
Study on the Natural Ventilation Model of a Single-Span Plastic Greenhouse in a High-Altitude Area


Abstract
:1. Introduction
2. Materials and Methods
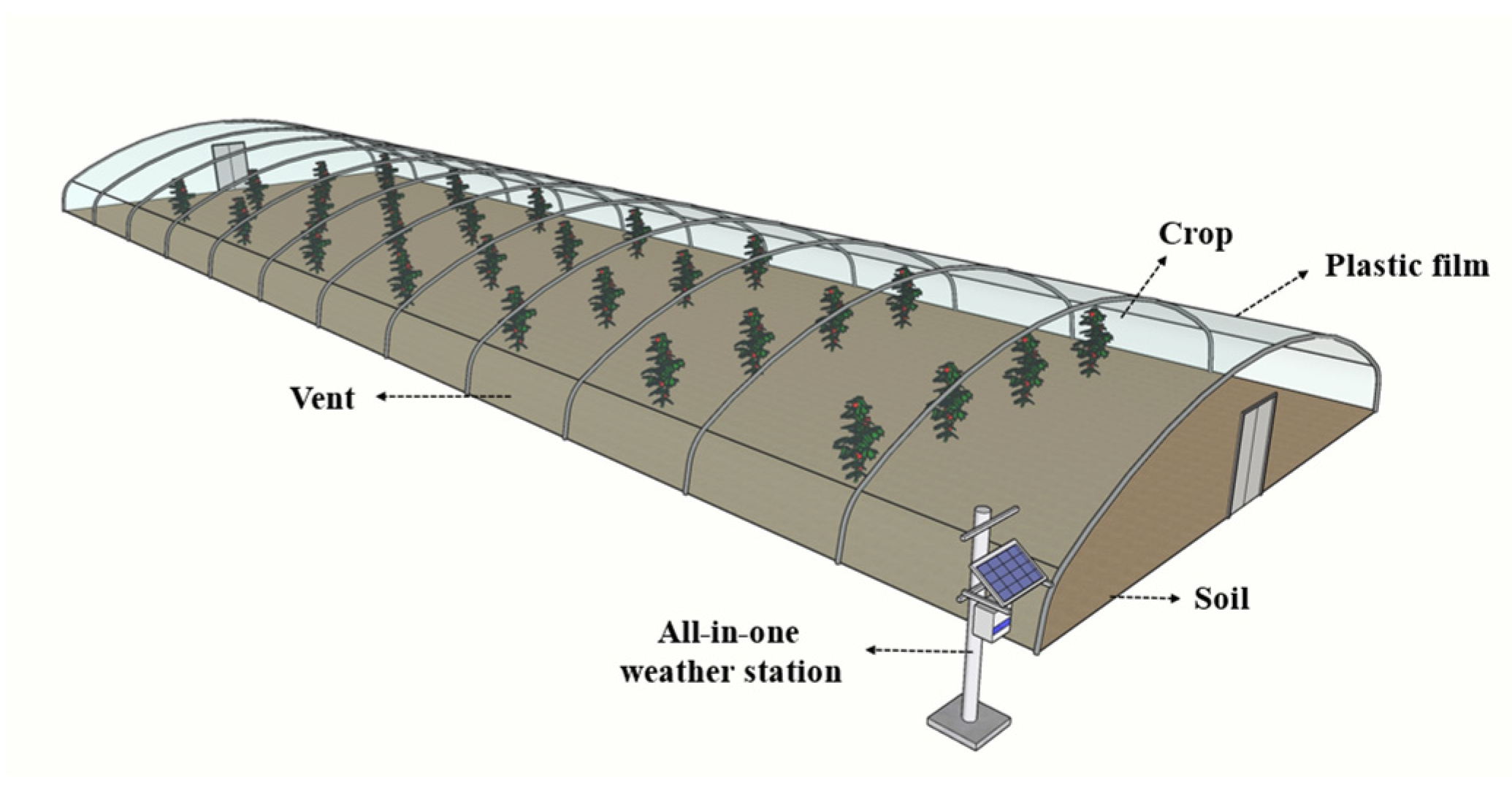
2.1. Description of the Experimental Greenhouse
2.2. Experimental Contents
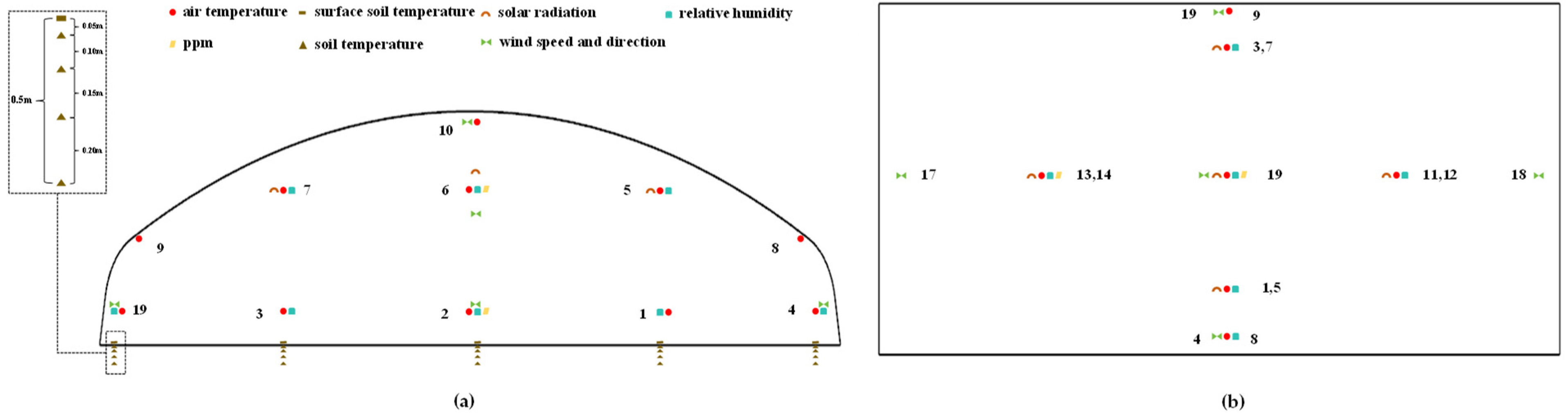
2.3. Environmental Data Collection
2.4. CFD Model
2.4.1. Basic Control Equations
2.4.2. Boussinesq Approximation
2.4.3. Turbulence Model
2.4.4. Solar Radiation Model
2.4.5. Species Transport Model
2.4.6. Crop Model
2.5. Numerical Calculation
2.5.1. Boundary Conditions and Calculation Parameters
2.5.2. Geometric Modeling and Grid Generation
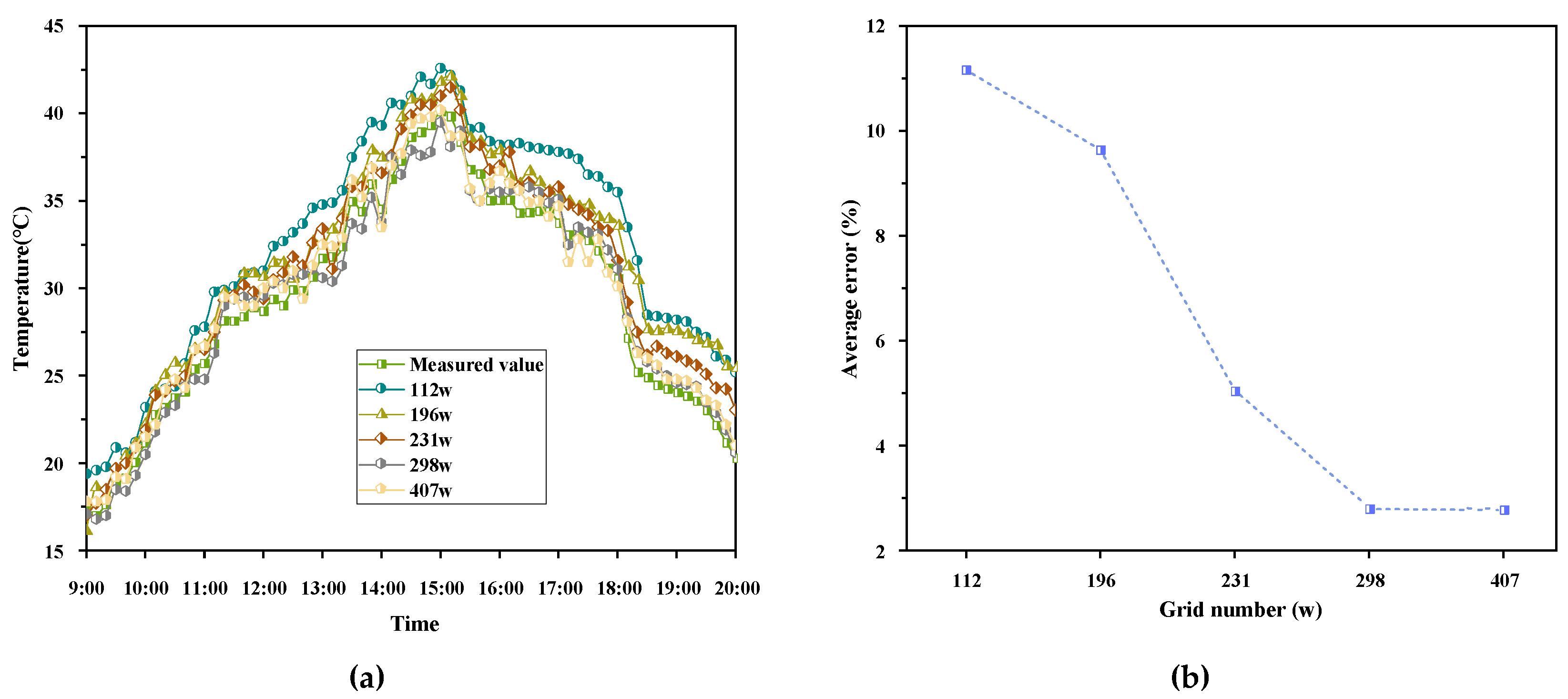
2.5.3. Grid Independence Validation
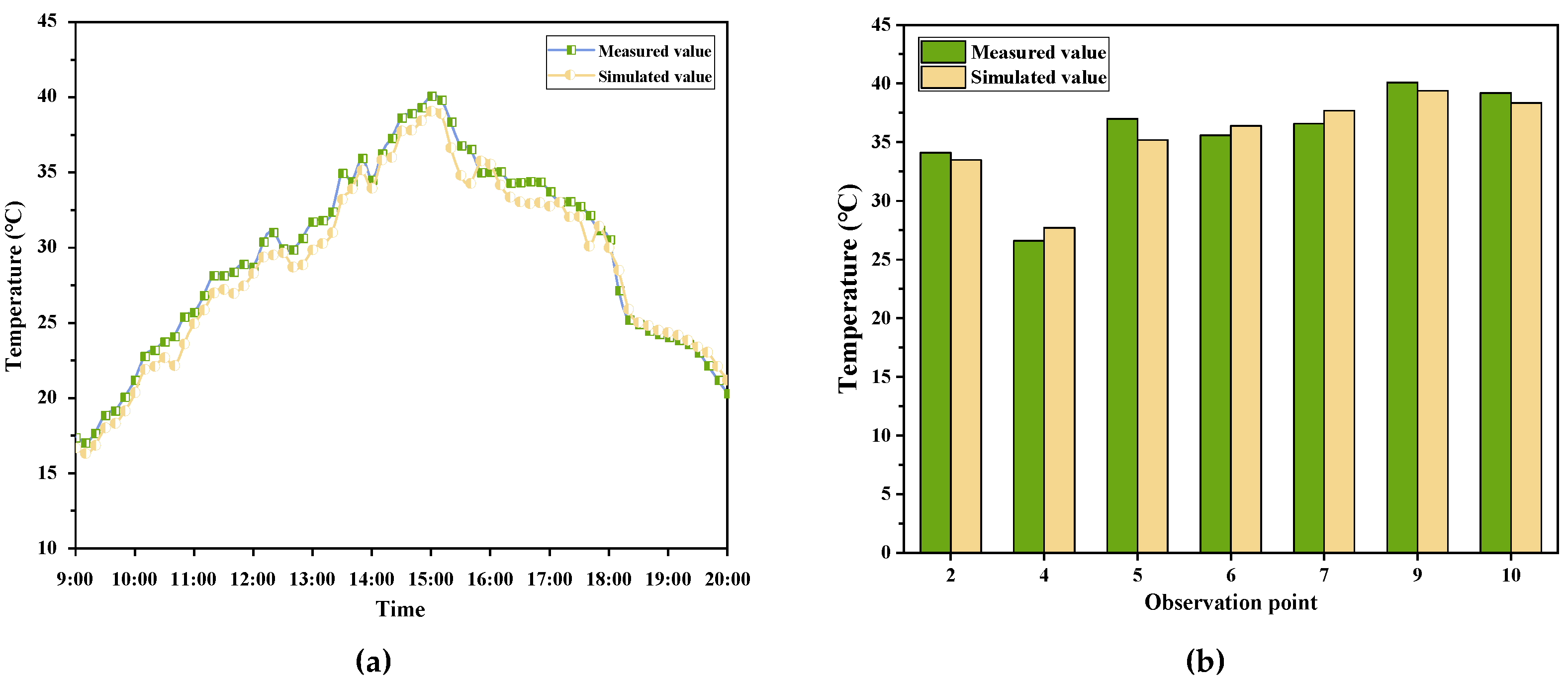
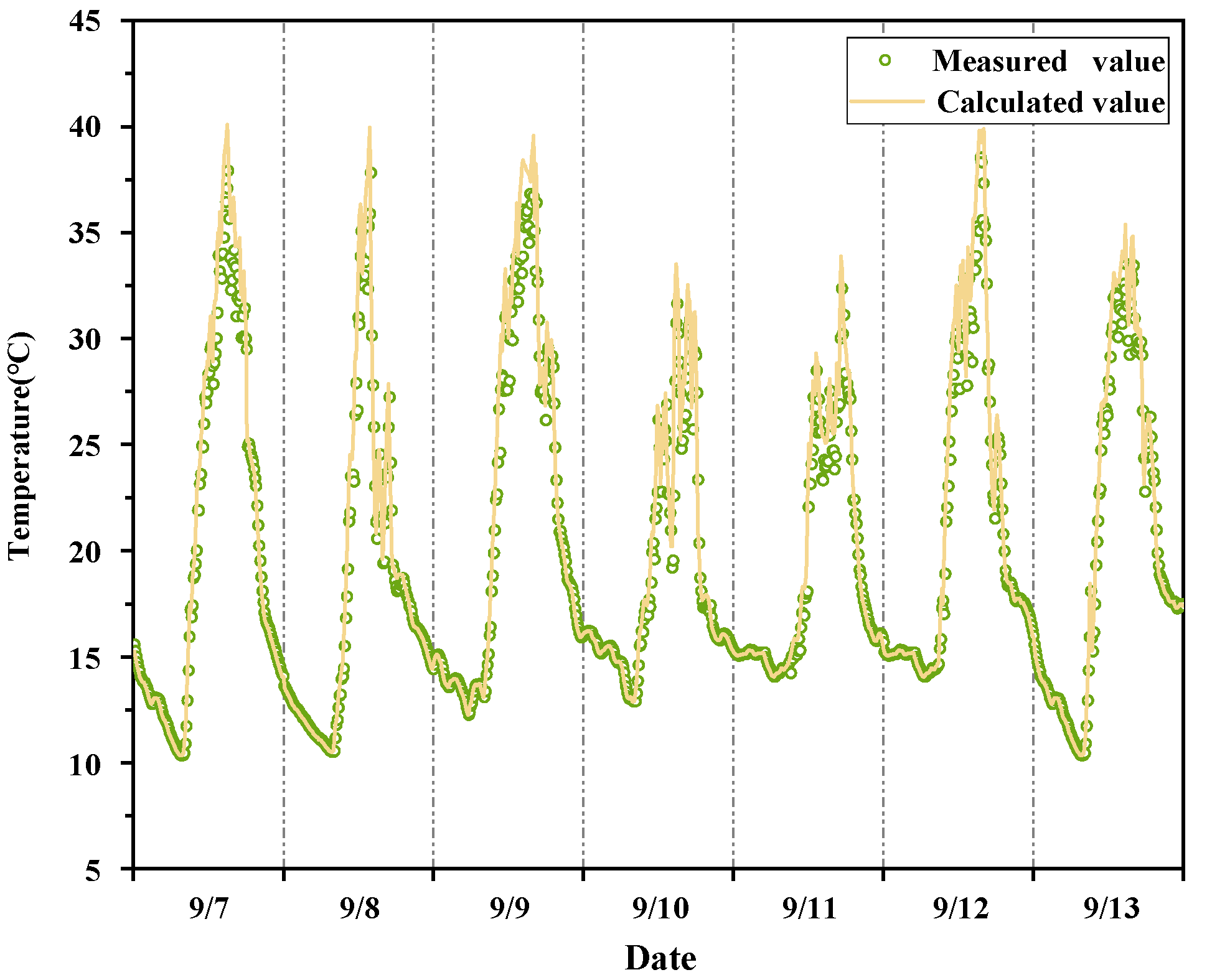
2.5.4. Model Validation
3. Results and Discussion
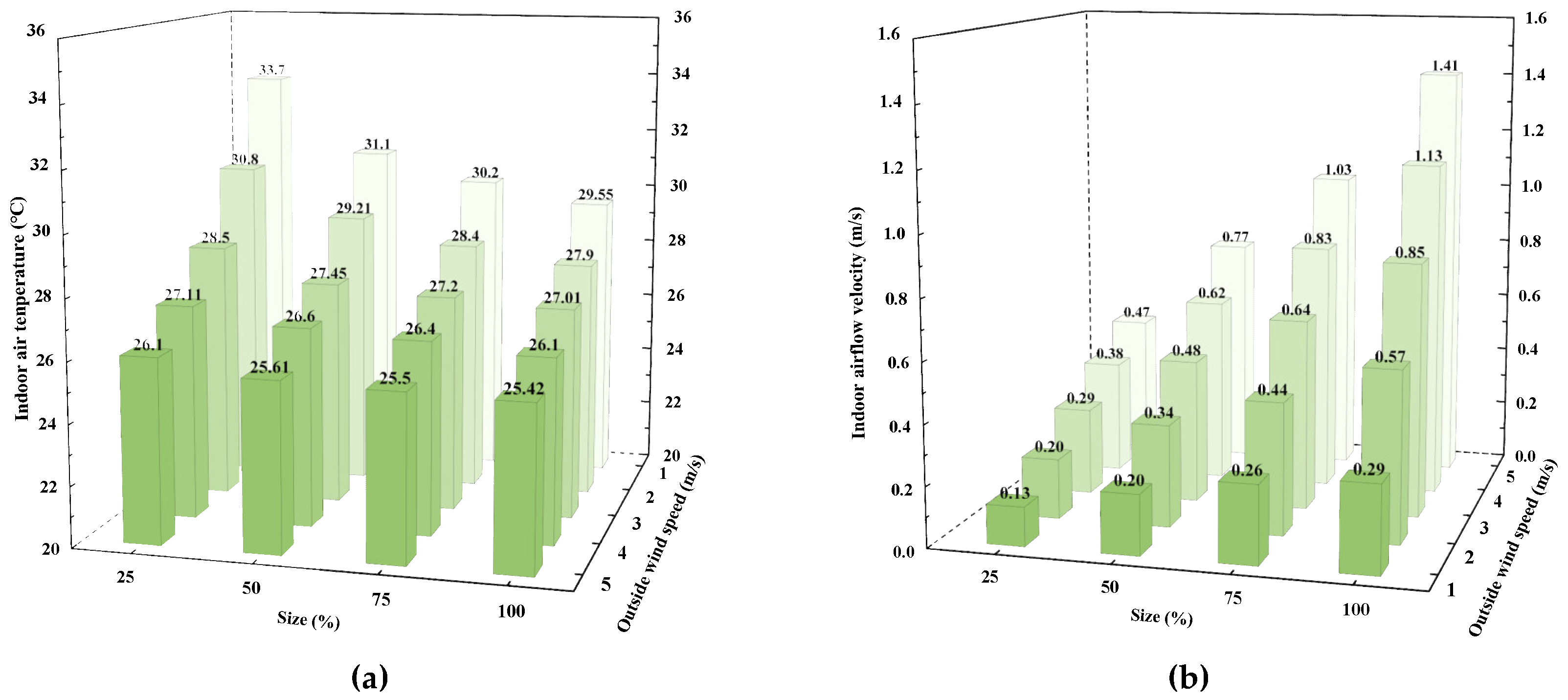
