Abstract
The plant pathogenic fungus Botrytis cinerea causes significant losses in agricultural production and it is rather difficult to control due to its broad host range and environmental persistence. The management of gray mold disease is still mainly based on the use of chemicals, which could have harmful effects not only due to impacts on the environment and human health, but also because they favor the development of fungicide-resistant strains. In this scenario, the strategy of RNA interference (RNAi) is being widely considered, and Spray-Induced Gene Silencing (SIGS) is gaining interest as a versatile, sustainable, effective, and environmentally friendly alternative to the use of chemicals in the protection of crops. The SIGS approach was evaluated to control B. cinerea infection on lettuce plants. In vitro-synthesized dsRNA molecules (BcBmp1-, BcBmp3-, and BcPls1-dsRNAs) were used naked, or complexed to small layered double hydroxide (sLDH) clay nanosheets. Therefore, treatments were applied by pressure spraying whole lettuce plants lately inoculated with B. cinerea. All sprayed dsRNAs proved effective in reducing disease symptoms with a notable reduction compared to controls. The effectiveness of SIGS in reducing disease caused by B. cinerea was high overall and increased significantly with the use of sLDH clay nanosheets. The sLDH clay nanosheet–dsRNA complexes showed better plant protection over time compared to the use of naked dsRNA and this was particularly evident at 27 days post-inoculation. RNAi-based biocontrol could be an excellent alternative to chemical fungicides, and several RNAi-based products are expected to be approved soon, although they will face several challenges before reaching the market.
1. Introduction
Plant diseases caused by fungi and, in particular, Botrytis cinerea (Pers., 1794), have overwhelming global economic and agronomic importance due to their effects in reducing agricultural productions [1,2,3]. Current fungal disease management still relies primarily on chemicals that, in addition to harmful effects on the environment and human health, can also promote the development of fungicide-resistant strains of the pathogen [4,5].
Lettuce (Lactuca sativa L.) is among the most important horticultural crops for fresh consumption worldwide, and its remarkable flexibility in cultivation systems makes it available practically all year round. Lettuce is a commonly consumed leafy vegetable and is considered a rich source of vitamins (A, C, E, and K), polyphenols, and antioxidant compounds [6]. The beginning of lettuce cultivation dates back more than 4500 years ago, and different morphotypes have been selected and cultivated. However, a depletion of genetic variability is now evident which poses problems for genetic improvement [7]. Among the infectious diseases that negatively affect the productivity of lettuce, there are about ten particularly fearsome fungal diseases [8]. Faced with this vulnerability, the control of plant diseases using fungicides poses serious practical problems such as the containment of residues in commercial products, which must be carefully managed [9]. Gray mold caused by the fungus B. cinerea has been considered a major disease in greenhouse-grown lettuce. The romaine lettuce variety and some iceberg lettuces are susceptible to B. cinerea even in the field [10].
Climate change will increase biotic stresses; in fact, it will contribute to creating environmental pressures that could foster new diseases caused by fungi [11]. The development of new control strategies is necessary to help increase food production. In this context, Spray-Induced Gene Silencing (SIGS), the non-transformative technology based on RNA interference (RNAi), is gaining ground as a sustainable, effective, and environmentally friendly alternative to chemicals in crop protection [12,13,14,15,16]. Notably, SIGS does not require transgenic modifications and allows the design of highly specific double-stranded RNA (dsRNA) molecules. Moreover, in this strategy, the choice of specific sequences targeting key pathogen genes allows for efficiency, selectivity, and avoidance of off-target problems [17]. Topical application of dsRNA is a promising tool against plant fungal diseases not only in the field but also in postharvest storage. In fact, RNAi-based biocontrol is considered a low-impact strategy due to its biodegradability in the environment [18]. Reported delivery techniques include foliar spraying, seed and fruit treatment, wood injection, and plant or root uptake [12,19,20,21]. In view of large-scale applications, the foliar spraying technique has the potential to be considered the most effective and feasible method of dsRNA delivery in terms of cost and time [22].
From the perspective of developing an RNAi-based product for field spray applications, two aspects of dsRNA production must first be considered: quantity and stability. The in vivo production of dsRNA involves the use of yeasts (e.g., Yarrowia lipolytica) or engineered bacteria (Escherichia coli or Pseudomonas syringae) as biofactories, which allow more dsRNA to be synthesized than in vitro production through enzymatic activity, with the advantage of reducing process costs [23,24,25,26]. Recently, the RNase III-deficient E. coli strain HT115(DE3) was successfully used to produce dsRNAs against B. cinerea and to target essential genes such as BcCYP51, Bcchs1, and BcEF2 [20,27], and the BcSas1 gene involved in the fungal secretory pathway [28].
