
2-Keto-L-Gulonic Acid Enhances Cold Stress Tolerance in Arabidopsis thaliana via Augmentation of Ascorbic Acid Biosynthesis and Antioxidant Defense Systems



Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Growth and Experimental Design
2.2. Growth Parameters and ASA Content
2.3. Photosynthetic Pigments
2.4. Contents of Malondialdehyde, Proline, Soluble Carbohydrate, and Soluble Protein
2.5. Antioxidant Enzyme Activity
2.6. Gene Expression Analysis
2.7. Statistical Analysis
3. Results
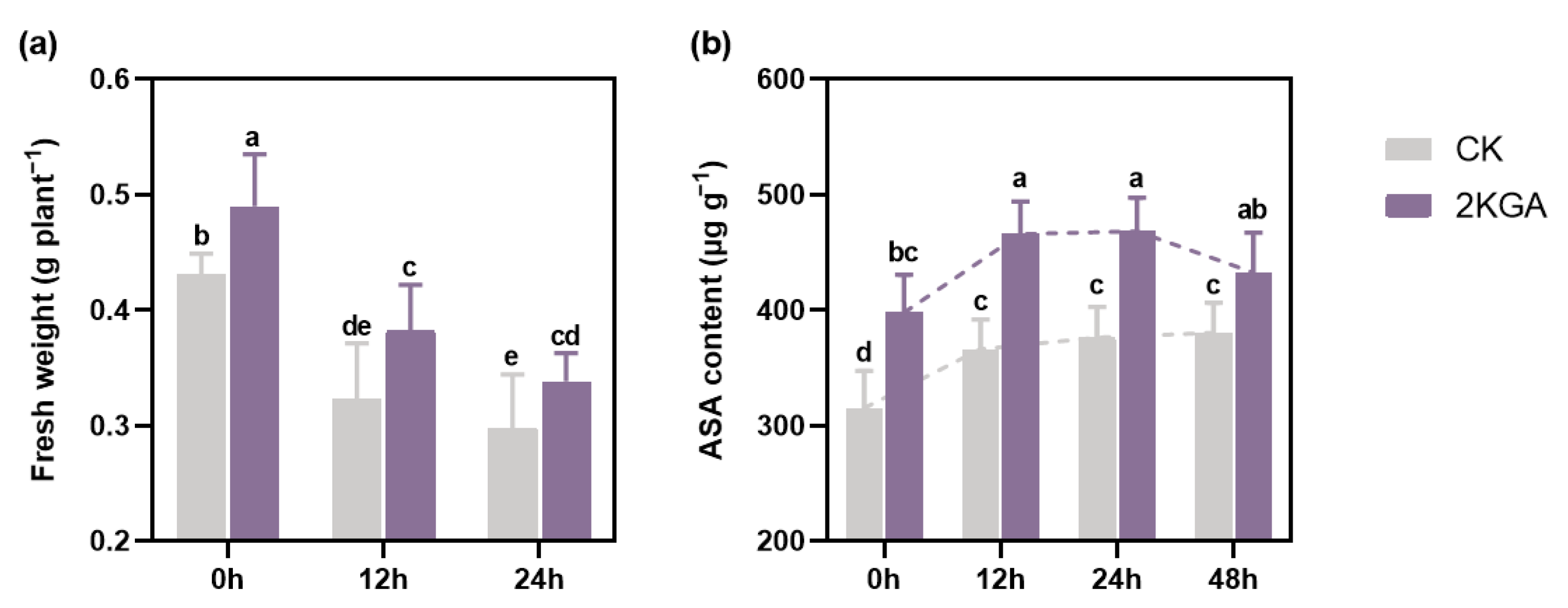
3.1. Effect of 2KGA on Plant Growth Parameters under Cold Stress
3.2. Effect of 2KGA on ASA Content under Cold Stress
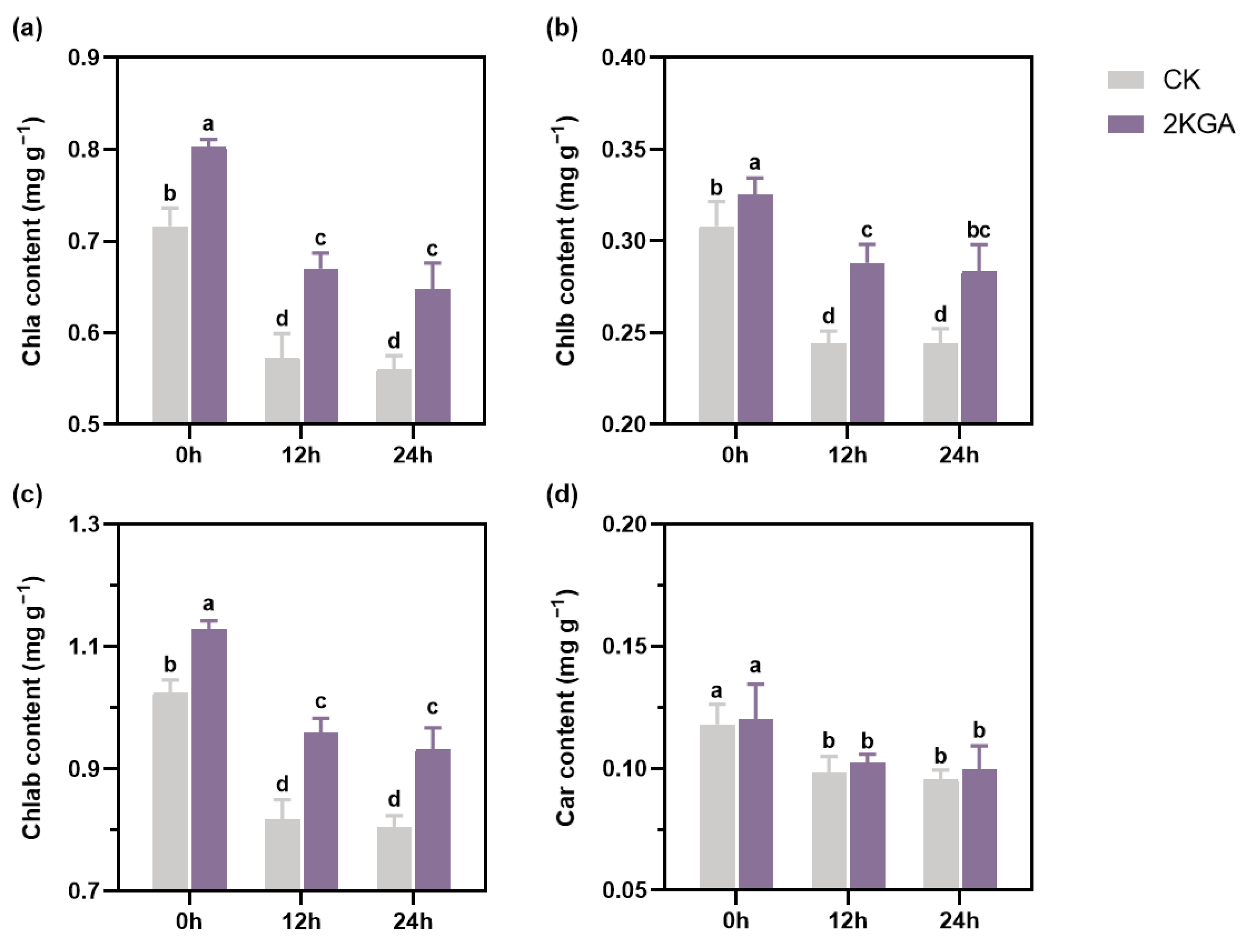
3.3. Effect of 2KGA on Photosynthetic Pigments under Cold Stress
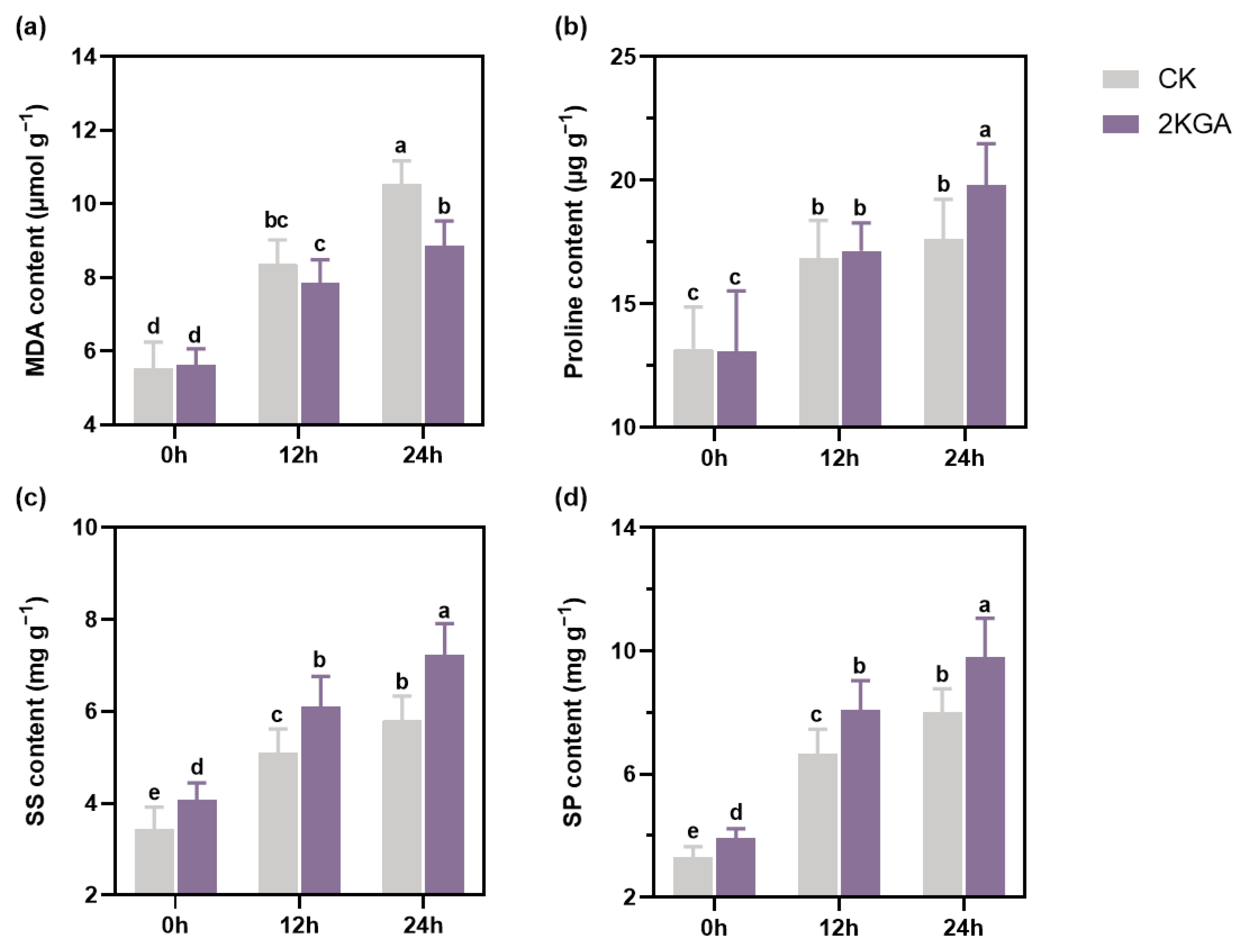
3.4. Effect of 2KGA on MDA, Proline, SS, and SP under Cold Stress
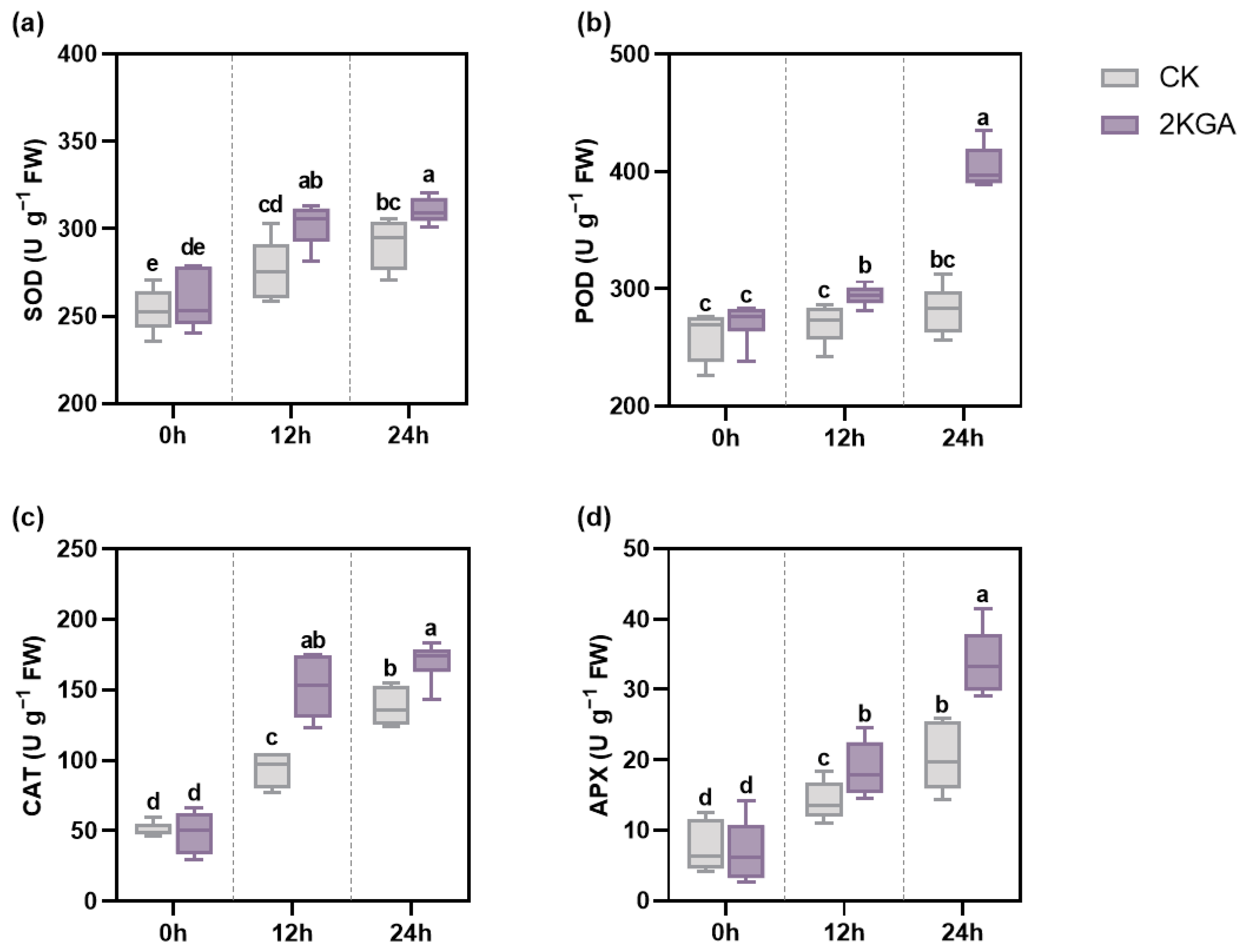
3.5. Effect of 2KGA on Enzymatic Antioxidant System under Cold Stress
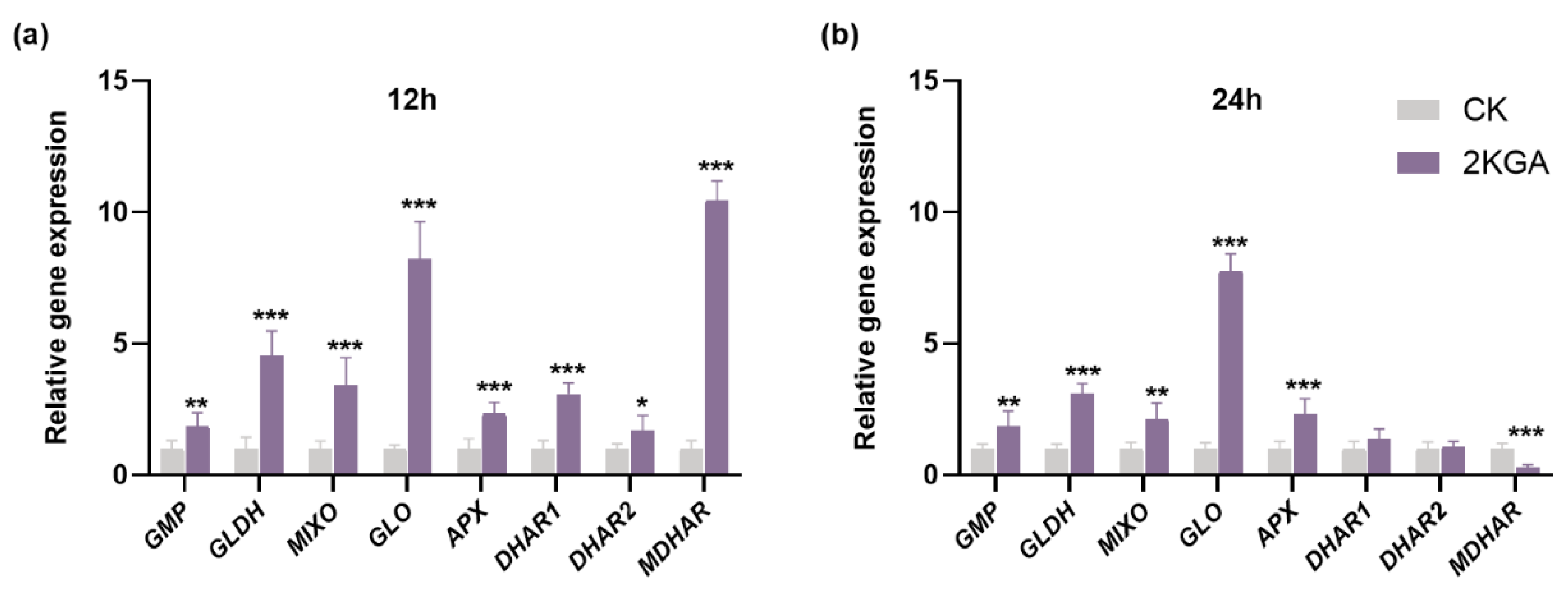
3.6. Effect of 2KGA on ASA Metabolism-Related Gene Expression under Cold Stress
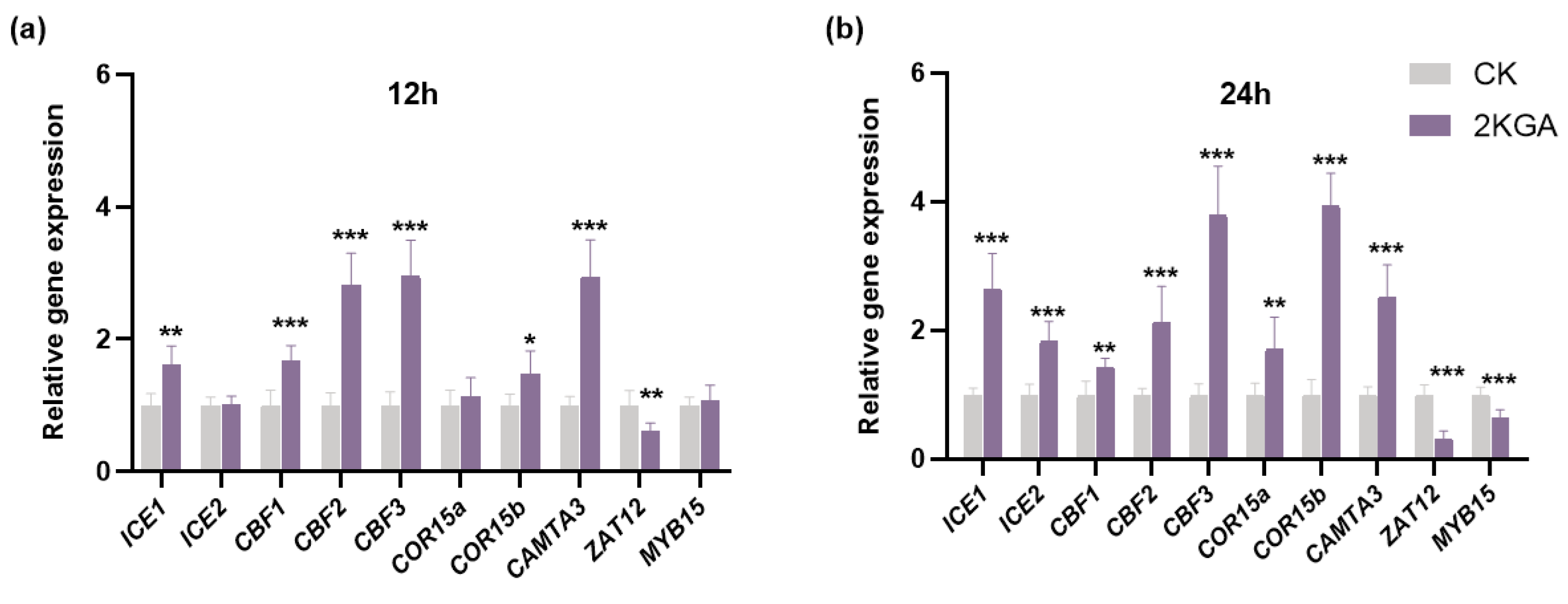
3.7. Effect of 2KGA on Cold Stress-Related Gene Expression under Cold Stress
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Fedoroff, N.V.; Battisti, D.S.; Beachy, R.N.; Cooper, P.J.M.; Fischhoff, D.A.; Hodges, C.N.; Knauf, V.C.; Lobell, D.; Mazur, B.J.; Molden, D.; et al. Radically Rethinking Agriculture for the 21st Century. Science 2010, 327, 833–834. [Google Scholar] [CrossRef]
- Nadarajah, K.K. ROS Homeostasis in Abiotic Stress Tolerance in Plants. Int. J. Mol. Sci. 2020, 21, 5208. [Google Scholar] [CrossRef]
- Li, S. Novel Insight into Functions of Ascorbate Peroxidase in Higher Plants: More than a Simple Antioxidant Enzyme. Redox Biol. 2023, 64, 102789. [Google Scholar] [CrossRef]
- Rudenko, N.N.; Vetoshkina, D.V.; Marenkova, T.V.; Borisova-Mubarakshina, M.M. Antioxidants of Non-Enzymatic Nature: Their Function in Higher Plant Cells and the Ways of Boosting Their Biosynthesis. Antioxidants 2023, 12, 2014. [Google Scholar] [CrossRef]
- Bajwa, V.S.; Shukla, M.R.; Sherif, S.M.; Murch, S.J.; Saxena, P.K. Role of Melatonin in Alleviating Cold Stress in Arabidopsis thaliana. J. Pineal Res. 2014, 56, 238–245. [Google Scholar] [CrossRef]
