Abstract
Sweet corn (Zea mays var. rugosa Bonaf.) is a crop with a high economic benefit in tropical and subtropical regions. Heat tolerance analysis and heat-tolerant gene mining are of great significance for breeding heat-resistant varieties. By combining improved genotyping using targeted sequencing (GBTS) with liquid chip (LC) technology, a high-density marker array containing 40 K multiple single polynucleotide polymorphisms (mSNPs) was used to genotype 376 sweet corn inbred lines and their heat-stress tolerance was evaluated in the spring and summer of 2019. In general, plant height, ear height and the number of lateral branches at the first level of the male flowers were reduced by 24.0%, 36.3%, and 19.8%, respectively. High temperatures in the summer accelerated the growth process of the sweet corn, shortening the days to shedding pollen by an average of 21.6% compared to the spring. A genome-wide association study (GWAS) identified 85 significant SNPs distributed on 10 chromosomes. Phenotypes in the spring and summer were associated with the 21 and 15 loci, respectively, and significant phenotypic differences between the two seasons caused by the temperature change were associated with the 49 SNP loci. The seed setting rate (SSR) was more susceptible to heat stress. An annotation analysis identified six candidate genes, which are either heat shock transcription factors (Hsfs) or heat shock proteins (Hsps) in Arabidopsis and rice (Oryza sativa), and these candidate genes were directly and indirectly involved in the heat-resistant response in the sweet corn. The current findings provide genetic resources for improving the heat-stress tolerance of sweet corn by molecular breeding.
1. Introduction
Maize is one of the major staple crops in the world and ranks among the top three in global food consumption [1]. At present, climate change poses a serious threat to maize production, and at the same time, maize is susceptible to abiotic stresses such as drought, excessive moisture, and high temperature (heat) [2,3,4]. Compared to other environmental stresses, heat stress can significantly impact and reduce crop yield. It is estimated that extreme heat stress during flowering would double global maize yield losses in the 2080s in comparison with that in the 1980s [5]. Therefore, it is particularly important to develop maize varieties that are tolerant to heat stress and discover heat-resistant genes [6,7,8,9].
The heat tolerance of maize is a complex trait that involves multiple genes and biological processes [9]. Maize plants are particularly sensitive to heat stress during flowering as high temperatures can reduce the pollen germination rate [10], cause late pollen tube growth arrest and sterility [11], and extend the anthesis-silking interval [12]. At the same time, heat stress also lowers the photosynthesis rate [13], decreases the chlorophyll content, and causes damage to the leaf [14] and root growth [15], negatively affecting the growth, development and yield of the maize. Moreover, heat stress also results in a heat shock response. The eukaryotic heat shock response is regulated by conserved heat shock transcription factors (Hsfs), and a group of identified proteins called heat shock proteins (Hsps) will be produced under heat stress. At the onset of heat stress, Hsps maintain cellular protein homeostasis by preventing protein folding and aggregation. To identify key cellular structures and functions that regulate the stress response, Gagliardi et al. studied heat shock factors and heat shock protein 70 (Hsp70) during the development of maize pollen [16], and Lin et al. identified and analyzed a family of Hsfs in maize [17]. However, more Hsfs and Hsps need to be mined as the global climate conditions become increasingly complex.
Through marker–trait association, phenotypic information related to crop stresses can be associated with the genomic regions for heat-responsive genes or quantitative trait loci (QTL). Applying genome-wide association studies (GWASs) can not only provide high-precision QTL localization but can also reveal the complex genetic networks behind the traits. Hence, GWASs have now become an indispensable tool for predicting allele function, pinpointing mutations, and identifying candidate genes responsible for agronomic traits. A GWAS is typically used together with high-density and high-quality single nucleotide polymorphism (SNP) markers. Genotyping by target sequencing (GBTS) is a high-throughput genotyping technique characterized by its low cost and high flexibility [18]. GBTS technology shows a higher polymorphism of markers, with a uniform distribution and technical repeatability of up to 99.9%. Such high-quality data are crucial for accurate GWASs. A GWAS could effectively identify genotype–phenotype associations in natural populations, providing a high-resolution method for detecting the QTL [19] and candidate genes responsible for the relevant traits [20]. In 2008, the first GWAS in maize identified 8590 SNP sites correlated with grain oleic acid content using 553 superior inbred lines [21]. The “B73” reference maize genome [22] and relevant hapmaps could be used with a GWAS to elucidate the genetic basis of complex traits [23] and to mine for significant loci. Many yield-related loci, such as plant height, grain length, grain width, ear length, and grain weight, have been discovered [19,24,25,26,27], and the effects of the candidate genes at significant loci on the yield have been confirmed using overexpression and silencing experiments [25,28]. GWASs have been also used to identify SNPs related to abiotic stresses such as cold and salt [29,30,31,32,33], as well as those related to disease and insect resistances for maize [34,35,36,37,38,39]. However, we still face a lack of research using GWASs to analyze the heat tolerance of maize [40,41].
In this study, a GWAS was performed on the yield-related traits of sweet corn in the environments of spring and summer. The main purposes were to explore the effects of spring and summer sowing on the agronomic traits of sweet corn and to identify the genomic regions and candidate genes associated with heat-stress tolerance.
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
The materials of this study included 376 inbred lines, which were developed from the tropical temperate hybrids through second-cycle line breeding for subtropical area, by the Sweet Corn Genetic Breeding Team of Foshan University. The experiment was conducted at the sweet corn planting base (E113°06′, N23°02′) in the middle of Nanhai District, Foshan City, Guangdong Province from March to November 2019, with a subtropical monsoon climate. The local spring is warm with abundant rain and the summer is hot and rainy. The annual average temperature is 23.2 °C, and the frost-free period is 350 to 355 days. According to the meteorological and ecological environment monitoring system near the test base, the meteorological data during the experiment were collected as a reference for assessing the heat tolerance performance of sweet corn under high temperature (Figure S1). The lowest temperature of the test base in the spring (March to July) was 12 °C, with an average temperature of 24 °C. In the summer (June to November), the lowest temperature was 24 °C and the highest was 40 °C, with an average temperature of 30 °C. The average maximum daily temperatures during the flowering periods for spring (15 days from 9 May to 24 May) and summer (18 days from 9 August to 22 August) seasons were 31.6 and 35.84 °C, respectively, indicating that the spring was suitable for the growth of sweet corn, while the summer had high enough temperatures to cause heat stress for the growth of sweet corn.
2.2. Phenotyping and Data Analysis
A total of eight traits were evaluated in the spring and summer fields and after harvest. These traits represented the indexes of heat tolerance for sweet corn. Three normal growing plants were selected from each inbred line to collect agronomic traits data, including plant height (PH), ear height (EH), internode number (IN), and the number of lateral branches at first level of the male flowers (MFLBL). These agronomic traits were evaluated according to NY/T 2232-2012 [42] Industry Standard for Maize Testing. The survival rate (SR), seed setting rate (SSR), and the successful ear rate of maize were investigated. Days to shedding pollen (DTS), when the anthers of the stamen split and released pollen, was recorded as follows: 60% of the plants were shedding pollen. Meanwhile, summer male fringing stage stress resistance index (FRI) was also assessed. The phenotypic expression of the traits measured in this study in a single environment were determined by taking the average from three replicates for each line. Differences in the same trait in the two crop seasons (spring and summer) were calculated. In this study, SPSS 20.0 software (IBM Corp., Armonk, NY, USA) was used to describe the basic information for each trait and to calculate Pearson correlation coefficient (r) among different traits. GraphPad Prism 8.0 was used to make the distribution map. QTL IciMapping 4.2 software was employed to calculate the heritability for the traits.
