
Effect of Maize Canopy Structure on Light Interception and Radiation Use Efficiency at Different Canopy Layers



Abstract
1. Introduction
2. Materials and Methods
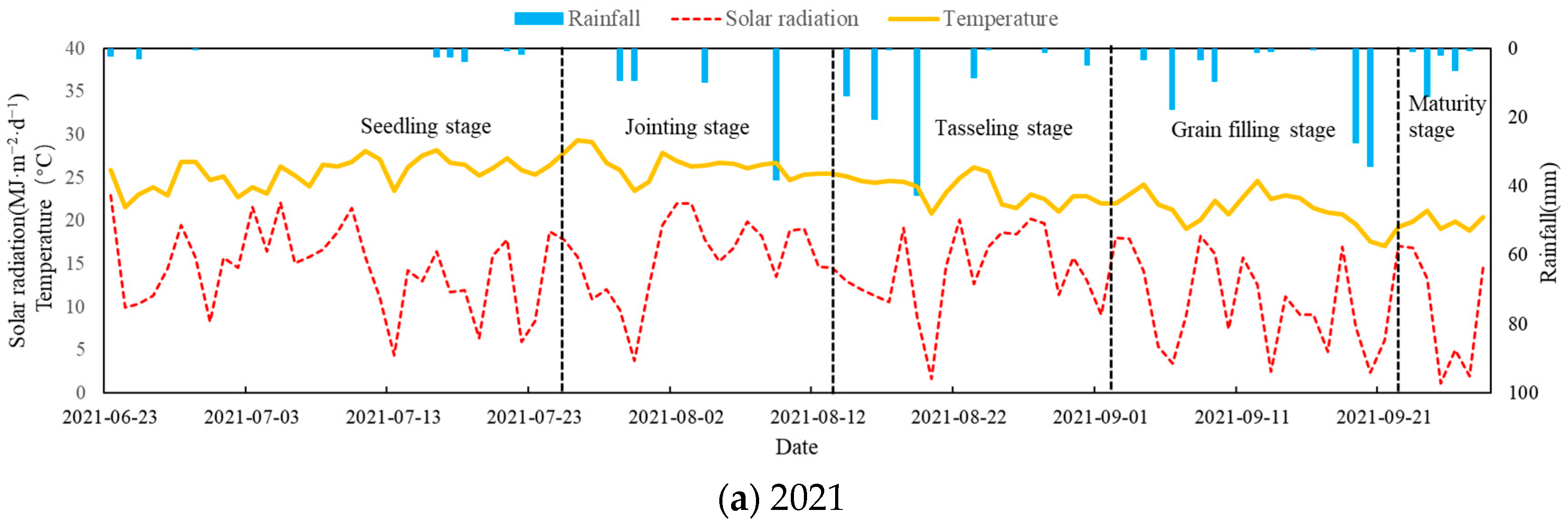
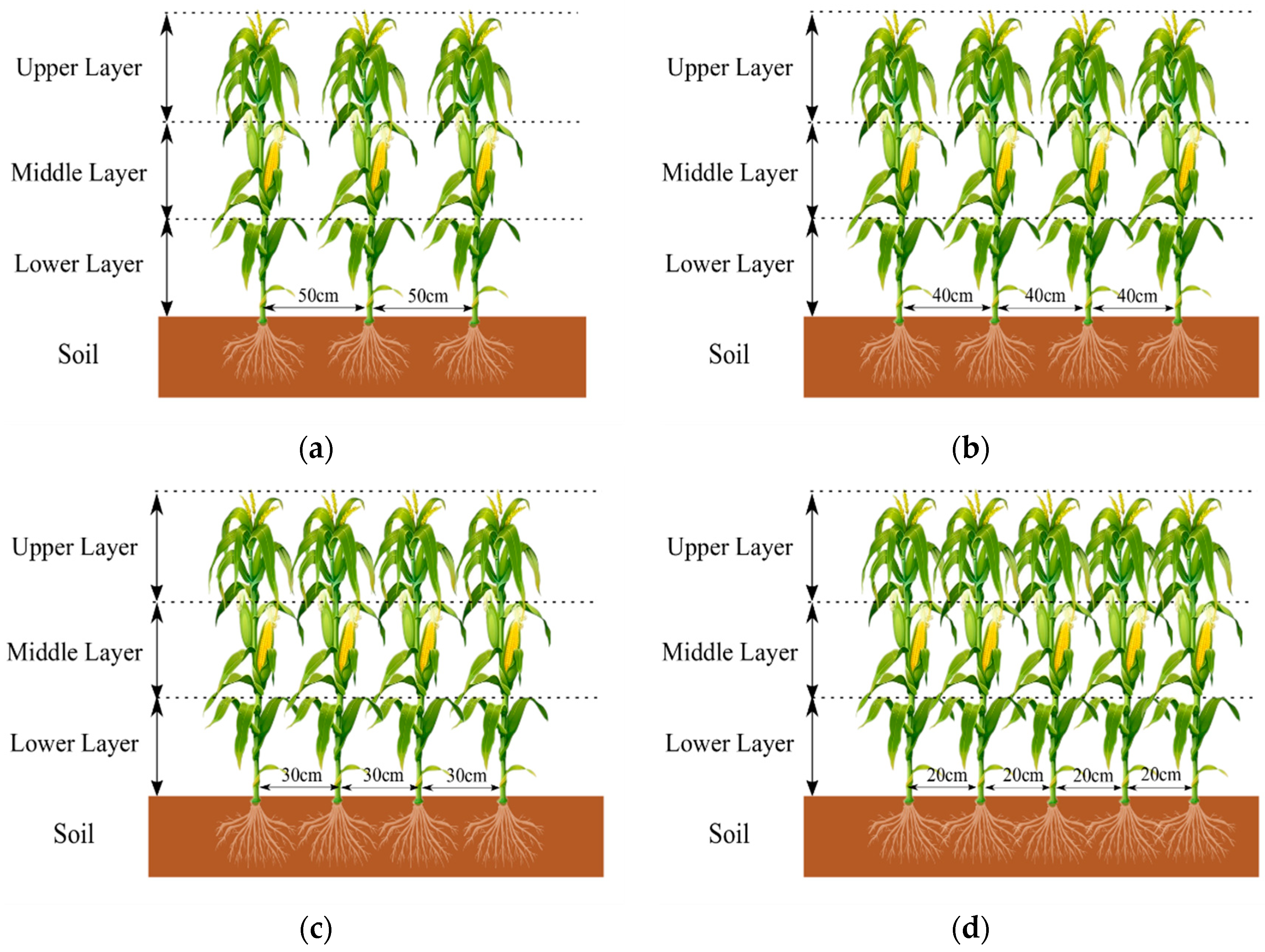
2.1. Experimental Conditions
2.2. Measurements and Calculations
2.2.1. Photosynthetically Active Radiation Measurements
2.2.2. Light Interception Rate
2.2.3. Above-Ground Biomass and Leaf Area Index
2.2.4. Radiation Use Efficiency and Precipitation Use Efficiency
2.2.5. Light Attenuation
2.3. Statistical Analysis
3. Results
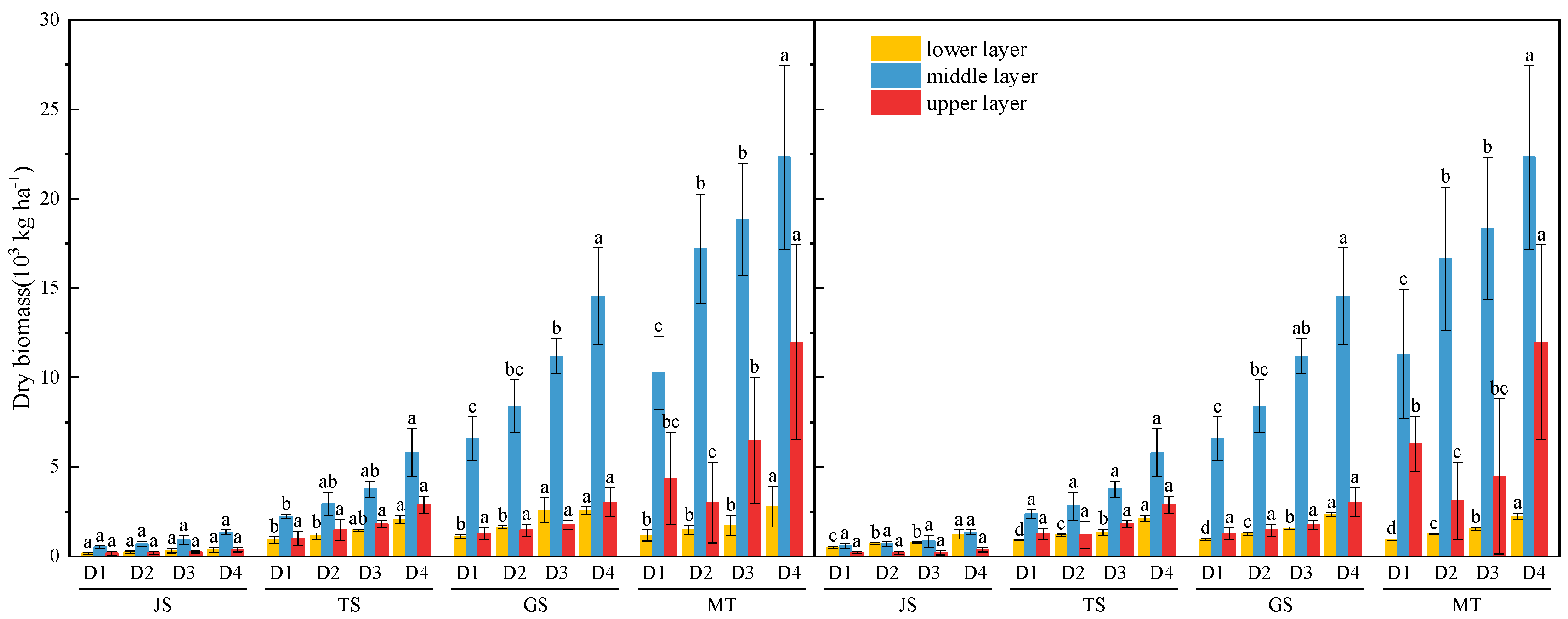
3.1. Biomass Yield and Grain Yield
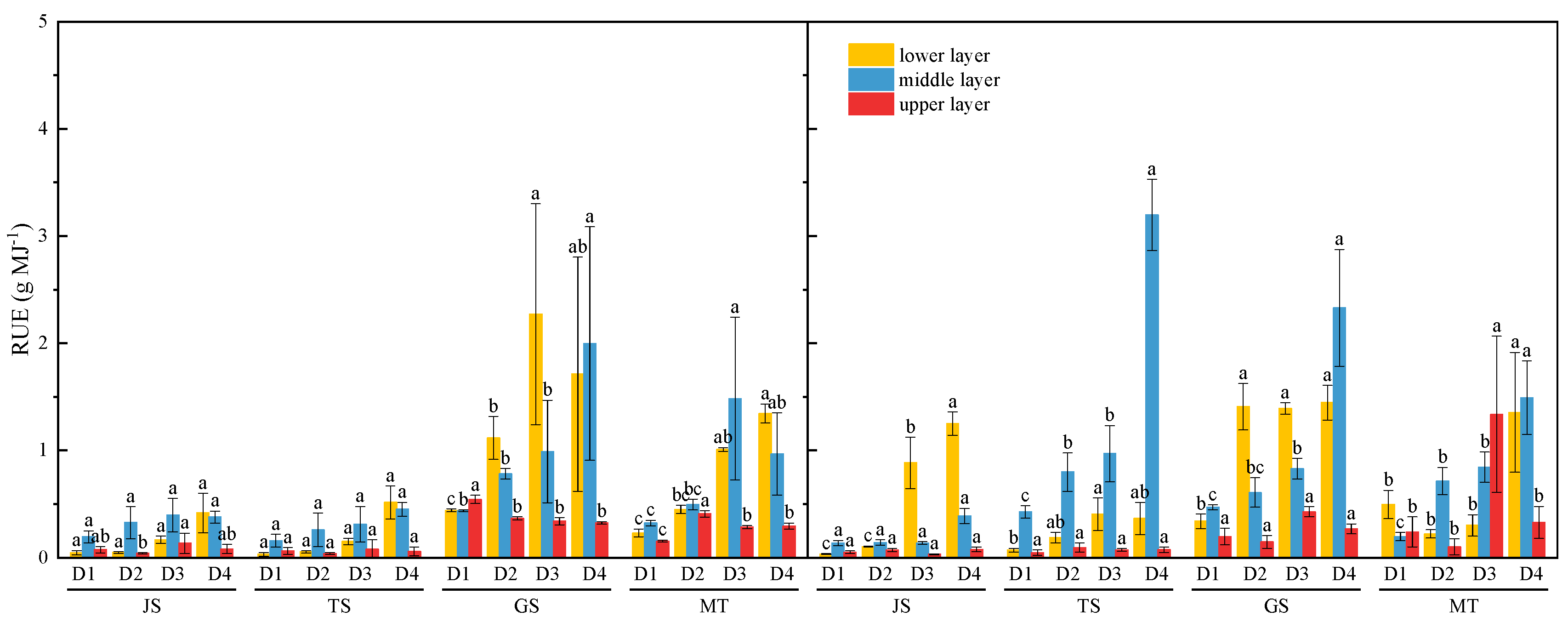
3.2. Radiation Use Efficiency
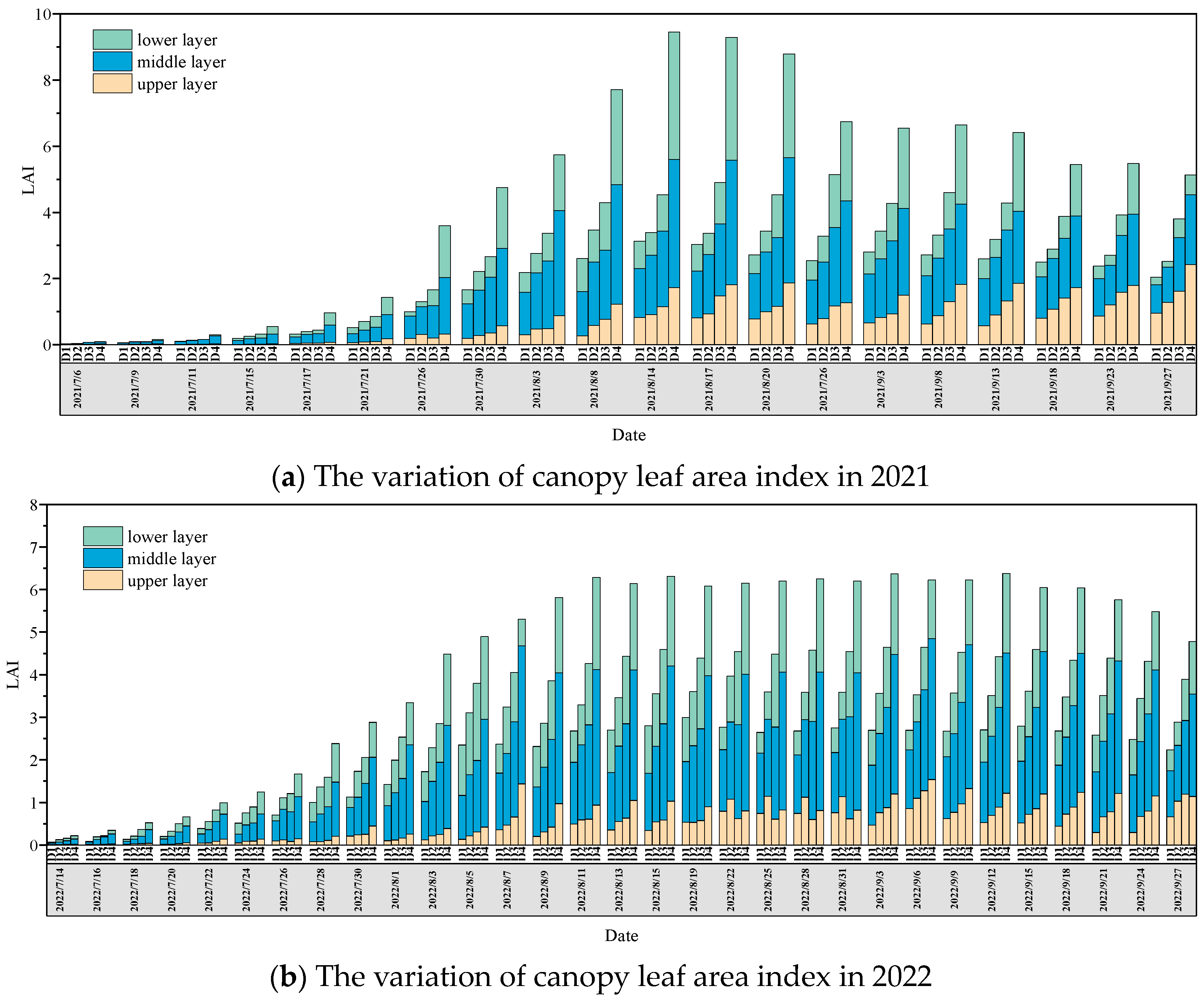
3.3. Leaf Area Index of Different Densities
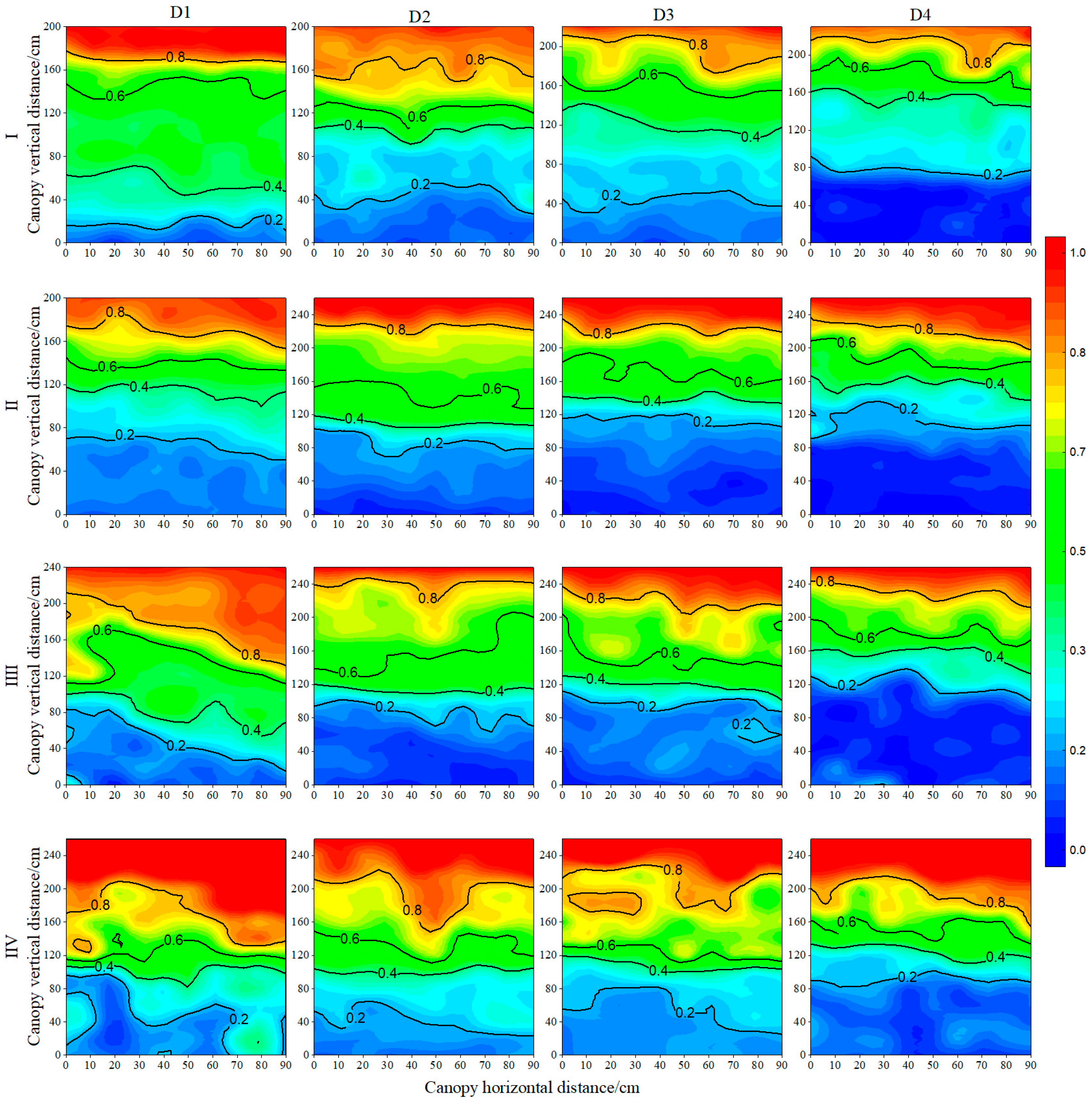
3.4. Maize Transmitted Photosynthetically Active Radiation of Entire Canopy throughout Growth Period
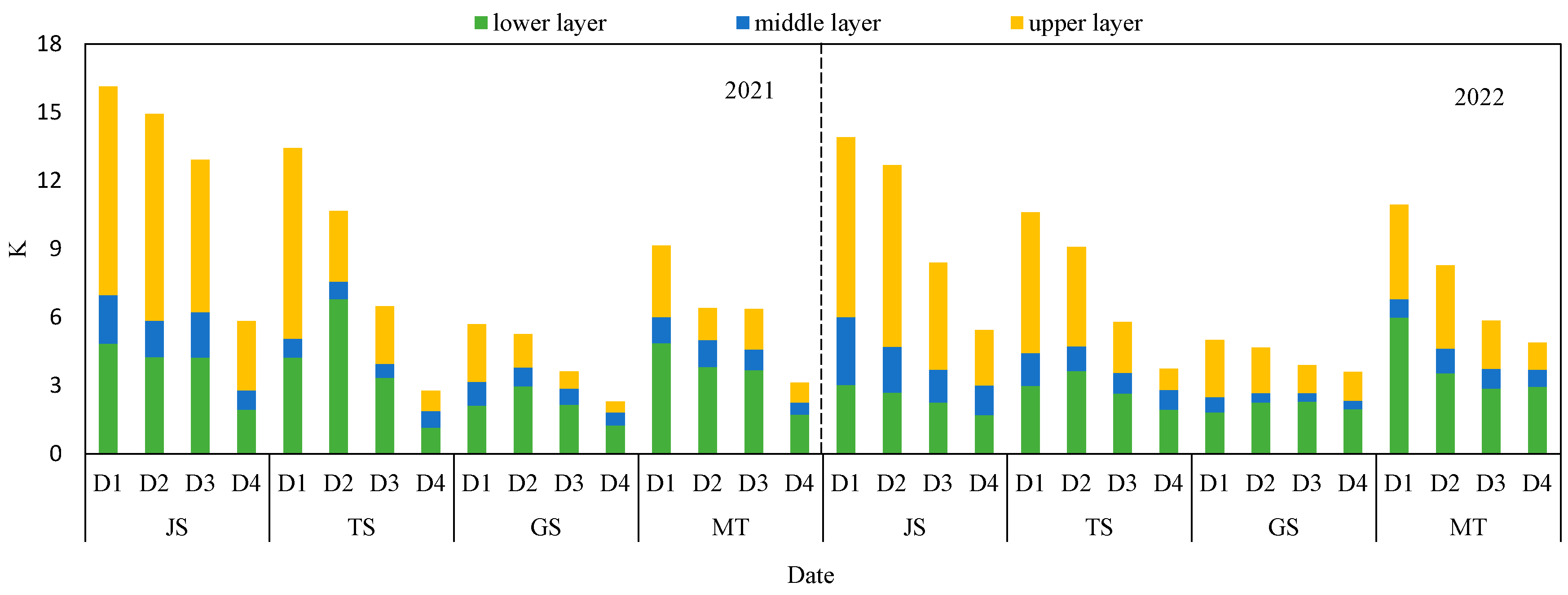
3.5. The Light Extinction Coefficient
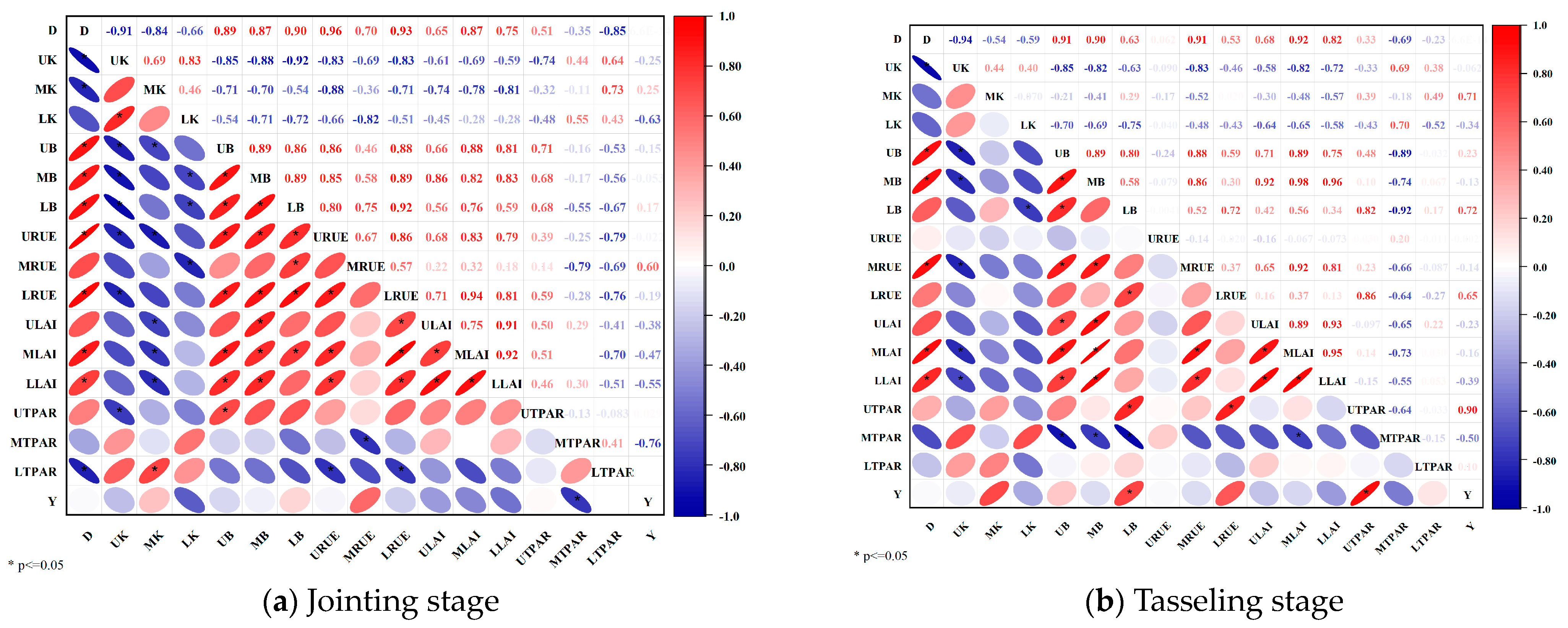
3.6. Relation between the K, B, RUE, LAI and TPAR to Density
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Erenstein, O.; Jaleta, M.; Sonder, K. Global maize production, consumption and trade: Trends and R & D implications. Food Secur. 2022, 14, 1295–1319. [Google Scholar]
- Sherry, A.T.; Laura, M.; Rachel, R. Maize agro-food systems to ensure food and nutrition security in reference to the Sustainable Development Goals. Glob. Food Secur. 2020, 25, 100327. [Google Scholar]
- Zhang, L.; Zhang, F.R.; Jiang, G.H.; Yao, H.M. Potential improvement of medium-low yielded farm land and guaran tee of food safety in China. Res. Agric. Mod. 2005, 26, 22–25. [Google Scholar]
- Gregory, P.J.; Ingram, J. Global change and food and forest production: Future scientific challenges. Agric. Ecosyst. Environ. 2000, 82, 3–14. [Google Scholar] [CrossRef]
- Mottaleb, K.A.; Kruseman, G.; Frija, A.; Sonder, K.; Santiago, L. Projecting wheat demand in China and India for 2030 and 2050: Implications for food security. Front. Nutr. 2022, 9, 10. [Google Scholar] [CrossRef] [PubMed]
