies between sites (ANOVA, p < 0.001), but CH4 and N2O do not (ANOVA, p > 0.05). The allotment soils have larger CO2 and N2O fluxes compared to Fenswood soil (Figure 5), but all sites have a mean negative flux in CH4, which interestingly is also larger for the permaculture soils, indicating a stronger sink potential. However, due to the high variability between samples in each site, due in part to the outliers shown in Figure 5, the N2O and CH4 fluxes were not significant.
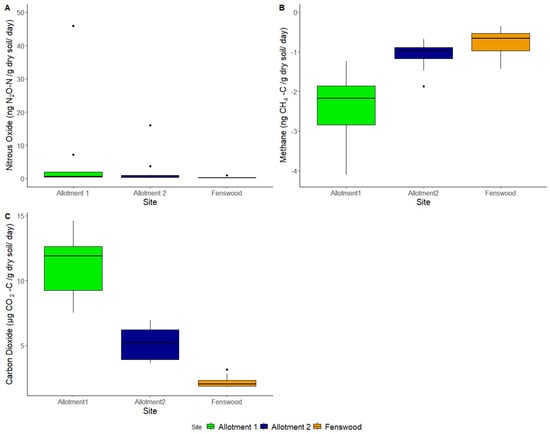
Figure 5.
Greenhouse gas flux of soils per site: (A) nitrous oxide (ng/g dry soil/day), (B) methane (ng/g dry soil/day), and (C) carbon dioxide (µg/g dry soil/day).
3.4. Controlling Factors
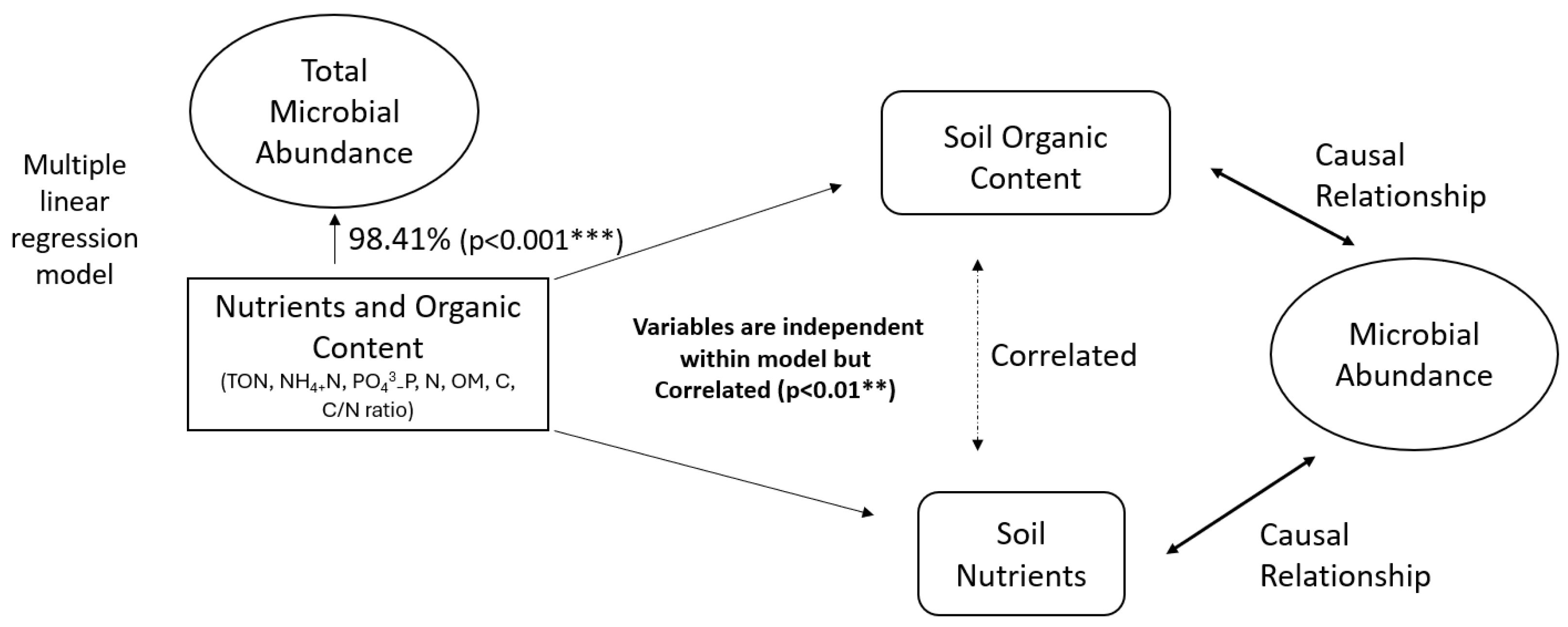
A multiple linear regression model investigated the potential control of soil nutrient availability and organic content on total microbial biomass. Soil nutrients and organic content (representative independent variables: total oxidised nitrogen, phosphate (PO43−), ammonium (NH4+), total nitrogen, organic matter, total carbon, and carbon/nitrogen ratio) have a significant 98.41% (r2, p < 0.001) influence on total microbial biomass. One of the assumptions of a linear regression model is that the residuals are independent. A Durbin–Watson test proves the model has no autocorrelation, but there are large positive Pearson correlations between organic matter and soil inorganic nutrients (ammonium, nitrate, and phosphate) (0.95, 0.96, 0.75, respectively, p < 0.01). Therefore, indicating that soil organic content and nutrients do have a relationship, but one that is not necessarily causal.
To further analyse the effect of organic amendments on microbial biomass and their subsequent effect on nutrient retention and organic matter and carbon breakdown under permaculture management, simple linear regression analysis of total microbial abundance with soil organic content, carbon, and nutrients was undertaken. The results show a strong relationship between total microbial biomass and organic and nutrient accumulation, with total microbial abundance explaining 64.81% variance in total carbon, 44.89% variance in organic matter, 43.37% variance in nitrogen percentage, and 93.04% variance in phosphate (p < 0.001). The soil physical properties (soil moisture and soil temperature) do not have a strong relationship with total soil microbial biomass with Pearson correlation tests of 0.5 (p < 0.01) and −0.2 (p < 0.01), respectively, which may be attributed to the varying climates and locations of the sampling sites.