2.3. DNA Extraction and Genotyping
Leaf samples of 3 to 4-week-old seedlings from each line were taken, and the genomic DNA was extracted from the fresh leaves using cetyltrimethyl ammonium bromide (CTAB) method [43]. An improved genotyping by target sequencing (GBTS) system combined with capture-in-solution (liquid chip) technology, which was used to develop 40K mSNPs in a previous study [44], was used to genotype the 376 sweet corn lines. The raw sequencing data from these lines were processed using the fastp tool (v 0.20.0) [45] with the main parameter of -n 10-q 20-U 40 to trim the library adapter sequence and remove reads with a low quality (phred score <20), base ratio >40%, and reads with >10 N bases (unknown bases). The filtered reads aligned against the B73 reference genome were compared by the BWA–MEM algorithm using the BWA 0.7.17 package [46]. The comparison results were sorted and duplicated sequences were removed using the Picard tool (version 2.1.1) to optimize the data quality. Finally, GATK tool (version v3.5-0-g36282e4) was used to detect the variation of the processed data, and the parameter was set at “-dcov 1000000-minIndelFrac 0.15-glm BOTH -l INFO” to identify SNPs and capture comprehensive variation information at the genome level. For GWAS, a set of 40 K SNP markers was used to genotype 376 samples and then we further removed markers with poor hybridization quality or unreliable genotyping, and manually adjusted markers with uncertain genotyping.
2.4. GWAS and the Prediction of Candidate Genes
The selected set of SNP markers with high quality and polymorphisms were used to conduct GWAS for eight target traits evaluated for 376 inbred lines using trait values collected for spring, summer, and their difference. The association analysis was performed using MLM linear model in TASSEL5.0 software. The threshold for significance was set at 0.1/n [47], where n represented the number of SNPs. The ggplot2 3.4.2 package of R 4.0.0 was employed to plot the physical locations and add the p-values of these high-quality variation loci on the maize genome into Manhattan map and QQ map [48].
The genes identified within the flanking 300 kb of these SNPs at each GWAS locus were considered as candidate genes for relevant heat tolerance traits [49]. EGCNOG-MAPPER website (http://eggnog-mapper.embl.de/, accessed on 9 August 2024) e.g., accessed on 15 March 2024. was used for GO annotation and KEGG enrichment analysis. The gene function annotation information was collected from Phytozome v13. In addition, the homologous genes in rice and Arabidopsis thaliana corresponding to the candidate genes were identified using BlastP comparison, as well as the gene function annotation information. According to the literature studies, the genes related to heat tolerance and heat stress were selected as candidate genes.
3. Results
3.1. Phenotypic Variation of Target Traits
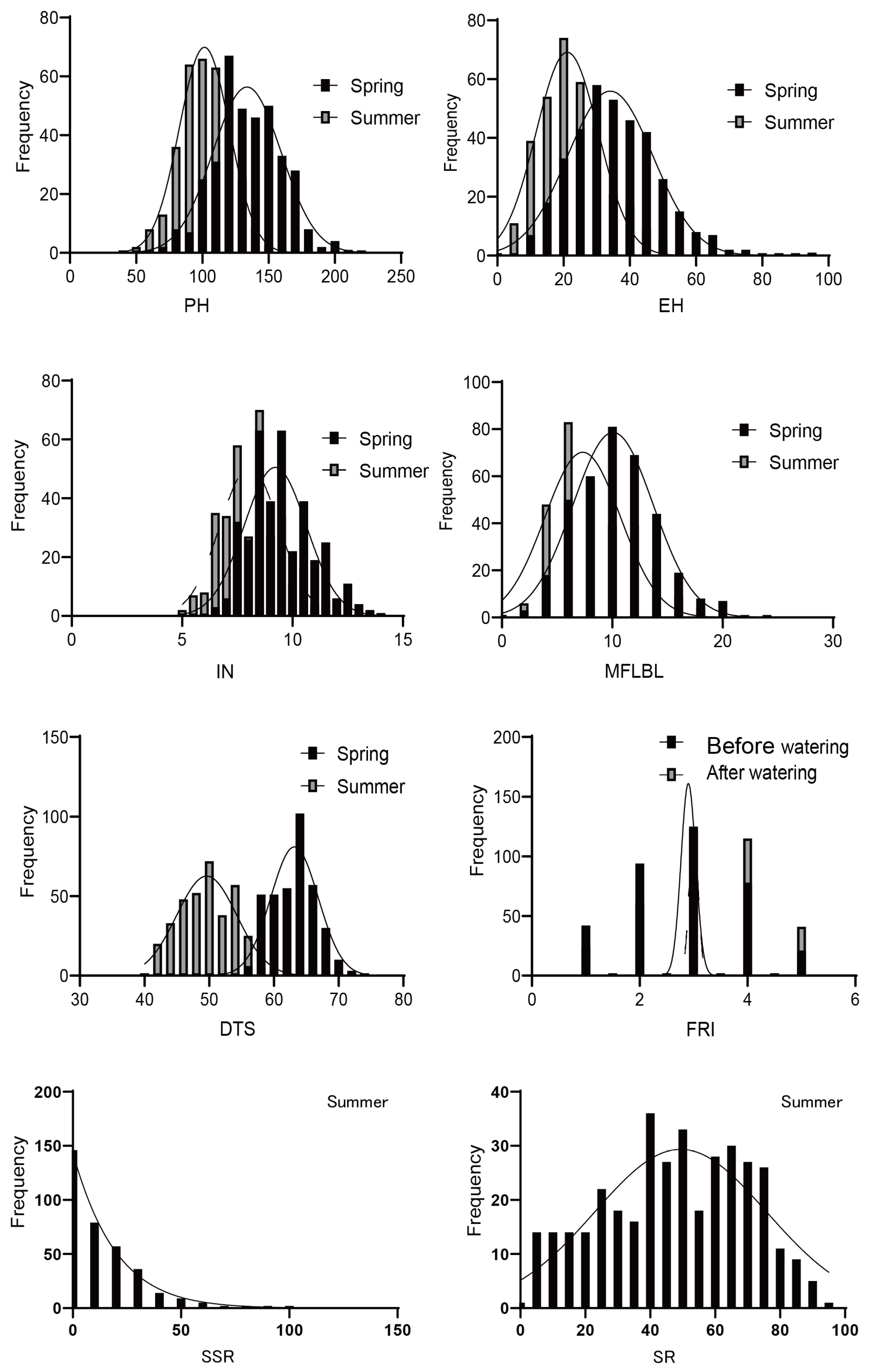
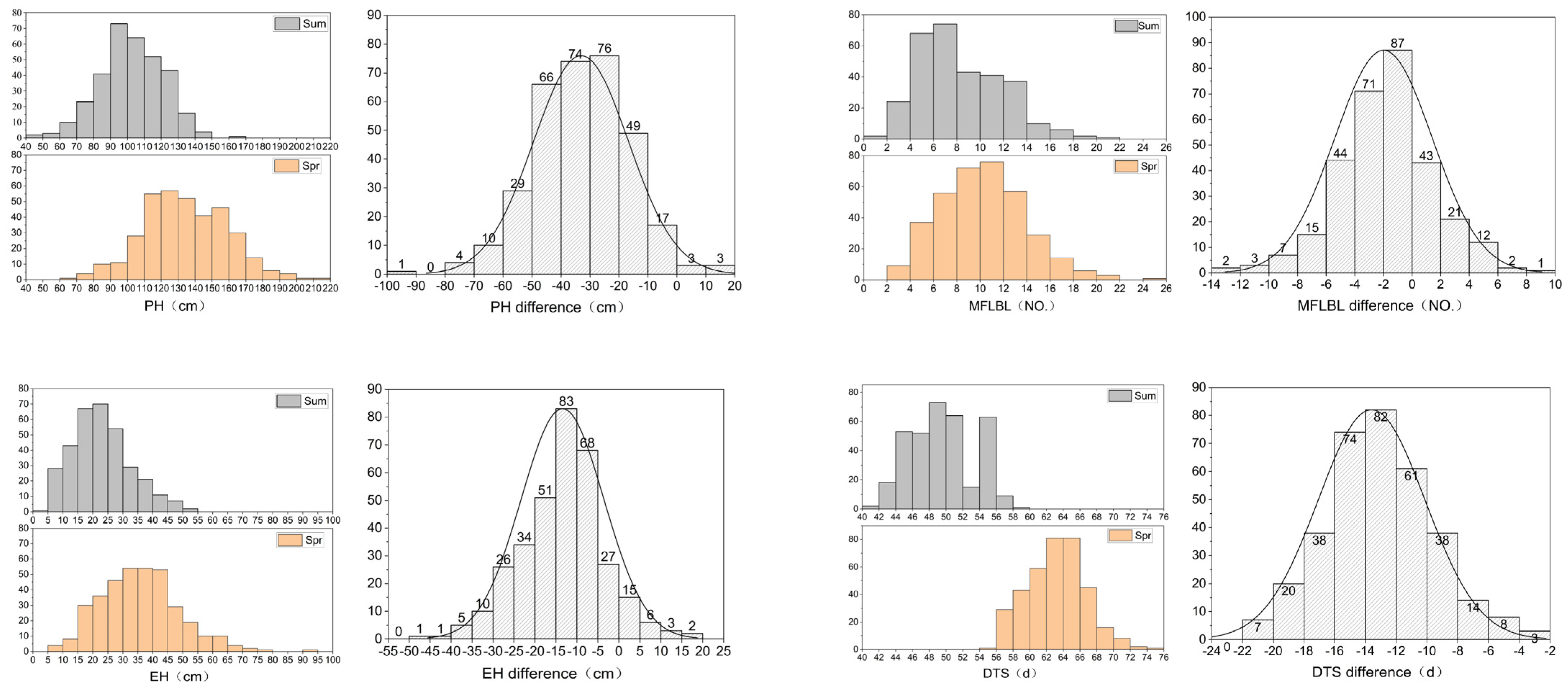