Regarding the stability of dsRNA, the use of nanoparticles could help protect dsRNA molecules during their delivery in topical applications [22,29,30,31,32,33]. The nanoparticles, ranging in size from 1 to 500 nm and positively charged, can bind dsRNA to form a complex that allows the sustained release of dsRNA over time and its protection from UV light and nuclease degradation [29,34]. Nanoparticles can confer greater persistence to dsRNA when sprayed onto the leaf surface, as has been demonstrated for layered double hydroxide clay (LDH) nanosheets called BioClay™ [29,35,36]. Very recently, dsRNA loading to LDH nanosheets was demonstrated to be effective in controlling B. cinerea infection. Notably, LDH nanosheets resulted in reduced fruit decay development and greater dsRNA integrity compared to the naked one [36,37].
In this work, the feasibility of spray application of in vitro-produced dsRNAs targeting the sensu stricto virulence genes BcBmp1, BcBmp3, and BcPls1 to reduce gray mold disease in in vivo experiments was evaluated. The two mitogen-activated protein kinases BcBmp1 and BcBmp3 genes were selected since they control many pathogenesis-related fungal processes through extracellular signal transduction. In addition, the tetraspanin BcPls1 gene was targeted, since it also plays an important role as a virulence factor in appressoria-forming pathogens such as B. cinerea.
Previously, the efficacy of topical applications of exogenous dsRNAs targeting the two mitogen-activated protein kinase genes (BcBmp1 and BcBmp3) and the tetraspanin gene BcPls1 was demonstrated both in vitro on axenic cultures of B. cinerea and in vivo on detached lettuce leaves. We also demonstrated that dsRNA treatments were able to effectively reduce the transcript levels of the three target genes (BcBmp1, BcBmp3, and BcPls1) and the necrotic areas caused by B. cinerea on detached lettuce leaves [38,39]. Here, dsRNA molecules were used naked or complexed with sLDH clay nanosheets and were applied by pressure spraying onto whole lettuce plants with the aim of testing the effectiveness of the RNAi strategy in a situation closer to crop protection in vivo, moving away from laboratory conditions. A chemical fungicide was also included among the treatments to evaluate whether SIGS strategy can compete with protection using chemical treatments. Disease assessment was carried out using phytopathometry (qualitative ordinal scale of disease symptoms, disease severity index or McKinney index) [40] and the effectiveness of treatments was evaluated using the Abbott Index.
2. Materials and Methods
2.1. Fungal Culture Conditions
The fungal pathogen used in this study is Botrytis cinerea B05.10, a haploid strain resulting from benomyl treatment of the wild-type isolate SAS56 [41,42]. It was incubated on Lettuce Malt Agar plates (LMA: 20 g/L of Malt extract (Biolife Italiana S.r.l., Milano, Italy), 20 g/L of agar (Sigma-Aldrich, L-3022, Saint Louis, MO, USA), and 100 g/L of homogenized healthy lettuce leaves) at 25 °C with a 12/12 NUV/light cycle. Conidial suspensions of B. cinerea B05.10 were prepared from 7- to 10-day-old LMA plates by gently scraping conidia from the surface of the LMA culture with a sterile spatula. The suspension was prepared in Sabouraud Maltose Broth [(SMB: 10 g/L mycological peptone (Sigma-Aldrich): 40 g/L maltose (Sigma-Aldrich), with MilliQ water (EASYpure® II LF, Thermo Scientific, Waltham, MA, USA; resistivity, 18.2 MΩ/cm)], pH 5.6 ± 0.2. The resulting conidial suspension was filtered through a layer of sterile Miracloth (Calbiochem, San Diego, CA, USA), and the conidia concentration was checked with a hemacytometer (Bürker, LO—Laboroptik Ltd., Lancing, UK) and adjusted to the desired value with SMB.
2.2. Preparation and Characterization of Small MgAl-Layered Double Hydroxide (sLDH) Clay Nanosheets
sLDH clay nanosheets were prepared according to a non-aqueous method described by Dong et al. [43], with minor changes. After precipitation, the heat treatment step was skipped by directly moving on to the purification and dispersion of the sLDH clay nanosheets in water to achieve an average particle size of ~40 nm.
Small aliquots of sLDH clay nanosheets suspension were dried at 65 °C in a microtubes thermoshaker (BioSan TS100C, Riga, Latvia) and the different tubes’ weights before and after drying were measured to estimate the sLDH clay nanosheets’ concentration.
The produced sLDH clay nanosheets were analyzed microscopically by Transmission Electron Microscopy (TEM) using a JEM 100 SX (JEOL) at 100 kV, which provided information on the morphology of the nanoparticles. In addition, TEM images allowed the evaluation of the average size and distribution of the sLDH clay nanosheets, where the lateral dimension was measured (n = 150) using ImageJ software, version 1.53a.