- Duvick, D.N.; Cassman, K.G. Post-Green revolution trends in yield potential of temperate maize in the north-central United States. Crop Sci. 1999, 39, 1622–1630. [Google Scholar] [CrossRef]
- Pimentel, D.; Harvey, C.; Resosudarmo, P.; Sinclair, K.; Kurz, D.; McNair, M.; Crist, S.; Shpritz, L.; Fitton, L.; Saffouri, R.; et al. Environmental and economic costs of soils erosion and conservation benefits. Science 1995, 267, 1117–1123. [Google Scholar] [CrossRef] [PubMed]
- Lobell, D.B.; Cassman, K.G.; Field, C.B. Crop yield gaps: Their importance, magnitudes, and causes. Annu. Rev. Environ. Resour. 2009, 34, 179–204. [Google Scholar] [CrossRef]
- Li, S.K.; Wang, C.T. The methods of obtaining and expressing information of crop plant shape and population structure. J. Shihezi Univ. Nat. Sci. 1997, 1, 250–256. [Google Scholar]
- Liu, J.H.; Wang, Z.M.; Li, L.J.; Zhang, H.M. Higher-yield is key technical method of maintaining future food security in China. Res. Agric. Mod. 2003, 24, 161–165. [Google Scholar]
- Ren, B.; Liu, W.; Zhang, J.; Dong, S.; Liu, P.; Zhao, B. Effects of plant density on the photosynthetic and chloroplast characteristics of maize under high-yielding conditions. Sci. Nat. 2017, 104, 3–4. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Liu, W.; Yang, Y. Marginal superiority of maize: An indicator for density tolerance under high plant density. Sci. Rep. 2020, 10, 15378. [Google Scholar] [CrossRef] [PubMed]
- Ciampitti, I.A.; Vyn, T.J. A comprehensive study of plant density consequences on nitrogen uptake dynamics of maize plants from vegetative to reproductive stages. Field Crops Res. 2011, 121, 2–18. [Google Scholar] [CrossRef]
- Hirose, T. Development of the Monsi-Saeki theory on canopy structure and function. Ann. Bot. 2005, 95, 483–494. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Rahman, T.; Song, C.; Yang, F.; Su, B.; Cui, L. Relationships among light distribution, radiation use efficiency and land equivalent ratio in maize-soybean strip intercropping. Field Crops Res. 2018, 224, 91–101. [Google Scholar] [CrossRef]
- Beruski, G.C.; Schiebelbein, L.M.; André, B.P. Maize Yield Components as Affected by Plant Population, Planting Date and Soil Coverings in Brazil. Agriculture 2020, 10, 579. [Google Scholar] [CrossRef]