Sweet corn inbred lines and their traits responded differently to high-temperature stress. Each trait showed a wealth of phenotypic variations, all of which were normally distributed (Figure 1, Table 1). The skewness of each trait in different environments was less than 1, indicating that the distribution of the phenotype was in line with a normal distribution. With the exception of IN in the spring, the heritability of the other traits was higher than 90% in both environments. Among them, the coefficient of variation of DTS was the smallest, and was lower than 10 in both seasons, while that of SSR was the highest at 119.29. The coefficient of variation of the PH, EH, IN, MFLBL, and FRI differences were higher than their performances in the two environments, but the coefficient of variation of the SSR difference was smaller than its value in the summer. The responses of ear height (EH), internode number (IN), number of lateral branches at the first-level of the male flowers (MFLBL), days to shedding pollen (50% of the plants shedding pollen) (DTS), and summer male fringing stage stress resistance index (FRI) to high-temperature stress were more obvious, highlighting the differences in heat tolerance among inbred lines. Among eight target traits, plant height (PH), EH, MFLBL, and DTS showed significant differences in their frequency distribution in the spring and summer (Figure 2). In general, the PH and EH under high-temperature stress (in the summer) decreased by 24.0% and 36.3% when compared to those the spring, respectively; both the IN and MFLBL decreased by 14.0% and 19.8% when compared to those in the spring, respectively, and the DTS shortened 21.6% on average compared with the spring. This indicates that high-temperature stress had a great effect on the development of sweet corn. The average survival rate of the 376 inbred lines in the summer was lower than 50%, and the average SSR was 12.7%. These results showed that the studied traits were largely affected by high temperatures.
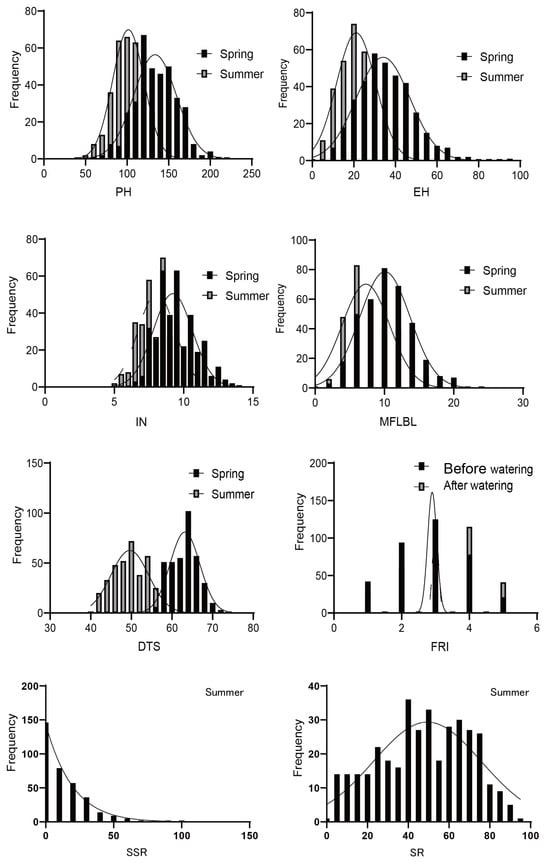
Figure 1.
Distribution of eight traits relevant to heat-stress tolerance. Plant height (PH), ear height (EH), internode number (IN), number of lateral branches at first level of male flowers (MFLBL), survival rate (SR), seed setting rate (SSR), days to shedding pollen (DTS), and summer male fringing stage stress resistance index (FRI).

Table 1.
Basic information on eight traits for sweet corn inbreds planted in spring and summer.
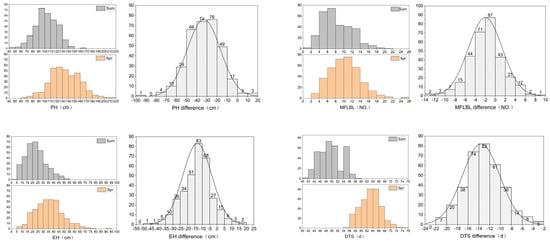
Figure 2.
Differences in four traits of sweet corn inbred lines between spring and summer. Plant height (PH), ear height (EH), internode number (IN), number of lateral branches at first level of male flowers (MFLBL), survival rate (SR), seed setting rate (SSR), days to shedding pollen (DTS), and summer male fringing stage stress resistance index (FRI).
Before watering means watering before high temperatures at the male flowering stage, while after watering means watering after high temperatures at the male flowering stage. The plant height (PH), ear height (EH), internode number (IN), number of lateral branches at the first level of the male flowers (MFLBL), survival rate (SR), seed setting rate (SSR), days to shedding pollen (DTS), and summer male fringing stage stress resistance index (FRI) were determined. H2 stands for heritability.
