Abstract
Cucumber plants (Cucumis sativus) displaying symptoms of phyllody, sterility, and flower virescence were observed in fields located in the Dianbai district of Guangdong province, China. Total DNA was extracted from the symptomatic plants; this was followed by molecular detection using a set of universal phytoplasma primer pairs, R16mF2/mR1, P1/P7, and SecYF1(II)/SecYR1(II). This resulted in the PCR amplification of products corresponding to expected sizes of 1.4 kb, 1.8 kb and 1.7 kb, respectively. The 16S rDNA sequence obtained exhibited 100% similarity with the eggplant phyllody phytoplasma, the ‘Cleome rutidosperma’ witches’ broom phytoplasma, and the ‘Desmodium ovalifolium’ witches’ broom phytoplasma strain DeOWB, all of which belong to the 16SrII group. Phylogenetic analysis, based on the 16S rDNA gene and SecY gene sequences, confirmed the close affiliation of the detected phytoplasma isolate, tentatively designated as cucumber phyllody phytoplasma (CuPh) China isolate, with the 16SrII-A subgroup. Additionally, virtual restriction fragment length polymorphism (RFLP) analysis of the 16S rDNA sequence revealed a pattern that was identical to that of the 16SrII-A subgroup. This is the first report of cucumber phyllody phytoplasma in China.
1. Introduction
Phytoplasmas are unique, non-helical prokaryotes that lack a cell wall and are known to be uncultivable outside of their hosts. They are characterized by small chromosome sizes, which vary from 500 to 1000 kilobases (kb) [1,2]. Phytoplasmas specifically colonize the phloem sieve elements of plants and are primarily transmitted by insects that feed on the phloem, such as leafhoppers, planthoppers, and psyllids [3,4,5]. In addition to insect vectors, phytoplasmas can also be transmitted through dodder (a type of parasitic plant) and via plant propagation methods [6,7,8,9]. In certain instances, seed transmission of phytoplasmas has been observed [10,11,12,13,14]. Phytoplasma infection in plants can result in a wide array of symptoms, including witches’ broom, phyllody, yellowing, virescence (greenish-white discoloration), little leaf, dwarfism, sterility of flowers, and purple tops [2,6,11,15].
Phytoplasmas are a diverse group of pathogens capable of infecting over 1000 different plant species, encompassing crop plants, fruit trees, ornamental plants, oil plants, and weeds [6,15,16,17]. Cucurbit crops are also vulnerable to these infections, with examples such as Lagenaria siceraria (bottle gourd), Cucurbita moschata (pumpkin), Momordica charantia (bitter gourd), and Cucumis sativus (cucumber) facing threats from phytoplasma diseases [18,19,20,21,22]. Phytoplasma-induced diseases in cucurbits have been documented globally, manifesting in various forms such as cucumber phyllody, cucumber flat stem, bitter melon witches’ broom, snake gourd witches’ broom, and pumpkin witches’ broom [19,22,23,24,25].
Cucumber, an annual climbing plant belonging to the Cucurbitaceae family, is prized for its fruit, which is rich in vitamins, amino acids, fiber, and minerals [26]. Cucumbers are not only consumed as a vegetable and fruit but are also widely used in skincare and health products [27]. China is the world’s largest cucumber producer, with a vast planting area of 1,309,422 hectares and an annual production reaching 77,258,256 metric tons (FAOSTAT, 2022, http://faostat.fao.org/, accessed on 1 May 2024). Phytoplasma infections are known to induce symptoms such as dwarfing, witches’ broom, phyllody, and yellowing in cucumber across several countries, including Iran, India and Turkey [22,28,29,30]. In China, the Candidatus phytoplasma asteris strain has been reported to cause symptoms of flat stem and witches’ broom in cucumber [23]. However, phytoplasma causing the phyllody symptom in cucumber has not been reported in China until now.
Guangdong province, situated in the south of China, boasts a tropical and subtropical climate. One benefit of this is that crop plants can be cultivated in Guangdong all year around. However, the hot and rainy climate also contributes to the frequent occurrence of various diseases. In the current study, cucumber plants exhibiting the typical phyllody symptom were observed within the Dianbai district of Guangdong province, China. To identify the causative agent, we employed PCR and sequencing techniques, followed by phylogenetic and RFLP analyses for classification. Our findings represent the first report of a phytoplasma associated with cucumber phyllody in China.
2. Materials and Methods
2.1. Sample Collection and Molecular Identification
In January 2024, cucumber plants exhibiting symptoms of phyllody, flower virescence, and sterility were identified in fields within the Dianbai district of Guangdong province, China (21°52′75′′ N, 111°16′37″ E). One asymptomatic and two symptomatic samples were collected and preserved in a −80 °C freezer for further analysis.