To explore the control of soil nutrient, organic, and total carbon content and bacterial and fungal biomass on soil gas fluxes, three multiple regression models containing the same algorithm of independent variables were assessed to explain variation in soil nitrous oxide, methane, and carbon dioxide flux, as shown in Table 2. The multiple regression models account for a similar significant explanation of variance for CH4 and CO2 flux but explain 15.52–23.54% greater variance in N2O flux. Ammonium is the greatest determinant for CH4 and N2O flux, whilst total carbon is the greatest determinant for CO2 flux, followed by bacterial biomass. Soil microbial abundance does not have as strong of an influence on the gas fluxes as compared to nutrients and carbon content but does account for some variance in CH4 and CO2 flux (14.57% and 16.41%, p < 0.1, respectively, assessed by simple linear regression). The measured soil physical properties, soil moisture and soil temperature, do not show a strong relationship with soil greenhouse gas fluxes, except for soil moisture’s positive Pearson correlation with CO2 flux (0.7, p < 0.01).
Table 2.
Multiple regression model of all three greenhouse gases explained by soil properties: the independent variables inputted into each model, the result, and a test of autocorrelation between the residuals. Influence expressed as % of variance (R2 value) (p < 0.001 ***).
4. Discussion
Permaculture management enriches soil fertility as shown by the significantly more nutrient and carbon-rich and microbiologically abundant soil samples collected from the permaculture practiced allotments compared to the soil samples from the conventionally practiced farm. The presence of AMF and G− bacteria within soils is crucial for exchanges of nutrients and minerals between the plant and microbes [20]. The permaculture managed allotments 1 and 2 had greater AMF (HSD, 2.90 µg g−1 and 3.69 µg g−1 difference, respectively) and G− bacteria (HSD, 9.52 µg g−1 and 13.5 µg g−1 difference, respectively) abundance, and a lower bacterial-to-fungal ratio compared to the conventionally managed Fenswood soil. These findings suggest a greater community and thus network of microbes within the permaculture soils, whereas conventional practices, such as tillage and the use of pesticides/fungicides, have reduced fungal abundance in the arable soil. Jiang et al. [43] also found greater AMF biomass (46%) within soils under organic fertiliser application compared to synthetic fertiliser only, within a meta-analysis of 162 field studies, and Oehl et al. [44] found the number of AMF in soil under bio-organic and bio-dynamic practices was greater than that of the conventionally practiced site (12.5, 14, and 10 AMF spores g−1 soil, respectively) in a long-term field study in Central Europe.
In addition, the greater AMF and total microbial biomass within the permaculture soils of this study correspond with a higher PO43− concentration, and total microbial abundance explains a significant 93.04% (p < 0.01) variance in PO43− concentration. PO43− is important for the exchange of minerals between the microbes and the plant, with certain microbes able to solubilise phosphorus [20]. Edlinger et al. [45] used a greenhouse experiment to test the acquisition of phosphorous by AMF and found a decrease in AMF within croplands caused a 43% decrease in phosphorus uptake. This may further explain why the conventional managed arable site has less PO43− in correspondence to its much lower bacterial and fungal biomass.
Furthermore, the greater abundance of G− bacteria within the permaculture soils corresponds with a higher total N, NH4+, and total oxidised N concentration compared to Fenswood soils. The use of crop residue after harvesting adds organic material, nutrients, and structure to the soil and thus promotes the mineralisation of organic nitrogen and subsequent nitrification of ammoniacal nitrogen to available nitrate and nitrite [21,46]. Rhizobia G− bacteria living within root nodules of legumes fix atmospheric nitrogen converting it to ammonium readily available for plant uptake [21,47]. Lazcano et al. [48] state how application of unprocessed manure, compost, biochar, and vermicompost can enhance the abundance of nitrifying and denitrifying microorganisms due to the high nitrogen and carbon content within these organic fertilisers. Therefore, the addition of organic material, mulch, and crop residue, under permaculture practices, promotes both a greater microbial biomass and nitrogen availability. This results in greater microbial activity, including G- bacteria, within the allotments, mineralising more of the available organic nitrogen for plant uptake.
These bacteria work alongside other microbes within the soil to enhance soil fertility [20]. Hestrin et al. [49] tested the potential synergistic relationship between AMF and other microbial communities with mycorrhizal plants to increase total nitrogen acquisition. They found soils treated with microbial biomass (by adding soil from perennial switchgrass to the organic matter, measured by PLFA analysis) inoculated with AMF acquired 18% of nitrogen from organic matter, double what the soil with just AMF acquisitioned. Their results further suggest a relationship between AMF, microbes, and the plant, acquiring nitrogen for the plant productivity. However, the present study represents a single snapshot of the soil’s conditions under permaculture and conventional arable practices during autumn 2022. Fenswood Farm applied 110 kg N/ha during the previous spring 2022; therefore, the soil N content may have been quite different if sampled in spring instead of autumn. Episodic fertiliser application often results in spikes of soil N, compared to a slow release of inorganic N from the use of organic amendments used within permaculture management [10,32]. This emphasises the need to increase temporal studies of permaculture and soil nutrient fluctuations. Nevertheless, despite the sampling time, the combination of these slowly induced amendments with no-dig practices enables soil microbial communities to develop and aid soil nutrient retention. By comparison, conventional management practices in Fenswood farm such asbiomass removal, tillage, and herbicide applications, disrupt the soil microbial network and inhibit the soils’ ability to retain nutrients [10]. This observation is strongly corroborated by simple linear regression results showing microbial abundance has a significant relationship with the accumulation of nitrogen, organic matter, and soil carbon (43.37%, 44.89%, and 64.81% variance, respectively, p < 0.001).
Interestingly, there is also an observed difference in G− bacteria abundance between the permaculture allotments themselves. Allotment 2 had a greater abundance of G− bacteria than allotment 1 (HSD, 7.69 µg g−1, p < 0.01), but allotment 1 had higher total nitrogen. Studies suggest the potential of biochar to enhance NH4+ and NO3- retention and overall nitrogen acquisition within soils [50,51,52,53,54,55]. Thus, despite these studies not investigating the effect of biochar use within permaculture managed soils, it could be one explanation for the higher total nitrogen found in allotment 1 having received more biochar addition compared to allotment 2. In addition to biochar application, there may be other factors that affect nitrogen cycling, such as types of crops planted and their respective C/N ratio.