3.2. SNP Quality Control
Firstly, 40K SNP markers were compared with the reference genome of maize (B73RefGen_v3 version) to count the number of SNPs within each window. On this basis, after the elimination and genotype identification, a total of 36,331 high-quality SNP markers with high polymorphisms were finally selected for subsequent analyses.
3.3. GWAS on the Eight Traits
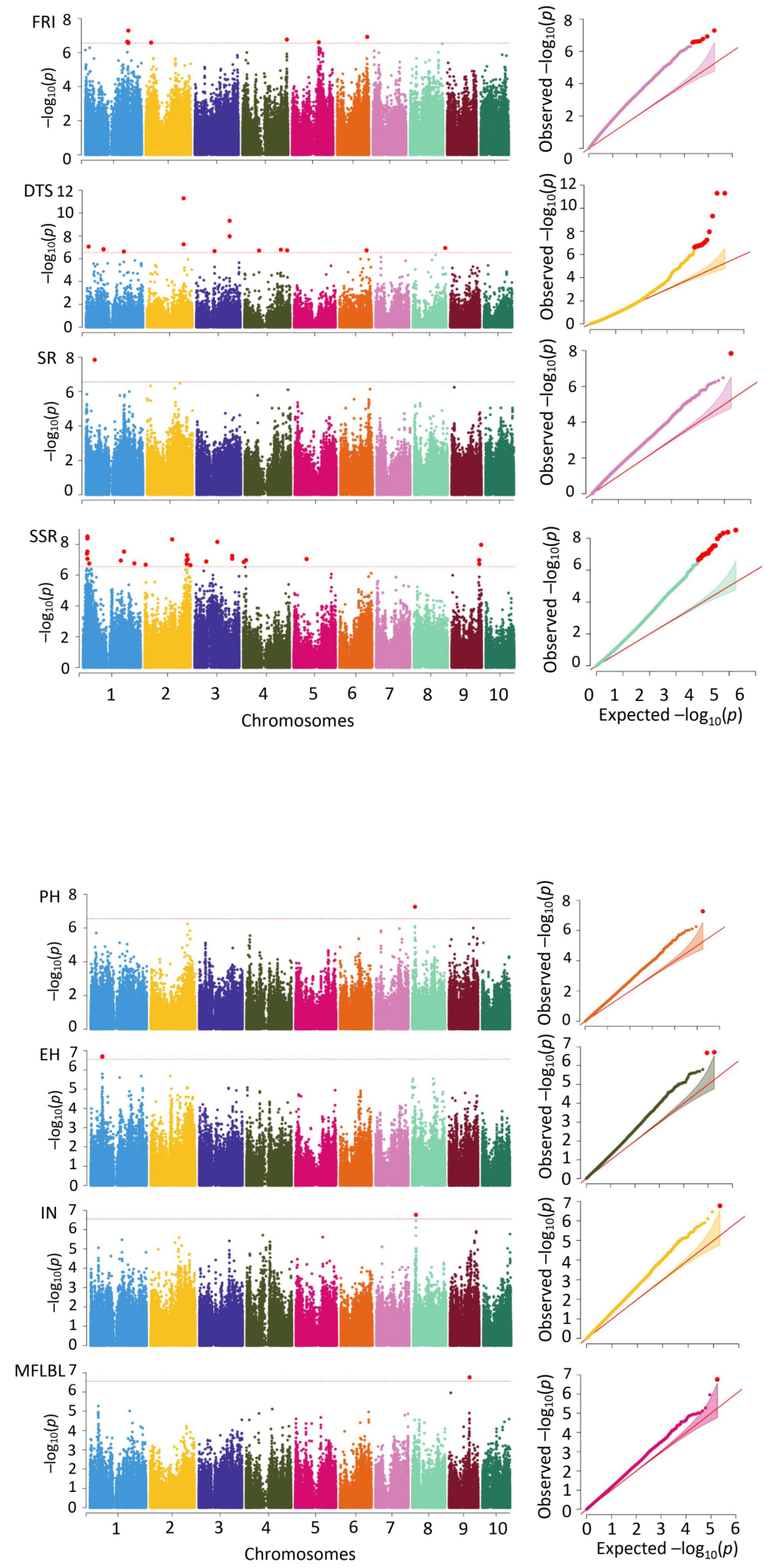
Applying the MLM method, the 36,331 SNP markers together with the PH, EH, IN, MFLBL, DTS, SR, SSR, and FRI were subjected to a correlation analysis, and 85 significant SNPs distributed across 10 chromosomes were identified. The spring data were associated with 21 significant SNPs, and the summer data were related to 15 significant SNPs. The differences between the data of the two environments were associated with 49 significant SNP sites, showing phenotypic variation-explained (PVE) rates from 5.1% to 15.3%, with an average of 8.8%.
The DTS was associated with 32 loci (Figure 3, Table 2), with 3 each on chromosomes 2, 6, and 10, 4 each on chromosomes 3 and 9, and 5 on chromosomes 1, 4, and 8. The PVE rate of these loci ranged from 5.1% to 15.3%, with an average of 8.3%. Fourteen loci were associated with the spring and four with the summer. The differences between the two environments were associated with 14 sites. The EH was associated with three loci, which were all associated with chromosome 1, and the PVE rate of each locus was 9.2%, 9.3%, and 9.4%. One locus was associated with the summer, and there were two loci associated with the seasonal differences. The IN was associated with two significant SNP loci and one of them, on chromosome 5, was associated with the spring, with a PVE rate of 5.6%. The seasonal difference was associated with one locus on chromosome 8, with a PVE rate of 9.5%. The MFLBL was associated with eight significant loci, two each on chromosomes 4 and 9, three on chromosome 3 and one on chromosome 8, with PVE rates ranging from 7.6% to 10.5%, with an average of 8.6%. The MFLBL was associated with eight significant loci, two each on chromosomes 4 and 9, three on chromosome 3 and one on chromosome 8. The PVE rates of these sites ranged from 7.6% to 10.5%, with an average of 8.6%. Two loci were associated with the spring, five with the summer, and one site with the difference. The PH was associated with six SNP loci, with two each on chromosomes 2 and 9 and one each on chromosomes 5 and 8, showing PVE rates from 6.5% to 9.9% and an average rate of 7.9%. Four loci were associated with the spring and one with the summer. The difference between the two environments was associated with one locus. The SR was only associated with a single locus on chromosome 1, with a PVE rate of 9.6%. The SSR was associated with 26 loci, with one locus on chromosome 5, two on chromosome 4, three on chromosome 9, four on chromosome 3, seven on chromosome 2, and nine on chromosome 1. All of these 26 loci were also identified using the difference between the two environments, with PVE rates from 9.1% to 12.0% and an average rate of 10.4%.
