Abstract
Rice–Xanthomonas oryzae pv. oryzicola (Xoc) is one of the commonly used rice models of host–pathogen interactions. Xoc causes bacterial leaf streak (BLS) and has quarantine status. As a Gram-negative pathogen, Xoc usually employs type III secretion effectors (T3SEs), including transcription activator-like effectors (TALEs) and non-TALEs, to interfere with the innate immunity of rice. However, few major resistance genes corresponding to Xoc are found in rice cultivations; only Rxo1-AvrRxo1 and Xo1-TALEs interactions have been discovered in rice–Xoc. In this review, we focus on the role of the T3S system (T3SS) in Xoc virulence and consider the reported non-TALEs, including AvrRxo1, AvrBs2, XopN, XopC2, XopAP, and XopAK, as well as TALEs including Tal2g/Tal5d, Tal2h, Tal2a, Tal7, Tal10a, TalI, Tal2b, and Tal2c. Interestingly, AvrRxo1, XopC2, and XopAP disturb stomatal opening to promote infection through targeting diverse signaling pathways in rice. Otherwise, Tal2b and Tal2c, respectively, activate two rice salicylic acid (SA) hydroxylation genes to redundantly suppress the SA-mediated basal defense, and TalI, which has unknown targets, suppresses the SA signaling pathway in rice. In addition, other Xoc virulence factors are discussed. In conclusion, several T3SEs from Xoc interfere with similar defense pathways in rice to achieve invasion, providing an outlook for the control of this disease through manipulating the conserved pathways.
1. Introduction
Xanthomonas oryzae pv. oryzicola (Xoc) is the causal agent of bacterial leaf streak (BLS) in rice, which frequently occurs in rice planting areas, causing devastating damage to rice production [1]. Unlike the similar Xanthomonas oryzae pv. oryzae (Xoo) pathogen, which causes rice bacterial blight (BB), a limited number of resistance (R) genes are used to control BLS [2]. Xoc carries the type III secretion system (T3SS), which delivers a diversity of T3S effectors (T3SEs), including transcription activator-like effectors (TALEs) and non-TALEs, into host cells [3,4]. TALEs belong to the AvrBs3 family and are a type of specific effectors which usually have the ability to bind to host gene promoters and activate gene expression. TALEs have a typical domain of repeat-variable diresidue (RVD) with different numbers, which usually exhibit the ability to specifically recognize host gene promoters according to positions 12 and 13 of the RVDs sequence. Non-TALEs, also named Xanthomonas outer proteins (Xops), are a type of effectors that have various functions toward host targets to interfere with host defense responses [5,6]. The Xoc genome contains dozens of T3SEs; a wide diversity of TALEs are found in different Xoc strains [7,8,9]. The first identified non-TALE and TALE were AvrRxo1 and Tal2g, respectively. These proteins work as avirulence proteins that recognize the maize nonhost R protein encoded by Rxo1 and target the sulfate transporter gene OsSULTR3;6 to improve rice susceptibility [10,11]. Over the past decades, a series of investigations on rice–Xoc interactions have led to the identification of several T3SEs, whereas, in general, they still lack sufficient cognition of the virulence function of the T3SEs in Xoc. In this review, we discuss the virulence role of T3SS in Xoc and collect the reported TALEs and non-TALEs involved in rice–Xoc interactions, which are listed in Figure 1 and Table 1.
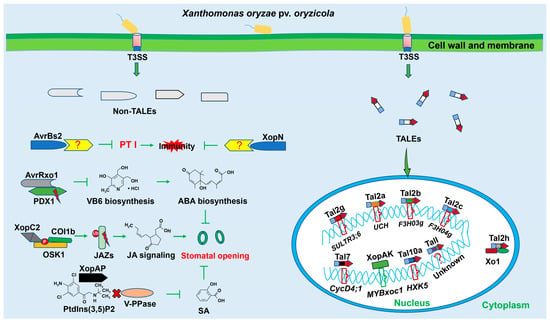
Figure 1.
Functions of T3SEs in rice–Xoc interactions. Xanthomonas oryzae pv. oryzicola (Xoc) employs the type III secretion system (T3SS) to deliver T3S effectors (T3SEs) into rice cells during invasion. One type of T3SE, transcription activator-like effectors (TALEs), has transcriptional activation ability and can enter into the cell nucleus to bind to effector-binding elements (EBEs) to activate host genes’ expressions. The models of TALE-Susceptible gene in rice–Xoc interactions are depicted, including Tal2gBLS256-SULTR3;6, Tal7RS105-CycD4;1, Tal2bHGA4-F3H03g, Tal2cHGA4-F3H04g, TalIGD41-Unknown, and Tal10aGX01-HXK5. Tal2aBLS256-ubiquitin carboxy-terminal hydrolase (UCH) is a model of TALE-resistant gene in rice–Xoc interactions; Tal2hBLS256-Xo1 is an abnormal case of TALE-resistant gene mediated by a direct protein–protein interaction. The other kind of T3SE, which has various functions, is called non-TALEs. The models of non-TALE-target interfere with stomatal immunity, including XopC2-Oryza sativa SKP1-like (OSK1)-OsCOI1b-JASMONATE ZIM-DOMAIN (JAZs)-jasmonic acid (JA) signaling, AvrRxo1-PDX1-vitamin B6 (VB6)-abscisic acid (ABA), XopAP-phosphatidylinositol-3,5-bisphosphate (PtdIns(3,5)P2)-vacuolar H+-pyrophosphatase (V-PPase)-salicylic acid (SA), and XopAK-MYBxoc1. AvrBs2 and XopN are virulence factors with unknown targets.
The T3SS-mediated interactions with hosts and Xanthomonas spp. have been thoroughly explored; however, other virulence factors, including extracellular polysaccharides (EPSs), diffusible signal molecules (DSFs), lipopolysaccharides (LPSs), and other virulence-associated secretion systems, also play important roles in pathogenicity and environmental fitness [12,13]. EPSs are involved in the formation of bacterial biofilms, which are essential for overcoming different environmental stresses [14,15]. Xanthan is one of the EPSs in Xanthomonas that helps the bacterium to survive under oxidative stress. EPSs have diverse roles in the pathogenicity of different Xanthomonas spp. [16,17,18]. DSF-mediated quorum sensing (QS) regulates bacterial biofilm agglomeration and distribution and also plays an important role in the pathogenicity of Xanthomonas [19,20]. LPS, which is composed of lipid A, a core oligosaccharide, and an O-antigen, is a component of the bacterial outer membrane and functions to protect the cells from unsuitable conditions [21]. LPS acts not only in virulence but also as a microbe-associated molecular pattern in Xanthomonas spp. [22,23]. Xanthomonas spp. usually contain six secretion systems (type I to VI), among which T3SS, T4SS, and T6SS assemble a needle-like structure to deliver bacterial effectors into host cells. T2SS translocates the cellular degradation enzymes to destroy the host–cell wall. However, our knowledge of T1SS and T5SS is limited [12,24,25,26]. To thoroughly promote our understanding of the virulence factors in Xoc, we summarized the roles of the DSF-mediated QS, EPS, LPS, regulatory systems, and molecules to determine their effect on the pathogenicity of Xoc, shown in Figure 2.
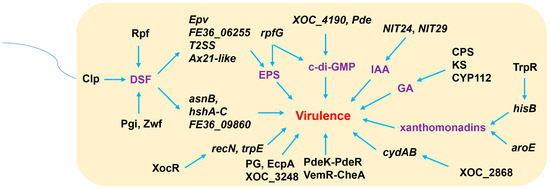
Figure 2.
The mechanisms of Xoc virulence. Xoc produces diffusible signal factors (DSFs), which are regulated by pathogenicity factor (Rpf), cyclic adenosine monophosphate receptor protein-like protein (Clp), glucose 6-phosphate isomerase (Pgi), and glucose-6-phosphate dehydrogenase (Zwf). DSFs improve the expressions of an extracellular polysaccharide (EPS)- and virulence-related gene (Epv), a TetR family of transcriptional regulator gene FE36_06255, type II secretion system (T2SS) genes, and an Ax21-like gene to regulate EPS production to facilitate the virulence of Xoc. Otherwise, DSFs also positively regulate an asparagine synthetase gene (asnB), virulence-associated genes hshA-C, and a TetR family of transcriptional regulator gene FE36_09860 to promote Xoc infection. Xoc also produces indole-3-acetic acid (IAA) by nitrilase enzyme family (NIT) genes NIT24 and NIT29, gibberellin (GA) by ent-copalyl diphosphate synthase (CPS), ent-kaurene synthase (KS), and cytochromes P450 (CYP112), c-di-GMP by a HD-GYP domain regulatory gene rpfG and GGDEF-EAL-domain genes XOC_4190 and pde, and xanthomonadins by aroE and histidine synthetase (his) gene hisB, which is positively regulated by a transcriptional regulator TrpR. These molecules are also involved in Xoc virulence. Two-component regulatory system (TCS) pairs PdeK-PdeR and VemR-CheA, a polygalacturonase PG, an extracellular protease EcpA, and an inner membrane protein XOC_3248 are involved in Xoc virulence. In addition, virulence-associated genes recN and trpE, for which their expression is positively regulated by plant-associated bacteria (PAB), LuxR protein XocR, and cytochrome bd oxidase operon cydAB, for which their expression is positively regulated by a transcriptional factor XOC_2868, are also involved in Xoc virulence.