Many of the indicators used within this analysis are not specific to one microbial group, and dormant microbes can recycle their PLFA cell membranes, which has the potential to lower the specificity of the PLFA indicator [31]. However, Romaniuk et al. [56] compared PLFA analysis with Catabolic Response Profiles (CRPs) and found similar sensitivity indices between both methods. Therefore, it is likely that another method of microbial analysis would have also revealed permaculture soils to have higher microbial abundance and thus come to the same conclusion.
Additionally, the PLFA approach used in this study assesses microbial biomass and structure; however, it does not provide an indication of diversity. Thus, a confident conclusion cannot be drawn for the permaculture soils having a greater microbial diversity. However, the lower G+/G− ratio and bacterial/fungal ratio (Figure 4) for the permaculture managed allotments, compared to the conventional site, do suggest a greater fungal biomass relative to bacterial biomass and a greater abundance of both G+ and G− bacteria in comparison to the conventional soil.
Other indicators of enhanced soil fertility under permaculture management are the soils’ higher organic matter and carbon contents. The permaculture managed allotments within this study use soil amendments such as crop residue, biochar, compost, and mulch. Roots are also left after harvesting, which provide continuous nutrition for soil biota between crop rotations and sowing [10]. In general, crop residue contains 4–45% C and 0.6–1% N [21]. Both permaculture managed allotments had significantly higher organic matter and carbon contents than Fenswood (HSD, mean 10.97% and 11.07%, respectively, p < 0.001). Schulz et al. [57] trialled biochar and compost ratios to discover the optimum biochar and compost mixture in response to oat growth and soil fertility for sand and loam soil. Overall, more composted biochar application caused greater plant growth attributed to increased organic carbon and nutrients released by the amendment. This supports the suggestion that the use of organic amendments within permaculture management of the allotments contributes to their greater organic matter and nutrient accumulation. This is in comparison to Fenswood Farm, which did not receive biochar, mulching, and crop residue application, where tillage was also applied, depleting the soil’s organic matter content, consequently impacting microbial abundance and nutrient release and maintenance of the soil structure [3,4,58]. In addition, Chitravadivu et al. [59] showed that soils inoculated with food compost had 30–500 times greater fungi populations compared to the use of commercial compost and nursery medium. More than double organic matter (56.8%) and total nitrogen (3.78%) were found in soils with food waste compost applied, indicating the influence of compost on organic matter and nutrient accumulation corresponding to fungi abundance.
In this lab incubation experiment where the allotment and arable farm soils were incubated at their ambient soil nitrogen, carbon, and moisture contents (reflecting the soil conditions at the time of sampling), it was observed that the permaculture managed soils had higher carbon dioxide and nitrous oxide fluxes. N2O is produced as an intermediate by both the processes of nitrification whereby organic nitrogen-derived ammonia is oxidised sequentially to nitrite and nitrate, with nitrate being subsequently used during denitrification as an electron acceptor by microbes under anaerobic conditions, producing N2O [60]. The permaculture management of the allotments encourages the continued recycling of soil nutrients and microbial activity all year long and does so by use of cover cropping, perennial crops, and organic amendments (such as manure and compost), which continuously and slowly recycle organic nitrogen to its mineral counterparts (ammonium and nitrate). The allotment soils have higher concentrations of NO3−-N and NH4+-N, partly attributed to this continuous supply of organic nitrogen, and ammonium is the greatest determinant of N2O flux within the multiple regression model explaining 72.44% (p < 0.001) variation (Table 2). Therefore, this may be one explanation for the higher N2O fluxes measured in this lab incubation, which contrasted with our initial hypothesis. Moreover, permaculture management uses no till practices, allowing the build-up of organic matter and carbon, of which the allotments’ soils had a much greater percentage compared to the conventional soil (Table 1). Therefore, ample carbon supply may further contribute to the observed higher N2O fluxes from the allotment soils as it is a key soil control factor of denitrification in natural and semi-natural UK soils [61]. The incubation study used soil samples taken within October 2022 and, therefore, only reflects the soil conditions at that time of the year. Therefore, seasonal variation and the effect of nutrient plant uptake are not considered in the present study. Moreover, it is highly likely that the commonly observed spike in N2O emissions following the application of synthetic nitrogen fertilisers under arable farming practices would have been observed if the soil sampling had occurred in spring months after fertiliser application at Fenswood Farm (110 kg N/ha in spring 2022). Akiyama et al. [62] measured N2O and NO fluxes over 3 months following a total of 20 g N m−2 nitrogen fertilisation application and discovered a high peak in both gas fluxes throughout the first month after application. Additionally, a meta-analysis investigating soil N2O emission factor (EF) with organic amendments discovered an EF of 0.57–0.3%; lower than the EF of one analysed by the IPCC for synthetic fertiliser [63]. This highlights the need for further investigations of permaculture managed soils with respect to their greenhouse gas emission and, subsequently, mitigation potential that are multiple year-round and conducted in situ to encompass seasonality, crop management, and fertilisation effects. In this context, it is likely to observe different N2O emission dynamics between continuous organic nitrogen soil amendment (permaculture) and episodic synthetic fertiliser applications (arable farming).