- Du, X.B.; Wang, Z.; Lei, W.X.; Kong, L.C. Increased planting density combined with reduced nitrogen rate to achieve high yield in maize. Sci. Rep. 2023, 11, 358. [Google Scholar] [CrossRef]
- Bahadur, M.M.; Ashrafuzzaman, M.; Chowdhury, M.F. Growth and Yield Component Responses of Maize as Affected by Population Density. Pak. J. Biol. Sci. 1999, 2, 1092–1095. [Google Scholar] [CrossRef]
- Sibonginkosi, N.; Mabuza, M.; Tana, T. Effect of Plant Density on Growth and Yield of Maize [Zea mays (L.)] Hybrids at Luyengo, Middleveld of Eswatini. Asian Plant Res. J. 2020, 3, 1–9. [Google Scholar] [CrossRef]
- Cai, F.; Mi, N.; Ming, H. Responses of dry matter accumulation and partitioning to drought and subsequent rewatering at different growth stages of maize in Northeast China. Front. Plant Sci. 2023, 14, 1110727. [Google Scholar] [CrossRef]
- Xu, C.L.; Li, R.D.; Song, W.W. High density and uniform plant distribution improve soybean yield by regulating population uniformity and canopy light interception. Agronomy 2021, 11, 1880. [Google Scholar] [CrossRef]
- Zhang, D.S.; Sun, Z.X.; Feng, L.S. Maize plant density affects yield, growth and source-sink relationship of crops in maize/peanut intercropping. Field Crops Res. 2020, 257, 107926. [Google Scholar] [CrossRef]
- Shi, D.Y.; Li, Y.H.; Zhang, J.W.; Liu, P.; Zhao, B.; Dong, S.T. Increased plant density and reduced N rate lead to more grain yield and higher resource utilization in summer maize. J. Integr. Agric. 2016, 15, 2515–2528. [Google Scholar] [CrossRef]
- Zheng, B.; Zhao, W.; Ren, T. Low Light Increases the Abundance of Light Reaction Proteins: Proteomics Analysis of Maize (Zea mays L.) Grown High Planting Density. Int. J. Mol. Sci. 2022, 23, 3015. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Jia, Q.; Li, J. Increased photosynthesis and grain yields in maize grown with less irrigation water combined with density adjustment in semiarid regions. PeerJ 2020, 8, 1–28. [Google Scholar]
- Wall, G.W.; Kanemasu, E.T. Carbon dioxide exchange rate in wheat canopies. part I, Influence of canopy geometry on trend in leaf area index, light interception and instantaneous exchange rates. Agric. For. Meteorol. 1990, 49, 81–102. [Google Scholar] [CrossRef]