- Hwarari, D.; Guan, Y.; Ahmad, B.; Movahedi, A.; Min, T.; Hao, Z.; Lu, Y.; Chen, J.; Yang, L. ICE-CBF-COR Signaling Cascade and Its Regulation in Plants Responding to Cold Stress. Int. J. Mol. Sci. 2022, 23, 1549. [Google Scholar] [CrossRef]
- Gusain, S.; Joshi, S.; Joshi, R. Sensing, Signalling, and Regulatory Mechanism of Cold-Stress Tolerance in Plants. Plant Physiol. Biochem. 2023, 197, 107646. [Google Scholar] [CrossRef]
- Wang, L.; Ma, R.; Yin, Y.; Jiao, Z. Role of Carbon Ion Beams Irradiation in Mitigating Cold Stress in Arabidopsis thaliana. Ecotoxicol. Environ. Saf. 2018, 162, 341–347. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Mahmud, J.A.; Fujita, M.; Fotopoulos, V. Reactive Oxygen Species and Antioxidant Defense in Plants under Abiotic Stress: Revisiting the Crucial Role of a Universal Defense Regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef]
- Jalili, I.; Ebadi, A.; Askari, M.A.; KalatehJari, S.; Aazami, M.A. Foliar Application of Putrescine, Salicylic Acid, and Ascorbic Acid Mitigates Frost Stress Damage in Vitis vinifera Cv. ‘Giziluzum’. BMC Plant Biol. 2023, 23, 135. [Google Scholar] [CrossRef]
- Broad, R.C.; Bonneau, J.P.; Hellens, R.P.; Johnson, A.A.T. Manipulation of Ascorbate Biosynthetic, Recycling, and Regulatory Pathways for Improved Abiotic Stress Tolerance in Plants. Int. J. Mol. Sci. 2020, 21, 1790. [Google Scholar] [CrossRef] [PubMed]
- Lisko, K.A.; Aboobucker, S.I.; Torres, R.; Lorence, A. Engineering Elevated Vitamin C in Plants to Improve Their Nutritional Content, Growth, and Tolerance to Abiotic Stress. In Phytochemicals—Biosynthesis, Function and Application; Jetter, R., Ed.; Springer International Publishing: Cham, Switzerland, 2014; Volume 44, pp. 109–128. ISBN 978-3-319-04045-5. [Google Scholar]
- Ali, S.; Nawaz, A.; Hussain, S.; Khan, S.M.; Ejaz, S.; Ahmad, S. Abiotic Stress Tolerance in Plants by Priming and Pretreatments with Ascorbic Acid. In Priming and Pretreatment of Seeds and Seedlings: Implication in Plant Stress Tolerance and Enhancing Productivity in Crop Plants; Hasanuzzaman, M., Fotopoulos, V., Eds.; Springer: Singapore, 2019; pp. 459–493. ISBN 9789811386251. [Google Scholar]
- Noreen, S.; Sultan, M.; Akhter, M.S.; Shah, K.H.; Ummara, U.; Manzoor, H.; Ulfat, M.; Alyemeni, M.N.; Ahmad, P. Foliar Fertigation of Ascorbic Acid and Zinc Improves Growth, Antioxidant Enzyme Activity and Harvest Index in Barley (Hordeum vulgare L.) Grown under Salt Stress. Plant Physiol. Biochem. 2021, 158, 244–254. [Google Scholar] [CrossRef] [PubMed]