Carbon dioxide is taken up during photosynthesis by plants, whereby some carbon is translocated and stored in root nodules [64] and some is emitted back to the atmosphere by the mineralisation of soil organic carbon and root and microbial respiration [65]. The CO2 flux in this lab incubation study, where living plants and roots were removed by sieving and soils were incubated in the dark, represents only the microbial respiration of readily available soil organic carbon. As hypothesised, the allotment soils showed a higher soil respiration CO2 flux compared to the conventional arable soils primarily attributed to their higher carbon content and microbial biomass, whereby soil total carbon accounts for the most significant independent variable within the multiple regression model, explaining 56.92% (p < 0.01) variance in CO2 flux (Table 2), followed by bacterial biomass. Permaculture managed allotments 1 and 2 also have a higher soil moisture compared to the conventional arable site (HSD, 18.99% and 10.97% difference, respectively). Otieno et al. [64] tested the effect of soil moisture on CO2 emissions from grasslands within the Kenyan savanna, using rain shelters to reduce rainfall by 0%, 10%, and 20%. They conclude the moister soils during the rainy season that cause a growth in above ground biomass is correlated with greater photosynthesis and net ecosystem CO2 exchange. In our study, there is also a strong causation between soil moisture and CO2 flux (0.7, p < 0.01, Pearson correlation). This, therefore, could be an additional explanation for the greater CO2 emissions from the allotment soils due to enhanced soil moisture retention capacity, mostly attributed to use of cover cropping, perennial crops, and mulch application in contrast to the harvested and tilled Fenswood Farm soils. However, the permaculture practiced allotments experience a much wetter climate situated within mid-west Wales, receiving an annual average of 1106.53 mm rainfall, compared to the arable site within Southwest England, receiving an annual average of 819.01 mm (rainfall calculations based on a 30-year average period 1991–2020 from the nearest weather station) [66]. Therefore, a confident conclusion cannot be drawn that the management of permaculture accounts for the enhanced water retention over climate.
Moreover, model simulations by Biala et al. [67] estimate an increase in carbon sequestration by 0.9% and 0.55% annually with manure and compost application, respectively, based on a 20-year continuous cropping average. Global high-resolution maps based on plant mycorrhizal associations and global biome distribution estimated AMF and ectomycorrhizal fungi stored 241 GT and 100 GT of carbon within above ground biomass, respectively, compared to 29 GT of carbon in vegetation of little to no mycorrhizal [68]. Additionally, Lange et al. [69] also discovered an increase in above ground plant diversity caused an increase in soil carbon storage by large inputs of organic material. Therefore, these findings could suggest that organic practices of crop cultivation most likely cause a greater carbon sink than source via CO2 remineralisation of microbes. This explains why despite the permaculture soils having a higher CO2 flux, they also have a significantly greater carbon content than Fenswood Farm soils. Additionally, the combined ratio of CO2/total carbon of both the permaculture allotment soils is lower than that of the Fenswood conventional arable site, 74.81 ± 10.24 (SE) and 83.40 ± 6.41 (SE), respectively. This indicates that permaculture soils are sequestering more carbon relative to their CO2 emission in comparison to the conventional site, thus building a greater carbon stock. Additionally, the permaculture managed allotments use no-dig raised beds and crop rotation for minimal disturbance to the soil and microbial community [10]. However, this study’s lab incubations only indicate CO2 respiration loss and thus does not account for the release of CO2 by use of tillage, which the conventional site practices.
In contrast, all soil samples, under permaculture management and conventional practice, have a negative CH4 flux, which was not statistically different between soil types due to the high sample variability. Soil CH4 flux is dependent on the balance between the activity of methanotrophs (aerobic consumers of methane) and methanogens (anaerobic producers of methane) [70] When investigating the effect nitrogen fertiliser had on soil methane flux from a rainforest over 4 years at four levels of application (N0:0, N25:25, N50:50, N100:100 kg N ha−1 y−1), Wu et al. [70] found a significant negative correlation between methane flux with soil temperature and soil moisture. They suggest fertilisation and warmer temperatures caused an increase in nitrogen mineralisation, thus more ammonium, and its inhibiting effect on methane oxidation. Similarly, Chang et al., [71] found large amounts of nitrogen fertilisation reduced methane uptake by 23.2%, but lower levels of fertilisation increased methane uptake by 35.6% within montane forest soils, claimed to be caused by the stimulation of nitrogen-limited bacteria-oxidising methane. Ammonium is the most significant contributor within this study’s multiple regression model explaining 48.9% (p < 0.01) variance in soil CH4 flux (Table 2). This would, therefore, imply that soils with high nitrogen content result in a higher CH4 flux due to their inhibiting effect on methane oxidation [72, 70). However, the permaculture practiced allotments within this study had higher N and NH4+ concentrations and a lower CH4 flux compared to the conventional arable soil at Fenswood. This may be a result of the soil’s greater bacterial and fungal abundance, as Bodelier and Laanborek [72] state an increase in nitrifying populations, such as ammonia-oxidising bacteria, can increase methane oxidation, and Li et al. [73] found an increase in microbial diversity correlated with lower methane emissions. Within this study, total microbial biomass accounts for 14.57% (p < 0.1) variance in CH4 flux, evaluated by simple linear regression. Permaculture manages the soil through aiding its natural biogeochemical cycles, such as nutrient retention and recycling by biological processes [11]. As shown in this study, permaculture promotes a greater microbial biomass, thus may be enhancing the allotment soil’s potential for CH4 oxidation, contributing to lower CH4 fluxes. Additionally, seasonal soil sampling would have tested the effect of fertilisation application during spring 2022 on the conventionally managed arable soil at Fenswood Farm, where there may have been a spike in CH4 flux due to a fertilisation effect, as previously highlighted within the literature.
Overall, a multiple linear regression model has shown a strong influence of soil organic content and nutrients on total microbial abundance (Figure 6). Permaculture management by use of organic amendments, mulch application, cover cropping and rotation, and no-dig practices provide a constant slow release of nutrients and build-up of organic matter and carbon and, by consequence, promote a growth of bacterial and fungal biomass within the soil. Therefore, our findings, alongside previous studies, suggest permaculture management enhances microbial abundance, soil nutrients, and carbon stocks [10,20,21]. Simple linear regression models of total microbial abundance against total carbon (64.81%, p < 0.001), organic matter (44.89%, p < 0.001), nitrogen (43.37%, p < 0.001), and phosphate (93.04%, p < 0.001) further suggest this.
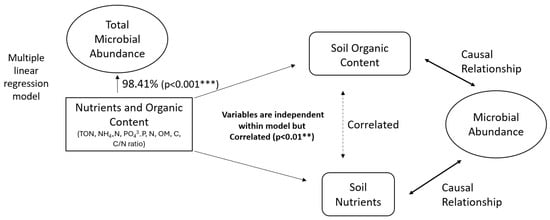
Figure 6.