2. Type III Secretion System Is Essential for Virulence of Xanthomonas oryzae pv. oryzicola
2.1. Type III Secretion System of Xoc
The T3SS is a Gram-negative bacterial secretion system that plays an important role in the interactions with hosts. The T3SS assembles a syringe-like apparatus to inject T3SEs into host cells [27,28]. Xanthomonas spp., which cause over 400 plant diseases, usually employ the T3SS to successfully invade into host plants [3,13]. The T3SSs of Xanthomonas spp. are encoded by a hypersensitive response and pathogenicity (hrp) and hrp-conserved (hrc) and hrp-associated (hpa) gene clusters [29,30,31]. A total of 10 hrp, 9 hrc, and 8 hpa genes are located in 27 kb hrp clusters, with a small cluster formed with hrpG and hrpX, which are conserved in different Xoc strains. hrpF is required for the pathogenicity of Xoc in rice. Additionally, Hpa2 can physically interact with HrpF to translocate the T3SEs into host cells, and the expression of hap2 is positively regulated by HrpG and HrpD6. The single mutant of hpa2 or hrpF partly impairs Xoc virulence, whereas both loss-of-function hpa2 and hrpF abolish the pathogenicity of Xoc in rice [32,33]. In addition to hpa2 expression, the expressions of hpa1, hpaB, hrcC, and hrcT are also positively regulated by HrpD6, which affects the secretion of T3SEs. The major roles of these genes in Xoc virulence in rice are also identified [4,34,35]. The expression of hrpE3, which, when encoding a T3SE protein, is positively regulated by HrpG and HrpX; hrpE3 is essential for Xoc achieving full virulence in rice [36]. In addition to hrp clusters, the plant-regulated hrp gene prhA also contributes to the full virulence of Xoc in rice [32].
The roles of HrpG, HrpX, and HrpD6 in the regulation of virulence functions, in addition to that of the T3SS, have also been reported. Xoryp_08180, which encodes a hypothetical protein, contributes to the full virulence of Xoc and is involved in extracellular protease production. The expression of Xoryp_08180 is also positively regulated by HrpG and HrpX [37]. fbaB and xanA, which encode a fructose–bisphophate aldolase and a phosphohexose mutase, respectively, are involved in carbon metabolism and are required for the full virulence of Xoc in rice and EPS production. The promoters of fbaB and xanA both contain a PIP-box, the expression of which is negatively regulated by HrpG and HrpX [38,39]. Five of the seventeen HrpX-regulated proteins (Xrps), Xrp1, Xrp2, Xrp5, Xrp8, and Xrp14, contribute to the pathogenicity of Xoc in rice. Of these five Xrp genes, the expression of xrp1, xrp2, and xrp5 is also negatively regulated by HrpD6 [40].
Several factors affect the T3SS and are involved in the pathogenicity of Xoc. wxocB is involved in LPS O-chain synthesis and is required for the full virulence of Xoc in rice. Additionally, the loss function of wxocB reduces type III secretion in Xoc [41,42]. A small regulatory noncoding RNA (sRNA) called Xonc3711 in Xoc directly binds the mRNA of a DNA-binding protein gene, xoc_3982, to suppress translation. Xoc_3982 targets the promoter of a T3SE gene, xopC2, which is a virulence factor of Xoc and negatively regulates expression of xopC2 [43]. The hypersensitive reaction (HR)- and pathogenicity-associated phosphatase gene hpaP, for which the protein carries the activity of ATPase and phosphatase, is essential to the full virulence of both Xoc and Xoo. The hpaP-regulated expression patterns of hrp genes in Xoc are different from those in Xoo, under the conditions of a plant inner-environment mimic medium and in planta [44].
2.2. Non-TALEs from Xoc
A range of non-TALEs are found in the Xoc genome, most of which are cognitively limited. AvrRxo1 is one of the first T3SEs identified in the Xoc genome that does not exist in the Xoo genome and acts as an avirulence effector that is recognized by maize resistance protein Rxo1 [10,45]. In addition to participation in the AvrRxo1-Rxo1 interaction, the virulence role of AvrRxo1 is also concerned and discovered. AvrRxo1 acts as an authentic phosphotransferase toward NAD to generate 3′-NADP, which is toxic to various cell types, including bacteria, fungi, and plants [46,47,48]. The avrRxo1 gene contains two open reading frames (ORFs), AvrRxo1-ORF1 and AvrRxo1-ORF2; the crystal structure indicates that AvrRxo1-ORF1 is formed with AvrRxo1-ORF2 as a tetramer that eliminates the toxicity of AvrRxo1-ORF1 [49]. Recently, AvrRxo1 was found to interact with and degrade the rice pyridoxal phosphate synthase OsPDX1, which blocks the vitamin B6 (VB6) biosynthesis pathway to promote infection in rice [50]. In addition, AvrRxo1 can interact with the Arabidopsis ubiquitin E3 ligase SINAT4 and facilitate activity, and it can also interact with and promote degradation of the cysteine protease RD21A, which positively regulates drought-induced immunity in Arabidopsis [51]. AvrBs2 is identified as a virulence factor in Xoc strain RS105 that has the ability to suppress pathogen-associated molecular pattern-triggered immunity (PTI). Additionally, this effector and another T3SE, XopN, both contribute to the full virulence of Xoc strain GX01 [52,53]. However, the targets of these two effectors in rice have not yet been identified. The AvrBs2 homologue in Xanthomonas euvesicatoria is corresponding to pepper R protein Bs2 and has a phosphodiesterase (GDE) domain, which is essential for its virulence but not for the Bs2-mediated defense [54]. The XopN homologue in Xanthomonas campestris pv. vesicatoria interacts with Tomato Atypical Receptor-Like Kinase1 (TARK1) as well as Tomato Fourteen–Three–Three isoforms 1 (TFT1) to overcome TARK1-mediated defense [55]. The targets of AvrBs2 and XopN in rice and how to interfere with rice defenses need more investigation. Another non-TALE, XopC2, from Xoc strain RS105, is annotated as a class of atypical kinases that phosphorylates adaptor protein OSK1 to facilitate the activation of jasmonate signaling and stomatal opening, which increases rice susceptibility to Xoc [56]. XopAP is as a lipase that works as a virulence factor of Xoc strain BLS256, which competitively binds to the molecule phosphatidylinositol 3,5-bisphosphate (PtdIns(3,5)P2) with vacuolar H+-pyrophosphatase (V-PPase) to compromise stomatal defense in rice [57]. Recently, XopAK was found to suppress the transcription of an MYB transcription factor gene, OsMYBxoc1, which works as a transcriptional repressor of ion-transport-related gene OsNRAMP5 to promote Xoc infection [58]. These findings regarding non-TALEs in Xoc indicate that several effectors disturb host stomatal immunity to promote infection, as summarized in Figure 1 and Table 1.
2.3. TALEs from Xoc
We depicted the TALEs identified from different strains of Xoc and the targets in rice (Figure 1). The first TALE identified in Xoc was Tal2g from BLS256. The deletion of Tal2g significantly reduces the virulence of BLS256 in rice [11]. Tal2g targets the effector-binding element (EBE) in the promoter of rice sulfate transporter gene OsSULTR3;6 to activate expression and promote invasion [11]. The homologous Tal5d from Xoc strain RS105 is different from Tal2g in the 10th RVD. Tal5d also targets OsSULTR3;6; gene editing of the EBE in the promoter of OsSULTR3;6 blocks the induction by Tal2g and Tal5d, thus developing rice broad-spectrum resistance to BLS [59]. Tal2h belongs to the truncated TALEs from Xoc strain BLS256 and has the ability to suppress Xo1-mediated resistance to African Xoc strains in the Carolina Gold Select rice variety. No potential recognition sites for Tal2h were predicted in rice, and the suppression function of Tal2h may directly or indirectly occur through interactions with the Xo1 protein [60,61]. Another TALE, Tal2a, from Xoc strain BLS256, can elicit HR in rice, as well as target and activate a ubiquitin carboxy-terminal hydrolase gene UCH, which is insufficient for triggering HR. Loss of Tal2a has no effect on virulence, whereas the overexpression of this effector reduces the virulence of BLS256 [62]. Tal7 from Xoc strain RS105 is able to suppress avrXa7-Xa7-triggered defense. Tal7 targets both a predicted Cyclin-D4-1 gene Os09g29100 and a GATA zinc finger family gene Os12g42970. Editing of the EBE in the Os09g29100 promoter confers enhanced resistance to RS105 [63]. Recently, Tal10a from Xoc strain GX01 targets and activates a rice hexokinase gene OsHXK5 to promote infection, and editing the EBEs in the promoters of OsHXK5 and OsSULTR3;6 significantly improves rice resistance to Xoc [64]. TalI deficiency in Xoc strain GD41 compromises full virulence, and TalI disturbs the salicylic acid (SA) signaling pathway to suppress rice defense but has no putative target genes [65]. In the latest studies, two TALEs, Tal2b and Tal2c, from the Xoc strain HGA4 are identified as virulence factors after being transferred into RS105, which is a strain that does not contain either of these two effectors. Generally, Tal2b and Tal2c exist together in the genome of many Xoc strains, whereas the mutant of either Tal2b or Tal2c homologues in Xoc strain BLS256 does not affect virulence [11]. Tal2b and Tal2c target rice flavanone 3-hydroxylase genes OsF3H03g and OsF3H04g, respectively, to suppress SA-mediated defense and to redundantly regulate pathogenic virulence in HGA4 [9,66]. These two genes and another homologous gene OsS3H are SA hydroxylases that reduce the SA levels and negatively regulate broad-spectrum resistance in rice [67,68,69,70]. Well known in rice–Xoo interactions, multiple TALEs independently target different members of rice clade III SWEETs to facilitate infection. Recently, clade III SWEETs were found to interact with OsHsp20L or OsHMGB1 to decrease rice resistance to Xoo and Magnaporthe oryzae [71,72,73]. As such, Xoc may employ different TALEs to target homologous SA hydroxylase genes to overcome broad-spectrum resistance.