Total DNA was extracted from these samples using the CTAB (hexadecyltrimethyl-ammonium bromide) method [31]. PCR-based detection was performed using R16mF2/mR1 [32] and P1/P7 [33,34] primer pairs targeting the 16S-23S ribosomal gene, as well as the SecYF1(II)/SecYR1(II) [35] primer pair for the SecY gene. Thermal protocols for the amplification of both genes are 94 °C for 10 min, 35 cycles each of 94 °C for 30 s, 55 °C for 30 s and 72 °C for 1 min, and a final extension at 72 °C for 10 min. The DNA of the asymptomatic cucumber was used as the negative control in PCR detection. PCR products were analyzed by electrophoresis on a 1% agarose gel, visualized under UV transilluminator (Bio-Rad, Hercules, CA, USA), and subsequently cloned into the pMD19T vector (Takara, Kusatsu, Japan). Transformation was carried out using Escherichia coli DH5α cells (Tsingke, Beijing, China). Confirmed clones were sequenced bidirectionally at Sangon Biotech Co. (Shanghai, China). The phytoplasma 16S group/subgroup classification and virtual RFLP (restriction fragment length polymorphism) analysis were conducted using iPhyClassifier (https://plantpathology.ba.ars.usda.gov/cgi-bin/resource/iphyclassifier.cgi, accessed on 26 January 2024) [36], based on the 16S rDNA sequence obtained from amplification.
2.2. Sequence Analysis
DNAMAN 9.0 software was utilized to perform sequence assembly and alignment. The resultant full-length sequences of the 16S rDNA and SecY genes were analyzed using a BLAST search to identify homologous sequences within the database. MEGA 11 software was employed to construct phylogenetic trees via the neighbor-joining method using the maximum composite likelihood model with 1000 bootstrapped replications, in accordance with the methodology described previously [37].
3. Results
3.1. Field Survey and Molecular Detection
In January 2024, an unusual phenomenon was observed in several cucumber plants on a farm situated in the Dianbai district of Guangdong province, China. These plants exhibited a symptom known as phyllody, characterized by the presence of green leaves with flower-like structures (Figure 1A–C). Additionally, they displayed a reduction in leaf size (Figure 1A–C), which heightened the suspicion of phytoplasma infection. Infected plants were unable to bear fruit, and the yield reduced by 100% compared to healthy plants. The incidence of this phenomenon was about 1%.

Figure 1.
Phyllody symptoms of the cucumber plants and molecular detection by PCR. Cucumber plants infected by phytoplasma showed phyllody and flower virescence symptoms (A,B) compared with flowers in asymptomatic plants (C). PCR amplification of 16S rDNA gene from the DNA of cucumber phyllody samples using the primer pairs R16mF2/mR1 (1.4 kb products) (D) and P1/P7 (1.8 kb products) (E). Amplification of SecY gene with primer pair SecYF1(II)/SecYR1(II) (1.7 kb products) (F). M: 2000 bp DNA marker (Takara). S1: sample 1 collected from cucumber plant infected by phytoplasma; S2: sample 2; AS: sample collected from asymptomatic cucumber plants.
To identify the causative agent of the phyllody symptoms, two phytoplasma 16S rDNA universal primer pairs, R16mF2/mR1 and P1/P7, were employed for phytoplasma detection. The PCR results revealed the presence of 1.4 kb and 1.8 kb amplification products exclusively in the symptomatic samples, but not in the asymptomatic control (Figure 1D,E). To confirm the presence of phytoplasma, we employed a specific primer pair, SecYF1(II)/SecYR1(II) [35], targeting the SecY gene. This analysis also yielded a positive result in the symptomatic samples, with no amplification in the asymptomatic sample (Figure 1F). Collectively, these findings strongly suggest that the abnormal growth observed in the cucumber plants was due to phytoplasma infection.
3.2. Phylogenetic Analysis
The phytoplasma responsible for the cucumber disease symptoms observed in China has been tentatively designated as cucumber phyllody phytoplasma (CuPh) China isolate. The nucleotide sequences of both the 16S rDNA and SecY genes were submitted to the National Center for Biotechnology Information (NCBI) to obtain the GenBank accession numbers PP479668 and PP498984, respectively.