2.3. Preparation of sLDH Clay Nanosheet–dsRNA Complexes and RNase A Protection Assay
To achieve improved and prolonged stability of the in vitro-synthesized dsRNA, it was loaded to sLDH clay nanosheets. Preparation of sLDH clay nanosheet–dsRNA complexes was conducted by mixing 500 ng of in vitro-produced dsRNAs (BcBmp1-, BcBmp3-, BcPls1-, and Green Fluorescent Protein (GFP)-dsRNAs) with pristine sLDH clay nanosheets at 25 °C for 30 min with gentle orbital shaking (300 rpm) in a microtubes thermoshaker (Biosan TS100C) at a final volume of 10 µL. To optimize dsRNA loading, intercalation experiments were carried out with different mass ratios of dsRNA/sLDH clay nanosheets (from 1:2 to 1:10). The capability of sLDH clay nanosheets to bind dsRNAs was evaluated by a gel retardation assay. Samples were analyzed by electrophoresis in 0.5× TBE buffer with 1% (w/v) agarose gels and detected by UV fluorescence after GelRed™ staining (Biotium Inc., Fremont, CA, USA), according to the manufacturer’s instructions. The 100 bp DNA ladder (Promega, Madison, WI, USA) was used as a molecular size marker. In the electrophoresis analysis, naked dsRNA (not loaded to sLDH clay nanosheets) and aliquots of sLDH clay nanosheets’ suspension alone were used as control samples.
The ability of sLDH clay nanosheets to protect dsRNA from probable environmental degradation was evaluated using an in vitro transcribed control dsRNA (500 bp) following the method described by Mitter et al. [29], with minor changes. An amount of 1.5 µg of control dsRNA and control dsRNA complexed with sLDH clay nanosheets (ratio 1:10) were exposed to RNase A (Sigma-Aldrich, R-4642). Samples (10 µL) were treated with 0.375 ng or 0.750 ng of RNase A (50 mM NaCl) for 5 min at 37 °C. The dsRNA from treated and untreated sLDH clay nanosheet–dsRNA complexes was released from the sLDH clay nanosheets prior to gel electrophoresis using the acid release buffer (4.11 mL of 0.2 M Na2HPO4 + 15.89 mL of 0.1 M citric acid; pH 3) in a volume ratio of 5:1. Samples were analyzed by electrophoresis as described above.
2.4. In Vitro Effects of sLDH Clay Nanosheets on the Growth of Botrytis cinerea
To evaluate the effects of sLDH clay nanosheets on fungal growth, 96-well polystyrene microtiter plates (Cellstar®, Greiner Bio-One, Frickenhausen, Germany) were used. Aliquots (5 × 102 spores) of a B. cinerea conidial suspension prepared in SMB were added to the wells (n = 8 for each treatment in a total volume of 100 µL per assay). Treatments were prepared in MilliQ water and consisted of: (i) B. cinerea conidia + TE buffer (CTRL, 10 µM Tris/1.0 µM EDTA, pH 7.0); (ii) B. cinerea conidia + sLDH clay nanosheets; and (iii) TE buffer (blank background control). The volume of TE used was equal to the volume of dsRNA used in the treatment, while the volume of sLDH clay nanosheets corresponded to the 1:2 ratio used in the sLDH clay nanosheet–dsRNA complexes. Plates were incubated at 25 ± 1 °C with a 12/12 NUV/light cycle and fungal growth was measured by analyzing the optical density (OD) at 595 nm using a microplate reader spectrophotometer (Bio-Rad, Model 680, Bio-Rad Laboratories, Hercules, CA, USA) at different times ranging from 0 to 96 h. The experiment was repeated twice. Absorbance values were converted to the percentage of fungal growth relative to the control (100%) using the following formula: {[(OD595Control − OD595Treated)/OD595Control)] × 100}.
2.5. Spray-Induced Gene Silencing (SIGS): Application of Naked dsRNAs or sLDH Clay Nanosheet–dsRNA Complexes and Botrytis cinerea Conidia by Pressure Spraying
The in vitro-produced dsRNAs were tested for their efficacy in controlling B. cinerea disease through spray applications on Lactuca sativa cv. ‘Romana’ plants (Romana Verde degli Ortolani, Sementi Dom Dotto S.p.A., Udine, Italy) after 4–5 weeks of cultivation in pots (capacity 150–200 mL). Lettuce plants were grown in a climate chamber (Fratelli Bertagnin, Bologna, Italy) with a 12 h photoperiod at 22 ± 1 °C with 65% relative humidity (RH).
The dsRNAs and fungicide were sprayed from a 2–4 cm distance on both the abaxial and adaxial sides of the leaves using a gravity airbrush gun (FENGDA FE-186K, Ningbo City, China) with a 0.3 mm nozzle at a pressure of 2.5 bar provided by a mini compressor (Model 2010900, LTF Spa, Antegnate, BG, Italy).