- Sharma, L.; Roy, S.; Satya, P.; Alam, N.M.; Goswami, T.; Barman, D.; Bera, A.; Saha, R.; Mitra, S.; Mitra, J. Exogenous Ascorbic Acid Application Ameliorates Drought Stress through Improvement in Morpho-Physiology, Nutrient Dynamics, Stress Metabolite Production and Antioxidant Activities Recovering Cellulosic Fibre Production in Jute (Corchorus olitorius L.). Ind. Crops Prod. 2024, 217, 118808. [Google Scholar] [CrossRef]
- Liu, X.; Bulley, S.M.; Varkonyi-Gasic, E.; Zhong, C.; Li, D. Kiwifruit bZIP Transcription Factor AcePosF21 Elicits Ascorbic Acid Biosynthesis during Cold Stress. Plant Physiol. 2023, 192, 982–999. [Google Scholar] [CrossRef]
- Elkelish, A.; Qari, S.H.; Mazrou, Y.S.A.; Abdelaal, K.A.A.; Hafez, Y.M.; Abu-Elsaoud, A.M.; Batiha, G.E.-S.; El-Esawi, M.A.; El Nahhas, N. Exogenous Ascorbic Acid Induced Chilling Tolerance in Tomato Plants Through Modulating Metabolism, Osmolytes, Antioxidants, and Transcriptional Regulation of Catalase and Heat Shock Proteins. Plants 2020, 9, 431. [Google Scholar] [CrossRef]
- Xiang, N.; Hu, J.; Wen, T.; Brennan, M.A.; Brennan, C.S.; Guo, X. Effects of Temperature Stress on the Accumulation of Ascorbic Acid and Folates in Sweet Corn (Zea mays L.) Seedlings. J. Sci. Food Agric. 2020, 100, 1694–1701. [Google Scholar] [CrossRef]
- Fu, Q.; Cao, H.; Wang, L.; Lei, L.; Di, T.; Ye, Y.; Ding, C.; Li, N.; Hao, X.; Zeng, J.; et al. Transcriptome Analysis Reveals That Ascorbic Acid Treatment Enhances the Cold Tolerance of Tea Plants through Cell Wall Remodeling. Int. J. Mol. Sci. 2023, 24, 10059. [Google Scholar] [CrossRef]
- Jha, U.C.; Bohra, A.; Jha, R. Breeding Approaches and Genomics Technologies to Increase Crop Yield under Low-Temperature Stress. Plant Cell Rep. 2017, 36, 1–35. [Google Scholar] [CrossRef]
- Bulley, S.M.; Cooney, J.M.; Laing, W. Elevating Ascorbate in Arabidopsis Stimulates the Production of Abscisic Acid, Phaseic Acid, and to a Lesser Extent Auxin (IAA) and Jasmonates, Resulting in Increased Expression of DHAR1 and Multiple Transcription Factors Associated with Abiotic Stress Tolerance. Int. J. Mol. Sci. 2021, 22, 6743. [Google Scholar] [CrossRef]