The nucleotide sequences of the 16S rDNA and SecY genes of CuPh China isolate were then subjected to a comprehensive search against the NCBI database using the Basic Local Alignment Search Tool 2.15.0 (BLAST). The BLAST analysis revealed that the 16S rDNA sequence (GenBank accession PP479668) of CuPh China isolate shared 100% identity with several phytoplasma strains, including eggplant phyllody phytoplasma (EPP) HZQZ1 (GenBank accession MH667642), ‘Cleome rutidosperma’ witches’ broom phytoplasma HN-Cr1 (GenBank accession OP875099), soybean witches’ broom phytoplasma s3 (GenBank accession MW393690), and ‘Desmodium ovalifolium’ witches’ broom phytoplasma strain DeOWB (GenBank accession GU113152). Furthermore, the BLAST search utilizing the SecY nucleotide sequence (GenBank accession PP498984) demonstrated 100% similarity with two isolates of Candidatus Phytoplasma australasiaticum (WF_GM2021, GenBank accession CP133702, and NCHU2022, GenBank accession CP097312).
The phylogenetic analysis of 16S rDNA gene revealed that CuPh China isolate is clustered into the 16SrII-A subgroup (Figure 2). In addition, the phylogenetic tree based on the SecY gene sequences (Table S1) also positioned the CuPh China isolate on the same branch as peanut witches’ broom phytoplasma (GU004331) (Figure S1). These findings collectively suggest that the CuPh China isolate is a member of the 16SrII-A subgroup of phytoplasmas.
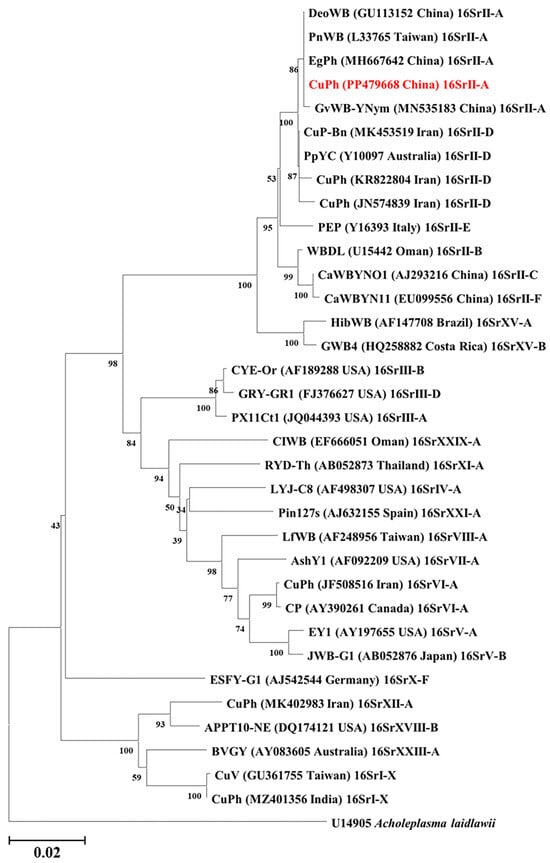
Figure 2.
Phylogenetic tree constructed from the full-length nucleotide sequences of 16S rDNA. The full name, GenBank accession number, the subgroup, and the origins of the phytoplasma strains or isolates used in this analysis are listed in Table 1. Acholeplasma laidlawii (U14905) was used as the outgroup. The cucumber phyllody phytoplasma China isolate identified in this study is marked in red. The phylogenetic tree was constructed with MEGA 11 software using the neighbor-joining method with 1000 bootstrapped replications. Branch lengths are measured using the relative number of substitutions per site, and bootstraps are shown on the main branches. Bars indicate two nucleotide substitutions per 100 nucleotides.
The iPhyClassifier, an online research tool, was used to simulate laboratory restriction enzyme digestion experiments and generate visual RFLP patterns. The computer-simulated RFLP analysis of the 16S rDNA sequence from CuPh China isolate exhibited a perfect match (with a similarity coefficient of 1.00) with the reference pattern of the 16SrII-A subgroup (GenBank accession L33765) (Figure S2), which further demonstrates that the CuPh China isolate belongs to the 16SrII-A subgroup.
4. Discussion
4.1. Phytoplasma Disease and Molecular Detection
Plants infected by phytoplasma display a multitude of symptoms, such as stunted growth, virescence, reduced internode length, big buds, little leaf, witches’ broom, phyllody, enlarged calyces, floral abnormalities, and vascular discoloration [2,15,38]. These infections can result in substantial yield reductions for crops like potatoes (with losses ranging from 30% to 80% [39,40]) tomatoes (with up to 60% loss [41]), eggplant (with a range of 40% to 100% [42,43]), sesame (with a range of 30% to 80% [44,45]), and cucumbers (with a potential 100% loss [23]), thereby affecting agricultural productivity globally.