- Watiki, J.M.; Fukai, S.; Banda, J.A. Radiation interception and growth of maize/cowpea intercrop as affected by maize plant density and cowpea cultivars. Field Crops Res. 1993, 35, 123–133. [Google Scholar] [CrossRef]
- Bai, Y.; Yang, Y.; Zhu, Y. Effect of planting density on light interception within canopy and grain yield of different plant types of maize. Acta Agron. Sin. 2019, 45, 1868–1879. [Google Scholar]
- Chen, C.Y.; Hou, H.P.; Li, Q.; Zhu, P.; Zhang, Z.Y.; Dong, Z.Q.; Zhao, M. Effects of panting density on photosynthetic characteristics and changes of carbon and nitrogen in leaf of different corn hybrids. Acta Agron. Sin. 2010, 36, 871–878. [Google Scholar] [CrossRef]
- Gao, J.; Lei, M.; Yang, L.J. Reduced row spacing improved yield by optimizing root distribution in maize. Eur. J. Agron. 2021, 127, 126291. [Google Scholar] [CrossRef]
- Liu, G.Z.; Liu, W.M.; Hou, P.; Ming, B.; Yang, Y.S.; Guo, X.X.; Xie, R.Z.; Wang, K.R.; Li, S.K. Reducing maize yield gap by matching plant density and solar radiation. J. Integr. Agric. 2021, 20, 363–370. [Google Scholar] [CrossRef]
- Varlet-Grancher, C.; Gosse, G.; Chartier, M. Solar radiation absorbed or intercepted by a crop. Agronomie 1989, 9, 419–439. [Google Scholar] [CrossRef]
- Tian, P.; Liu, J.; Zhao, Y. Nitrogen rates and plant density interactions enhance radiation interception, yield, and nitrogen use efficiencies of maize. Front. Plant Sci. 2022, 13, 974714. [Google Scholar] [CrossRef]
- Durand, M.; Murchie, E.H.; Lindfors, A.V.; Urban, O.; Aphalo, P.J.; Robson, T.M. Diffuse solar radiation and canopy photosynthesis in a changing environment. Agric. For. Meteorol. 2021, 311, 108684. [Google Scholar] [CrossRef]
- Kar, G.; Kumar, A.; Mohapatra, S. Radiation utilization efficiency, nitrogen uptake and modeling crop growth and yield of rainfed rice under different nitrogen rates. J. Indian Soc. Soil Sci. 2014, 62, 108–117. [Google Scholar]
- Kiniry, J.R.; Simpson, C.E.; Schuber, A.M.; Reed, J.D. Peanut leaf area index, light interception, radiation use efficiency, and harvest index at three sites in texas. Field Crops Res. 2005, 91, 297–306. [Google Scholar] [CrossRef]