Table 1.
The identified T3SE-targets in rice–Xoc interaction.
Table 1.
The identified T3SE-targets in rice–Xoc interaction.
| Effectors | Targets | Function and Mechanism of Effector-Targets | References |
|---|---|---|---|
| Non-TALEs | |||
| AvrRxo1 | NAD; OsPDX1 | Phosphotransferase; cytotoxicity; degradation; suppression of VB6 biosynthesis; activation of stomatal opening | [46,47,48,49,50] |
| AvrBs2 | Unknown | Suppression of PAMP-triggered defense | [52,53] |
| XopN | Unknown | Unknown | [53] |
| XopC2 | OSK1 | Phosphorylation; activation of JA signaling and stomatal opening | [56] |
| XopAP | PtdIns(3,5)P2 | Lipase; binding and competing; suppression of stomatal defense | [57] |
| XopAk | OsMYBxoc1 | Transcriptional suppression; accumulation of iron ions | [58] |
| TALEs | |||
| Tal2g/Tal5d | OsSULTR3;6 | Transcriptional activation | [11,59] |
| Tal2h | Xo1 | Protein–protein interaction; suppression of Xo1-mediated resistance | [60,61] |
| Tal2a | UCH | Transcriptional activation; induction of hypersensitive reaction | [62] |
| Tal7 | CycD4;1 | Transcriptional activation; suppression of avrXa7-Xa7-mediated resistance | [63] |
| Tal10a | OsHXK5 | Transcriptional activation; suppression of PR gene expression and MAPK activation | [64] |
| TalI | Unknown | Transcriptional activation; interference of SA signaling | [65] |
| Tal2b | OsF3H03g | Transcriptional activation; reduction in SA level | [9] |
| Tal2c | OsF3H04g | Transcriptional activation; reduction in SA level | [66] |
Notes: JA (jasmonic acid), MAPK (mitogen-activated protein kinase), NAD (nicotinamide adenine dinucleotide), OSK1 (Oryza sativa SKP1-like), PAMP (pathogen-associated molecular pattern), PR (pathogenesis-related), PtdIns(3,5)P2 (phosphatidylinositol-3,5-bisphosphate), SA (salicylic acid), UCH (ubiquitin carboxy-terminal hydrolase), VB6 (vitamin B6).
3. Diffusible Signal Factor (DSF)-Mediated Quorum Sensing (QS) and Extracellular Polysaccharides (EPSs) Are Involved in Xoc Virulence
In bacteria, DSF-mediated QS is involved in virulence-associated functions and diverse biological progresses [74]. In Xanthomonas, DSFs positively regulate EPS production, which is involved in virulence [75,76]. In Xoc, rpfF, which encodes DSF synthase, plays an essential role in virulence, and loss of rpfF reduces EPS production [77,78]. DSFs and Clp positively regulate the expression of the nodB-rghB locus genes hshA-C, which are essential for the full virulence of Xoc [77,79,80]. The expressions of an EPS- and virulence-related gene (epv) encoding a hypothetical protein and asnB encoding an asparagine synthetase are both positively regulated by DSFs and the global regulator Clp. Loss of epv substantially impairs the pathogenic virulence of Xoc; however, EPS production is decreased in the epv mutant but not in the asnB mutant [81,82]. Moreover, DSFs regulate three T2SS-related genes and an Ax21-like gene, which contribute to the full virulence of Xoc; these genes are also involved in EPS production [83]. FE36_09860 and FE36_06255, which belong to the TetR family of transcriptional regulators, are required for Xoc virulence and are regulated by DSFs and Clp. FE36_09860 and FE36_06255 mutants both decrease the virulence of Xoc, and a significant reduction in EPS production was observed with FE36_06255 deletion but not with FE36_09860 deletion [84]. Additionally, sugar-metabolizing enzymes, including glucose 6-phosphate isomerase (Pgi) and glucose-6-phosphate dehydrogenase (Zwf), catalyze glucose 6-phosphate into gluconate 6-phosphate and 6-phosphate into fructose 6-phosphate, respectively. Both Pgi and Zwf are required for the virulence of Xoc in rice. The mutant of Pgi or Zwf in Xoc results in reductions in DSF signals as well as EPS production [39,85]. The DSF-regulated virulence of Xoc is displayed in Figure 2.
4. Other Factors Affect the Virulence of Xoc
4.1. Molecules
Many microorganisms have the ability to generate gibberellin (GA) through the ent-kaurene synthesis pathway [86]. GA-synthesis-related enzymes, ent-copalyl diphosphate synthase (CPS), ent-kaurene synthase (KS), and cytochromes P450 (CYP112) in the same operon, are required for the full virulence of Xoc. These synthetases catalyze the production of bioactive GA4 to suppress jasmonic acid (JA)-mediated defense in rice [87,88,89]. In addition to GA, Xoc also has the ability to biosynthesize indole-3-acetic acid (IAA). Two nitrilase enzyme family (NIT) genes, NIT24 and NIT29, are involved in IAA synthesis, which contribute to the virulence of Xoc [90]. c-di-GMP is one of the important second messengers in bacterial signaling transduction, and the GGDEF-EAL and HD-GYP domain proteins are involved in c-di-GMP biosynthesis [91]. One of the HD-GYP domain regulatory proteins, rpfG, is involved in the virulence of Xoc. A rpfG mutant reduces EPS production and virulence but increases biofilm formation and the expression of T3SS [92]. Of the eleven GGDEF-EAL proteins in the Xoc genome, one of this family gene, XOC_4190, plays as a positive role in the virulence of Xoc, but the other 10 homologous genes do not. The loss function of XOC_4190 impairs the virulence and increases the sliding motility of Xoc but has little effect on the endogenous c-di-GMP density, even though purified XOC_4190 proteins can bind to c-di-GMP [93]. Another EAL domain encoding protein gene pde may be involved in c-di-GMP biosynthesis in Xoc. The pathogenic factors in the pde mutant are reduced, which impairs the virulence of Xoc [94]. The histidine biosynthesis operon in Xoc contains nine genes; among them, trpR and hisB are predominantly required for the virulence of Xoc. trpR, encoding a transcriptional regulator, negatively regulates the expression of histidine synthetase (his) genes. hisB is required for xanthomonadins biosynthesis and positively regulates the expression of pigment-associated genes. One of these xanthomonadins biosynthesis genes, aroE, is also essential for the production of xanthomonadins and the full virulence of Xoc [95].
4.2. Two-Component Regulatory System (TCS)
TCS is the main signal transduction system in bacteria [96]. A total of 110 TCS genes have been annotated in the genome of Xoc strain GX01, including 56 histidine kinases (HKs) and 54 response regulators (RRs). Seventeen of the RR genes contribute to the full virulence of Xoc [97]. Another TCS pair, PdeK-PdeR, can interact with the c-di-GMP binding protein FimX to assemble the type IV pili complex to promote virulence. Moreover, loss-of-function pdeK impairs the virulence and swimming motility of Xoc, whereas loss-of-function pdeR does not significantly affect virulence [98]. In addition, an RR gene, vemR, is extraordinarily required for the virulence of Xoc in rice, and the VemR/CheA pair functions as a TCS HK/RR for phosphorylation. Otherwise, VemR interacts with OGDH, SirA, FlgA, and HrpG to regulate different processes in Xoc [99].