PCR amplification of conserved genes has become a pivotal technique in the detection and classification of phytoplasmas, offering a significant advantage over traditional methods such as electron microscopy, histochemical staining, and serological detection. By employing universal primers that target the conserved regions of genes like 16S rDNA, SecY, SecA, Tuf, and rp [2] followed by sequencing, the classification of phytoplasmas can be efficiently achieved. The International Research Project for Comparative Mycoplasmology (IRPCM) Phytoplasma/Spiroplasma Working Team has recently updated the guidelines for phytoplasma classification in 2022 [46]. These refined guidelines recommend that the 16S rDNA sequence used for classification ideally spans between 1200 to 1500 base pairs. In the current study, we utilized specific primer pairs, such as R16mF2/mR1 [32] and P1/P7 [33,34] to amplify the 16S rDNA gene and the SecYF1(II)/SecYR1(II) [35] primer pair to amplify the SecY gene. The resulting PCR products were approximately 1.4 kb, 1.8 kb, and 1.7 kb in length, respectively. After sequencing, we obtained nucleotide sequences for the 16S rDNA (GenBank accession PP479668) and SecY genes (GenBank accession PP498984). Phylogenetic analysis of these sequences revealed that the phytoplasma responsible for cucumber phyllody in the Dianbai district is a member of the 16SII-A subgroup. RFLP pattern analysis conducted by the iPhyClassifier [36] also confirmed this result.
4.2. Phytoplasmas in China
In cucumber, phytoplasmas belonging to various groups and subgroups have been documented in several countries and districts. For instance, the cucumber phyllody phytoplasma has been identified in Iran as belonging to the 16SrII m subgroup [21,22], while in India, it falls into the 16SrI-X subgroup [29]. Iran has also reported the cucumber regional yellowing phytoplasma within the 16SrVI-A subgroup [28]. In China, the cucumber flat stem and witches’ broom phytoplasma is classified under the 16SrI subgroup [23], and the cucumber virescence phytoplasma in Taiwan is similarly placed in the 16SrI subgroup [47]. Our study is the first report of cucumber phyllody caused by a distinct phytoplasma from the 16SrII-A subgroup in China.
Reports indicate that over 100 phytoplasma diseases have been identified in China, affecting a wide array of plants. They are associated with phytoplasmas from 11 distinct phytoplasma groups: 16SrI, 16SrII, 16SrIII, 16SrV, 16SrVI, 16SrVII, 16SrVIII, 16SrX, 16SrXI, 16SrXII, and 16SrXIV. Among these, the 16SrI group has the broadest host range, with 16SrII being the next most diverse in terms of plant hosts [48,49,50]. Phytoplasmas which belong to the 16SrII group are mainly distributed in southern parts of China and can infect cowpea, areca palm, cinnamon, and other plants, causing certain economic losses [49,51,52,53,54,55,56,57]. It is noteworthy that similar phytoplasmas can trigger a variety of symptoms across different plant species, while distinct phytoplasmas may elicit similar responses. For instance, tomato big bud disease has been linked to multiple phytoplasma groups, including 16SrI, 16SrII, 16SrV, 16SrVI, and 16SrXII [39,58,59,60]. Conversely, the 16SrII-D group of phytoplasmas has been identified as a culprit in Oman, affecting multiple crops and causing symptoms such as phyllody, virescence, and big bud [61].
4.3. Prospects
Phytoplasma in cucumber can result in floral malformations, phyllody, and sterility, leading to significant crop losses. Given that China is the world’s largest producer of cucumbers, it is imperative that we conduct further research into the epidemiology of this disease, identify potential natural sources of its spread, and develop effective management strategies to mitigate its impact. The most effective strategy for preventing phytoplasma diseases may involve disrupting their transmission pathways. This includes eliminating weeds that could serve as phytoplasma reservoirs, controlling insect vectors such as leafhoppers, and disinfecting the tools used for grafting and pruning. Given the transmission characteristics of phytoplasmas, it is necessary to adhere to a policy that prioritizes prevention and integrates it with control measures.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/agronomy14081873/s1, Table S1. SecY gene sequences of phytoplasmas employed in this analysis. Figure S1. Phylogenetic tree constructed from the full-length nucleotide sequences of SecY. Phytoplasma strains or isolates used in this analysis are listed in Table S1. Figure S2. Virtual RFLP patterns from in silico digestion of the 16S rDNA sequence.
Author Contributions
Z.L. and Z.H. conceived and designed the experiments. Y.X., Z.L., Y.T. and L.Y. conducted the experiments. M.D., X.S., G.L. and S.D. analyzed the data. Y.X., Z.L. and Z.H. wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (32272509), the Science and Technology Program of Guangzhou (2024A04J7051), the Young Talent Support Project of Guangzhou Association for Science and Technology (QT20230101154), and the Special Fund for Scientific Innovation Strategy—Construction of High-Level Academy of Agriculture Science (R2023PY-JX011).