- Tollenaar, M. Physiological Basis of Genetic Improvement of Maize Hybrids in Ontario from 1959 to 1988. Crop Sci. 1991, 31, 119–124. [Google Scholar] [CrossRef]
- Maddonni, G.A.; Otegui, M.E. Intra-specific competition in maize: Early establishment of hierarchies among plants affects final kernel set. Field Crops Res. 2004, 85, 1–13. [Google Scholar] [CrossRef]
- Blessing, C.; Nhamo, M.; Rangarirai, M. The impact of plant density and spatial arrangement on light interception on cotton crop and seed cotton yield: An overview. J. Cotton Res. 2020, 3, 210–215. [Google Scholar]
- Mathieu, A.; Cournède, P.H.; Letort, V. A dynamic model of plant growth with interactions between development and functional mechanisms to study plant structural plasticity related to trophic competition. Ann. Bot. 2009, 103, 1173–1186. [Google Scholar] [CrossRef]
- Niinemets, U.; Keenan, T.F.; Hallik, L. A worldwide analysis of within-canopy variations in leaf structural, chemical and physiological traits across plant functional types. New Phytol. 2015, 205, 973–993. [Google Scholar] [CrossRef] [PubMed]
- Yan, P.; Chen, Y.Q.; Sui, P.; Vogel, A.; Zhang, X.P. Effect of maize plant morphology on the formation of apical kernels at different sowing dates and under different plant densities. Field Crops Res. 2018, 223, 83–92. [Google Scholar] [CrossRef]
- Azhar, J.N.; Ehsanzadeh, P. Evaluation of Interrelationship of Growth Indices and Grain Yield of Five Maize Hybrids under Two Irrigation Regimes in Isfahan. J. Sci. Technol. Agric. Nat. Resour. 2007, 41, 261–273. [Google Scholar]
- Huang, W.J.; Yang, Q.Y.; Pu, R.L.; Yang, S.Y. Estimation of nitrogen vertical distribution by bi-directional canopy reflectance in winter wheat. Sensors 2014, 14, 20347–20359. [Google Scholar] [CrossRef] [PubMed]
- Ying, J.; Lee, E.A.; Tollenaar, M. Response of maize leaf photosynthesis to low temperature during the grain-filling period. Field Crops Res. 2000, 68, 87–96. [Google Scholar] [CrossRef]
- Begna, S.H.; Hamilton, R.I.; Dwyer, L.M. Weed Biomass Production Response to Plant Spacing and Corn (Zea mays) Hybrids Differing in Canopy Architecture. Weed Technol. 2009, 15, 647–653. [Google Scholar] [CrossRef]