4.3. Other Factors
EcpA is the first identified extracellular protease and is a virulence factor in Xoc. Purified EcpA proteins can cause chlorosis- and necrosis-like symptoms that are similar to those achieved with the injection of Xoc in rice leaves [100]. Polygalacturonases (PGs) are involved in the degradation of plant cell wall, which are secreted by bacteria, fungi, and pests to destroy the physical defenses of plant surface layer. To resist the destruction of cell walls by the PGs of phytopathogens, plants employ polygalacturonase-inhibiting proteins (PGIPs) to recognize PGs and inhibit the complete digestion of polygalacturonic acid, thus generating oligogalacturonides (OGs) to activate host immunity responses [101,102]. In Xoc strain RS105, the loss function of the unique pg gene reduces the virulence in both susceptible rice varieties and moderately resistant rice varieties. And the overexpression of rice OsPGIP1 increases resistance to Xoc beyond response to the PG of Xoc [103]. The mutant of xoc_3248, which encodes an inner membrane protein (Imp), impairs the virulence of Xoc in rice and attenuates carbohydrate utilization [104]. xocR, encoding the plant-associated bacteria (PAB) LuxR protein, participates in the cell swimming of Xoc. XocR can bind to the promoters of two Xoc virulence-associated genes, recN and trpE, to regulate their transcription; the loss function of xocR severely reduces the virulence of Xoc [105]. The horizontal transfer gene xoc_2868, encoding a transcriptional factor in Xoc, may be derived from a Burkholderiaceae ancestor. XOC_2868 directly activates the expression of the cydAB operon, which encodes two subunits of cytochrome bd oxidase to overcome oxidative stress. The xoc_2868, cydA, or cydAB mutants reduce the virulence of Xoc [106,107].
5. Discussion and Perspectives
Xoc contains various types of T3SEs, but their functions and host targets are unknown. Here, we discussed the six non-TALEs, AvrRxo1, AvrBs2, XopN, XopC2, XopAP, and XopAK, and eight TALEs, Tal2g, Ta2h, Tal2a, Ta7, TalI, Tal10a, Tal2b, and Tal2c (Figure 1). Three non-TALEs, XopC2, AvrRxo1, and XopAP, hinder stomatal immunity in rice [50,56,57]. This implies that Xoc possesses many complex effectors that interfere with rice stomatal-mediated defense. As Xoc is a type of bacteria which can invade into hosts through stoma, how to overcome rice stomatal immunity is important for Xoc to successfully infect rice. Thus, Xoc may develop XopC2, AvrRxo1, and XopAP to target rice different defense response signaling pathways to combat stomatal immunity. Several dozens of T3SEs are present in the Xoc genome that may have others of the three non-TALEs to interfere with rice stomatal immunity. However, a single mutant of the 23 non-TALEs in Xoc strain RS105 and GX01 only identified the virulence role of AvrBs2 and XopN [52,53]. This indicates that the non-TALEs in Xoc may work as virulence factors redundantly. How to effectively identify the non-TALEs which are involved in the intrusion of stomatal immunity is a problem for the investigations of rice–Xoc interaction. In rice, the overexpression of VB6 biosynthesis gene OsPDX1.2, for which the protein is a target of AvrRxo1, promotes stomatal closure and increases rice resistance to Xoc [50]. Thus, modifying the stomatal immunity-related genes may improve resistance against Xoc in rice.
In addition to the similar composition of non-TALEs, Xoc has different numbers of TALEs in different strains, even as many as almost 30 TALEs in some strains [8,9]. Similar to non-TALEs, the mutant of TALEs in Xoc strain BLS256 one by one only identifies a few virulent TALEs. Among these TALEs in BLS256, the mutant of either Tal2c or Tal2d has no significant influence on virulence [11]. However, we identified the virulence roles of Tal2b and Tal2c in Xoc strain HGA4, which are homologous to Tal2c and Tal2d in BLS256, respectively. We performed a comparison of the TALomes between Xoc strains of strongly pathogenic HGA4 and weakly pathogenic RS105 to identify that Tal2b and Tal2c pair-existed in HGA4 but not in RS105. Gain-of-function Tal2b or Tal2c in RS105 both improve virulence in rice [9,66]. This indicates that Tal2b and Tal2c have functional redundancy. These two TALEs target and activate two homologous SA hydroxylase genes, respectively, to suppress rice SA-mediated defenses redundantly [9,11,66]. Furthermore, another TALE, TalI, destroys SA signaling to promote infection in rice with unknown target genes [65]. Thus, Xoc employs multiple effectors to manipulate the similar rice susceptible pathways to improve pathogenicity. This may explain why attempts to identify the virulence roles with individual T3SEs have been unsuccessful in Xoc. Researchers have proposed that consideration of whole effectomes and plant target repertoire may efficiently understand pathogen–host interaction [108]. As such, the diversity of effectomes from different Xoc strains should be systematically investigated to identify the interactions among effectors and targets.
TALEs have the ability to bind to the EBEs of host target genes and activate genes’ expressions. If blocking the binding of EBEs of the susceptibility genes, it may prevent host susceptible signaling pathways to overcome the infection. In rice–Xoo interaction, the members of rice clade III SWEETs are important susceptibility genes which activate by different TALEs of Xoo. Genome editing of the EBEs of these OsSWEETs confers rice broad-spectrum resistance to different Xoo strains [109,110]. In rice–Xoc interaction, OsSULTR3;6 is a major susceptibility gene which is proved to be targeted and activated by the Tal2g of Xoc strain BLS256 and the Tal5d of Xoc strain RS105 [11,59]. The Tal2g homologues ubiquitously exist in different Xoc strains [8,9]. Thus, genome editing of the EBEs of OsSULTR3;6 may also confer rice broad-spectrum resistance to Xoc. Recent research has validated this hypothesis. Editing the EBEs of OsSULTR3;6 gene in rice varieties IRBB10, Guihong 1, Zhonghua 11, Nipponbare, and 58B improves resistance to different Xoc strains [59,64,111,112,113]. Moreover, the EBE of OsSULTR3;6 gene is lacked only in 1% of genome-sequenced rice cultivars, which are increasingly resistant to Xoc [114]. Thus, the screening and breeding of rice cultivars with the deletion of the EBEs in OsSULTR3;6 gene may be an important strategy for BLS disease-resistant breeding. In addition, the Tal2b and Tal2c from Xoc strain HGA4 work as virulence factors by targeting and activating two SA hydroxylase genes OsF3H03g and OsF3H04g, respectively [9,66]. However, the loss function either of Tal2b or Tal2c homologues do not affect Xoc virulence and genome editing either of the EBEs of OsF3H03g or OsF3H04g also do not increase rice resistance to Xoc strain, for which the genome has both Tal2b and Tal2c [11,66]. This indicates that the virulence function of these two TALEs is redundant by targeting two homologous genes. Thus, gene editing both of the EBEs of OsF3H03g and OsF3H04g may enhance rice resistance to the Xoc strains, which contain both Tal2b and Tal2c. In conclusion, using comparative effectomes to discover the virulent T3SEs in Xoc and the common targets in rice, and modifying the pathways that are commonly hijacked by Xoc, are important strategies to control this disease in the future.
In this review, we summarized the reported TALEs and non-TALEs from Xoc that work as virulence factors to interfere with rice defense (Figure 1), as well as the virulence factors, including DSF-mediated QS and EPS, molecules, regulation systems, etc., in Xoc (Figure 2). Several non-TALEs disturb rice stomatal immunity, and several TALEs attenuate rice SA-mediated defense. This may remind us that manipulating these defense signaling pathways attacked by T3SEs of Xoc can protect rice from this pathogen.
Author Contributions
Writing—original draft preparation, N.T.; writing—literature collection, Y.H.; writing—review and editing, T.W. and W.M.; supervision, T.W.; funding acquisition, Q.Z. and T.W. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (32302366).