- Yang, W.; Han, L.; Mandlaa, M.; Zhang, H.; Zhang, Z.; Xu, H. A Plate Method for Rapid Screening of Ketogulonicigenium vulgare Mutants for Enhanced 2-Keto-l-Gulonic Acid Production. Braz. J. Microbiol. 2017, 48, 397–402. [Google Scholar] [CrossRef]
- Kong, T.; Xu, H.; Wang, Z.; Sun, H.; Wang, L. Effect of a Residue after Evaporation from Industrial Vitamin C Fermentation on Chemical and Microbial Properties of Alkali-Saline Soil. Pak. J. Pharm. Sci. 2014, 27, 1069–1074. [Google Scholar] [PubMed]
- Wang, B.; Sun, H.; Yang, W.; Gao, M.; Zhong, X.; Zhang, L.; Chen, Z.; Xu, H. Potential Utilization of Vitamin C Industrial Effluents in Agriculture: Soil Fertility and Bacterial Community Composition. Sci. Total Environ. 2022, 851, 158253. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.; Zhang, Z.; Yang, W.; Sun, H.; Xu, H. Application of Organic Waste Derived from Vitamin C Industry Increases Yield and Bioactive Constituents of Medicinal Food Plant Purslane (Portulaca oleracea L.). Horticulturae 2024, 10, 683. [Google Scholar] [CrossRef]
- Gao, M.; Han, X.; Yang, W.; Sun, H.; Zhang, L.; Xu, H. A Strategy for Improving Saline-Alkali Soil Properties and Cotton Stress Tolerance Using Vitamin C Industrial Wastes: A “Prebiotics-Probiotics” Interaction between Organic Acids and Bacillus Endophyticus. Ind. Crops Prod. 2024, 220, 119187. [Google Scholar] [CrossRef]
- Shi, M.; Gao, M.; Sun, H.; Yang, W.; Zhao, H.; Zhang, L.; Xu, H. Exogenous 2-Keto-L-Gulonic Acid Supplementation as a Novel Approach to Enhancing L-Ascorbic Acid Biosynthesis in Zebrafish (Danio rerio). Animals 2023, 13, 2502. [Google Scholar] [CrossRef]
- Gao, M.; Sun, H.; Shi, M.; Wu, Q.; Ji, D.; Wang, B.; Zhang, L.; Liu, Y.; Han, L.; Ruan, X.; et al. 2-Keto-L-Gulonic Acid Improved the Salt Stress Resistance of Non-Heading Chinese Cabbage by Increasing L-Ascorbic Acid Accumulation. Front. Plant Sci. 2021, 12, 697184. [Google Scholar] [CrossRef]
- Sivasubramanian, R.; Mukhi, N.; Kaur, J. Arabidopsis Thaliana: A Model for Plant Research. In Plant Biology and Biotechnology: Volume II: Plant Genomics and Biotechnology; Bahadur, B., Venkat Rajam, M., Sahijram, L., Krishnamurthy, K.V., Eds.; Springer India: New Delhi, India, 2015; pp. 1–26. ISBN 978-81-322-2283-5. [Google Scholar]