For each plant, 500 µL of dsRNAs, naked or complexed to sLDH clay nanosheet solutions (20 ng/µL: BcBmp1-, BcBmp3-, BcPls1-, and GFP-dsRNAs), or sterile MilliQ water + TE buffer (CTRL) were sprayed at the beginning of the experiment (T) (Figure 1a). The volume of TE exactly corresponded to the volume of dsRNA added for each treatment. In the case of using sLDH clay nanosheet–dsRNA complexes, the 1:2 ratio was selected in our experiments. The treated lettuce plants were maintained in the climate chamber as described before.

Figure 1.
(a) Schematic representation of SIGS experiments against B. cinerea performed in this study. T = Treatment; dpt = days post-treatment; dpi = days post-inoculation. (b) Empirical qualitative ordinal scale used to evaluate gray mold disease symptoms on infected lettuce plants.
B. cinerea B05.10 was inoculated (1 mL of conidial suspension = 1 × 104 spores) by spraying the plants 2 days after the treatments (2 dpt) using the gravity airbrush gun and the same conditions described before. Artificially inoculated plants were incubated in transparent plastic propagation boxes at 25 ± 1 °C with 98–99% RH under natural lighting conditions in the laboratory for 6 days (6 dpi). After this time, they were transferred to the climate chamber where they remained for up to 27 dpi (Figure 1a).
For each experiment, eight or sixteen plants (n = 8; n = 16) per treatment were used, unless otherwise specified. The experiments performed were the following:
- Plants were treated with single dsRNAs (BcBmp1-, BcBmp3-, BcPls1-, and GFP-dsRNAs; 10 µg per plant) or sterile MilliQ water + TE buffer (CTRL);
- Plants were treated with all three dsRNAs mixed together (BcBmp1-dsRNA + BcBmp3-dsRNA + BcPls1-dsRNA; 10 µg each for a total of 30 µg per plant), or GFP-dsRNA (30 µg per plant), or sterile MilliQ water + TE buffer (CTRL);
- Plants were treated with single dsRNAs (BcBmp1-, BcBmp3-, BcPls1-, and GFP-dsRNAs; 10 µg per plant) complexed to sLDH clay nanosheets, or sterile MilliQ water + TE buffer (CTRL), or sLDH clay nanosheets.
- Plants were treated with the fungicide Switch® 62.5 WG (Syngenta, 37.5% cyprodinil and 25% fludioxonil, WG = water-dispersible granules) at the recommended dose (0.7 g/L) for lettuce.
2.6. Disease Assessment
In all SIGS experiments, disease symptoms were assessed and photographed 6 and 27 days after B. cinerea inoculation (6 dpi and 27 dpi) by evaluating individual leaves. At 6 dpi, we scored disease symptoms on the 4/5 outer true leaves present on the plants at the beginning of the experiment when dsRNAs or water were applied. Immediately after the disease assessment, these leaves were removed. The second sampling was conducted at 27 dpi on the 10 outermost new leaves that were still forming at the beginning of the experiment. In the latter case, the inoculum of B. cinerea was represented by the conidia present on the plants and which spread into the growth chamber, simulating what happens naturally.
We assessed the gray mold symptoms on lettuce leaves using an empirical qualitative ordinal scale with 9 infection classes, described and depicted in Figure 1b.
Each lettuce leaf was assigned to one of nine infection classes, and we calculated the McKinney index (MKI), which incorporates both the incidence and severity of the disease at 6 dpi and 27 dpi for each plant [44]. The MKI was calculated as follows:
where d is the class of disease infection scored, f is the disease frequency, N is the total number of organs examined, and D is the highest class of disease infection that occurred in the empirical qualitative ordinal scale.
The efficacy of the treatments was calculated from the MKI data using the Abbott Index (AI) [45] with the following formula:
where MKIuntreated stands for the mean of MKI in the untreated control and MKItreated stands for the mean of a given treatment.
Efficacy (%) = [(MKIuntreated − MKItreated)/MKIuntreated] × 100
2.7. Statistical Analysis
Data from SIGS experiments were expressed as MKI (%) and AI (%) and those from the in vitro assay in 96-microtiter plates were converted as growth percentages of the untreated control. All data were subjected to an analysis of variance (ANOVA) using the statistical program CoStat 6.4 (Cohort Software, Monterey, CA, USA). Percentage data were transformed into arcsine √% before the ANOVA. All means were separated by Tukey’s honestly significant difference post hoc (HSD) test (p ≤ 0.001 for SIGS experiments and p ≤ 0.05 for in vitro assay). The normality of the data was tested using the Shapiro–Wilk test, while the homoscedasticity was tested using the Bartlett test.