- Vargas, L.A.; Andersen, M.N.; Jensen, C.R.; Jørgensen, U. Estimation of leaf area index, light interception and biomass accumulation of Miscanthus sinensis ‘Goliath’ from radiation measurements. Biomass Bioenergy 2002, 22, 1–14. [Google Scholar] [CrossRef]
- Evers, J.B.; Vos, J.; Yin, X. Simulation of wheat growth and development based on organ-level photosynthesis and assimilate allocation. J. Exp. Bot. 2010, 61, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Tahiri, A.Z.; Anyoji, H.; Yasuda, H. Fixed and variable light extinction coefficients for estimating plant transpiration and soil evaporation under irrigated maize. Agric. Water Manag. 2006, 84, 186–192. [Google Scholar] [CrossRef]
- Kandel, T.P.; Elsgaard, L.; Andersen, M.N. Influence of harvest time and frequency on light interception and biomass yield of festulolium and tall fescue cultivated on a peatland. Eur. J. Agron. 2016, 81, 150–160. [Google Scholar] [CrossRef]
- Koetz, B.; Baret, F.; Poilvé, H.; Hill, J. Use of coupled canopy structure dynamic and radiative transfer models to estimate biophysical canopy characteristics. Remote Sens. Environ. 2005, 95, 115–124. [Google Scholar] [CrossRef]
- Osawa, A. Plant canopies: Their growth, form and function. For. Ecol. Manag. 1992, 49, 168–170. [Google Scholar] [CrossRef]
- Maddonni, G.A.; Cirilo, A.G.; Otegui, M.E. Row Width and Maize Grain Yield. Agron. J. 2006, 98, 1532–1543. [Google Scholar] [CrossRef]
- Meng, Q.F.; Hou, P.; Wu, L.Q.; Chen, X.P.; Cui, Z.L.; Zhang, F.S. Understanding production potentials and yields gaps in intensive maize production in China. Field Crops Res. 2013, 143, 91–97. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Growth Period | Treatment | Kernel Weight (g 100 Seed−1) | Grain Yield (kg·ha−1) | Biomass Yield (kg·ha−1) | HI (%) | PUE (kg·ha−1 mm−1) |
|---|---|---|---|---|---|---|
| 2021 | D1 | 31.2 d | 5497.2 d | 9247.6 d | 59.4 a | 18.8 d |
| D2 | 33.0 c | 6142.1 c | 12,383.0 c | 49.6 b | 21.0 c | |
| D3 | 37.9 a | 6996.9 b | 14,570.1 b | 48.0 c | 24.0 b | |
| D4 | 34.6 b | 9257.3 a | 20,937.6 a | 44.2 d | 31.7 a | |
| 2022 | D1 | 38.5 b | 7233.2 d | 14,519.2 d | 49.8 a | 15.8 d |