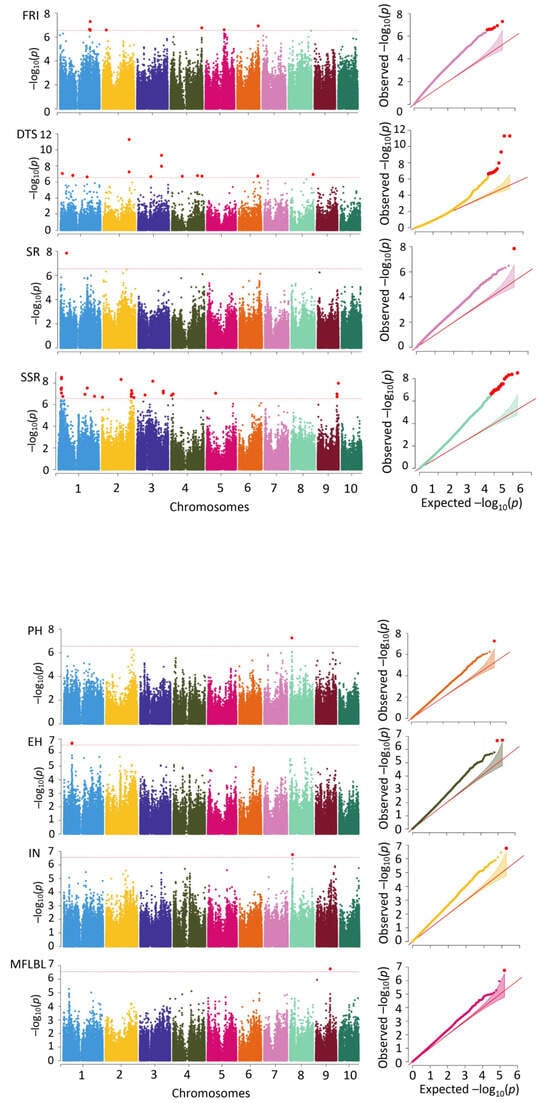
Figure 3.
Results of correlation analysis between molecular markers and fiber quality traits based on MLM method. Note: on the left is the Manhattan map and on the right is the QQ map. Plant height (PH), ear height (EH), internode number (IN), number of lateral branches at first level of male flowers (MFLBL), survival rate (SR), seed setting rate (SSR), days to shedding pollen (DTS), and summer male fringing stage stress resistance index (FRI).
Table 2.
Genome-wide association study on eight traits using three sets of data (datasets collected in spring and summer and their differences).
3.4. Candidate Genes Associated with Heat-Stress Tolerance
A total of 629 candidate genes were obtained according to the locations of the significant SNP loci on the reference genome and then enriched by the Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) analyses. The GO analysis results showed that these genes were significantly enriched to two GO terms (p-adjusted value ≤0.01). Specifically, the annotated genes were mainly located on the vacuole in the cell component term and were enriched for programmed cell death and cellular senescence during cell development in the biological process term. The KEGG analysis did not enrich any result (p-adjusted ≤0.05).
These candidate genes were compared to the homologous genes of Arabidopsis and rice. Based on their annotation information, six candidate genes were obtained (Table 3), and they were directly or indirectly involved in the heat shock response. Specifically, GRMZM2G149647 belonged to the Hsp21 in Arabidopsis; GRMZM2G065355 and AC216247.3_FG001 belonged to the heat shock factor binding protein and Hsfs in Arabidopsis; the homologous genes of GRMZM2G016734, GRMZM2G042133 and GRMZM2G4483685 in Arabidopsis were the heat shock n-terminal domain protein genes. The homologous genes of GRMZM2G149647, GRMZM2G065355, GRMZM2G016734, GRMZM2G042133, and GRMZM2G448368 in rice were also heat shock-related protein genes.
Table 3.
Candidate genes for heat-stress tolerance and their annotation information and homologous genes in Arabidopsis and rice.
4. Discussion
Due to the rapid emissions of greenhouse gases, the global average temperature is increasing rapidly and is estimated to be 1.5 °C above the pre-industrial level by 2050 [10]. Maize is susceptible to heat stress, especially during the flowering and grain filling periods, resulting in significant yield losses due to grain abortion and reduced starch storage [50,51,52]. At the same time, delayed sowing means that the maize will be exposed to the long-term accumulation of high temperatures, prolonging the transition period from vegetative development to the reproductive phase and eventually leading to yield loss [53]. Heat-stress tolerance is a complex trait that is more prone to genotype–environment interactions, which requires plants to respond from multiple aspects. Breeding heat-tolerant varieties has become a critical challenge. Currently, using GWASs to mine potential QTL and genes is a widely employed approach in crop improvement. In this study, 342 inbred lines with abundant genetic variations were genotyped using a 40K-mSNP chip and various traits were evaluated in the spring and summer. In terms of genotyping, this study used mSNP and GBTS platform integration, and has several advantages in this respect. Because SNP calls can be mutually confirmed by multiple SNPs within a single mSNP site, more than 99.9% repeatability is guaranteed for both construction and sequencing. Conducted by two different systems, multiplex PCR-based GenoPlexs and solution-based capture gene baits cover a very flexible range of marker numbers, from multiple markers to 40K mSNP sites. QTL maps were plotted using a GWAS, and SNP markers related to the target traits under both conditions were analyzed. Based on the significant marker–trait associations revealed by the GWAS, these variations can be used to introgress heat-tolerant alleles into the elite germplasm through marker-assisted selection, broadening the genetic basis of the functions of heat tolerance-related genes.
The coefficient of variation of the EH, IN, MFLBL, and DTS in the summer was higher than that in the spring, and the coefficient of variation of the SRR in the summer was 119.29, indicating that when compared with spring sowing, high temperatures had different degrees of influence on all traits in the summer sowing. Comparing the average of the two seasons, it was found that the PH, EH, IN, MFLBL, and DTS had different degrees of decrease. In this study, the PH and EH of the sweet corn plants under high-temperature stress decreased by 24.0% and 36.3%, respectively, compared with those in the spring. Longmei et al. also reported reduced PH and EH under the condition of high temperatures [31]. At the same time, the SR and SSR also declined, and the coefficient of variation of the SRR in the summer reached 119.29, indicating that the temperature had different degrees of influence on each inbred line. A lower SSR may be caused by reduced assimilates in the ear and a low pollen viability. Impairment of pollen and anther development by elevated high temperature are two important factors contributing to decreased seed set in many crops [10,54,55]. Therefore, it is particularly important to explore new heat-resistant sites and varieties.
Plant morphological traits (PH and EH) were related to 6 and 3 significant SNPs, respectively. A total of 32 significant SNPs were detected in terms of DTS, with more SNPs related to the spring than to the summer. A large number of studies have shown that these traits are significantly related to maize grain yield [56,57,58,59]. The most significant loci of the SSR were all obtained by difference correlation, indicating that the SSR was more susceptible to the influence of temperature, and all loci could provide new research directions and candidate regions for the subsequent mining of heat-resistant candidate genes to improve these above traits to improve sweet corn grain yield. These QTL were all detected in one environment, indicating that different genomic regions were involved in the expression of specific traits under normal (spring) and heat-stress (summer) conditions.