3. Results
3.1. Spray-Induced Gene Silencing (SIGS): Application of Naked dsRNAs by Pressure Spraying Reduces Botrytis cinerea Symptoms on Lettuce Plants
The in vitro-produced BcBmp1-, BcBmp3-, and BcPls1-dsRNAs were used to challenge B. cinerea in vivo during spray application experiments on whole lettuce plants. The efficacy of the dsRNA treatment was evaluated starting from an empirical qualitative ordinal scale, which helped in the calculation of the McKinney index (MKI%) and Abbott Index (AI%), two useful indices during the analysis of the disease symptoms. Hence, all data presented below are in percentage (%) form.
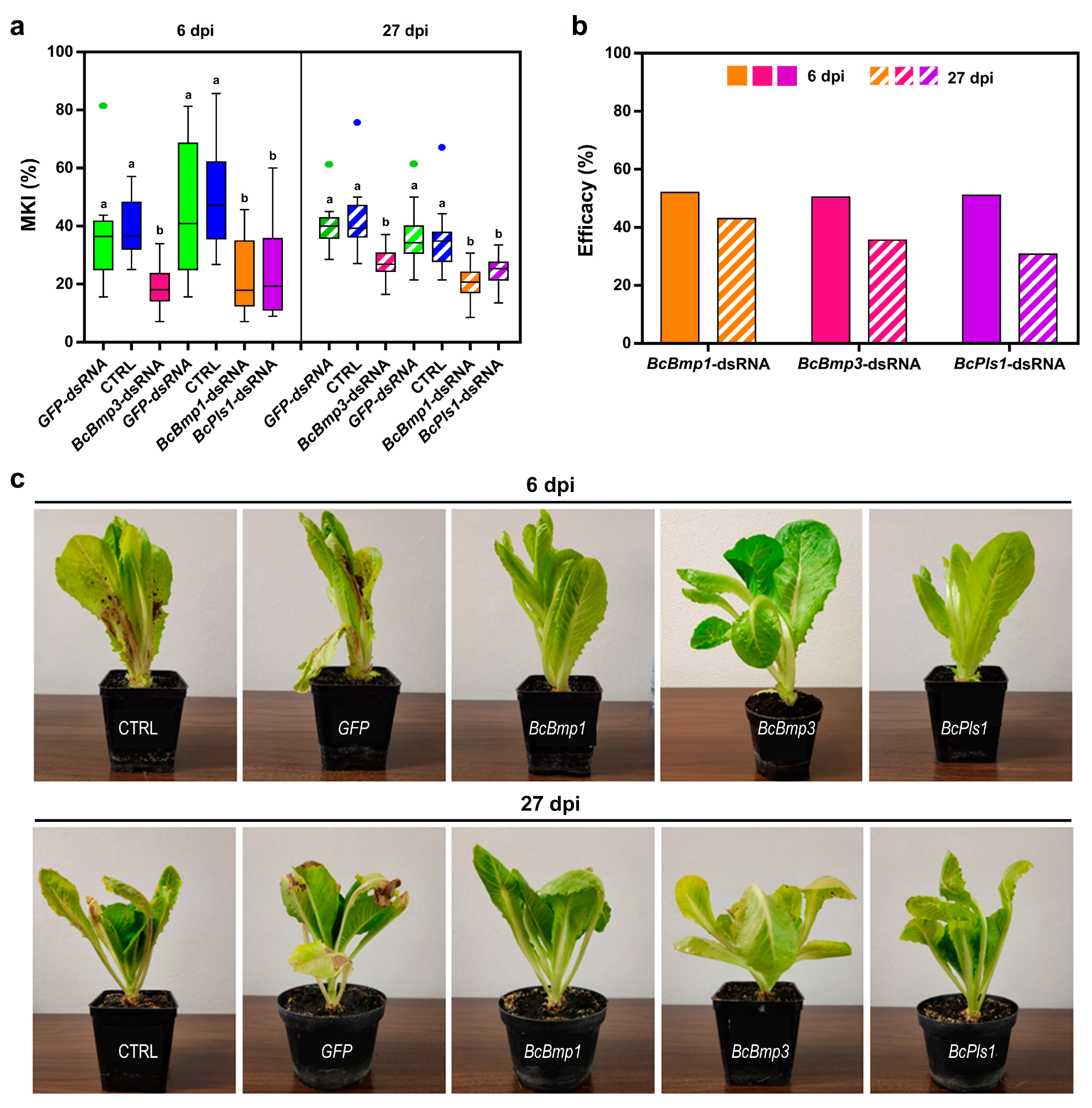
In the case of naked dsRNA, from the MKI analysis at 6 dpi, it was found that the BcBmp1-, BcBmp3-, and BcPls1-dsRNAs were all effective in reducing disease symptoms caused by B. cinerea. In particular, for BcBmp3-dsRNA, the MKI was 19.8 ± 1.9 compared to controls (MKICTRL = 39.9 ± 2.5 and MKIGFP-dsRNA = 36.2 ± 3.7). In the case of BcBmp1-dsRNA and BcPls1-dsRNA, the MKI was 23.9 ± 3.3 and 24.4 ± 3.7, respectively, compared to controls (MKICTRL = 45.4 ± 5.5 and MKIGFP-dsRNA = 49.9 ± 4.4 (Figure 2a)).