Statistical analysis indicating relationships between soil organic nutrients, organic content, and microbial abundance.
Therefore, the fertile soils of both allotments show the ability to enrich soils in both rural and urban settings and, in accordance with similar studies, prove the hypothesised increased organic matter, carbon, and greater nutrient retention within the permaculture soils to be true and attributed to permaculture management mimicking a whole ecosystem design approach, applying organic amendments, creating minimal disturbance to the soil and mycelium network, and promoting microbial activity.
Current movements of community-based organisations and projects are implementing localised food systems that enrich soil fertility through using the principles of permaculture. A localised food system where communities provide for themselves may have the potential to significantly improve rural and urban soils [74]. However, the reality of implementing a localised food system globally requires a complete change in societal behaviour and diet to fit the food that could be grown within the local environment [2]. Education of permaculture and its benefits would need to be adopted on a global platform, with increased research in permaculture within academia to support the evidence of its benefits [2,12]. Additionally, implementing local food systems does not support the current global system and economy of transnational companies [2]. Therefore, a bottom-up approach is most likely to give rise to the agroecology movement, such as community-based organisations or non-governmental organisations [74,75].
5. Conclusions
This study found permaculture management of allotments leads to significantly more fertile soils than the conventionally managed arable soils, thus significantly contributing to the currently limited peer-reviewed literature on the influence of permaculture management on soil properties.
The management of permaculture focused on mimicking the natural recycling of an ecosystem with addition of organic amendments, little disturbance to the soil using no-dig raised beds, and crop diversity and rotation to aid microbial activity and synergy with the plant, creating a dense network of hyphae within the soil, contributing to a much greater carbon, nutrient, and organic content compared to the conventionally managed soil. Soil CO2 and N2O fluxes were greater within the permaculture practiced soils, unlike CH4 flux, which was lower, potentially due to the increased nitrogen and carbon contents caused by the application of organic amendments and cover cropping. However, synthetic nitrogen fertiliser used on the conventional soils is a large source of GHGs, but it did not affect the arable soils of this study because fertiliser was applied in spring and soil samples were taken during autumn. Therefore, future temporal sampling of soils under both managements would reduce this uncertainty.
Overall analysis has shown significantly more fertile soils under permaculture management compared to conventional practices. Methods of agroecology and holistic management of the permaculture allotments, such as no-dig raised beds, organic amendments (compost, mulch, biochar, seaweed), cover cropping and rotations, and perennial crops, maximise the edge and biodiversity. Thereby, they create a food system where microbial communities can grow and thrive, breaking down organic matter, promoting carbon sequestration, and exchanging vital nutrients with the plant, creating rich and fertile soils.
The study, therefore, highlights the potential benefits of permaculture management for crop growth and soil fertility, reducing the use of fertiliser needs and, in the long-term, reducing greenhouse gas emissions. The comparable conventional site within this study could adopt this approach through using an agroforestry permaculture design framework, using alleyways of tree lines and companion planting, splitting the field into multiple no-dig beds, to create more biodiverse edges attracting pollinators and diversity and enhancing soil fertility and carbon storage. Accordingly, we present one example of how permaculture could be implemented on a larger arable farm scale and provide an alternative agriculture system that promotes carbon sequestration and soil restoration.
Author Contributions
R.F.W., M.R. and F.S. contributed to the study conceptualisation and experimental design. R.F.W. conducted the fieldwork and laboratory experimentation as well as data analysis and manuscript preparation. R.F.W., M.R. and F.S. edited, read, and agreed to the published version of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
Data available upon request to the corresponding author.
Acknowledgments
The authors are grateful to J. Williamson for his enthusiasm and permission to use the permaculture managed allotments for the basis of this study and to the University of Bristol Fenswood Farm for comparison of management. We are also thankful to the University of Bristol Geography department and the Organic Geochemistry Unity at the School of Chemistry for their facilities to conduct soil analysis.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Faltin, J. Failing Food Supply: Permaculture’s Potential. Bachelor’s Thesis, University of Nebraska-Lincoln, Lincoln, Nebraska, 2017. Available online: https://digitalcommons.unl.edu/envstudtheses/213/ (accessed on 29 January 2024).
- Hathaway, M.D. Agroecology and permaculture: Addressing key ecological problems by rethinking and redesigning agricultural systems. J. Environ. Stud. Sci. 2016, 6, 239–250. [Google Scholar] [CrossRef]
- Reganold, J.P. Comparison of soil properties as influenced by organic and conventional farming systems. Am. J. Altern. Agric. 1988, 3, 144–155. [Google Scholar] [CrossRef]
- Vanwalleghem. Soil erosion and conservation. In International Encyclopedia of Geography: People, the Earth, Environment and Technology: People, the Earth, Environment and Technology; 2017; pp. 1–10. Available online: https://onlinelibrary.wiley.com/doi/abs/10.1002/9781118786352.wbieg0381 (accessed on 29 January 2024). [CrossRef]
- United Nations, 2022. FAO Warns 90 per Cent of Earth’s Topsoil at Risk by 2050. UN News, Global Perspective Human Stories. United Nations. [Online]. Available online: https://news.un.org/en/story/2022/07/1123462 (accessed on 29 January 2024).
- Montgomery. Soil erosion and agricultural sustainability. Proc. Natl. Acad. Sci. USA 2007, 104, 13268–13272. [Google Scholar] [CrossRef]
- Gomiero, T.; Pimentel, D.; Paoletti, M.G. Environmental impact of different agricultural management practices: Conventional vs. organic agriculture. Crit. Rev. Plant Sci. 2011, 30, 95–124. [Google Scholar] [CrossRef]
- Reeve, J.R.; Hoagland, L.A.; Villalba, J.J.; Carr, P.M.; Atucha, A.; Cambardella, C.; Davis, D.R.; Delate, K. Organic farming, soil health, and food quality: Considering possible links. Adv. Agron. 2016, 137, 319–367. [Google Scholar]
- Tully, K.L.; McAskill, C. Promoting soil health in organically managed systems: A review. Org. Agric. 2020, 10, 339–358. [Google Scholar] [CrossRef]
- Hemenway, T. GAIA’S GARDEN a Guide to Home Scale Permaculture, 2nd ed.; Chelsea Green Publishing: White River Junction, VT, USA, 2009. [Google Scholar]
- Mollison, B.C.; Slay, R.M.; Girard, J.L.; Girard, J.L. Introduction to Permaculture. Yankee Permaculture. Publisher and Distributor of Permaculture Publications POB 52, Sparr FL 32192-0052 USA. 1991. Available online: https://www.academia.edu/download/62514467/Bill_Mollison_-_Permaculture_Design_Course_20200328-35259-mdyiv7.pdf (accessed on 15 October 2022).