- Lichtenthaler, H.K. [34] Chlorophylls and Carotenoids: Pigments of Photosynthetic Biomembranes. In Methods in Enzymology; Plant Cell Membranes; Academic Press: London, UK, 1987; Volume 148, pp. 350–382. [Google Scholar]
- Ali, S.; Gill, R.A.; Ulhassan, Z.; Zhang, N.; Hussain, S.; Zhang, K.; Huang, Q.; Sagir, M.; Tahir, M.B.; Gill, M.B.; et al. Exogenously Applied Melatonin Enhanced the Tolerance of Brassica Napus against Cobalt Toxicity by Modulating Antioxidant Defense, Osmotic Adjustment, and Expression of Stress Response Genes. Ecotoxicol. Environ. Saf. 2023, 252, 114624. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid Determination of Free Proline for Water-Stress Studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Zhang, W.H.; Wu, J.; Weng, L.; Zhang, H.; Zhang, J.; Wu, A. An Improved Phenol-Sulfuric Acid Method for the Determination of Carbohydrates in the Presence of Persulfate. Carbohydr. Polym. 2020, 227, 115332. [Google Scholar] [CrossRef]
- Mm, B. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Xiang, H.; Wang, S.; Liang, X.; Wang, X.; Xie, H.; Wang, D.; Gai, Z.; Wang, N.; Xiang, P.; Han, D.; et al. Foliar Spraying of Exogenous Uniconazole (S3307) at the Flowering Stage as an Effective Method to Resist Low-Temperature Stress on Mung Bean [Vigna radiata (L.) Wilczek]. Sci. Rep. 2023, 13, 22331. [Google Scholar] [CrossRef] [PubMed]
- Beauchamp, C.; Fridovich, I. Superoxide Dismutase: Improved Assays and an Assay Applicable to Acrylamide Gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef] [PubMed]
- Castillo, F.J.; Penel, C.; Greppin, H. Peroxidase Release Induced by Ozone in Sedum album Leaves: Involvement of Ca2+. Plant Physiol. 1984, 74, 846–851. [Google Scholar] [CrossRef]
- Anderson, M.D.; Prasad, T.K.; Stewart, C.R. Changes in Isozyme Profiles of Catalase, Peroxidase, and Glutathione Reductase during Acclimation to Chilling in Mesocotyls of Maize Seedlings. Plant Physiol. 1995, 109, 1247–1257. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen Peroxide Is Scavenged by Ascorbate-Specific Peroxidase in Spinach Chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing Real-Time PCR Data by the Comparative C(T) Method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Raza, A.; Bashir, S.; Khare, T.; Karikari, B.; Copeland, R.G.R.; Jamla, M.; Abbas, S.; Charagh, S.; Nayak, S.N.; Djalovic, I.; et al. Temperature-Smart Plants: A New Horizon with Omics-Driven Plant Breeding. Physiol. Plant. 2024, 176, e14188. [Google Scholar] [CrossRef]