3.1. Analysis of Factors Affecting Ventilation Rate
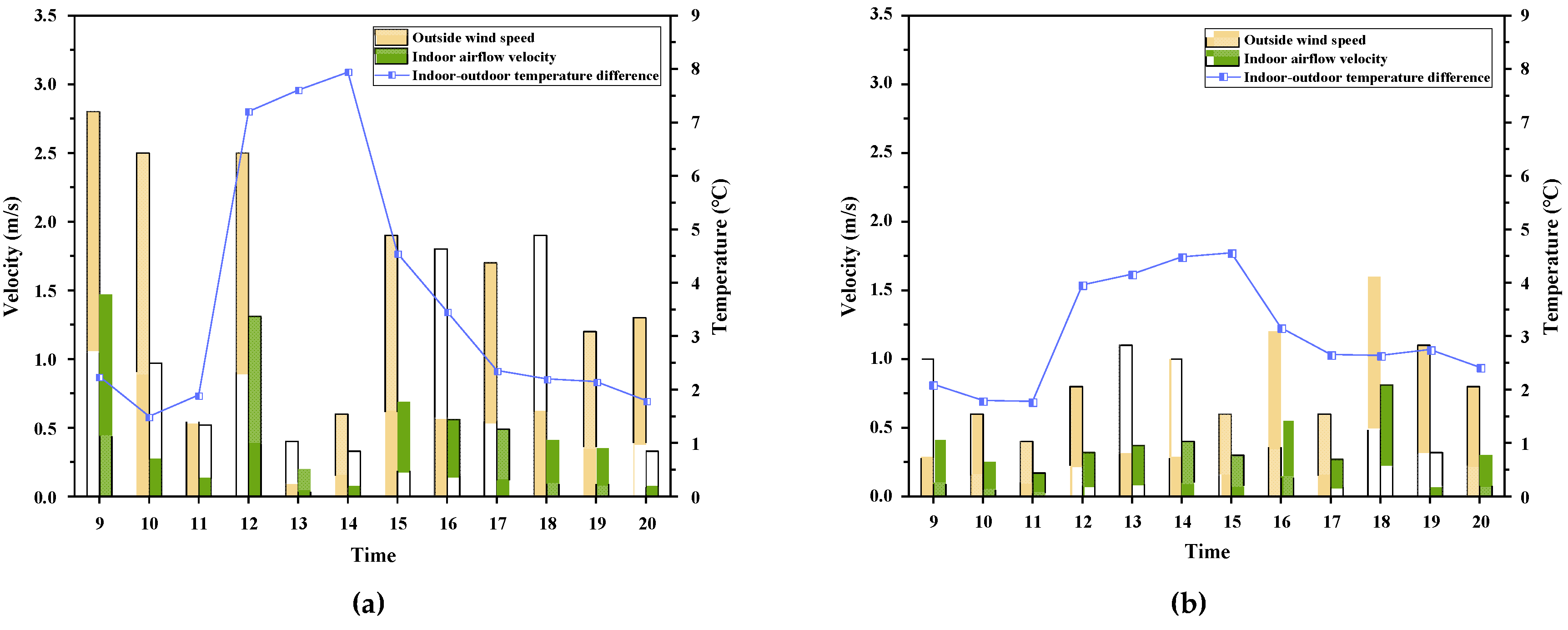
3.1.1. Daily Variation Patterns of Indoor Air Velocity
3.1.2. Correlation Analysis and Significance Testing
3.2. Ventilation Rate Model
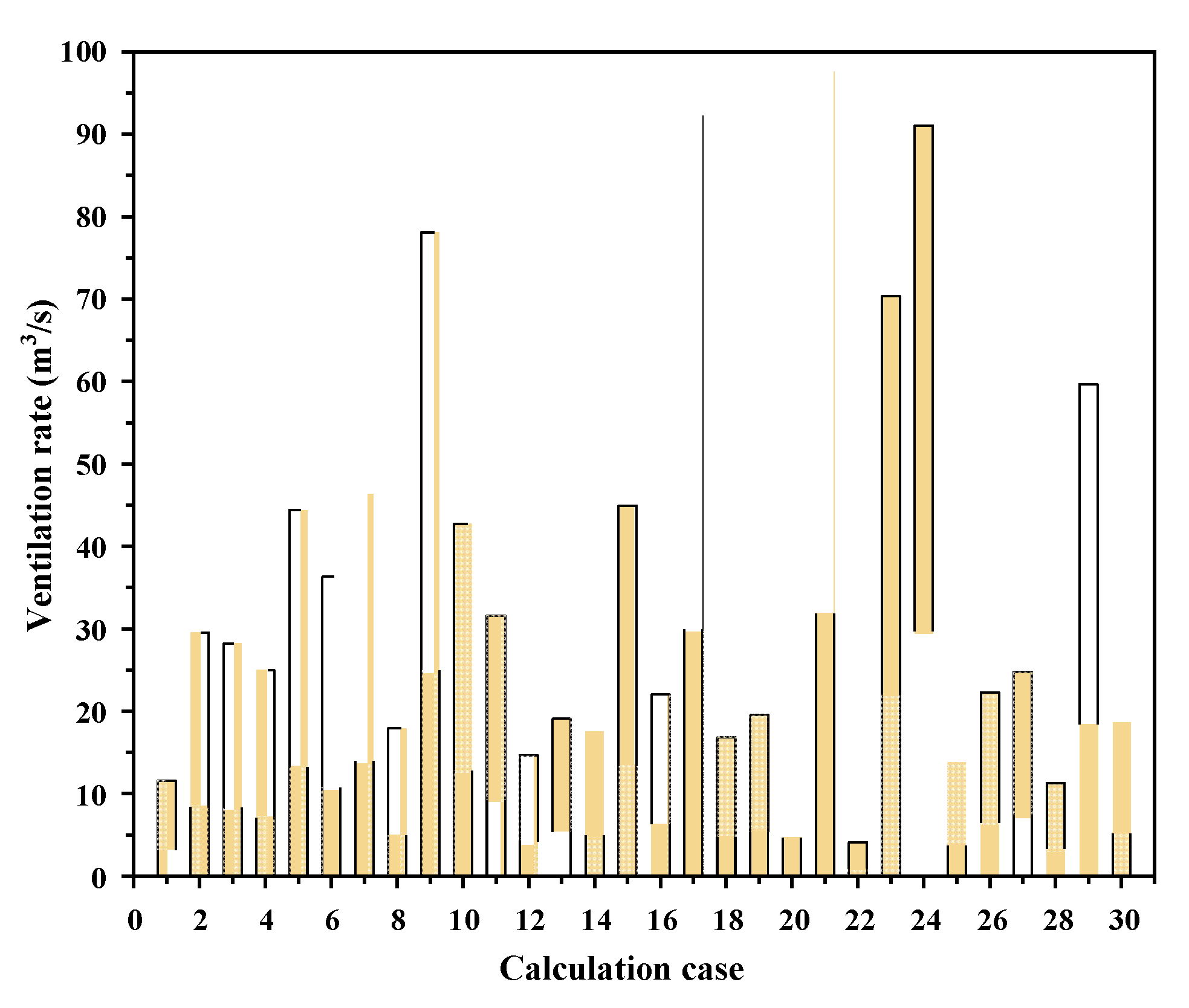
3.2.1. Ventilation Rate Sample Values
3.2.2. Ventilation Rate Model Establishment
3.3. Ventilation Rate Model Validation
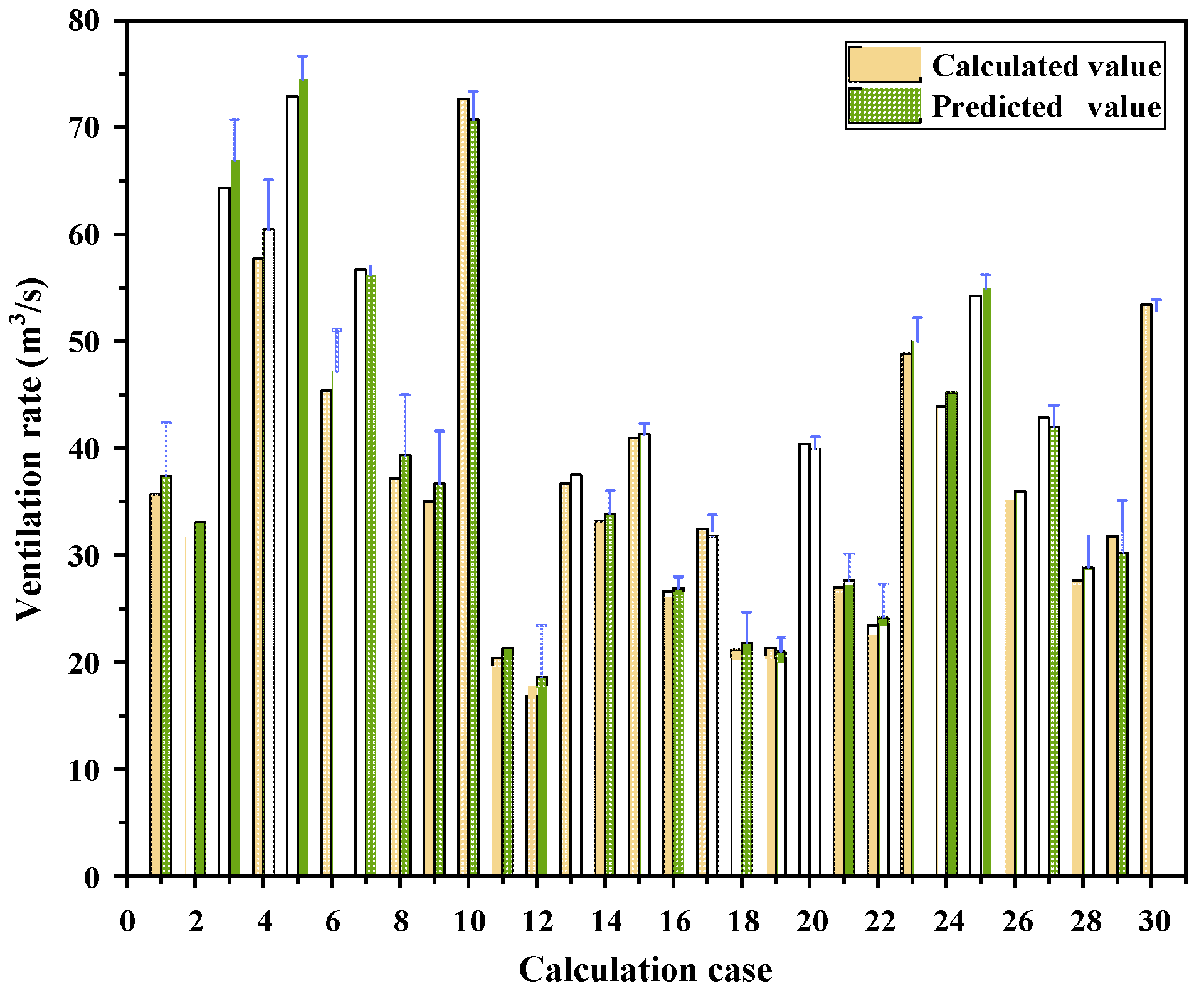
3.3.1. Accuracy Validation
3.3.2. Applicability Validation
3.4. The Impact of Ventilation Opening Sizes
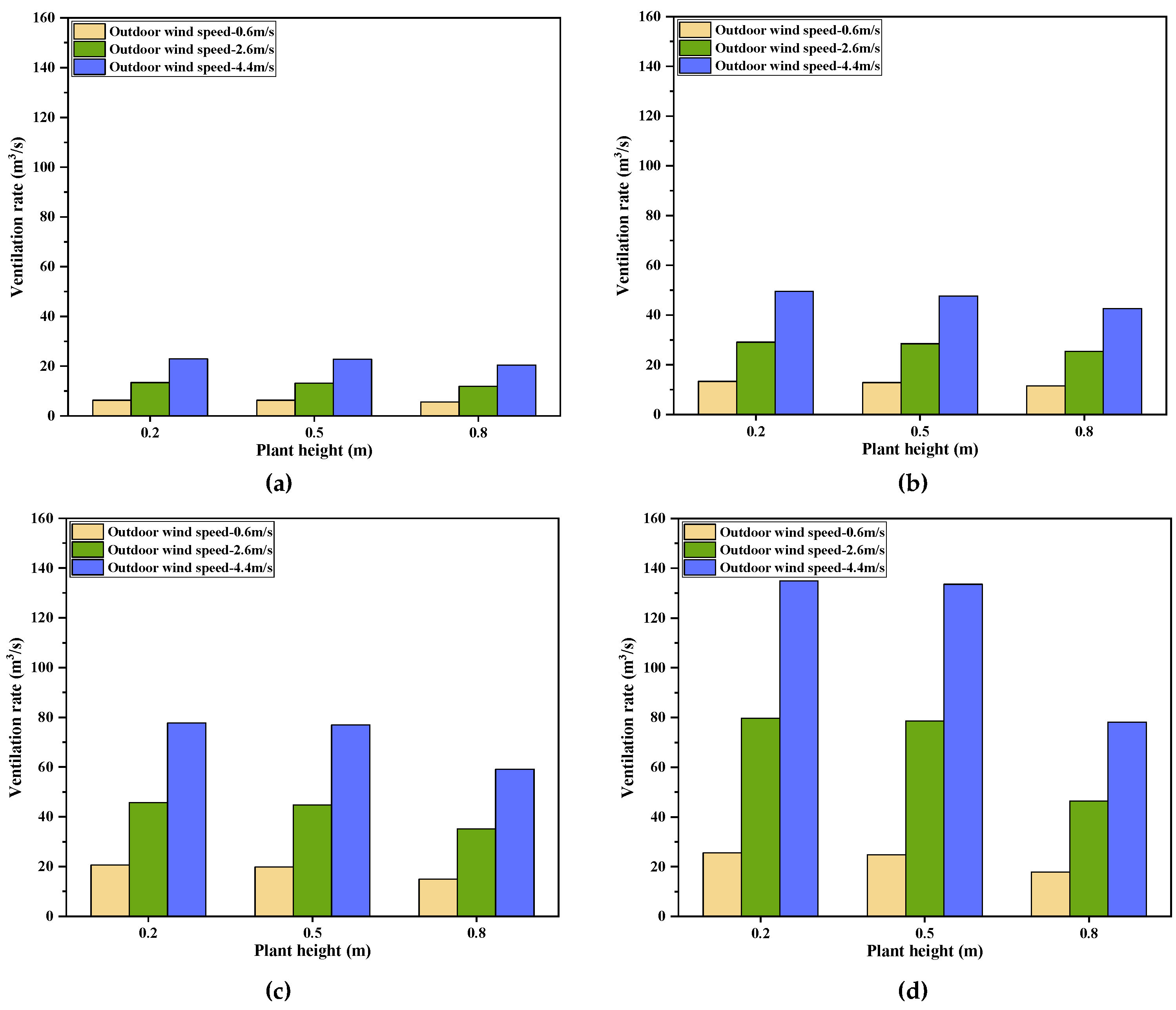
3.5. The Impact of Plant Heights
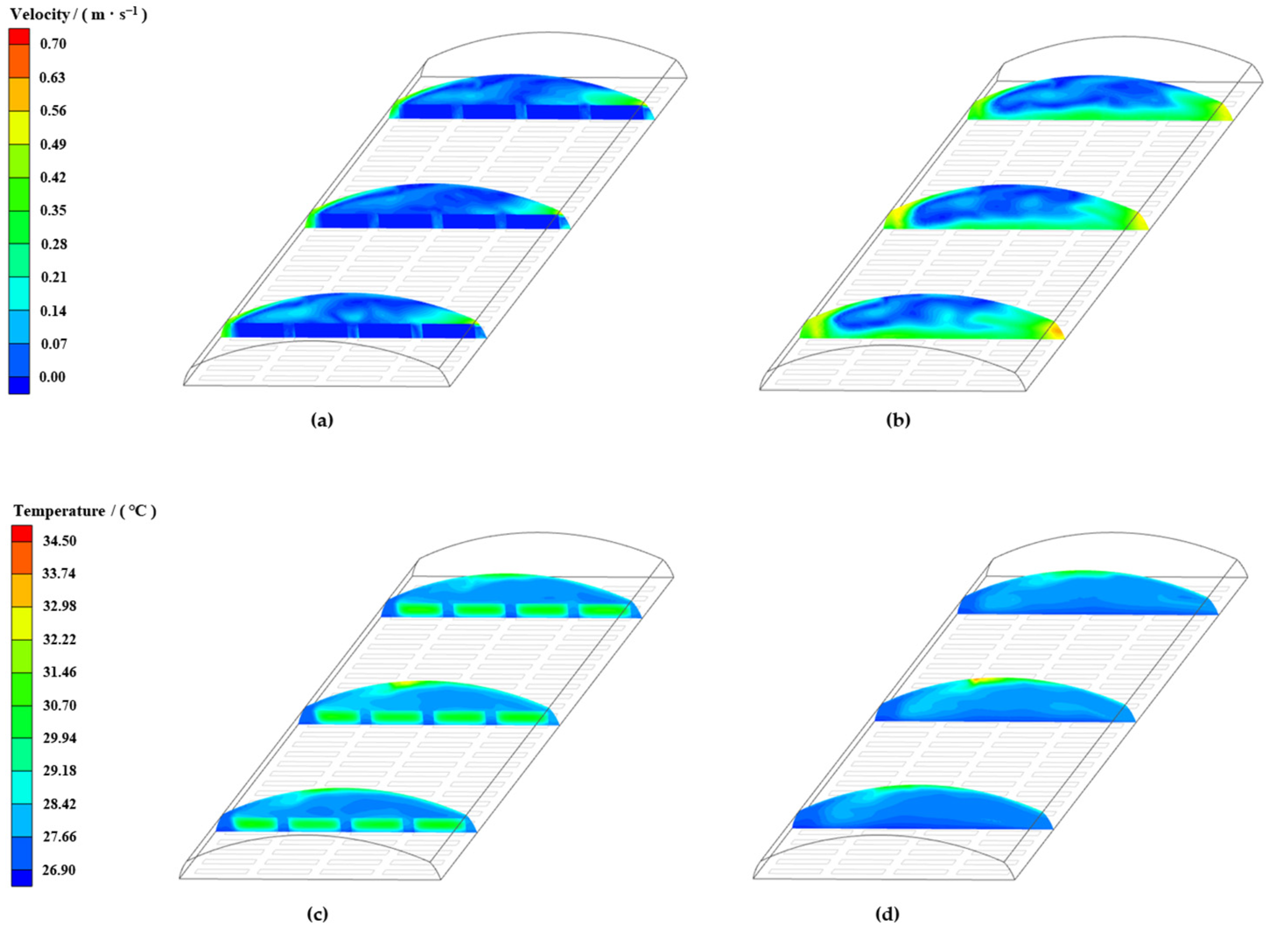
3.6. Analysis of Natural Ventilation Characteristics
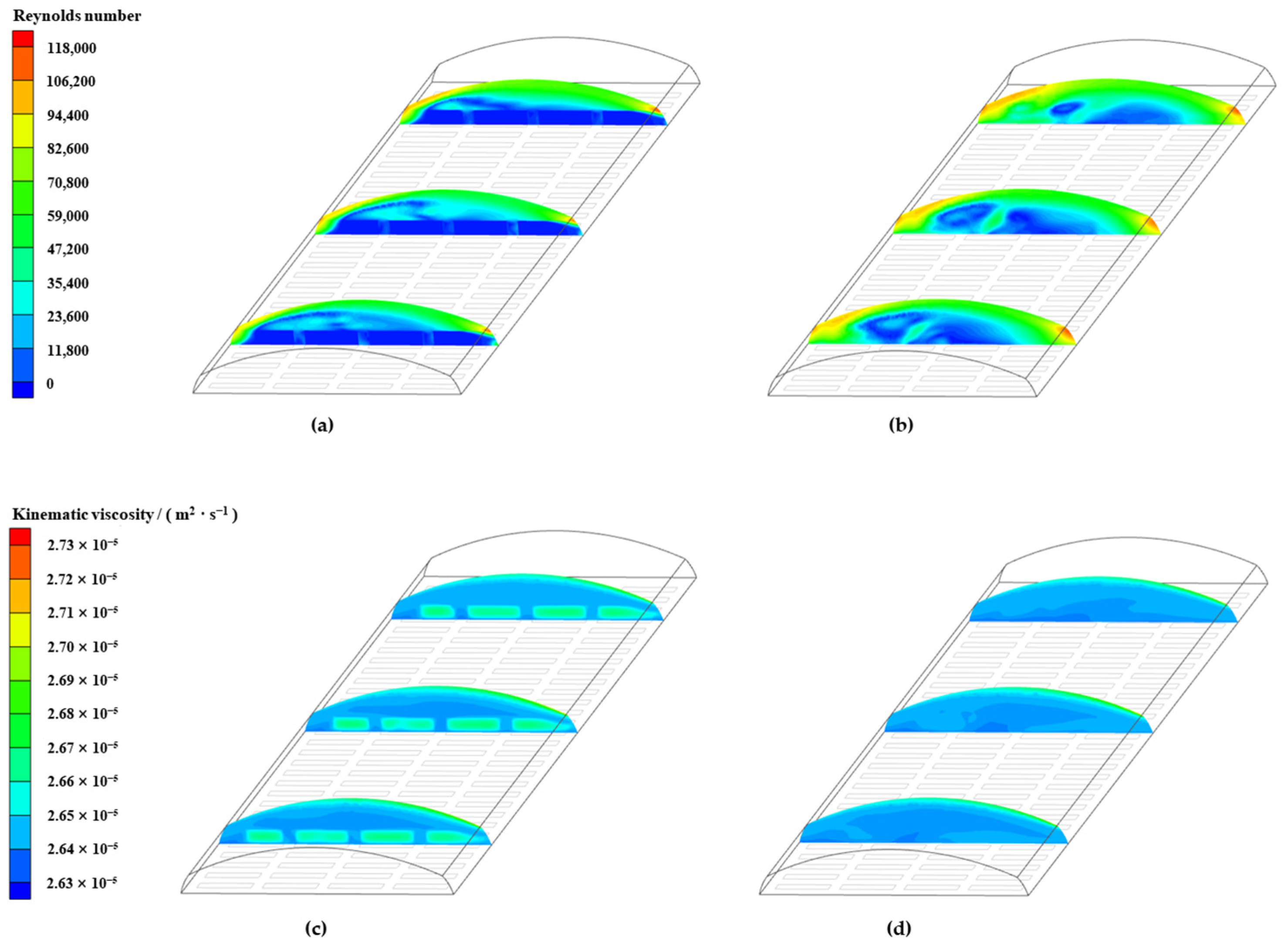
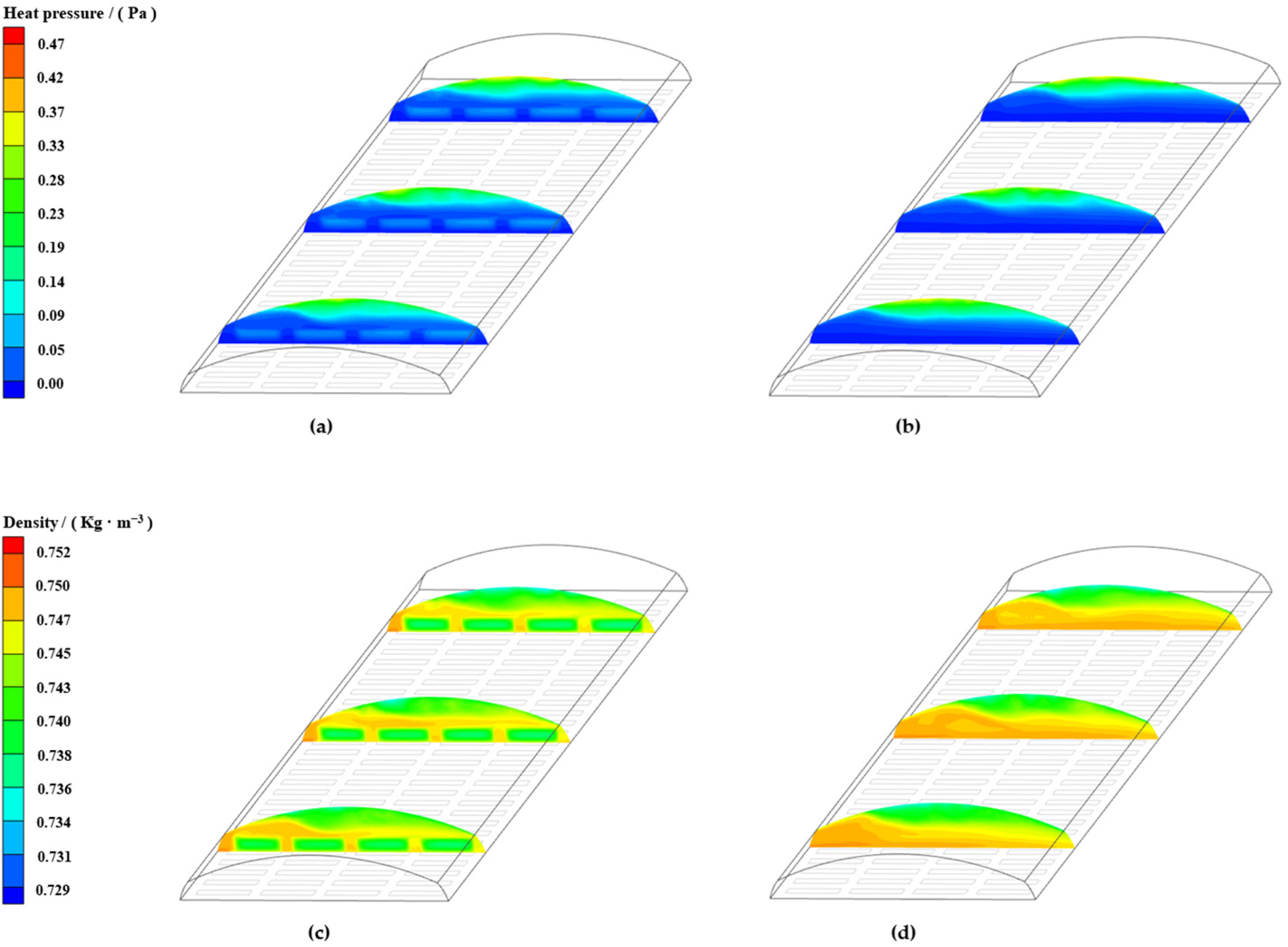
3.6.1. Wind Pressure Ventilation
3.6.2. Thermal Pressure Ventilation
3.6.3. Wind Pressure-Thermal Pressure Coupled Ventilation