| D2 | 38.9 ab | 7718.4 c | 16,177.9 c | 47.7 b | 16.9 c | |
| D3 | 39.8 a | 9204.6 b | 20,263.0 b | 45.4 c | 20.2 b | |
| D4 | 36.7 c | 9599.1 a | 21,771.8 a | 44.1 d | 21.0 a |
| Growth Period | Treatment | LAI (m2 m−2) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| 2021 | 2022 | ||||||||
| Upper Layer of Canopy | Middle Layer of Canopy | Lower Layer of Canopy | Sum | Upper Layer of Canopy | Middle Layer of Canopy | Lower Layer of Canopy | Sum | ||
| Jointing stage | D1 | 0.57 b | 0.79 b | 0.27 b | 1.64 | 0.37 b | 0.52 c | 0.14 b | 1.02 |
| D2 | 0.77 b | 0.98 ab | 0.42 ab | 2.17 | 0.69 ab | 0.71 bc | 0.24 ab | 1.65 | |
| D3 | 0.79 b | 1.39 ab | 0.42 ab | 2.61 | 0.80 ab | 0.90 b | 0.25 ab | 1.95 | |
| D4 | 2.93 a | 1.62 a | 0.77 a | 5.33 | 0.91 a | 1.32 a | 0.45 a | 2.68 | |
| Tasseling stage | D1 | 0.64 b | 1.35 d | 0.62 b | 2.60 | 0.82 c | 1.33 c | 0.48 b | 2.62 |
| D2 | 0.83 b | 1.80 c | 0.76 b | 3.40 | 1.11 bc | 1.57 bc | 0.64 ab | 3.32 | |
| D3 | 1.03 b | 2.14 b | 1.08 b | 4.25 | 1.40 ab | 2.15 b | 0.83 ab | 4.38 | |
| D4 | 5.36 a | 3.84 a | 1.92 a | 11.12 | 1.80 a | 3.05 a | 1.11 a | 5.97 | |
| Grain-filling stage | D1 | 0.53 b | 1.49 c | 0.75 c | 2.77 | 0.69 c | 1.25 d | 0.69 b | 2.63 |
| D2 | 0.71 b | 1.80 bc | 0.89 bc | 3.40 | 0.72 c | 1.82 c | 0.99 b | 3.53 | |
| D3 | 1.04 b | 2.02 b | 1.09 b | 4.16 | 1.16 b | 2.37 b | 0.97 b | 4.50 | |
| D4 | 3.41 a | 3.43 a | 1.69 a | 8.54 | 1.47 a | 2.91 a | 1.63 a | 6.02 | |
| Maturity stage | D1 | 0.39 b | 0.73 c | 0.87 b | 1.99 | 0.74 a | 0.83 c | 0.66 b | 2.24 |
| D2 | 0.47 b | 0.96 bc | 1.04 b | 2.47 | 0.63 a | 1.22 bc | 1.03 ab | 2.88 | |
| D3 | 1.04 a | 1.25 b | 1.38 b | 3.68 | 0.97 a | 1.47 ab | 1.28 ab | 3.72 | |
| D4 | 1.13 a | 1.92 a | 2.47 a | 5.52 | 0.92 a | 1.89 a | 1.45 a | 4.26 | |
| Growth Period | Treatment | K | |||||||
|---|---|---|---|---|---|---|---|---|---|
| 2021 | 2022 | ||||||||
| Lower Layer of Canopy | Middle Layer of Canopy | Upper Layer of Canopy | Average | Lower Layer of Canopy | Middle Layer of Canopy | Upper Layer of Canopy | Average | ||
| Jointing stage | D1 | 4.85 a | 2.13 a | 9.16 a | 5.38 | 3.02 a | 2.98 a | 7.89 a | 4.63 |