Data Availability Statement
Data are contained within the article and Supplementary Materials.
Conflicts of Interest
The authors declare that this research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Wei, W.; Zhao, Y. Phytoplasma Taxonomy: Nomenclature, Classification, and Identification. Biology 2022, 11, 1119. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Bai, B.; Li, D.; Wang, J.; Huang, W.; Wu, Y.; Zhao, L. Phytoplasma: A plant pathogen that cannot be ignored in agricultural production—Research progress and outlook. Mol. Plant Pathol. 2024, 25, e13437. [Google Scholar] [CrossRef]
- Hogenhout, S.A.; Oshima, K.; Ammar el, D.; Kakizawa, S.; Kingdom, H.N.; Namba, S. Phytoplasmas: Bacteria that manipulate plants and insects. Mol. Plant Pathol. 2008, 9, 403–423. [Google Scholar] [CrossRef]
- Koinuma, H.; Maejima, K.; Tokuda, R.; Kitazawa, Y.; Nijo, T.; Wei, W.; Kumita, K.; Miyazaki, A.; Namba, S.; Yamaji, Y. Spatiotemporal dynamics and quantitative analysis of phytoplasmas in insect vectors. Sci. Rep. 2020, 10, 4291. [Google Scholar] [CrossRef]
- Rashidi, M.; D’Amelio, R.; Galetto, L.; Marzachì, C.; Bosco, D. Interactive transmission of two phytoplasmas by the vector insect. Ann. Appl. Biol. 2014, 165, 404–413. [Google Scholar] [CrossRef]
- Bertaccini, A. Phytoplasmas: Diversity, taxonomy, and epidemiology. Front. Biosci. A J. Virtual Libr. 2007, 12, 673–689. [Google Scholar] [CrossRef]
- Akhtar, K.P.; Sarwar, G.R.; Dickinson, M.J.; Ahmad, M.; Haq, M.A.; Hameed, S.; Iqbal, M.J. Sesame phyllody disease: Its symptomatology, etiology, and transmission in Pakistan. Turk. J. Agric. For. 2009, 33, 477–486. [Google Scholar] [CrossRef]
- Přibylová, J.; Spak, J. Dodder transmission of phytoplasmas. Methods Mol. Biol. 2013, 938, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.-C.; Chen, J.-C. An efficient grafting method for phytoplasma transmission in Catharanthus roseus. Plant Methods 2024, 20, 13. [Google Scholar] [CrossRef] [PubMed]
- Mateeti, S.; Checchi, G.; Messina, N.; Feduzi, G.; Bertaccini, A.; Contaldo, N. Presence and seed transmission of phytoplasmas in tomato fields in Italy. Phytopathog. Mollicutes 2022, 12, 1–6. [Google Scholar] [CrossRef]
- Kirdat, K.; Tiwarekar, B.; Sathe, S.; Yadav, A. From sequences to species: Charting the phytoplasma classification and taxonomy in the era of taxogenomics. Front. Microbiol. 2023, 14, 1123783. [Google Scholar] [CrossRef] [PubMed]
- Kirdat, K.; Tiwarekar, B.; Swetha, P.; Padma, S.; Thorat, V.; Manjula, K.N.; Kavya, N.; Sundararaj, R.; Yadav, A. Nested real-time PCR assessment of vertical transmission of sandalwood spike phytoplasma (‘Ca. phytoplasma asteris’). Biology 2022, 11, 1494. [Google Scholar] [CrossRef] [PubMed]
- Priya, M.; Yadav, A.; Thorat, V.; Rao, G. Molecular detection of 16SrI-B and 16SrII-D subgroups of phytoplasma associated with flat stem and witches’ broom disease of Celosia argentea L. 3 Biotech 2017, 7, 311. [Google Scholar] [CrossRef]
- Contaldo, N.; D’Amico, G.; Paltrinieri, S.; Diallo, H.A.; Bertaccini, A.; Arocha Rosete, Y. Molecular and biological characterization of phytoplasmas from coconut palms affected by the lethal yellowing disease in Africa. Microbiol. Res. 2019, 223–225, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Asudi, G.O.; Omenge, K.M.; Paulmann, M.K.; Reichelt, M.; Grabe, V.; Mithöfer, A.; Oelmüller, R.; Furch, A.C.U. The physiological and biochemical effects on napier grass plants following napier grass stunt phytoplasma infection. Phytopathology 2021, 111, 703–712. [Google Scholar] [CrossRef]