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Li, Q. Discussion on the Development of Facility Vegetables in Tibet under the Rural Revitalization Strategy. Tibet Agric. Sci. Technol. 2020, 2, 62–65. (In Chinese) [Google Scholar]
- Cheng, F. Current Situation and Countermeasures of Facility Vegetable Development in Tibet Region. Anhui Agric. Bull. 2021, 27, 57–58. [Google Scholar] [CrossRef]
- Lei, W.; Lu, H.; Qi, X.; Tai, C.; Fan, X.; Zhang, L. Field measurement of environmental parameters in solar greenhouses and analysis of the application of passive ventilation. Sol. Energy 2023, 263, 111851. [Google Scholar] [CrossRef]
- Park, D.; Kang, B.; Cho, K.; Shin, C.; Cho, S.; Park, J.; Yang, W. A Study on Greenhouse Automatic Control System Based on Wireless Sensor Network. Wirel. Pers. Commun. 2011, 56, 117–130. [Google Scholar] [CrossRef]
- Cao, K.; Xu, H.; Zhang, R.; Xu, D.; Yan, L.; Sun, Y.; Xia, L.; Zhao, J.; Zou, Z.; Bao, E. Renewable and sustainable strategies for improving the thermal environment of Chinese solar greenhouses. Energy Build. 2019, 202, 109414. [Google Scholar] [CrossRef]
- Teitel, M.; Wenger, E. Air exchange and ventilation efficiencies of a monospan greenhouse with one inflow and one outflow through longitudinal side openings. Biosyst. Eng. 2014, 119, 98–107. [Google Scholar] [CrossRef]
- Tusi, A.; Shimazu, T.; Ochiai, M.; Suzuki, K. Continuous Measurement of Greenhouse Ventilation Rate in Summer and Autumn via Heat and Water Vapor Balance Methods. Environ. Control Biol. 2021, 59, 41–48. [Google Scholar] [CrossRef]
- Mashonjowa, E.; Ronsse, F.; Milford, J.R.; Lemeur, R.; Pieters, J.G. Measurement and simulation of the ventilation rates in a naturally ventilated Azrom-type greenhouse in Zimbabwe. Appl. Eng. Agric. 2010, 26, 475–488. [Google Scholar] [CrossRef]