- Celi, G.E.A.; Gratão, P.L.; Lanza, M.G.D.B.; Reis, A.R.D. Physiological and Biochemical Roles of Ascorbic Acid on Mitigation of Abiotic Stresses in Plants. Plant Physiol. Biochem. 2023, 202, 107970. [Google Scholar] [CrossRef]
- Cheng, P.; Feng, L.; Zhang, S.; Li, L.; Guan, R.; Long, W.; Xian, Z.; Zhang, J.; Shen, W. Ammonia Borane Positively Regulates Cold Tolerance in Brassica Napus via Hydrogen Sulfide Signaling. BMC Plant Biol. 2022, 22, 585. [Google Scholar] [CrossRef]
- Dai, T.; Ban, S.; Han, L.; Li, L.; Zhang, Y.; Zhang, Y.; Zhu, W. Effects of Exogenous Glycine Betaine on Growth and Development of Tomato Seedlings under Cold Stress. Front. Plant Sci. 2024, 15, 1332583. [Google Scholar] [CrossRef]
- Wang, Q.; Shi, H.; Huang, R.; Ye, R.; Luo, Y.; Guo, Z.; Lu, S. AIR12 Confers Cold Tolerance through Regulation of the CBF Cold Response Pathway and Ascorbate Homeostasis. Plant Cell Environ. 2021, 44, 1522–1533. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Lu, T.; Yan, W.; Yu, P.; Fu, W.; Li, J.; Su, X.; Chen, T.; Fu, G.; Wu, Z.; et al. Transcriptome and Metabolome Analyses Reveal Ascorbic Acid Ameliorates Cold Tolerance in Rice Seedling Plants. Agronomy 2024, 14, 659. [Google Scholar] [CrossRef]
- Heidari, P.; Reza Amerian, M.; Barcaccia, G. Hormone Profiles and Antioxidant Activity of Cultivated and Wild Tomato Seedlings under Low-Temperature Stress. Agronomy 2021, 11, 1146. [Google Scholar] [CrossRef]
- Rahman, M.d.M.; Mostofa, M.G.; Rahman, M.d.A.; Islam, M.d.R.; Keya, S.S.; Das, A.K.; Miah, M.d.G.; Kawser, A.Q.M.R.; Ahsan, S.M.; Hashem, A.; et al. Acetic Acid: A Cost-Effective Agent for Mitigation of Seawater-Induced Salt Toxicity in Mung Bean. Sci. Rep. 2019, 9, 15186. [Google Scholar] [CrossRef]
- Mittler, R.; Vanderauwera, S.; Gollery, M.; Van Breusegem, F. Reactive Oxygen Gene Network of Plants. Trends Plant Sci. 2004, 9, 490–498. [Google Scholar] [CrossRef]
- Tang, K.; Zhao, L.; Ren, Y.; Yang, S.; Zhu, J.-K.; Zhao, C. The Transcription Factor ICE1 Functions in Cold Stress Response by Binding to the Promoters of CBF and COR Genes. J. Integr. Plant Biol. 2020, 62, 258–263. [Google Scholar] [CrossRef]