- Gurr, G.M.; Johnson, A.C.; Ash, G.J.; Wilson, B.A.L.; Ero, M.M.; Pilotti, C.A.; Dewhurst, C.F.; You, M.S. Coconut lethal yellowing diseases: A phytoplasma threat to palms of global economic and social significance. Front. Plant Sci. 2016, 7, 1521. [Google Scholar] [CrossRef] [PubMed]
- Maejima, K.; Oshima, K.; Namba, S. Exploring the phytoplasmas, plant pathogenic bacteria. J. Gen. Plant Pathol. 2014, 80, 210–221. [Google Scholar] [CrossRef]
- Ashwathappa, K.V.; Reddy, L.R.C.N.; Venkataravanappa, V.; Sastry, S.V.; Reddy, M.G.; Rao, G.P. A new strain of ‘Candidatus Phytoplasma asteris’ associated with bottle gourd phyllody disease in South India. Phytopathog. Mollicutes 2020, 10, 43–49. [Google Scholar] [CrossRef]
- Wang, X.; Wang, C.-g.; Li, X.-Y.; Li, Z. Molecular detection and identification of a ‘Candidatus Phytoplasma solani’-related strain associated with pumpkin witches’ broom in Xinjiang, China. Phytopathol. Mediterr. 2021, 60, 63–68. [Google Scholar] [CrossRef]
- Venkataravanappa, V.; Reddy, L.; Polam, S.; Subbanna, S.K.; Manem, K.R. Detection and characterization of ‘Candidatus Phytoplasma asteris’ associated with little leaf disease of bitter gourd from India by 16S rRNA phylogenetic and RFLP (in vitro and virtual) analysis. Arch. Biol. Sci. 2017, 69, 707–714. [Google Scholar] [CrossRef]
- Dehghan, H.; Salehi, M.; Khanchezar, A.; Afshar, H. Biological and molecular characterization of a phytoplasma associated with greenhouse cucumber phyllody in Fars province. Iran. J. Plant Pathol. 2015, 50, 393–401. [Google Scholar]
- Salehi, M.; Siampour, M.; Hosseini, S.A.E.; Bertaccini, A. Characterization and vector identification of phytoplasmas associated with cucumber and squash phyllody in Iran. Bull. Insectol. 2015, 68, 311–319. [Google Scholar]
- Wang, X.; Hu, Q.; Wang, J.; Lou, L.; Xu, X.; Chen, X. Comparative biochemical and transcriptomic analyses provide new insights into phytoplasma infection responses in cucumber. Genes 2022, 13, 1903. [Google Scholar] [CrossRef]
- Alves, M.S.; Ribeiro, G.M.; Souza, A.N.; Carvalho, C.M. First report of a 16SrIII-B phytoplasma associated with Momordica charantia witches’-broom in Brazil. Plant Dis. 2017, 101, 1314. [Google Scholar] [CrossRef]
- Weng, Y.-Y.; Liou, W.-C.; Chien, Y.-Y.; Liao, P.-Q.; Wang, C.-J.; Chiu, Y.-C.; Chen, Y.-K.; Yang, J.-Y. First report of 16SrII-V peanut witches’ broom phytoplasma in snake gourd (Trichosanthes cucumerina) in Taiwan. Plant Dis. 2021, 105, 2236. [Google Scholar] [CrossRef] [PubMed]
- Qi, J.; Liu, X.; Shen, D.; Miao, H.; Xie, B.; Li, X.; Zeng, P.; Wang, S.; Shang, Y.; Gu, X.; et al. A genomic variation map provides insights into the genetic basis of cucumber domestication and diversity. Nat. Genet. 2013, 45, 1510–1515. [Google Scholar] [CrossRef]
- Li, C.; Ma, H.; Li, P.; Zhang, S.; Xu, J.; Wang, L.; Sheng, W.; Xu, T.; Shen, L.; Wang, W.; et al. Cucumber (Cucumis sativus L.) with heterologous poly-γ-glutamic acid has skin moisturizing, whitening and anti-wrinkle effects. Int. J. Biol. Macromol. 2024, 262, 130026. [Google Scholar] [CrossRef] [PubMed]
- Ghayeb Zamharir, M.; Azimi, H. Detection and characterisation of a phytoplasma associated with cucumber (Cucumis sativus) regional yellows disease in Iran. Arch. Phytopathol. Plant Prot. 2018, 51, 889–893. [Google Scholar] [CrossRef]
- Muttappagol, M.; Kumar, H.D.V.; Hiremath, S.; Nandan, M.; Basha, C.R.J.; Shankarappa, K.S.; Venkataravanappa, V.; Reddy, C.N.L. Detection and molecular characterization of a phytoplasma associated with cucumber (Cucumis sativus) and its first report from India. bioRxiv 2022. [Google Scholar] [CrossRef]