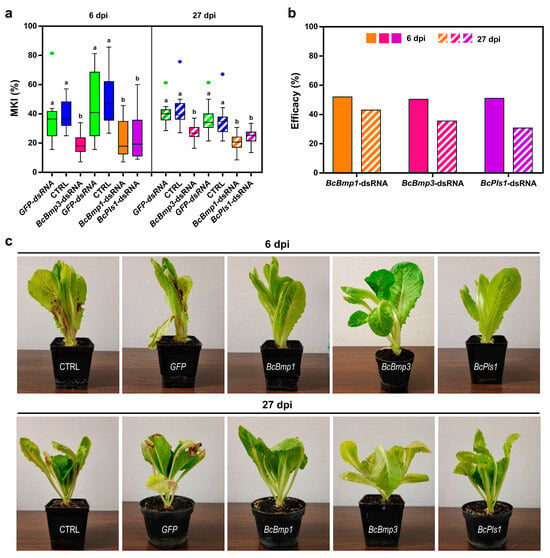
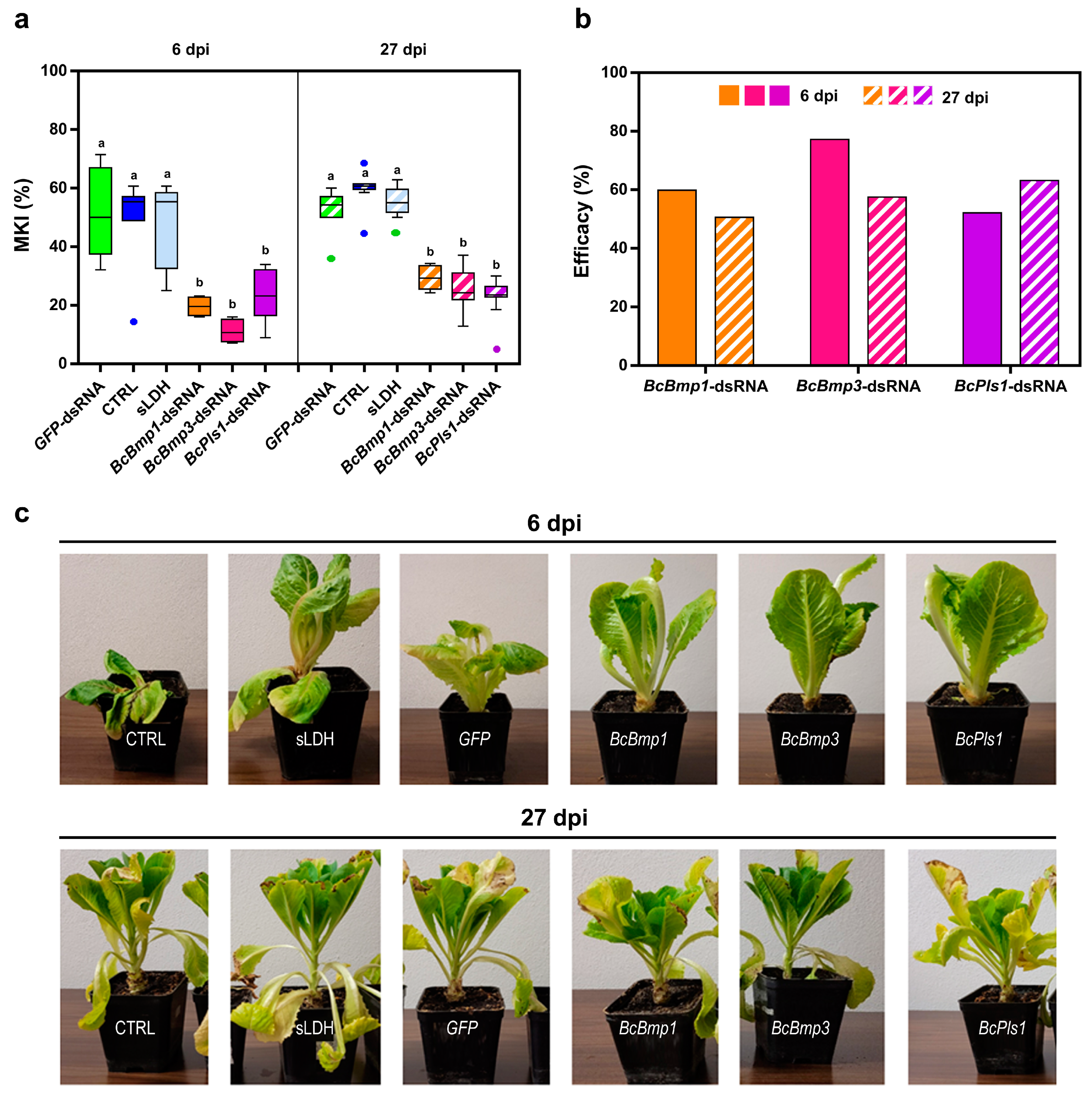
Figure 2.