- Akrami, M.; Javadi, A.A.; Hassanein, M.J.; Farmani, R. Study of the effects of vent configuration on mono-span greenhouse ventilation using computational fluid dynamics. Sustainability 2020, 12, 986. [Google Scholar] [CrossRef]
- Chu, C.R.; Lan, T.W.; Tasi, R.K.; Wu, T.R.; Yang, C.K. Wind-driven natural ventilation of greenhouses with vegetation. Biosyst. Eng. 2017, 164, 221–234. [Google Scholar] [CrossRef]
- Yang, Z.; Zou, Z.; Chen, S.; Wang, J. Indoor Wind Speed Distribution in Northwestern Solar Greenhouses and its Relationship with Outdoor Wind Speed and Ventilation Area. J. Northwest AF Univ. 2006, 34, 36–40. [Google Scholar] [CrossRef]
- Liang, B.; Zhao, S.; Li, Y.; Wang, P.; Liu, Z.; Zhang, J.; Ding, T. Study on the Natural Ventilation Characteristics of a Solar Greenhouse in a High-Altitude Area. Agronomy 2022, 12, 2387. [Google Scholar] [CrossRef]
- Fuller, R.; Aye, L.; Zahnd, A.; Thakuri, S. Thermal evaluation of a greenhouse in a remote high-altitude area of Nepal. Int. Energy 2009, 10, 71–80. [Google Scholar]
- Sun, X.; Wu, L.; Ma, Y.; Wang, R.; Song, B. Analysis of Temperature Variation in High-altitude Solar Greenhouses. Agric. Sci. 2023, 13, 257–263. [Google Scholar] [CrossRef]
- Cemek, B.; Atis, A.; Küçüktopcu, E. Evaluation of temperature distribution in different greenhouse models using computational fluid dynamics (CFD). Anadolu J. Agric. Sci. 2017, 32, 54–63. [Google Scholar] [CrossRef]
- Xu, F.; Cai, Y.; Chen, J.; Zhang, L. Temperature/flow field simulation and parameter optimal design for greenhouses with fan-pad evaporative cooling system. Trans. Chin. Soc. Agric. Eng. 2015, 31, 201–208. [Google Scholar] [CrossRef]
- Wu, F.; Xu, F.; Zhang, L.; Ma, X. Numerical simulation on thermal environment of heated glass greenhouse based on porous medium. Trans. Chin. Soc. Agric. Eng. 2011, 42, 180–185. [Google Scholar]
- Zhong, H.; Sun, Y.; Shang, J.; Qian, F.; Zhao, F.; Kikumoto, H.; Jimenez-Bescos, C.; Liu, X. Single-sided natural ventilation in buildings: A critical literature review. Build. Environ. 2022, 212, 108797. [Google Scholar] [CrossRef]
- Velazquez, J.; Montero, J.; Montero, J.; Baeza, E.; Lopez, J. Mechanical and natural ventilation systems in a greenhouse designed using computational fluid dynamics. Int. J. Agric. Biol. Eng. 2014, 7, 1–16. [Google Scholar] [CrossRef]
- Cemek, B.; Atis, A.; Küçüktopcu, E. Evaluation of various turbulence models performance for greenhouse simulation. In Proceedings of the Second International Symposium on Agricultural Engineering, Belgrade, Serbia, 9–10 October 2015; pp. 39–51. Available online: https://www.researchgate.net/publication/330114969 (accessed on 9 October 2015).