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Niño-Liu, D.O.; Ronald, P.C.; Bogdanove, A.J. Xanthomonas oryzae pathovars: Model pathogens of a model crop. Mol. Plant Pathol. 2006, 7, 303–324. [Google Scholar] [CrossRef] [PubMed]
- Jiang, N.; Yan, J.; Liang, Y.; Shi, Y.; He, Z.; Wu, Y.; Zeng, Q.; Liu, X.; Peng, J. Resistance genes and their interactions with bacterial blight/leaf streak pathogens (Xanthomonas oryzae) in rice (Oryza sativa L.)-an updated review. Rice 2020, 13, 3. [Google Scholar] [CrossRef] [PubMed]
- Kay, S.; Bonas, U. How Xanthomonas type III effectors manipulate the host plant. Curr. Opin. Microbiol. 2009, 12, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zou, H.; Che, Y.; Cui, Y.; Guo, W.; Zou, L.; Chatterjee, S.; Biddle, E.M.; Yang, C.; Chen, G. A novel regulatory role of HrpD6 in regulating hrp-hrc-hpa genes in Xanthomonas oryzae pv. oryzicola. Mol. Plant Microbe Interact. 2011, 24, 1086–1101. [Google Scholar] [CrossRef]
- White, F.F.; Potnis, N.; Jones, J.B.; Koebnik, R. The type III effectors of Xanthomonas. Mol. Plant Pathol. 2009, 10, 749–766. [Google Scholar] [CrossRef]
- Boch, J.; Scholze, H.; Schornack, S.; Landgraf, A.; Hahn, S.; Kay, S.; Lahaye, T.; Nickstadt, A.; Bonas, U. Breaking the code of DNA binding specificity of TAL-type III effectors. Science 2009, 326, 1509–1512. [Google Scholar] [CrossRef]
- Ji, Z.; Zakria, M.; Zou, L.; Xiong, L.; Li, Z.; Ji, G.; Chen, G. Genetic diversity of transcriptional activator-like effector genes in Chinese isolates of Xanthomonas oryzae pv. oryzicola. Phytopathology 2014, 104, 672–682. [Google Scholar] [CrossRef]
- Wilkins, K.E.; Booher, N.J.; Wang, L.; Bogdanove, A.J. TAL effectors and activation of predicted host targets distinguish Asian from African strains of the rice pathogen Xanthomonas oryzae pv. oryzicola while strict conservation suggests universal importance of five TAL effectors. Front. Plant Sci. 2015, 6, 536. [Google Scholar] [CrossRef]
- Wu, T.; Zhang, H.; Yuan, B.; Liu, H.; Kong, L.; Chu, Z.; Ding, X. Tal2b targets and activates the expression of OsF3H03g to hijack OsUGT74H4 and synergistically interfere with rice immunity. New Phytol. 2022, 233, 1864–1880. [Google Scholar] [CrossRef]
- Zhao, B.; Ardales, E.Y.; Raymundo, A.; Bai, J.; Trick, H.N.; Leach, J.E.; Hulbert, S.H. The avrRxo1 gene from the rice pathogen Xanthomonas oryzae pv. oryzicola confers a nonhost defense reaction on maize with resistance gene Rxo1. Mol. Plant Microbe Interact. 2004, 17, 771–779. [Google Scholar] [CrossRef]
- Cernadas, R.A.; Doyle, E.L.; Niño-Liu, D.O.; Wilkins, K.E.; Bancroft, T.; Wang, L.; Schmidt, C.L.; Caldo, R.; Yang, B.; White, F.F.; et al. Code-assisted discovery of TAL effector targets in bacterial leaf streak of rice reveals contrast with bacterial blight and a novel susceptibility gene. PLoS Pathog. 2014, 10, e1003972. [Google Scholar] [CrossRef] [PubMed]
- Büttner, D.; Bonas, U. Regulation and secretion of Xanthomonas virulence factors. FEMS Microbiol. Rev. 2010, 34, 107–133. [Google Scholar] [CrossRef]
- Timilsina, S.; Potnis, N.; Newberry, E.A.; Liyanapathiranage, P.; Iruegas-Bocardo, F.; White, F.F.; Goss, E.M.; Jones, J.B. Xanthomonas diversity, virulence and plant-pathogen interactions. Nat. Rev. Microbiol. 2020, 18, 415–427. [Google Scholar] [CrossRef] [PubMed]
- Limoli, D.H.; Jones, C.J.; Wozniak, D.J. Bacterial extracellular polysaccharides in biofilm formation and function. Microbiol. Spectr. 2015, 3, 1–30. [Google Scholar] [CrossRef]
- Whitfield, C.; Wear, S.S.; Sande, C. Assembly of bacterial capsular polysaccharides and exopolysaccharides. Annu. Rev. Microbiol. 2020, 74, 521–543. [Google Scholar] [CrossRef] [PubMed]
- Dunger, G.; Relling, V.M.; Tondo, M.L.; Barreras, M.; Ielpi, L.; Orellano, E.G.; Ottado, J. Xanthan is not essential for pathogenicity in citrus canker but contributes to Xanthomonas epiphytic survival. Arch. Microbiol. 2007, 188, 127–135. [Google Scholar] [CrossRef]
- Kim, S.Y.; Kim, J.G.; Lee, B.M.; Cho, J.Y. Mutational analysis of the gum gene cluster required for xanthan biosynthesis in Xanthomonas oryzae pv. oryzae. Biotechnol. Lett. 2009, 31, 265–270. [Google Scholar] [CrossRef]
- Bianco, M.I.; Toum, L.; Yaryura, P.M.; Mielnichuk, N.; Gudesblat, G.E.; Roeschlin, R.; Marano, M.R.; Ielpi, L.; Vojnov, A.A. Xanthan pyruvilation is essential for the virulence of Xanthomonas campestris pv. campestris. Mol. Plant Microbe Interact. 2016, 29, 688–699. [Google Scholar] [CrossRef]
- Ryan, R.P.; An, S.Q.; Allan, J.H.; McCarthy, Y.; Dow, J.M. The DSF family of cell-cell signals: An expanding class of bacterial virulence regulators. PLoS Pathog. 2015, 11, e1004986. [Google Scholar] [CrossRef]
- Feng, Y.; Long, Z.; Xiang, H.; Ran, J.; Zhou, X.; Yang, S. Research on diffusible signal factor-mediated quorum sensing in Xanthomonas: A mini-review. Molecules 2023, 28, 876. [Google Scholar] [CrossRef]
- Simpson, B.W.; Trent, M.S. Pushing the envelope: LPS modifications and their consequences. Nat. Rev. Microbiol. 2019, 17, 403–416. [Google Scholar] [CrossRef] [PubMed]
- Di Lorenzo, F.; Silipo, A.; Andersen Gersby, L.B.; Palmigiano, A.; Lanzetta, R.; Garozzo, D.; Boyer, C.; Pruvost, O.; Newman, M.A.; Molinaro, A. Xanthomonas citri pv. citri pathotypes: LPS structure and function as microbe-associated molecular patterns. ChemBioChem 2017, 18, 772–781. [Google Scholar] [CrossRef]
- Singh, A.; Bansal, K.; Kumar, S.; Patil, P.B. Deep population genomics reveals systematic and parallel evolution at a lipopolysaccharide biosynthetic locus in Xanthomonas pathogens that infect rice and sugarcane. Appl. Environ. Microbiol. 2022, 88, e0055022. [Google Scholar] [CrossRef]
- Green, E.R.; Mecsas, J. Bacterial secretion systems: An overview. Microbiol. Spectr. 2016, 4, 213–239. [Google Scholar] [CrossRef]
- Kanonenberg, K.; Spitz, O.; Erenburg, I.N.; Beer, T.; Schmitt, L. Type I secretion system-it takes three and a substrate. FEMS Microbiol. Lett. 2018, 365, fny094. [Google Scholar] [CrossRef]
- Meuskens, I.; Saragliadis, A.; Leo, J.C.; Linke, D. Type V secretion systems: An overview of passenger domain functions. Front. Microbiol. 2019, 10, 1163. [Google Scholar] [CrossRef] [PubMed]
- Büttner, D.; He, S.Y. Type III protein secretion in plant pathogenic bacteria. Plant Physiol. 2009, 150, 1656–1664. [Google Scholar] [CrossRef] [PubMed]
- Puhar, A.; Sansonetti, P.J. Type III secretion system. Curr. Biol. 2014, 24, R784–R791. [Google Scholar] [CrossRef]
- Rossier, O.; Wengelnik, K.; Hahn, K.; Bonas, U. The Xanthomonas Hrp type III system secretes proteins from plant and mammalian bacterial pathogens. Proc. Natl. Acad. Sci. USA 1999, 96, 9368–9373. [Google Scholar] [CrossRef]
- Tampakaki, A.P.; Fadouloglou, V.E.; Gazi, A.D.; Panopoulos, N.J.; Kokkinidis, M. Conserved features of type III secretion. Cell. Microbiol. 2004, 6, 805–816. [Google Scholar] [CrossRef]
- Gürlebeck, D.; Thieme, F.; Bonas, U. Type III effector proteins from the plant pathogen Xanthomonas and their role in the interaction with the host plant. J. Plant Physiol. 2006, 163, 233–255. [Google Scholar] [CrossRef]
- Zou, L.; Wang, X.; Xiang, Y.; Zhang, B.; Li, Y.; Xiao, Y.; Wang, J.; Walmsley, A.R.; Chen, G. Elucidation of the hrp clusters of Xanthomonas oryzae pv. oryzicola that control the hypersensitive response in nonhost tobacco and pathogenicity in susceptible host rice. Appl. Environ. Microbiol. 2006, 72, 6212–6224. [Google Scholar] [CrossRef]