The assessment of disease symptoms caused by B. cinerea and the relative effectiveness of dsRNA treatment are shown. (a) The McKinney index (MKI) was used to evaluate disease symptoms after spray applications of in vitro-produced BcBmp1-, BcBmp3-, and BcPls1-dsRNAs (10 µg per plant) and controls (CTRL and GFP-dsRNA) at 6 dpi and 27 dpi. Graphs and box plots show data from sixteen biological replicates (n = 16) of the experiment. The circles on box plots indicate 5th and 95th percentile outliers. Same letters above the bars indicate no significant differences from each other (ANOVA) according to Tukey’s test (p ≤ 0.001). (b) The efficacy of spray applications of in vitro-produced BcBmp1-, BcBmp3-, and BcPls1-dsRNAs was assessed at 6 dpi and 27 dpi by calculating the Abbott Index (AI) from the McKinney index (MKI). (c) Representative images of potted lettuce plants showing symptoms caused by B. cinerea when sampled at 6 dpi and 27 dpi. Symptoms of gray mold disease were analyzed in controls [CTRL and GFP-dsRNA (GFP)] and in lettuce plants treated with BcBmp1-, BcBmp3-, and BcPls1-dsRNAs (BcBmp1, BcBmp3, and BcPls1).
At 27 dpi, the MKI analysis showed that all sprayed naked BcBmp1-, BcBmp3- and BcPls1-dsRNAs were still effective in reducing disease symptoms caused by B. cinerea. In particular, for BcBmp3-dsRNA, the MKI was 27.6 ± 1.4 compared to controls (MKICTRL = 42.9 ± 2.7 and MKIGFP-dsRNA = 41.8 ± 2.2), and in the case of BcBmp1-dsRNA and BcPls1-dsRNA, the MKI was 20.3 ± 1.3 and 24.6 ± 1.3, respectively, compared to controls (MKICTRL = 35.6 ± 2.6 and MKIGFP-dsRNA = 36.6 ± 2.5) (Figure 2a).
The efficacy of dsRNA treatments calculated with the Abbot Index was 52.1 at 6 dpi and 43.0 at 27 dpi for BcBmp1-dsRNA, 50.5 at 6 dpi and 35.6 at 27 dpi for BcBmp3-dsRNA, and 51.1 at 6 dpi and 30.8 at 27 dpi for BcPls1-dsRNA. The effectiveness of the dsRNA treatment is slightly reduced at 27 dpi but is still consistent, and the trend shown at 6 dpi still persists after 3 weeks during the second sampling (Figure 2b,c).
The in vitro-produced BcBmp1-, BcBmp3-, and BcPls1-dsRNAs were also used together during spray application experiments to evaluate their possible synergistic activity. However, in the case of the naked dsRNAs mix, MKI analysis at 6 dpi and 27 dpi showed no significant differences between the treatment (dsRNAs) and controls (CTRL and GFP-dsRNA) (Figure 3).
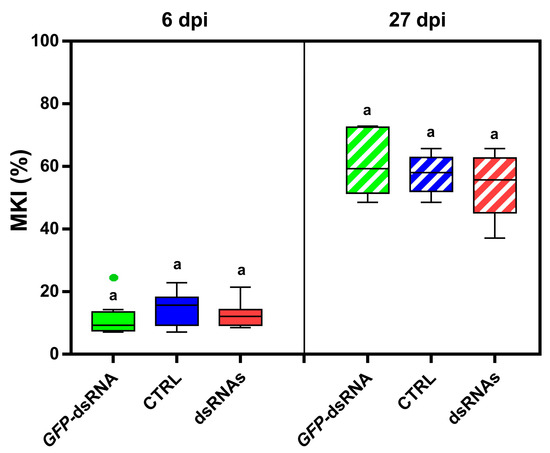
Figure 3.