- Leni-Konig. Beyond School Gardens: Permaculture Food Forests Enhance Ecosystem Services While Achieving Education for Sustainable Development Goals. Ph.D. Thesis, Harvard University, Cambridge, MA, USA, 2020.
- Minckler, S.J. Permaculture: The Need for Increased Science. Ph.D. Thesis, Washington State University, Pullman, WA, USA, 2019. [Google Scholar]
- Fiebrig, I.; Zikeli, S.; Bach, S.; Gruber, S. Perspectives on permaculture for commercial farming: Aspirations and realities. Org. Agric. 2020, 10, 379–394. [Google Scholar] [CrossRef]
- Morel, K.; Léger, F.; Ferguson, R.S. Permaculture. In Encyclopedia of Ecology, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 559–567. [Google Scholar] [CrossRef]
- De Tombeur, F.; Sohy, V.; Chenu, C.; Colinet, G.; Cornelis, J.T. Effects of permaculture practices on soil physicochemical properties and organic matter distribution in aggregates: A case study of the Bec-Hellouin Farm (France). Front. Environ. Sci. 2018, 6, 116. [Google Scholar] [CrossRef]
- French, E.; Kaplan, I.; Iyer-Pascuzzi, A.; Nakatsu, C.H.; Enders, L. Emerging strategies for precision microbiome management in diverse agroecosystems. Nat. Plants 2021, 7, 256–267. [Google Scholar] [CrossRef]
- Bak, G.R.; Lee, G.J.; Lee, J.T.; Jee, S.N. Crop rotation affects biological properties of rhizosphere soil and productivity of Kimchi cabbage (Brassica rapa ssp. pekinensis) compared to monoculture. Hortic. Environ. Biotechnol. 2022, 63, 613–625. [Google Scholar] [CrossRef]
- Lori, M.; Symnaczik, S.; Mäder, P.; De Deyn, G.; Gattinger, A. Organic farming enhances soil microbial abundance and activity—A meta-analysis and meta-regression. PLoS ONE 2017, 12, e0180442. [Google Scholar] [CrossRef] [PubMed]
- Rhodes, C.J. Feeding and healing the world: Through regenerative agriculture and permaculture. Sci. Prog. 2012, 95, 345–446. [Google Scholar] [CrossRef] [PubMed]
- Sneha, S.; Anitha, B.; Sahair, R.A.; Raghu, N.; Gopenath, T.S.; Chandrashekrappa, G.K.; Basalingappa, M.K. Biofertilizer for crop production and soil fertility. Acad. J. Agric. Res. 2018, 6, 299–306. [Google Scholar]
- Smith, S.E.; Jakobsen, I.; Grønlund, M.; Smith, F.A. Roles of arbuscular mycorrhizas in plant phosphorus nutrition: Interactions between pathways of phosphorus uptake in arbuscular mycorrhizal roots have important implications for understanding and manipulating plant phosphorus acquisition. Plant Physiol. 2011, 156, 1050–1057. [Google Scholar] [CrossRef] [PubMed]
- Mäder, P.; Edenhofer, S.; Boller, T.; Wiemken, A.; Niggli, U. Arbuscular mycorrhizae in a long-term field trial comparing low-input (organic, biological) and high-input (conventional) farming systems in a crop rotation. Biol. Fertil. Soils 2000, 31, 150–156. [Google Scholar] [CrossRef]
- Sainju, U.M.; Stevens, W.B.; Caesar-TonThat, T.; Liebig, M.A. Soil greenhouse gas emissions affected by irrigation, tillage, crop rotation, and nitrogen fertilization. J. Environ. Qual. 2012, 41, 1774–1786. [Google Scholar] [CrossRef]
- Huang, Y.; Ren, W.; Wang, L.; Hui, D.; Grove, J.H.; Yang, X.; Tao, B.; Goff, B. Greenhouse gas emissions and crop yield in no-tillage systems: A meta-analysis. Agric. Ecosyst. Environ. 2018, 268, 144–153. [Google Scholar] [CrossRef]
- Gao, Y.; Cabrera Serrenho, A. Greenhouse gas emissions from nitrogen fertilizers could be reduced by up to one-fifth of current levels by 2050 with combined interventions. Nat. Food 2023, 4, 170–178. [Google Scholar] [CrossRef]
- Sanz-Cobena, A.; Lassaletta, L.; Aguilera, E.; del Prado, A.; Garnier, J.; Billen, G.; Iglesias, A.; Sanchez, B.; Guardia, G.; Abalos, D.; et al. Strategies for greenhouse gas emissions mitigation in Mediterranean agriculture: A review. Agric. Ecosyst. Environ. 2017, 238, 5–24. [Google Scholar] [CrossRef]
- Hu, Y.; Li, D.; Wu, Y.; Liu, S.; Li, L.; Chen, W.; Wu, S.; Meng, Q.; Feng, H.; Siddique, K.H. Mitigating greenhouse gas emissions by replacing inorganic fertilizer with organic fertilizer in wheat–maize rotation systems in China. J. Environ. Manag. 2023, 344, 118494. [Google Scholar] [CrossRef]
- IPCC. IPCC, 2023: Summary for Policymakers. In Climate Change 2023: Synthesis Report. Contribution of Working Groups I, II and III to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Lee, H.H., Romero, J., Eds.; IPCC: Geneva, Switzerland, 2023; pp. 1–34. [Google Scholar] [CrossRef]
- Frostegård, Å.; Tunlid, A.; Bååth, E. Microbial biomass measured as total lipid phosphate in soils of different organic content. J. Microbiol. Methods 1991, 14, 151–163. [Google Scholar] [CrossRef]
- Joergensen. Phospholipid fatty acids in soil—Drawbacks and future prospects. Biol. Fertil. Soils 2022, 58, 1–6. [Google Scholar] [CrossRef]
- Jensen, E.S.; Peoples, M.B.; Boddey, R.M.; Gresshoff, P.M.; Hauggaard-Nielsen, H.; JRAlves, B.; Morrison, M.J. Legumes for mitigation of climate change and the provision of feedstock for biofuels and biorefineries. A review. Agron. Sustain. Dev. 2012, 32, 329–364. [Google Scholar] [CrossRef]
- Digimap, 2023. Digimap. [Online]. Available online: https://digimap.edina.ac.uk (accessed on 12 January 2023).