- Küçüktopcu, E.; Cemek, B. Evaluating the influence of turbulence models used in computational fluid dynamics for the prediction of airflows inside poultry houses. Biosyst. Eng. 2019, 183, 1–12. [Google Scholar] [CrossRef]
- Saberian, A.; Sajadiye, S. The effect of dynamic solar heat load on the greenhouse microclimate using CFD simulation. Renew. Energy 2019, 138, 722–737. [Google Scholar] [CrossRef]
- Nurmalisa, M.; Tokairin, T.; Kumazaki, T.; Takayama, K.; Inoue, T. CO2 Distribution under CO2 Enrichment Using Computational Fluid Dynamics Considering Photosynthesis in a Tomato Greenhouse. Appl. Sci. 2022, 12, 7756. [Google Scholar] [CrossRef]
- Kibwika, A.; Seo, H.; Seo, I. CFD Model Verification and Aerodynamic Analysis in Large-Scaled Venlo Greenhouse for Tomato Cultivation. AgriEngineering 2023, 5, 1395–1414. [Google Scholar] [CrossRef]
- Kang, L.; Zhang, Y.; Kacira, M.; Hooff, T. CFD simulation of air distributions in a small multi-layer vertical farm: Impact of computational and physical parameters. Biosyst. Eng. 2024, 243, 148–174. [Google Scholar] [CrossRef]
- Jiang, C.; Xu, K.; Rao, J.; Liu, J.; Li, Y.; Song, Y.; Li, M.; Zheng, Y.; Lu, W. Establishment and Solution of a Finite Element Gas Exchange Model in Greenhouse-Grown Tomatoes for Two-Dimensional Porous Media with Light Quantity and Light Direction. Agriculture 2024, 14, 1209. [Google Scholar] [CrossRef]
- Miao, Z. Numerical Simulation and Optimization of Vent Configuration for Natural Ventilation of the Large Space Exhibition Greenhouse. Refrig. Air Cond. 2020, 34, 29–38. [Google Scholar] [CrossRef]
- Li, H.; Li, Y.; Yue, X.; Liu, X.; Tian, S.; Li, T. Evaluation of airflow pattern and thermal behavior of the arched greenhouses with designed roof ventilation scenarios using CFD simulation. PLoS ONE 2020, 15, e0239851. [Google Scholar] [CrossRef]
- Lee, S.; Lee, I.; Kim, R. Evaluation of wind-driven natural ventilation of single-span greenhouses built on reclaimed coastal land. Biosyst. Eng. 2018, 171, 120–142. [Google Scholar] [CrossRef]
- Zhai, Z.; Mankibi, M.; Zoubir, A. Review of Natural Ventilation Models. Energy Procedia 2015, 78, 2700–2705. [Google Scholar] [CrossRef]
- Teitel, M.; Liran, O.; Tanny, J.; Barak, M. Wind driven ventilation of a mono-span greenhouse with a rose crop and continuous screened side vents and its effect on flow patterns and microclimate. Biosyst. Eng. 2008, 101, 111–122. [Google Scholar] [CrossRef]
- Boulard, T.; Baille, B. Modelling of air exchange rate in a greenhouse equipped with continuous roof vents. J. Agric. Eng. Res. 1995, 61, 37–48. [Google Scholar] [CrossRef]
- Sun, W.; Wei, X.; Zhou, B.; Lu, C.; Guo, W. Greenhouse heating by energy transfer between greenhouses: System design and implementation. Appl. Energy 2022, 325, 119815. [Google Scholar] [CrossRef]
- Li, H.; Lu, J.; He, X.; Zong, C.; Song, W.; Zhao, S. Effect of installation factors on the environment uniformity of multifunctional fan-coil unit system in Chinese solar greenhouse. Case Stud. Therm. Eng. 2024, 60, 104818. [Google Scholar] [CrossRef]
- Singh, M.; Sharma, K.; Prasad, V. Impact of ventilation rate and its associated characteristics on greenhouse microclimate and energy use. Arab. J. Geosci. 2022, 15, 288. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Instrument | Measurement Data | Measurement Range | Precision |
|---|---|---|---|
| PT100 | Temperature | −40~80 °C | ±0.2 °C (25 °C) |
| Humidity sensor | Humidity | 0~99%RH | ±3%RH (5%RH~95%RH, 25 °C) |
| Thermal bulb wind speed sensor | Velocity | 0~10 m/s | ±(0.03 m/s + 2%reading) |
| Light sensor | Total indoor radiation | 0~2000 W/m2 | ±10 W/m2 |
| Material | Density (kg·m−3) | Specific Heat Capacity (J·kg−1·K−1) | Thermal Conductivity (W·m−1·K−1) |
|---|---|---|---|
| Air | 0.77 | 1005.57 | 0.026 |
| Soil | 1700 | 1010 | 0.80 |
| Crop | 560 | 2100 | 0.19 |
| Plastic Film | 950 | 1600 | 0.29 |
| Cq | Cq (Cw)0.5 | MAE (m3·s−1) | ARE (%) | RMSE (m3·s−1) | R2 |
|---|---|---|---|---|---|
| 0.0955 | 0.6258 | 0.5257 | 3.27 | 0.6501 | 0.9996 |
| Outside Wind Speed (m·s−1) | Outside Air Temperature (°C) | Outside Soil Temperature (°C) | Inside Air Temperature (°C) | Inside Soil Temperature (°C) | Plastic Film Temperature (°C) |
|---|---|---|---|---|---|
| 1.6 | 19.4 | 23.6 | 21.5 | 19.3 | 30.0 |
| 0.2 | 26.4 | 24.9 | 32.5 | 28.0 | 39.1 |
| 0.7 | 27.5 | 26.3 | 29.4 | 26.9 | 32.9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Zhao, S.; Dai, A.; Zhang, J.; Fan, Z.; Ding, T. Study on the Natural Ventilation Model of a Single-Span Plastic Greenhouse in a High-Altitude Area. Agronomy 2024, 14, 2166. https://doi.org/10.3390/agronomy14092166
Li Y, Zhao S, Dai A, Zhang J, Fan Z, Ding T. Study on the Natural Ventilation Model of a Single-Span Plastic Greenhouse in a High-Altitude Area. Agronomy. 2024; 14(9):2166. https://doi.org/10.3390/agronomy14092166
Chicago/Turabian StyleLi, Youyu, Shumei Zhao, Anguo Dai, Jingfu Zhang, Zilong Fan, and Tao Ding. 2024. "Study on the Natural Ventilation Model of a Single-Span Plastic Greenhouse in a High-Altitude Area" Agronomy 14, no. 9: 2166. https://doi.org/10.3390/agronomy14092166