- Usta, M.; Güller, A.; Sipahioglu, H. Detection and characterization of two phytoplasma lineages on cucumber (Cucumis sativus L.) with same symptomatology based on virtual RFLP and nucleotide sequence analysis of 16S rDNA. Yuz. Yil Univ. J. Agric. Sci. 2017, 27, 299–308. [Google Scholar]
- Doyle, J.; Doyle, J. Genomic plant DNA preparation from fresh tissue-CTAB method. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Gundersen, D.E.; Lee, I.-M. Ultrasensitive detection of phytoplasmas by nested-PCR assays using two universal primer pairs. Phytopathol. Mediterr. 1996, 35, 144–151. [Google Scholar]
- Deng, S.; Hiruki, C. Amplification of 16S rRNA genes from culturable and nonculturable Mollicutes. J. Microbiol. Methods 1991, 14, 53–61. [Google Scholar] [CrossRef]
- Smart, C.D.; Schneider, B.; Blomquist, C.L.; Guerra, L.J.; Harrison, N.A.; Ahrens, U.; Lorenz, K.H.; Seemuller, E.; Kirkpatrick, B.C. Phytoplasma-specific PCR primers based on sequences of the 16S-23S rRNA spacer region. Appl. Environ. Microbiol. 1996, 62, 2988–2993. [Google Scholar] [CrossRef]
- Lee, I.-M.; Bottner-Parker, K.D.; Zhao, Y.; Davis, R.E.; Harrison, N.A. Phylogenetic analysis and delineation of phytoplasmas based on SecY gene sequences. Int. J. Syst. Evol. Microbiol. 2010, 60, 2887–2897. [Google Scholar] [CrossRef]
- Zhao, Y.; Wei, W.; Lee, I.M.; Shao, J.; Suo, X.; Davis, R.E. Construction of an interactive online phytoplasma classification tool, iPhyClassifier, and its application in analysis of the peach X-disease phytoplasma group (16SrIII). Int. J. Syst. Evol. Microbiol. 2009, 59, 2582–2593. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Bertaccini, A.; Duduk, B.; Paltrinieri, S.; Contaldo, N. Phytoplasmas and phytoplasma diseases: A severe threat to agriculture. Am. J. Plant Sci. 2014, 5, 1763–1788. [Google Scholar] [CrossRef]
- Ember, I.; Acs, Z.; Munyaneza, J.E.; Crosslin, J.M.; Kolber, M. Survey and molecular detection of phytoplasmas associated with potato in Romania and southern Russia. Eur. J. Plant Pathol. 2011, 130, 367–377. [Google Scholar] [CrossRef]
- Castillo Carrillo, C.; Paltrinieri, S.; Bustamante, J.B.; Bertaccini, A. Detection and molecular characterization of a 16SrI-F phytoplasma in potato showing purple top disease in Ecuador. Australas. Plant Pathol. 2018, 47, 311–315. [Google Scholar] [CrossRef]
- Navrátil, M.; Válová, P.; Fialová, R.; Lauterer, P.; Šafářová, D.; Starý, M. The incidence of stolbur disease and associated yield losses in vegetable crops in South Moravia (Czech Republic). Crop Prot. 2009, 28, 898–904. [Google Scholar] [CrossRef]
- Rao, G.P.; Kumar, M. World status of phytoplasma diseases associated with eggplant. Crop Prot. 2017, 96, 22–29. [Google Scholar] [CrossRef]
- Rao, G.; Mall, S.; Raj, S.; Snehi, S. Phytoplasma diseases affecting various plant species in India. Acta Phytopathol. Entomol. Hung. 2011, 46, 59–99. [Google Scholar] [CrossRef]
- Salehi, M.; Esmailzadeh Hosseini, S.A.; Salehi, E.; Bertaccini, A. Genetic diversity and vector transmission of phytoplasmas associated with sesame phyllody in Iran. Folia Microbiol. 2017, 62, 99–109. [Google Scholar] [CrossRef]
- Ghulam Sarwar, G.S.; Akhtar, K. Performance of some sesame mutants to phyllody an leaf curl virus disease under natural field conditions. Pak. J. Phytopathol. 2009, 21, 18–25. [Google Scholar]
- Bertaccini, A.; Arocha-Rosete, Y.; Contaldo, N.; Duduk, B.; Fiore, N.; Montano, H.G.; Kube, M.; Kuo, C.-H.; Martini, M.; Oshima, K.; et al. Revision of the ‘Candidatus Phytoplasma’ species description guidelines. Int. J. Syst. Evol. Microbiol. 2022, 72, 005353. [Google Scholar] [CrossRef]