- Rhodes, C.J. The imperative for regenerative agriculture. Sci. Prog. 2017, 100, 80–129. [Google Scholar] [CrossRef]
- OpenStreetMap.org 2023. OpenStreetMap. [Online]. Available online: www.openstreetmap.org/copyright/ (accessed on 23 February 2023).
- Pennock, D.; Yates, T.; Braidek, J. Soil Sampling Designs. In Soil Sampling and Methods of Analysis: Chapter 1; CRC Press: Boca Raton, FL, USA, 2008; pp. 863–869. [Google Scholar]
- Malvern Instruments Ltd. Mastersizer 3000 User Manual; Transport; Malvern Analytical Ltd.: Worcestershire, UK, 2017. [Google Scholar]
- Natural Resources Conservation Service USDA, nd. Soil Textural Calculator. Natural Resources Conservation Service. U.S. Department of Agriculutre. [Online]. Available online: https://www.nrcs.usda.gov/resources/education-and-teaching-materials/soil-texture-calculator (accessed on 19 December 2022).
- Buyer, J.S.; Sasser, M. High throughput phospholipid fatty acid analysis of soils. Appl. Soil Ecol. 2012, 61, 127–130. [Google Scholar] [CrossRef]
- Kroppenstedt, R.M. The genus Nocardiopsis. In The Prokaryotes 2; Balows, A., Trüper, H.G., Dworkin, M., Harder, W., Schleifer, K.H., Eds.; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 1992; pp. 1139–1156. [Google Scholar]
- Frostegård, A.; Bååth, E. The use of phospholipid fatty acid analysis to estimate bacterial and fungal biomass in soil. Biol. Fertil. Soils 1996, 22, 59–65. [Google Scholar] [CrossRef]
- Zelles, L. Phospholipid fatty acid profiles in selected members of soil microbial communities. Chemosphere 1997, 35, 275–294. [Google Scholar] [CrossRef]
- Jiang, S.; An, X.; Shao, Y.; Kang, Y.; Chen, T.; Mei, X.; Dong, C.; Xu, Y.; Shen, Q. Responses of arbuscular mycorrhizal fungi occurrence to organic fertilizer: A meta-analysis of field studies. Plant Soil 2021, 469, 89–105. [Google Scholar] [CrossRef]
- Oehl, F.; Sieverding, E.; Mäder, P.; Dubois, D.; Ineichen, K.; Boller, T.; Wiemken, A. Impact of long-term conventional and organic farming on the diversity of arbuscular mycorrhizal fungi. Oecologia 2004, 138, 574–583. [Google Scholar] [CrossRef] [PubMed]
- Edlinger, A.; Garland, G.; Hartman, K.; Banerjee, S.; Degrune, F.; García-Palacios, P.; Hallin, S.; Valzano-Held, A.; Herzog, C.; Jansa, J.; et al. Agricultural management and pesticide use reduce the functioning of beneficial plant symbionts. Nat. Ecol. Evol. 2022, 6, 1145–1154. [Google Scholar] [CrossRef]
- Fu, B.; Chen, L.; Huang, H.; Qu, P.; Wei, Z. Impacts of crop residues on soil health: A review. Environ. Pollut. Bioavailab. 2021, 33, 164–173. [Google Scholar] [CrossRef]
- Brussaard, L.; De Ruiter, P.C.; Brown, G.G. Soil biodiversity for agricultural sustainability. Agric. Ecosyst. Environ. 2007, 121, 233–244. [Google Scholar] [CrossRef]
- Lazcano, C.; Zhu-Barker, X.; Decock, C. Effects of organic fertilizers on the soil microorganisms responsible for N2O emissions: A review. Microorganisms 2021, 9, 983. [Google Scholar] [CrossRef] [PubMed]
- Hestrin, R.; Hammer, E.C.; Mueller, C.W.; Lehmann, J. Synergies between mycorrhizal fungi and soil microbial communities increase plant nitrogen acquisition. Commun. Biol. 2019, 2, 233. [Google Scholar] [CrossRef] [PubMed]
- Fidel, R.B.; Laird, D.A.; Spokas, K.A. Sorption of ammonium and nitrate to biochars is electrostatic and pH-dependent. Sci. Rep. 2018, 8, 17627. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Hoffman-Krull, K.; Bidwell, A.L.; DeLuca, T.H. Locally produced wood biochar increases nutrient retention and availability in agricultural soils of the San Juan Islands, USA. Agric. Ecosyst. Environ. 2016, 233, 43–54. [Google Scholar] [CrossRef]
- Zheng, H.; Wang, Z.; Deng, X.; Herbert, S.; Xing, B. Impacts of adding biochar on nitrogen retention and bioavailability in agricultural soil. Geoderma 2013, 206, 32–39. [Google Scholar] [CrossRef]
- Peng, J.; Han, X.; Li, N.; Chen, K.; Yang, J.; Zhan, X.; Luo, P.; Liu, N. Combined application of biochar with fertilizer promotes nitrogen uptake in maize by increasing nitrogen retention in soil. Biochar 2021, 3, 367–379. [Google Scholar] [CrossRef]
- Ding, Y.; Liu, Y.X.; Wu, W.X.; Shi, D.Z.; Yang, M.; Zhong, Z.K. Evaluation of biochar effects on nitrogen retention and leaching in multi-layered soil columns. Water Air Soil Pollut. 2010, 213, 47–55. [Google Scholar] [CrossRef]
- Atkinson, C.J.; Fitzgerald, J.D.; Hipps, N.A. Potential mechanisms for achieving agricultural benefits from biochar application to temperate soils: A review. Plant Soil 2010, 337, 1–18. [Google Scholar] [CrossRef]
- Romaniuk, R.; Giuffré, L.; Costantini, A.; Nannipieri, P. Assessment of soil microbial diversity measurements as indicators of soil functioning in organic and conventional horticulture systems. Ecol. Indic. 2011, 11, 1345–1353. [Google Scholar] [CrossRef]