The candidate genes identified by GO near the GWAS loci were enriched for programmed cell death and cellular senescence during cell development, suggesting that heat stress causes programmed cell death and senescence [31,60]. Also, the candidate genes were compared with the homologous genes in rice and Arabidopsis and annotated to obtain six candidate Hsfs or Hsps related to heat shock responses. It has been reported that increased expression of the Hsfs and Hsps could improve the heat resistance of transgenic maize [6]. Hsfs play a wide range of roles in anti-stress processes, for example, they activate the expression of protein chaperones by recognizing simple promoter binding sites, that is, heat shock elements, to prevent protein misfolding and inflammation, etc. [9,61,62]. In maize, a total of 31 Hsfs have been identified to regulate the expression of different heat shock proteins (Hsps), which contribute to heat-stress tolerance [17]. Qin et al. cloned a Hsf gene (ZmHsf11) from maize and found that the overexpression of ZmHsf11 in Arabidopsis and rice could significantly reduce the survival rate of transgenic plants under heat shock treatment and reduce their abscisic acid (ABA) sensitivity [63]. Li et al. found that overexpressed ZmHsf06 (cloned from maize leaves) in Arabidopsis plants results in enhanced basal heat tolerance and acquisition, stronger drought tolerance, and growth advantages under mild heat-stress conditions [64]. Hsps are a widespread class of heat-stress proteins that are thermally stimulated to protect the organism exposed to high temperatures. It has been reported that 22 ZmHsp70 genes are responsive to heat stress [65], and that the overexpression of Hsp101 in anthers can enhance the heat tolerance of maize [66]. The six candidate genes identified in this study would play key roles in enhancing the heat tolerance of maize and can be used in heat-tolerance breeding.
5. Conclusions
We evaluated eight yield-related agronomic traits as heat tolerance indexes using 376 sweet corn inbred lines as a natural population. Compared to the spring, heat stress in the summer had a negative effect on both plant height and ear height. The modified 40K mSNP chip was used to genotype the population, and 85 significant SNPs that were associated with the eight traits were identified. Six Hsfs or Hsps were selected as heat-tolerant candidate genes for sweet corn.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/agronomy14092171/s1. Figure S1 Temperature diagram of spring and summer in 2019 at the test base of sweet corn.
Author Contributions
Conceptualization, Q.Y., Y.X. and H.N.; Methodology, J.Z.; investigation, Q.Y.; Formal Analysis, J.Z.; Data Curation, Z.G.; Writing—Original Draft Preparation, Q.Y.; Writing—Review & Editing, H.N. and Y.X. Y.W. participated the project and revised the article. All authors have read and agreed to the published version of the manuscript.
Funding
This supported research was funded by the Key Research Area and Development Program of Guangdong Province (2018B020202008),the Agricultural Science and Technology Innovation Program (ASTIP) of the Chinese Academy of Agricultural Sciences, Shenzhen Science and Technology Program (KQTD202303010928390070), the Provincial Technology Innovation Program of Shandong, China, the Science and Technology Program of Hebei (215A7612D) and the National Key Research and Development Program of China (2023YFD1202500). Development Program of Guangdong Province funded the A.P.C.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author/s.
Acknowledgments
We are grateful to MolBreeding Biotechnology Co., Ltd., helped develop the chip.
Conflicts of Interest
Author Jianan Zhang was employed by the company MolBreeding Biotechnology Co., Ltd. The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest. Author Yunbi Xu was employed by the company BGI Bioverse. The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Luo, N.; Mueller, N.; Zhang, Y.; Feng, P.; Huang, S.; Liu, D.L.; Yu, Y.; Wang, X.; Wang, P.; Meng, Q. Short-term extreme heat at flowering amplifies the impacts of climate change on maize production. Environ. Res. Lett. 2023, 18, 084021. [Google Scholar] [CrossRef]
- Dos Santos, T.B.; Ribas, A.F.; de Souza, S.G.H.; Budzinski, I.G.F.; Domingues, D.S. Physiological responses to drought, salinity, and heat stress in plants: A review. Stresses 2022, 2, 113–135. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, J.; Li, R.; Ge, Y.; Li, Y.; Li, R. Plants’ response to abiotic stress: Mechanisms and strategies. Int. J. Mol. Sci. 2023, 24, 10915. [Google Scholar] [CrossRef] [PubMed]
- Praveen, A.; Dubey, S.; Singh, S.; Sharma, V.K. Abiotic stress tolerance in plants: A fascinating action of defense mechanisms. 3 Biotech 2023, 13, 102. [Google Scholar] [CrossRef]
- Siebert, S.; Ewert, F. Future crop production threatened by extreme heat. Environ. Res. Lett. 2014, 9, 041001. [Google Scholar] [CrossRef]
- Djalovic, I.; Kundu, S.; Bahuguna, R.N.; Pareek, A.; Raza, A.; Singla-Pareek, S.L.; Prasad, P.V.V.; Varshney, R.K. Maize and heat stress: Physiological, genetic, and molecular insights. Plant Genome 2024, 17, e20378. [Google Scholar] [CrossRef]
- Chen, Y.; Du, T.; Zhang, J.; Chen, S.K.; Fu, J.; Li, H.H.; Yang, Q. Genes and pathways correlated with heat stress responses and heat tolerance in maize kernels. Front. Plant Sci. 2023, 14, 1228213. [Google Scholar] [CrossRef]
- Sabagh, A.E.L.; Hossain, A.; Iqbal, M.A.; Barutçular, C.; Islam, M.S.; Çiğ, F.; Erman, M.; Sytar, O.; Brestic, M.; Wasaya, A.; et al. Maize adaptability to heat stress under changing climate. In Plant Stress physiology; IntechOpen: London, UK, 2020; Volume 9. [Google Scholar]
- Li, Z.; Howell, S.H. Heat stress responses and thermotolerance in Maize. Int. J. Mol. Sci. 2021, 22, 948. [Google Scholar] [CrossRef]
- Gong, W.; Oubounyt, M.; Baumbach, J.; Dresselhaus, T. Heat-stress-induced ROS in maize silks cause late pollen tube growth arrest and sterility. iScience. 2024, 27, 110081. [Google Scholar] [CrossRef]
- Wang, Y.; Tao, H.; Tian, B.; Sheng, D.; Xu, C.; Zhou, H.; Huang, S.; Wang, P. Flowering dynamics, pollen, and pistil contribution to grain yield in response to high temperature during maize flowering. Environ. Exp. Bot. 2019, 158, 80–88. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, X.; Hou, X.; Sheng, D.; Dong, X.; Gao, Y.; Wang, P.; Huang, S. Maximum lethal temperature for flowering and seed set in maize with contrasting male and female flower sensitivities. J. Agron. Crop Sci. 2021, 207, 679–689. [Google Scholar] [CrossRef]
- Qu, L.; Gu, X.; Li, J.; Guo, J.; Lu, D. Leaf photosynthetic characteristics of waxy maize in response to different degrees of heat stress during grain filling. BMC Plant Biol. 2023, 23, 469. [Google Scholar] [CrossRef] [PubMed]
- Karim, M.D.A.; Fracheboud, Y.; Stamp, P. Photosynthetic activity of developing leaves of Zea mays is less affected by heat stress than that of developed leaves. Physiol. Plant. 1999, 105, 685–693. [Google Scholar] [CrossRef]