- Yang, J.-Y.; Chien, Y.-Y.; Chiu, Y.-C.; Mejia, H.M.; Tan, C.M. Diversity, distribution, and status of phytoplasma diseases in Taiwan. In Diversity, Distribution, and Current Status; Tiwari, A.K., Caglayan, K., Al-Sadi, A.M., Azadvar, M., Abeysinghe, S., Eds.; Academic Press: Cambridge, MA, USA, 2023; Volume 1, pp. 149–168. [Google Scholar]
- Lai, F.; Li, Y.; Xu, Q.C.; Tian, G.Z. The present status on classification of phytoplasmas. Microbiology 2008, 35, 291–295. [Google Scholar] [CrossRef]
- Wang, X.-Y.; Zhang, R.-Y.; Li, J.; Li, Y.-H.; Shan, H.-L.; Li, W.-F.; Huang, Y.-K. The diversity, distribution and status of phytoplasma diseases in China. Front. Sustain. Food Syst. 2022, 6, 943080. [Google Scholar] [CrossRef]
- Che, H.; Yu, S.; Chen, W.; Zheng, W.; Cao, X.; Luo, D. Molecular identification and characterization of novel taxonomic subgroups and new host plants in 16SrI and 16SrII group phytoplasmas and their evolutionary diversity on Hainan island, China. Plant Dis. 2024, 108, 1703–1718. [Google Scholar] [CrossRef]
- Yu, S.-S.; Song, W.-W. Ipomoea obscura represents a new host of phytoplasma belonging to 16SrII group associated with witches’ broom symptoms in China. Plant Dis. 2024, 108, 780. [Google Scholar] [CrossRef]
- Wang, B.; Lin, Z.W.; Zhao, Z.X.; Yan, W.R.; Tan, S.M.; Huang, W.K.; Ji, X.C. First report of 16SrII group related phytoplasma associated with witches’-broom disease on cowpea (Vigna unguiculata) in Hainan province, China. Plant Dis. 2024, 108, 783. [Google Scholar] [CrossRef]
- Li, Z.; Zhanbiao, L.; Xiaoman, S.; Farooq, T.; Zhenggang, L.; Lin, Y.; Guobing, L.; Zifu, H.; Tang, Y. First report of 16SrII-V phytoplasma associated with cowpea (Vigna unguiculata) phyllody in China. Can. J. Plant Pathol. 2023, 45, 103–109. [Google Scholar] [CrossRef]
- Che, H.; Li, Z.; Zhang, L.; Wu, Y.; Luo, D. Detection and identification of 16SrII group phytoplasmas infecting stylosanthes in China. J. Phytopathol. 2012, 160, 437–439. [Google Scholar] [CrossRef]
- Li, Y.; Chen, W. First report of a 16SrII-A phytoplasma infecting Spermacoce exilis in China. J. Plant Pathol. 2018, 100, 347. [Google Scholar] [CrossRef]
- Lin, Z.; Song, W.; Tang, Q.; Meng, X. First report of 16SrII group related phytoplasma associated with areca palm yellow leaf disease on Areca catechu in China. Plant Dis. 2023, 107, 3275. [Google Scholar] [CrossRef]
- Yang, Y.; Jiang, L.; Che, H.-Y.; Cao, X.-R.; Yang, J.-Y.; Sang, L.-W.; Liu, A.-Q.; Luo, D.-Q. Molecular identification of a 16SrII-A group-related phytoplasma associated with cinnamon yellow leaf disease in China. J. Phytopathol. 2015, 164, 52–55. [Google Scholar] [CrossRef]
- Gungoosingh-Bunwaree, A.; Bertaccini, A.; Benimadhu, S. Presence of phytoplasma infections in tomato plants in Mauritius. Bull. Insectol. 2007, 60, 151–152. [Google Scholar]
- Vali Sichani, F.; Bahar, M.; Zirak, L. Characterization of phytoplasmas related to aster yellows group infecting annual plants in Iran, based on the studies of 16S rRNA and rp Genes. J. Plant Prot. Res. 2014, 54, 1–8. [Google Scholar] [CrossRef]
- Madhupriya; Kumari, D.A.; Vennila, D.S.; Narayana Bhat, M.; Rao, G.P. Increasing incidence of tomato big bud phytoplasma in Ranga Reddy District of Telangana State, India. Indian Phytopathol. 2018, 71, 207–211. [Google Scholar] [CrossRef]
- Al-Subhi, A.M.; Hogenhout, S.A.; Al-Yahyai, R.A.; Al-Sadi, A.M. Detection, identification, and molecular characterization of the 16SrII-D phytoplasmas infecting vegetable and field crops in Oman. Plant Dis. 2018, 102, 576–588. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).