- Vogel, J.T.; Zarka, D.G.; Van Buskirk, H.A.; Fowler, S.G.; Thomashow, M.F. Roles of the CBF2 and ZAT12 Transcription Factors in Configuring the Low Temperature Transcriptome of Arabidopsis. Plant J. 2005, 41, 195–211. [Google Scholar] [CrossRef]
- Doherty, C.J.; Van Buskirk, H.A.; Myers, S.J.; Thomashow, M.F. Roles for Arabidopsis CAMTA Transcription Factors in Cold-Regulated Gene Expression and Freezing Tolerance. Plant Cell 2009, 21, 972–984. [Google Scholar] [CrossRef]
- Wang, X.; Ding, Y.; Li, Z.; Shi, Y.; Wang, J.; Hua, J.; Gong, Z.; Zhou, J.-M.; Yang, S. PUB25 and PUB26 Promote Plant Freezing Tolerance by Degrading the Cold Signaling Negative Regulator MYB15. Dev. Cell 2019, 51, 222–235.e5. [Google Scholar] [CrossRef]
- Ritonga, F.N.; Chen, S. Physiological and Molecular Mechanism Involved in Cold Stress Tolerance in Plants. Plants 2020, 9, 560. [Google Scholar] [CrossRef]
- Xu, Y.; Huang, B. Exogenous Ascorbic Acid Mediated Abiotic Stress Tolerance in Plants. In Ascorbic Acid in Plant Growth, Development and Stress Tolerance; Hossain, M.A., Munné-Bosch, S., Burritt, D.J., Diaz-Vivancos, P., Fujita, M., Lorence, A., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 233–253. ISBN 978-3-319-74057-7. [Google Scholar]
- Liang, C.; Prins, T.W.; van de Wiel, C.C.M.; Kok, E.J. Safety Aspects of Genetically Modified Crops with Abiotic Stress Tolerance. Trends Food Sci. Technol. 2014, 40, 115–122. [Google Scholar] [CrossRef]
- Hu, Z.; Fan, J.; Xie, Y.; Amombo, E.; Liu, A.; Gitau, M.M.; Khaldun, A.B.M.; Chen, L.; Fu, J. Comparative Photosynthetic and Metabolic Analyses Reveal Mechanism of Improved Cold Stress Tolerance in Bermudagrass by Exogenous Melatonin. Plant Physiol. Biochem. 2016, 100, 94–104. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Li, D.; Wang, Q.; Song, X.; Wang, Y.; Yang, X.; Qin, D.; Xie, T.; Yang, D. Exogenous Salicylic Acid Improves Chilling Tolerance in Maize Seedlings by Improving Plant Growth and Physiological Characteristics. Agronomy 2021, 11, 1341. [Google Scholar] [CrossRef]
- Sun, H.; Rang, X.; Han, H.; Pei, Z.; Zhao, J.; Zhu, Z.; Li, J.; Zhang, P.; Zhao, Y.; Duan, Y. Exogenous Gibberellic Acid Ameliorates Chilling Injury in Peach (Prunus persica L.) by Improving the Antioxidant System. Agronomy 2024, 14, 816. [Google Scholar] [CrossRef]