- Li, Y.; Che, Y.; Zou, H.; Cui, Y.; Guo, W.; Zou, L.; Biddle, E.M.; Yang, C.; Chen, G. Hpa2 required by HrpF to translocate Xanthomonas oryzae transcriptional activator-like effectors into rice for pathogenicity. Appl. Environ. Microbiol. 2011, 77, 3809–3818. [Google Scholar] [CrossRef] [PubMed]
- Zou, L.; Li, Y.; Chen, G. A non-marker mutagenesis strategy to generate poly-hrp gene mutants in the rice pathogen Xanthomonas oryzae pv. oryzicola. Agric. Sci. China 2011, 10, 12. [Google Scholar] [CrossRef]
- Guo, W.; Cui, Y.; Li, Y.; Che, Y.; Yuan, L.; Zou, L.; Zou, H.; Chen, G. Identification of seven Xanthomonas oryzae pv. oryzicola genes potentially involved in pathogenesis in rice. Microbiology 2012, 158, 505–518. [Google Scholar] [CrossRef]
- Cui, Y.; Zou, L.; Zou, H.; Li, Y.; Zakria, M.; Chen, G. HrpE3 is a type III effector protein required for full virulence of Xanthomonas oryzae pv. oryzicola in rice. Mol. Plant Pathol. 2013, 14, 678–692. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Zou, L.; Li, Y.; Zou, H.; Liu, X.; Chen, G. Xoryp_08180 of Xanthomonas oryzae pv. oryzicola, encoding a hypothetical protein, is regulated by HrpG and HrpX and required for full virulence in rice. J. Integr. Agric. 2012, 11, 600–610. [Google Scholar] [CrossRef]
- Guo, W.; Zou, L.; Li, Y.; Cui, Y.; Ji, Z.; Cai, L.; Zou, H.; Hutchins, W.C.; Yang, C.; Chen, G. Fructose-bisphophate aldolase exhibits functional roles between carbon metabolism and the hrp system in rice pathogen Xanthomonas oryzae pv. oryzicola. PLoS ONE 2012, 7, e31855. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Chu, C.; Yang, X.; Fang, Y.; Liu, X.; Chen, G.; Liu, J. Phosphohexose mutase of Xanthomonas oryzae pv. oryzicola is negatively regulated by HrpG and HrpX, and required for the full virulence in rice. Eur. J. Plant Pathol. 2014, 140, 353–364. [Google Scholar] [CrossRef]
- Xue, X.; Zou, L.; Ma, W.; Liu, Z.; Chen, G. Identification of 17 HrpX-regulated proteins including two novel type III effectors, XOC_3956 and XOC_1550, in Xanthomonas oryzae pv. oryzicola. PLoS ONE 2014, 9, e93205. [Google Scholar] [CrossRef]
- Wang, L.; Makino, S.; Subedee, A.; Bogdanove, A.J. Novel candidate virulence factors in rice pathogen Xanthomonas oryzae pv. oryzicola as revealed by mutational analysis. Appl. Environ. Microbiol. 2007, 73, 8023–8027. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Vinogradov, E.V.; Bogdanove, A.J. Requirement of the lipopolysaccharide O-chain biosynthesis gene wxocB for type III secretion and virulence of Xanthomonas oryzae pv. oryzicola. J. Bacteriol. 2013, 195, 1959–1969. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Wang, S.; Nie, W.; Wang, P.; Fu, L.; Ahmad, I.; Zhu, B.; Chen, G. A key antisense sRNA modulates the oxidative stress response and virulence in Xanthomonas oryzae pv. oryzicola. PLoS Pathog. 2021, 17, e1009762. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Ren, P.; Zhang, D.; Cui, P.; Zhu, G.; Xian, X.; Tang, J.; Lu, G. HpaP divergently regulates the expression of hrp genes in Xanthomonas oryzae pathovars oryzae and oryzicola. Mol. Plant Pathol. 2023, 24, 44–58. [Google Scholar] [CrossRef]
- Zhao, B.; Lin, X.; Poland, J.; Trick, H.; Leach, J.; Hulbert, S. A maize resistance gene functions against bacterial streak disease in rice. Proc. Natl. Acad. Sci. USA 2005, 102, 15383–15388. [Google Scholar] [CrossRef]
- Liu, H.; Chang, Q.; Feng, W.; Zhang, B.; Wu, T.; Li, N.; Yao, F.; Ding, X.; Chu, Z. Domain dissection of AvrRxo1 for suppressor, avirulence and cytotoxicity functions. PLoS ONE 2014, 9, e113875. [Google Scholar] [CrossRef]
- Triplett, L.R.; Shidore, T.; Long, J.; Miao, J.; Wu, S.; Han, Q.; Zhou, C.; Ishihara, H.; Li, J.; Zhao, B.; et al. AvrRxo1 is a bifunctional type III secreted effector and toxin-antitoxin system component with homologs in diverse environmental contexts. PLoS ONE 2016, 11, e0158856. [Google Scholar] [CrossRef] [PubMed]
- Schuebel, F.; Rocker, A.; Edelmann, D.; Schessner, J.; Brieke, C.; Meinhart, A. 3′-NADP and 3′-NAADP, two metabolites formed by the bacterial type III effector AvrRxo1. J. Biol. Chem. 2016, 291, 22868–22880. [Google Scholar] [CrossRef]
- Han, Q.; Zhou, C.; Wu, S.; Liu, Y.; Triplett, L.; Miao, J.; Tokuhisa, J.; Deblais, L.; Robinson, H.; Leach, J.E.; et al. Crystal structure of Xanthomonas AvrRxo1-ORF1, a type III effector with a polynucleotide kinase domain, and its interactor AvrRxo1-ORF2. Structure 2015, 23, 1900–1909. [Google Scholar] [CrossRef]
- Liu, H.; Lu, C.; Li, Y.; Wu, T.; Zhang, B.; Liu, B.; Feng, W.; Xu, Q.; Dong, H.; He, S.; et al. The bacterial effector AvrRxo1 inhibits vitamin B6 biosynthesis to promote infection in rice. Plant Commun. 2022, 3, 100324. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, K.; Cheng, Q.; Kong, D.; Zhang, X.; Wang, Z.; Wang, Q.; Xie, Q.; Yan, J.; Chu, J.; et al. Cysteine protease RD21A regulated by E3 ligase SINAT4 is required for drought-induced resistance to Pseudomonas syringae in Arabidopsis. J. Exp. Bot. 2020, 71, 5562–5576. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Wang, Y.; Wang, S.; Fang, A.; Wang, J.; Liu, L.; Zhang, K.; Mao, Y.; Sun, W. The type III effector AvrBs2 in Xanthomonas oryzae pv. oryzicola suppresses rice immunity and promotes disease development. Mol. Plant Microbe Interact. 2015, 28, 869–880. [Google Scholar] [CrossRef] [PubMed]
- Liao, Z.; Li, J.; Mo, X.; Ni, Z.; Jiang, W.; He, Y.; Huang, S. Type III effectors xopN and avrBS2 contribute to the virulence of Xanthomonas oryzae pv. oryzicola strain GX01. Res. Microbiol. 2020, 171, 102–106. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Dahlbeck, D.; Krasileva, K.V.; Fong, R.W.; Staskawicz, B.J. Computational and biochemical analysis of the Xanthomonas effector AvrBs2 and its role in the modulation of Xanthomonas type three effector delivery. PLoS Pathog. 2011, 7, e1002408. [Google Scholar] [CrossRef]
- Kim, J.G.; Li, X.; Roden, J.A.; Taylor, K.W.; Aakre, C.D.; Su, B.; Lalonde, S.; Kirik, A.; Chen, Y.; Baranage, G.; et al. Xanthomonas T3S effector XopN suppresses PAMP-triggered immunity and interacts with a Tomato Atypical Receptor-Like Kinase and TFT1. Plant Cell 2009, 21, 1305–1323. [Google Scholar] [CrossRef]
- Wang, S.; Li, S.; Wang, J.; Li, Q.; Xin, X.F.; Zhou, S.; Wang, Y.; Li, D.; Xu, J.; Luo, Z.Q.; et al. A bacterial kinase phosphorylates OSK1 to suppress stomatal immunity in rice. Nat. Commun. 2021, 12, 5479. [Google Scholar] [CrossRef]
- Liu, L.; Li, Y.; Xu, Z.; Chen, H.; Zhang, J.; Manion, B.; Liu, F.; Zou, L.; Fu, Z.Q.; Chen, G. The Xanthomonas type III effector XopAP prevents stomatal closure by interfering with vacuolar acidification. J. Integr. Plant Biol. 2022, 64, 1994–2008. [Google Scholar] [CrossRef]
- Zhang, H.; Sun, B.; Wu, W.; Li, Y.; Yin, Z.; Lu, C.; Zhao, H.; Kong, L.; Ding, X. The MYB transcription factor OsMYBxoc1 regulates resistance to Xoc by directly repressing transcription of the iron transport gene OsNRAMP5 in rice. Plant Commun. 2024, 5, 100859. [Google Scholar] [CrossRef]
- Xu, X.; Xu, Z.; Li, Z.; Zakria, M.; Zou, L.; Chen, G. Increasing resistance to bacterial leaf streak in rice by editing the promoter of susceptibility gene OsSULRT3;6. Plant Biotechnol. J. 2021, 19, 1101–1103. [Google Scholar] [CrossRef]
- Read, A.C.; Rinaldi, F.C.; Hutin, M.; He, Y.Q.; Triplett, L.R.; Bogdanove, A.J. Suppression of Xo1-mediated disease resistance in rice by a truncated, non-DNA-binding TAL effector of Xanthomonas oryzae. Front. Plant Sci. 2016, 7, 1516. [Google Scholar] [CrossRef]