- Elashtokhy, M.M.A.; Soaud, N.; Koul, M.; Mir, R.R.; Yan, K.; Li, J.; El-Tarabily, K.A.; Abbas, M. Heat stress-mediated constraints in maize (Zea mays) production: Challenges and solutions. Front. Plant Sci. 2022, 13, 879366. [Google Scholar]
- Gagliardi, D.; Breton, C.; Chaboud, A.; Vergne, P.; Dumas, C. Expression of heat shock factor and heat shock protein 70 genes during maize pollen development. Plant Mol. Biol. 1995, 29, 841–856. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.X.; Jiang, H.-Y.; Chu, Z.X.; Tang, X.L.; Zhu, S.W.; Cheng, B.J. Genome-wide identification, classification and analysis of heat shock transcription factor family in maize. BMC Genom. 2011, 12, 76. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Zheng, Z.; Qi, F.; Wang, J.; Wang, M.; Zhao, R.; Liu, H.; Xu, J.; Qin, L.; Dong, W.; et al. Development and evaluation of the utility of GenoBaits Peanut 40K for a peanut MAGIC population. Mol. Breed. 2023, 43, 72. [Google Scholar] [CrossRef]
- Adak, A.; Conrad, C.; Chen, Y.; Wilde, S.C.; Murray, S.C.; Anderson, I.S.L.; Subramanian, N.K. Validation of functional polymorphisms affecting maize plant height by unoccupied aerial systems discovers novel temporal phenotypes. G3 (Bethesda) 2021, 11, 6. [Google Scholar] [CrossRef]
- Ma, L.; Zhang, M.; Chen, J.; Qing, C.; He, S.; Zou, C.; Yuan, G.; Yang, C.; Peng, H.; Pan, G.; et al. GWAS and WGCNA uncover hub genes controlling salt tolerance in maize (Zea mays L.) seedlings. Theor. Appl. Genet. 2021, 134, 3305–3318. [Google Scholar] [CrossRef]
- Beló, A.; Zheng, P.; Luck, S.; Shen, B.; Meyer, D.J.; Li, B.; Tingey, S.; Rafalski, A. Whole genome scan detects an allelic variant of fad2 associated with increased oleic acid levels in maize. Mol. Genet. Genom. 2008, 279, 1–10. [Google Scholar] [CrossRef]
- Schnable, P.S.; Ware, D.; Fulton, R.S.; Stein, J.C.; Wei, F.; Pasternak, S.; Liang, C.; Zhang, J.; Fulton, L.; Graves, T.A.; et al. The B73 maize genome: Complexity, diversity, and dynamics. Science 2009, 326, 1112–1115. [Google Scholar] [CrossRef] [PubMed]
- Sahito, J.H.; Zhang, H.; Gishkori, Z.G.; Ma, C.; Wang, Z.; Ding, D.; Zhang, X.; Tang, J. Advancements and prospects of genome-wide association studies (GWAS) in maize. Int. J. Mol. Sci. 2024, 25, 1918. [Google Scholar] [CrossRef] [PubMed]
- Duan, H.; Xue, Z.; Ju, X.; Yang, L.; Gao, J.; Sun, L.; Xu, S.; Li, J.; Xiong, X.; Sun, Y.; et al. The genetic architecture of prolificacy in maize revealed by association mapping and bulk segregant analysis. Theor. Appl. Genet. 2023, 136, 182. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Zhang, M.; Liu, Y.; Liu, J.; Li, W.; Chen, G.; Peng, Y.; Jin, M.; Wei, W.; Jian, L.; et al. Genetic variation in YIGE1 contributes to ear length and grain yield in maize. New Phytol. 2022, 23, 513–526. [Google Scholar] [CrossRef]
- Qian, F.; Jing, J.; Zhang, Z.; Chen, S.; Sang, Z.; Li, W. GWAS and Meta-QTL analysis of yield-related ear traits in maize. Plants 2023, 12, 3806. [Google Scholar] [CrossRef]
- Zeng, T.; Meng, Z.; Yue, R.; Lu, S.; Li, W.; Li, W.; Meng, H.; Sun, Q. Genome wide association analysis for yield related traits in maize. BMC Plant Biol. 2022, 22, 449. [Google Scholar] [CrossRef]
- Yan, P.; Du, Q.; Chen, H.; Guo, Z.; Wang, Z.; Tang, J.; Li, W.X. Biofortification of iron content by regulating a NAC transcription factor in maize. Science 2023, 382, 1159–1165. [Google Scholar] [CrossRef]
- Wang, X.; Wang, H.; Liu, S.; Ferjani, A.; Li, J.; Yan, J.; Yang, X.; Qin, F. Genetic variation in ZmVPP1 contributes to drought tolerance in maize seedlings. Nat. Genet. 2016, 48, 1233–1241. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, J.; Xu, Q.; Wang, D.; Di, H.; Huang, J.; Yang, X.; Wang, Z.; Zhang, L.; Dong, L.; et al. Identification of candidate tolerance genes to low-temperature during maize germination by GWAS and RNA-seqapproaches. BMC Plant Biol. 2020, 20, 333. [Google Scholar] [CrossRef]
- Guo, J.; Li, C.; Zhang, X.; Li, Y.; Zhang, D.; Shi, Y.; Song, Y.; Li, Y.; Yang, D.; Wang, T. Transcriptome and GWAS analyses reveal candidate gene for seminal root length of maize seedlings under drought stress. Plant Sci. 2020, 292, 110380. [Google Scholar] [CrossRef]
- Li, C.; Guo, J.; Wang, D.; Chen, X.; Guan, H.; Li, Y.; Zhang, D.; Liu, X.; He, G.; Wang, T.; et al. Genomic insight into changes of root architecture under drought stress in maize. Plant Cell Environ. 2023, 46, 1860–1872. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Wang, S.; Zhao, S.; Chen, D.; Tian, H.; Li, J.; Zhang, L.; Li, S.; Liu, L.; Shi, C.; et al. Global crotonylatome and GWAS revealed a TaSRT1-TaPGK model regulating wheat cold tolerance through mediating pyruvate. Sci. Adv. 2023, 9, 1012. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Zou, C.; Liu, X.; Wang, S.; Li, W.X.; Jeffers, D.; Fan, X.; Xu, M.; Xu, Y. Complex genetic system involved in fusarium ear rot resistance in maize as revealed by GWAS, bulked sample analysis, and genomic prediction. Plant Dis. 2020, 10, 1725–1735. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Liu, S.; Pei, Y.; Jiang, X.; Jaqueth, J.S.; Li, B.; Han, J.; Jeffers, D.; Wang, J.; Song, X. Identification of genetic loci associated with rough dwarf disease resistance in maize by integrating GWAS and linkage mapping. Plant Sci. 2022, 315, 111100. [Google Scholar] [CrossRef] [PubMed]
- Shu, G.; Wang, A.; Wang, X.; Ding, J.; Chen, R.; Gao, F.; Wang, A.; Li, T.; Wang, Y. Identification of southern corn rust resistance QTNs in Chinese summer maize germplasm via multi-locus GWAS and post-GWAS analysis. Front. Plant Sci. 2023, 14, 1221395. [Google Scholar] [CrossRef] [PubMed]
- Warburton, M.L.; Womack, E.D.; Tang, J.D.; Thrash, A.; Smith, J.S.; Xu, W.; Murray, S.C.; Williams, W.P. Genome-wide association and metabolic pathway analysis of corn earworm resistance in maize. Plant Genome 2018, 11, 1. [Google Scholar] [CrossRef]
- Tamiru, A.; Paliwal, R.; Manthi, S.J.; Odeny, D.A.; Midega, C.A.O.; Khan, Z.R.; Pickett, J.A.; Bruce, T.J.A. Genome wide association analysis of a stemborer egg induced “call-for-help” defence trait in maize. Sci. Rep. 2020, 10, 11205. [Google Scholar] [CrossRef]
- Washburn, J.D.; LaFond, H.F.; Lapadatescu, M.C.; Pereira, A.E.; Erb, M.; Hibbard, B.E. GWAS analysis of maize host plant resistance to western corn rootworm (Coleoptera: Chrysomelidae) reveals candidate small effect loci for resistance breeding. J. Econ. Entomol. 2023, 116, 2184–2192. [Google Scholar] [CrossRef]
- Ahmed, Z.; Khalid, M.; Ghafoor, A.; Shah, M.K.N.; Raja, G.K.; Rana, R.M.; Mahmood, T.; Thompson, A.M. SNP-based genome-wide association mapping of pollen viability under heat stress in tropical Zea mays L. Inbred Lines. Front. Genet. 2022, 13, 819849. [Google Scholar] [CrossRef]
- Longmei, N.; Gill, G.K.; Zaidi, P.H.; Kumar, R.; Nair, S.K.; Hindu, V.; Vinayan, M.T.; Vikal, Y. Genome wide association mapping for heat tolerance in sub-tropical maize. BMC Genom. 2021, 22, 154. [Google Scholar] [CrossRef]
- NY/T 2232-2012; Guidelines for the Conduct of Tests for Distinctness, Uniformity and Stability. Maize (Zea mays L.). National public Service Platform for Standards Information. Ministry of Agriculture of the People’s Republic of China: Beijing, China, 2012.