- Schulz, H.; Dunst, G.; Glaser, B. Positive effects of composted biochar on plant growth and soil fertility. Agron. Sustain. Dev. 2013, 33, 817–827. [Google Scholar] [CrossRef]
- Krauss, M.; Wiesmeier, M.; Don, A.; Cuperus, F.; Gattinger, A.; Gruber, S.; Haagsma, W.K.; Peigné, J.; Palazzoli, M.C.; Schulz, F.; et al. Reduced tillage in organic farming affects soil organic carbon stocks in temperate Europe. Soil Tillage Res. 2022, 216, 105262. [Google Scholar] [CrossRef]
- Chitravadivu, C.; Balakrishnan, V.; Manikandan, J.; Elavazhagan, T.; Jayakumar, S. Application of food waste compost on soil microbial population in groundnut cultivated soil, India. Middle-East J. Sci. Res. 2009, 4, 90–93. [Google Scholar]
- Kumar, A.; Medhi, K.; Fagodiya, R.K.; Subrahmanyam, G.; Mondal, R.; Raja, P.; Malyan, S.K.; Gupta, D.K.; Gupta, C.K.; Pathak, H. Molecular and ecological perspectives of nitrous oxide producing microbial communities in agro-ecosystems. Rev. Environ. Sci. Bio/Technol. 2020, 19, 717–750. [Google Scholar] [CrossRef]
- Sgouridis, F.; Ullah, S. Soil greenhouse gas fluxes, environmental controls, and the partitioning of N2O sources in UK natural and seminatural land use types. J. Geophys. Res. Biogeosci. 2017, 122, 2617–2633. [Google Scholar] [CrossRef]
- Akiyama, H.; Tsuruta, H.; Watanabe, T. N2O and NO emissions from soils after the application of different chemical fertilizers. Chemosphere-Glob. Chang. Sci. 2000, 2, 313–320. [Google Scholar] [CrossRef]
- Charles, A.; Rochette, P.; Whalen, J.K.; Angers, D.A.; Chantigny, M.H.; Bertrand, N. Global nitrous oxide emission factors from agricultural soils after addition of organic amendments: A meta-analysis. Agric. Ecosyst. Environ. 2017, 236, 88–98. [Google Scholar] [CrossRef]
- Otieno, D.O.; K’Otuto, G.O.; Maina, J.N.; Kuzyakov, Y.; Onyango, J.C. Responses of ecosystem carbon dioxide fluxes to soil moisture fluctuations in a moist Kenyan savanna. J. Trop. Ecol. 2010, 26, 605–618. [Google Scholar] [CrossRef]
- Plaza-Bonilla, D.; Cantero-Martínez, C.; Bareche, J.; Arrúe, J.L.; Álvaro-Fuentes, J. Soil carbon dioxide and methane fluxes as affected by tillage and N fertilization in dryland conditions. Plant Soil 2014, 381, 111–130. [Google Scholar] [CrossRef]
- UK Climate Averages (nd). UK Climate Averages. MetOffice.gov.uk. [Online]. Available online: https://www.metoffice.gov.uk/research/climate/maps-and-data/uk-climate-averages (accessed on 4 May 2024).
- Biala, J.; Wilkinson, K.; Henry, B.; Singh, S.; Bennett-Jones, J.; De Rosa, D. The potential for enhancing soil carbon levels through the use of organic soil amendments in Queensland, Australia. Reg. Environ. Chang. 2021, 21, 1–15. [Google Scholar] [CrossRef]
- Soudzilovskaia, N.A.; van Bodegom, P.M.; Terrer, C.; Zelfde, M.V.T.; McCallum, I.; Luke McCormack, M.; Fisher, J.B.; Brundrett, M.C.; de Sá, N.C.; Tedersoo, L. Global mycorrhizal plant distribution linked to terrestrial carbon stocks. Nat. Commun. 2019, 10, 5077. [Google Scholar] [CrossRef] [PubMed]
- Lange, M.; Eisenhauer, N.; Sierra, C.A.; Bessler, H.; Engels, C.; Griffiths, R.I.; Mellado-Vázquez, P.G.; Malik, A.A.; Roy, J.; Scheu, S.; et al. Plant diversity increases soil microbial activity and soil carbon storage. Nat. Commun. 2015, 6, 6707. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Peng, C.; Wang, C.; Chen, H.; Liu, W.; Liu, Z.; Wang, H.; Li, H.; Chen, D.; Li, Y.; et al. Responses of soil CH4 fluxes to nitrogen addition in two tropical montane rainforests in southern China. For. Ecosyst. 2022, 9, 100031. [Google Scholar] [CrossRef]
- Chang, R.; Liu, X.; Wang, T.; Li, N.; Bing, H. Stimulated or inhibited response of methane flux to nitrogen addition depends on nitrogen levels. J. Geophys. Res. Biogeosciences 2021, 126, e2021JG006600. [Google Scholar] [CrossRef]
- Bodelier, L.; Laanbroek, H.J. Nitrogen as a regulatory factor of methane oxidation in soils and sediments. FEMS Microbiol. Ecol. 2004, 47, 265–277. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Peng, C.; Zhang, J.; Li, Y.; Song, X. Nitrogen addition decreases methane uptake caused by methanotroph and methanogen imbalances in a Moso bamboo forest. Sci. Rep. 2021, 11, 5578. [Google Scholar] [CrossRef]
- Bell, G. The Permaculture Way: Practical Steps to Create a Self-Sustaining World; Chelsea Green Publishing: White River Junction, VT, USA, 2005. [Google Scholar]
- Suh, J. Towards sustainable agricultural stewardship: Evolution and future directions of the permaculture concept. Environ. Values 2014, 23, 75–98. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).