- Read, A.C.; Hutin, M.; Moscou, M.J.; Rinaldi, F.C.; Bogdanove, A.J. Cloning of the rice Xo1 resistance gene and interaction of the Xo1 protein with the defense-suppressing Xanthomonas effector Tal2h. Mol. Plant Microbe Interact. 2020, 33, 1189–1195. [Google Scholar] [CrossRef]
- Hummel, A.W.; Wilkins, K.E.; Wang, L.; Cernadas, R.A.; Bogdanove, A.J. A transcription activator-like effector from Xanthomonas oryzae pv. oryzicola elicits dose-dependent resistance in rice. Mol. Plant Pathol. 2017, 18, 55–66. [Google Scholar] [CrossRef]
- Cai, L.; Cao, Y.; Xu, Z.; Ma, W.; Zakria, M.; Zou, L.; Cheng, Z.; Chen, G. A transcription activator-like effector Tal7 of Xanthomonas oryzae pv. oryzicola activates rice gene Os09g29100 to suppress rice immunity. Sci. Rep. 2017, 7, 5089. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Liao, Z.; Jin, X.; Liao, L.; Zhang, Y.; Zhang, R.; Zhao, X.; Qin, H.; Chen, J.; He, Y.; et al. Xanthomonas oryzae pv. oryzicola effector Tal10a directly activates rice OsHXK5 expression to facilitate pathogenesis. Plant J. 2024. [Google Scholar] [CrossRef] [PubMed]
- Long, J.; Wang, Z.; Chen, X.; Liu, Y.; Zhang, M.; Song, C.; Dong, H. Identification of a TAL effector in Xanthomonas oryzae pv. oryzicola enhancing pathogen growth and virulence in plants. Physiol. Mol. Plant Pathol. 2021, 114, 101620. [Google Scholar] [CrossRef]
- Wu, T.; Zhang, H.; Bi, Y.; Yu, Y.; Liu, H.; Yang, H.; Yuan, B.; Ding, X.; Chu, Z. Tal2c activates the expression of OsF3H04g to promote infection as a redundant TALE of Tal2b in Xanthomonas oryzae pv. oryzicola. Int. J. Mol. Sci. 2021, 22, 13628. [Google Scholar] [CrossRef]
- Liang, B.; Wang, H.; Yang, C.; Wang, L.; Qi, L.; Guo, Z.; Chen, X. Salicylic acid is required for broad-spectrum disease resistance in rice. Int. J. Mol. Sci. 2022, 23, 1354. [Google Scholar] [CrossRef]
- Zhang, Y.; Yu, Q.; Gao, S.; Yu, N.; Zhao, L.; Wang, J.; Zhao, J.; Huang, P.; Yao, L.; Wang, M.; et al. Disruption of the primary salicylic acid hydroxylases in rice enhances broad-spectrum resistance against pathogens. Plant Cell Environ. 2022, 45, 2211–2225. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Bi, Y.; Yu, Y.; Zhou, Z.; Yuan, B.; Ding, X.; Zhang, Q.; Chen, X.; Yang, H.; Liu, H.; et al. Activated expression of rice DMR6-like gene OsS3H partially explores the susceptibility to bacterial leaf streak mediated by knock-out OsF3H04g. Int. J. Mol. Sci. 2023, 24, 13263. [Google Scholar] [CrossRef]
- Liu, X.; Yu, Y.; Yao, W.; Yin, Z.; Wang, Y.; Huang, Z.; Zhou, J.Q.; Liu, J.; Lu, X.; Wang, F.; et al. CRISPR/Cas9-mediated simultaneous mutation of three salicylic acid 5-hydroxylase (OsS5H) genes confers broad-spectrum disease resistance in rice. Plant Biotechnol. J. 2023, 21, 1873–1886. [Google Scholar] [CrossRef]
- Gupta, P.K.; Balyan, H.S.; Gautam, T. SWEET genes and TAL effectors for disease resistance in plants: Present status and future prospects. Mol. Plant Pathol. 2021, 22, 1014–1026. [Google Scholar] [CrossRef]
- Wang, X.; Ju, Y.; Wu, T.; Kong, L.; Yuan, M.; Liu, H.; Chen, X.; Chu, Z. The clade III subfamily of OsSWEETs directly suppresses rice immunity by interacting with OsHMGB1 and OsHsp20L. Plant Biotechnol. J. 2024, 22, 2186–2200. [Google Scholar] [CrossRef]
- Shi, X.; Xie, X.; Guo, Y.; Zhang, J.; Gong, Z.; Zhang, K.; Mei, J.; Xia, X.; Xia, H.; Ning, N.; et al. A fungal core effector exploits the OsPUX8B.2-OsCDC48-6 module to suppress plant immunity. Nat. Commun. 2024, 15, 2559. [Google Scholar] [CrossRef] [PubMed]
- Mole, B.M.; Baltrus, D.A.; Dangl, J.L.; Grant, S.R. Global virulence regulation networks in phytopathogenic bacteria. Trends Microbiol. 2007, 15, 363–371. [Google Scholar] [CrossRef]
- Slater, H.; Alvarez-Morales, A.; Barber, C.E.; Daniels, M.J.; Dow, J.M. A two-component system involving an HD-GYP domain protein links cell-cell signalling to pathogenicity gene expression in Xanthomonas campestris. Mol. Microbiol. 2000, 38, 986–1003. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, S.; Sonti, R.V. rpfF mutants of Xanthomonas oryzae pv. oryzae are deficient for virulence and growth under low iron conditions. Mol. Plant Microbe Interact. 2002, 15, 463–471. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Qian, G.; Yin, F.; Fan, J.; Zhai, Z.; Liu, C.; Hu, B.; Liu, F. Proteomic analysis of the regulatory function of DSF-dependent quorum sensing in Xanthomonas oryzae pv. oryzicola. Microb. Pathog. 2011, 50, 48–55. [Google Scholar] [CrossRef]
- Rai, R.; Javvadi, S.; Chatterjee, S. Cell-cell signalling promotes ferric iron uptake in Xanthomonas oryzae pv. oryzicola that contribute to its virulence and growth inside rice. Mol. Microbiol. 2015, 96, 708–727. [Google Scholar] [CrossRef]
- Song, Z.; Zhao, Y.; Zhou, X.; Wu, G.; Zhang, Y.; Qian, G.; Liu, F. Identification and characterization of two novel DSF-controlled virulence-associated genes within the nodB-rhgB locus of Xanthomonas oryzae pv. oryzicola Rs105. Phytopathology 2015, 105, 588–596. [Google Scholar] [CrossRef]
- Song, Z.; Zhao, Y.; Qian, G.; Odhiambo, B.O.; Liu, F. Novel insights into the regulatory roles of gene hshB in Xanthomonas oryzae pv. oryzicola. Res. Microbiol. 2017, 168, 165–173. [Google Scholar] [CrossRef]
- Qian, G.; Zhang, Y.; Zhou, Y.; Liu, C.; Zhao, Y.; Song, Z.; Fan, J.; Hu, B.; Liu, F. epv, encoding a hypothetical protein, is regulated by DSF-mediating quorum sensing as well as global regulator Clp and is required for optimal virulence in Xanthomonas oryzae pv. oryzicola. Phytopathology 2012, 102, 841–847. [Google Scholar] [CrossRef]
- Qian, G.; Liu, C.; Wu, G.; Yin, F.; Zhao, Y.; Zhou, Y.; Zhang, Y.; Song, Z.; Fan, J.; Hu, B.; et al. AsnB, regulated by diffusible signal factor and global regulator Clp, is involved in aspartate metabolism, resistance to oxidative stress and virulence in Xanthomonas oryzae pv. oryzicola. Mol. Plant Pathol. 2013, 14, 145–157. [Google Scholar] [CrossRef] [PubMed]
- Qian, G.; Zhou, Y.; Zhao, Y.; Song, Z.; Wang, S.; Fan, J.; Hu, B.; Venturi, V.; Liu, F. Proteomic analysis reveals novel extracellular virulence-associated proteins and functions regulated by the diffusible signal factor (DSF) in Xanthomonas oryzae pv. oryzicola. J. Proteome Res. 2013, 12, 3327–3341. [Google Scholar] [CrossRef] [PubMed]
- Wan, X.; Ahmed, W.; Yang, J.; Meng, H.; Wei, L.; Ji, G. Identification and characterization of two transcriptional regulators in Xanthomonas oryzae pv. oryzicola YM15. Physiol. Mol. Plant Pathol. 2023, 124, 101964. [Google Scholar] [CrossRef]
- Guo, W.; Zou, L.; Ji, Z.; Cai, L.; Chen, G. Glucose 6-phosphate isomerase (Pgi) is required for extracellular polysaccharide biosynthesis, DSF signals production and full virulence of Xanthomonas oryzae pv. oryzicola in rice. Physiol. Mol. Plant Pathol. 2017, 100, 209–219. [Google Scholar] [CrossRef]
- Salazar-Cerezo, S.; Martínez-Montiel, N.; García-Sánchez, J.; Pérez-Y-Terrón, R.; Martínez-Contreras, R.D. Gibberellin biosynthesis and metabolism: A convergent route for plants, fungi and bacteria. Microbiol. Res. 2018, 208, 85–98. [Google Scholar] [CrossRef]
- Lu, X.; Hershey, D.M.; Wang, L.; Bogdanove, A.J.; Peters, R.J. An ent-kaurene-derived diterpenoid virulence factor from Xanthomonas oryzae pv. oryzicola. New Phytol. 2015, 206, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Nagel, R.; Peters, R.J. Investigating the phylogenetic range of gibberellin biosynthesis in bacteria. Mol. Plant Microbe Interact. 2017, 30, 343–349. [Google Scholar] [CrossRef]