- Murray, M.G.; Thompson, W.F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res. 1980, 8, 4321–4325. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Yang, Q.; Huang, F.; Zheng, H.; Sang, Z.; Xu, Y.; Zhang, C.; Wu, K.; Tao, J.; Prasanna, B.M.; et al. Development of high-resolution multiple-SNP arrays for genetic analyses and molecular breeding through genotyping by target sequencing and liquid chip. Plant Commun. 2021, 2, 100230. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef]
- Shi, J.; Wang, Y.; Wang, C.; Wang, L.; Zeng, W.; Han, G.; Qiu, C.; Wang, T.; Tao, Z.; Wang, K.; et al. Linkage mapping combined with GWAS revealed the genetic structural relationship and candidate genes of maize flowering time-related traits. BMC Plant Biol. 2022, 22, 328. [Google Scholar] [CrossRef]
- Lipka, A.E.; Tian, F.; Wang, Q.; Peiffer, J.; Li, M.; Bradbury, P.J.; Gore, M.A.; Buckler, E.S.; Zhang, Z. GAPIT: Genome association and prediction integrated tool. Bioinformatics 2012, 28, 2397–2399. [Google Scholar] [CrossRef]
- Schoper, J.B.; Lambert, R.J.; Vasilas, B.L. Maize pollen viability and ear receptivity under water and high-temperature stress. Crop Sci. 1986, 26, 1029–1033. [Google Scholar] [CrossRef]
- Zhang, X.; Guan, Z.; Li, Z.; Liu, P.; Ma, L.; Zhang, Y.; Pan, L.; He, S.; Zhang, Y.; Li, P.; et al. A combination of linkage mapping and GWAS brings new elements on the genetic basis of yield-related traits in maize across multiple environments. Theor. Appl. Genet. 2020, 133, 2881–2895. [Google Scholar] [CrossRef]
- Edreira, J.I.R.; Otegui, M.E. Heat stress in temperate and tropical maize hybrids: A novel approach for assessing sources of kernel loss in field conditions. Field Crop. Res. 2013, 142, 58–67. [Google Scholar] [CrossRef]
- Dhaliwal, D.S.; Williams, M.M. Evidence of sweet corn yield losses from rising temperatures. Sci. Rep. 2022, 12, 18218. [Google Scholar] [CrossRef]
- Cao, Z.-Y.; Chen, Z.H.; Tang, B.; Zeng, Q.; Guo, H.L.; Huang, W.H.; Luo, Y.; Shen, S.; Zhou, S.L. The effects of sowing date on maize: Phenology, morphology, and yield formation in a hot subtropical monsoon region. Field Crop. Res. 2024, 309, 109309. [Google Scholar] [CrossRef]
- Peet, M.M.; Sato, S.; Gardner, R.G. Comparing heat stress effects on male-fertile and male-sterile tomatoes. Plant Cell Environ. 1998, 21, 225–231. [Google Scholar] [CrossRef]
- Sato, S.; Kamiyama, M.; Iwata, T.; Makita, N.; Furukawa, H.; Ikeda, H. Moderate increase of mean daily temperature adversely affects fruit set of Lycopersicon esculentum by disrupting specific physiological processes in male reproductive development. Ann. Bot. 2006, 97, 731–738. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Guo, W.; Le, L.; Yu, J.; Wu, Y.; Li, D.; Wang, Y.; Wang, H.; Lu, X.; Qiao, H.; et al. Integration of high-throughput phenotyping, GWAS, and predictive models reveals the genetic architecture of plant height in maize. Mol. Plant. 2023, 1, 354–373. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Bi, Y.; Jiang, F.; Guo, R.; Zhang, Y.; Fan, J.; Kang, M.S.; Fan, X. Fine mapping of candidate quantitative trait loci for plant and ear height in a maize nested-association mapping population. Front. Plant Sci. 2022, 13, 963985. [Google Scholar] [CrossRef]
- Shu, G.; Wang, A.; Wang, X.; Chen, R.; Gao, F.; Wang, A.; Li, T.; Wang, Y. Identification of QTNs, QTN-by-environment interactions for plant height and ear height in maize multi-environment GWAS. Front. Plant Sci. 2023, 14, 1284403. [Google Scholar] [CrossRef]
- Du, L.; Zhang, H.; Xin, W.; Ma, K.; Du, D.; Yu, C.; Liu, Y. Dissecting the genetic basis of flowering time and height related-traits using two doubled haploid populations in maize. Plants 2021, 10, 1585. [Google Scholar] [CrossRef]
- McNellie, J.P.; Chen, J.; Li, X.; Yu, J. Genetic mapping of foliar and tassel heat stress tolerance in maize. Crop Sci. 2018, 58, 2484–2493. [Google Scholar] [CrossRef]
- Gomez-Pastor, R.; Burchfiel, E.T.; Thiele, D.J. Regulation of heat shock transcription factors and their roles in physiology and disease. Nat. Rev. Mol. Cell Bio. 2018, 19, 4–19. [Google Scholar] [CrossRef]
- Tan, K.; Fujimoto, M.; Takii, R.; Takaki, E.; Hayashida, N.; Nakai, A. Mitochondrial SSBP1 protects cells from proteotoxic stresses by potentiating stress-induced Hsf1 transcriptional activity. Nat. Commun. 2015, 6, 6580. [Google Scholar] [CrossRef]
- Qin, Q.; Zhao, Y.; Zhang, J.; Chen, L.; Si, W.; Jiang, H. A maize heat shock factor ZmHsf11 negatively regulates heat stress tolerance in transgenic plants. BMC Plant Biol. 2022, 22, 406. [Google Scholar] [CrossRef] [PubMed]
- Li, H.C.; Zhang, H.N.; Li, G.L.; Liu, Z.H.; Zhang, Y.M.; Zhang, H.M.; Guo, X.L. Expression of maize heat shock transcription factor gene ZmHsf06 enhances the thermotolerance and drought-stress tolerance of transgenic Arabidopsis. Funct. Plant Biol. 2015, 42, 1080–1091. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Hu, W.; Qian, Y.; Ren, Q.; Zhang, J. Genome-wide identification, classification and expression analysis of the Hsf and Hsp70 gene families in maize. Gene 2021, 770, 145348. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Huang, Y.; Sun, H.; Wang, T.; Ru, W.; Pan, L.; Zhao, X.; Dong, Z.; Huang, W.; Jin, W. Heat shock protein 101 contributes to the thermotolerance of male meiosis in maize. Plant Cell 2022, 34, 3702–3717. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).