| D2 | 4.25 b | 1.59 b | 9.07 a | 4.97 | 2.68 b | 2.03 b | 7.97 a | 4.22 | |
| D3 | 4.23 b | 2.00 a | 6.70 b | 4.31 | 2.26 b | 1.44 c | 4.69 b | 2.80 | |
| D4 | 1.94 c | 0.85 c | 3.03 c | 1.94 | 1.70 c | 1.30 c | 2.43 c | 1.81 | |
| Tasseling stage | D1 | 4.23 b | 0.82 a | 8.38 a | 4.48 | 2.99 b | 1.44 a | 6.18 a | 3.54 |
| D2 | 6.79 a | 0.77 b | 3.11 b | 3.56 | 3.64 a | 1.08 b | 4.36 b | 3.03 | |
| D3 | 3.35 c | 0.60 c | 2.54 c | 2.16 | 2.65 b | 0.90 bc | 2.23 c | 1.93 | |
| D4 | 1.15 d | 0.72 b | 0.89 d | 0.92 | 1.93 c | 0.87 c | 0.94 c | 1.25 | |
| Grain-filling stage | D1 | 2.11 b | 1.06 a | 2.52 a | 1.90 | 1.82 b | 0.67 a | 2.50 a | 1.67 |
| D2 | 2.96 a | 0.82 b | 1.48 b | 1.75 | 2.25 a | 0.41 b | 2.00 a | 1.55 | |
| D3 | 2.15 b | 0.71 b | 0.76 c | 1.21 | 2.30 a | 0.37 b | 1.24 b | 1.30 | |
| D4 | 1.24 c | 0.59 b | 0.47 c | 0.77 | 1.95 b | 0.38 b | 1.26 b | 1.20 | |
| Maturity stage | D1 | 4.87 a | 1.13 a | 3.15 a | 3.05 | 5.99 a | 0.82 bc | 4.13 a | 3.65 |
| D2 | 3.80 b | 1.20 a | 1.41 b | 2.14 | 3.54 b | 1.09 a | 3.65 b | 2.76 | |
| D3 | 3.68 b | 0.90 b | 1.78 b | 2.12 | 2.87 c | 0.87 b | 2.11 c | 1.95 | |
| D4 | 1.72 c | 0.54 c | 0.86 c | 1.04 | 2.94 bc | 0.76 c | 1.18 d | 1.63 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duan, M.; Zhang, X.; Wei, Z.; Chen, X.; Zhang, B. Effect of Maize Canopy Structure on Light Interception and Radiation Use Efficiency at Different Canopy Layers. Agronomy 2024, 14, 1511. https://doi.org/10.3390/agronomy14071511
Duan M, Zhang X, Wei Z, Chen X, Zhang B. Effect of Maize Canopy Structure on Light Interception and Radiation Use Efficiency at Different Canopy Layers. Agronomy. 2024; 14(7):1511. https://doi.org/10.3390/agronomy14071511
Chicago/Turabian StyleDuan, Meng, Xiaotao Zhang, Zheng Wei, Xu Chen, and Baozhong Zhang. 2024. "Effect of Maize Canopy Structure on Light Interception and Radiation Use Efficiency at Different Canopy Layers" Agronomy 14, no. 7: 1511. https://doi.org/10.3390/agronomy14071511
APA StyleDuan, M., Zhang, X., Wei, Z., Chen, X., & Zhang, B. (2024). Effect of Maize Canopy Structure on Light Interception and Radiation Use Efficiency at Different Canopy Layers. Agronomy, 14(7), 1511. https://doi.org/10.3390/agronomy14071511