- Nagel, R.; Turrini, P.C.; Nett, R.S.; Leach, J.E.; Verdier, V.; Van Sluys, M.A.; Peters, R.J. An operon for production of bioactive gibberellin A4 phytohormone with wide distribution in the bacterial rice leaf streak pathogen Xanthomonas oryzae pv. oryzicola. New Phytol. 2017, 214, 1260–1266. [Google Scholar] [CrossRef]
- Zhang, H.; Rong, Z.; Li, Y.; Yin, Z.; Lu, C.; Zhao, H.; Kong, L.; Meng, L.; Ding, X. NIT24 and NIT29-mediated IAA synthesis of Xanthomonas oryzae pv. oryzicola suppresses immunity and boosts growth in rice. Mol. Plant Pathol. 2024, 25, e13409. [Google Scholar] [CrossRef]
- Dow, J.M.; Fouhy, Y.; Lucey, J.F.; Ryan, R.P. The HD-GYP domain, cyclic di-GMP signaling, and bacterial virulence to plants. Mol. Plant Microbe Interact. 2006, 19, 1378–1384. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wei, C.; Jiang, W.; Wang, L.; Li, C.; Wang, Y.; Dow, J.M.; Sun, W. The HD-GYP domain protein RpfG of Xanthomonas oryzae pv. oryzicola regulates synthesis of extracellular polysaccharides that contribute to biofilm formation and virulence on rice. PLoS ONE 2013, 8, e59428. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.; Jiang, W.; Zhao, M.; Ling, J.; Zeng, X.; Deng, J.; Jin, D.; Dow, J.M.; Sun, W. A systematic analysis of the role of GGDEF-EAL domain proteins in virulence and motility in Xanthomonas oryzae pv. oryzicola. Sci. Rep. 2016, 6, 23769. [Google Scholar] [CrossRef]
- Wan, X.; Yang, J.; Ahmed, W.; Liu, Q.; Wang, Y.; Wei, L.; Ji, G. Functional analysis of pde gene and its role in the pathogenesis of Xanthomonas oryzae pv. oryzicola. Infect. Genet. Evol. 2021, 94, 105008. [Google Scholar] [CrossRef]
- Su, P.; Song, Z.; Wu, G.; Zhao, Y.; Zhang, Y.; Wang, B.; Qian, G.; Fu, Z.Q.; Liu, F. Insights into the roles of two genes of the histidine biosynthesis operon in pathogenicity of Xanthomonas oryzae pv. oryzicola. Phytopathology 2018, 108, 542–551. [Google Scholar] [CrossRef] [PubMed]
- Jacob-Dubuisson, F.; Mechaly, A.; Betton, J.M.; Antoine, R. Structural insights into the signalling mechanisms of two-component systems. Nat. Rev. Microbiol. 2018, 16, 585–593. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhao, M.; Xu, L.; Niu, X.; Qin, H.; Li, Y.; Li, M.; Jiang, Z.; Yang, X.; Huang, G.; et al. Genome-wide screening for novel candidate virulence related response regulator genes in Xanthomonas oryzae pv. oryzicola. Front. Microbiol. 2018, 9, 1789. [Google Scholar] [CrossRef]
- Wei, C.; Wang, S.; Liu, P.; Cheng, S.T.; Qian, G.; Wang, S.; Fu, Y.; Qian, W.; Sun, W. The PdeK-PdeR two-component system promotes unipolar localization of FimX and pilus extension in Xanthomonas oryzae pv. oryzicola. Sci. Signal. 2021, 14, eabi9589. [Google Scholar] [CrossRef]
- Cai, L.; Ma, W.; Zou, L.; Xu, X.; Xu, Z.; Deng, C.; Qian, W.; Chen, X.; Chen, G. Xanthomonas oryzae pv. oryzicola response regulator VemR is co-opted by the sensor kinase CheA for phosphorylation of multiple pathogenicity-related targets. Front. Microbiol. 2022, 13, 928551. [Google Scholar] [CrossRef]
- Zou, H.; Song, X.; Zou, L.; Yuan, L.; Li, Y.; Guo, W.; Che, Y.; Zhao, W.; Duan, Y.; Chen, G. EcpA, an extracellular protease, is a specific virulence factor required by Xanthomonas oryzae pv. oryzicola but not by X. oryzae pv. oryzae in rice. Microbiology 2012, 158, 2372–2383. [Google Scholar] [CrossRef]
- Kalunke, R.M.; Tundo, S.; Benedetti, M.; Cervone, F.; De Lorenzo, G.; D’Ovidio, R. An update on polygalacturonase-inhibiting protein (PGIP), a leucine-rich repeat protein that protects crop plants against pathogens. Front. Plant Sci. 2015, 6, 146. [Google Scholar] [CrossRef]
- Davidsson, P.; Broberg, M.; Kariola, T.; Sipari, N.; Pirhonen, M.; Palva, E.T. Short oligogalacturonides induce pathogen resistance-associated gene expression in Arabidopsis thaliana. BMC Plant Biol. 2017, 17, 19. [Google Scholar] [CrossRef]
- Wu, T.; Peng, C.; Li, B.; Wu, W.; Kong, L.; Li, F.; Chu, Z.; Liu, F.; Ding, X. OsPGIP1-mediated resistance to bacterial leaf streak in rice is beyond responsive to the polygalacturonase of Xanthomonas oryzae pv. oryzicola. Rice 2019, 12, 90. [Google Scholar] [CrossRef]
- Cai, L.; Zou, L.; Ling, G.; Xue, X.; Zou, H.; Chen, G. An inner membrane protein (Imp) of Xanthomonas oryzae pv. oryzicola functions in carbon acquisition, EPS production, bacterial motility and virulence in rice. J. Integr. Agric. 2014, 13, 2656–2668. [Google Scholar] [CrossRef]
- Xu, H.; Zhao, Y.; Qian, G.; Liu, F. XocR, a LuxR solo required for virulence in Xanthomonas oryzae pv. oryzicola. Front. Cell. Infect. Microbiol. 2015, 5, 37. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Wang, H.; Liu, X.; Xin, D.; Rao, Y.; Zhu, B. Transcriptome analysis of Xanthomonas oryzae pv. oryzicola exposed to H2O2 reveals horizontal gene transfer contributes to its oxidative stress response. PLoS ONE 2019, 14, e0218844. [Google Scholar] [CrossRef]
- Wang, P.; Wang, S.; Nie, W.; Wu, Y.; Iftikhar, A.; Ayizekeranmu, Y.; Huang, J.; Chen, G.; Zhu, B. A transferred regulator that contributes to Xanthomonas oryzae pv. oryzicola oxidative stress adaptation and virulence by regulating the expression of cytochrome bd oxidase genes. J. Integr. Agric. 2022, 21, 1673–1682. [Google Scholar] [CrossRef]
- Arroyo-Velez, N.; González-Fuente, M.; Peeters, N.; Lauber, E.; Noël, L.D. From effectors to effectomes: Are functional studies of individual effectors enough to decipher plant pathogen infectious strategies? PLoS Pathog. 2020, 16, e1009059. [Google Scholar] [CrossRef]
- Xu, Z.; Xu, X.; Gong, Q.; Li, Z.; Li, Y.; Wang, S.; Yang, Y.; Ma, W.; Liu, L.; Zhu, B.; et al. Engineering broad-spectrum bacterial blight resistance by simultaneously disrupting variable TALE-binding elements of multiple susceptibility genes in rice. Mol. Plant 2019, 12, 1434–1446. [Google Scholar] [CrossRef]
- Oliva, R.; Ji, C.; Atienza-Grande, G.; Huguet-Tapia, J.C.; Perez-Quintero, A.; Li, T.; Eom, J.S.; Li, C.; Nguyen, H.; Liu, B.; et al. Broad-spectrum resistance to bacterial blight in rice using genome editing. Nat. Biotechnol. 2019, 37, 1344–1350. [Google Scholar] [CrossRef]
- Ni, Z.; Cao, Y.; Jin, X.; Fu, Z.; Li, J.; Mo, X.; He, Y.; Tang, J.; Huang, S. Engineering resistance to bacterial blight and bacterial leaf streak in rice. Rice 2021, 14, 38. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Fang, Y.; Wu, H.; Zhao, N.; Guo, X.; Mackon, E.; Peng, H.; Huang, S.; He, Y.; Qin, B.; et al. Improvement of resistance to rice blast and bacterial leaf streak by CRISPR/Cas9-mediated mutagenesis of Pi21 and OsSULTR3;6 in rice (Oryza sativa L.). Front. Plant Sci. 2023, 14, 1209384. [Google Scholar] [CrossRef] [PubMed]
- Scinto-Madonich, N.J.; Baruah, S.; Young, S.; Vignona, K.; Read, A.C.; Carpenter, S.C.D.; Wang, L.; Shi, X.; Chang, G.; Piñeros, M.A.; et al. Initial characterization of a bacterial leaf streak susceptibility gene suggests it encodes a membrane transporter that influences seed nutrition and germination. Physiol. Mol. Plant Pathol. 2023, 126, 102031. [Google Scholar] [CrossRef]
- Shafique, M.S.; Liu, Y.; Li, M.; Wang, H.; Su, R.; Wang, C.; Ji, Z. Coevolution unveiled: Sulfate transporters mediate rice resistance and susceptibility to Xanthomonas oryzae pv. oryzicola. Plant Biotechnol. J. 2024, 22, 2632–2634. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).