- Savvides, A.; Ali, S.; Tester, M.; Fotopoulos, V. Chemical Priming of Plants Against Multiple Abiotic Stresses: Mission Possible? Trends Plant Sci. 2016, 21, 329–340. [Google Scholar] [CrossRef]
- Jaglo, K.R.; Kleff, S.; Amundsen, K.L.; Zhang, X.; Haake, V.; Zhang, J.Z.; Deits, T.; Thomashow, M.F. Components of the Arabidopsis C-Repeat/Dehydration-Responsive Element Binding Factor Cold-Response Pathway Are Conserved inBrassica Napus and Other Plant Species. Plant Physiol. 2001, 127, 910–917. [Google Scholar] [CrossRef]
- Jiang, F.; Wang, F.; Wu, Z.; Li, Y.; Shi, G.; Hu, J.; Hou, X. Components of the Arabidopsis CBF Cold-Response Pathway Are Conserved in Non-Heading Chinese Cabbage. Plant Mol. Biol. Rep. 2011, 29, 525–532. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time | Leaf Length (mm) | Leaf Width (mm) | Maximum Rosette Radius (mm) | |||
|---|---|---|---|---|---|---|
| CK | 2KGA | CK | 2KGA | CK | 2KGA | |
| 0 h | 20.53 ± 1.08 bc | 22.03 ± 0.60 a | 9.61 ± 0.33 a | 9.82 ± 0.38 a | 30.61 ± 1.33 bc | 32.79 ± 0.88 a |
| 12 h | 20.33 ± 0.42 c | 21.98 ± 0.58 a | 9.54 ± 0.34 a | 9.77 ± 0.31 a | 28.94 ± 1.69 cd | 31.34 ± 1.13 b |
| 24 h | 20.06 ± 1.09 c | 21.54 ± 0.94 ab | 9.19 ± 0.52 a | 9.73 ± 0.23 a | 28.85 ± 0.69 d | 31.18 ± 0.99 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Q.; Gao, M.; Sun, H.; Yang, W.; Xu, H. 2-Keto-L-Gulonic Acid Enhances Cold Stress Tolerance in Arabidopsis thaliana via Augmentation of Ascorbic Acid Biosynthesis and Antioxidant Defense Systems. Agronomy 2024, 14, 2149. https://doi.org/10.3390/agronomy14092149
Wu Q, Gao M, Sun H, Yang W, Xu H. 2-Keto-L-Gulonic Acid Enhances Cold Stress Tolerance in Arabidopsis thaliana via Augmentation of Ascorbic Acid Biosynthesis and Antioxidant Defense Systems. Agronomy. 2024; 14(9):2149. https://doi.org/10.3390/agronomy14092149
Chicago/Turabian StyleWu, Qiqi, Mingfu Gao, Hao Sun, Weichao Yang, and Hui Xu. 2024. "2-Keto-L-Gulonic Acid Enhances Cold Stress Tolerance in Arabidopsis thaliana via Augmentation of Ascorbic Acid Biosynthesis and Antioxidant Defense Systems" Agronomy 14, no. 9: 2149. https://doi.org/10.3390/agronomy14092149
APA StyleWu, Q., Gao, M., Sun, H., Yang, W., & Xu, H. (2024). 2-Keto-L-Gulonic Acid Enhances Cold Stress Tolerance in Arabidopsis thaliana via Augmentation of Ascorbic Acid Biosynthesis and Antioxidant Defense Systems. Agronomy, 14(9), 2149. https://doi.org/10.3390/agronomy14092149