The assessment of disease symptoms caused by B. cinerea and the relative effectiveness of dsRNA treatment are shown. The McKinney index (MKI) was used to evaluate disease symptoms after spray applications of a dsRNA mix (dsRNAs) including all the three in vitro-produced BcBmp1-, BcBmp3-, and BcPls1-dsRNAs (10 µg each, 30 µg total dsRNA per plant) and controls (CTRL and GFP-dsRNA) at 6 dpi and 27 dpi. Graphs and box plots show data from nine biological replicates (n = 9) of the experiment. The circles on box plots indicate 5th and 95th percentile outliers. Same letters above the bars indicate no significant differences from each other (ANOVA) according to Tukey’s test (p ≤ 0.001).
3.2. Characterization of Small Layered Double Hydroxide (sLDH) Clay Nanosheets
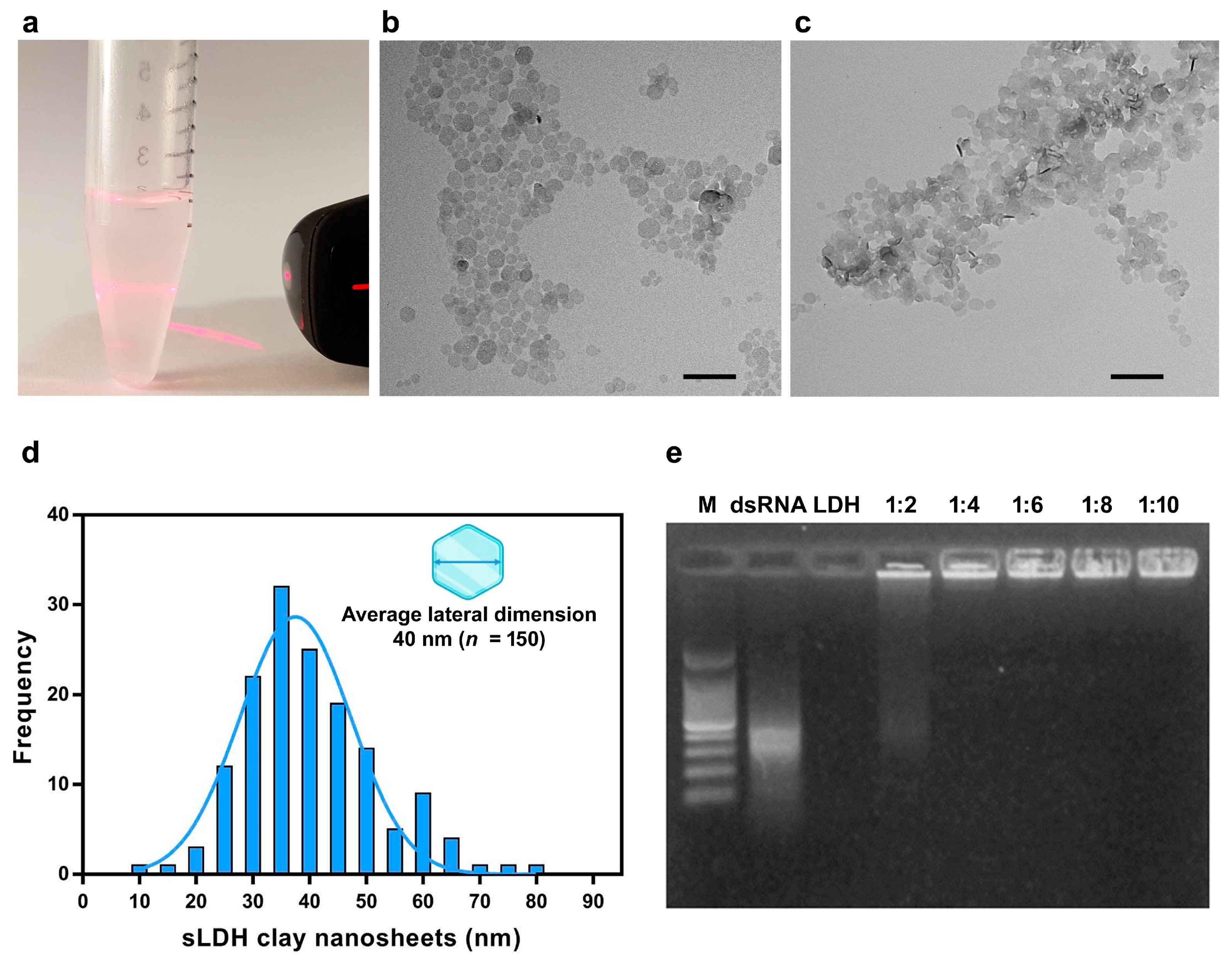
To prolong the stability of dsRNA, it was functionalized using sLDH clay nanosheets that we synthesized for its delivery using non-aqueous conditions. The sLDH clay nanosheets were dispersed in MilliQ water and the suspension visually appears very transparent and homogeneous. The suspension exhibits a Tyndall light-scattering effect, which indicates the presence of well-dispersed exfoliated nanosheets and confirms the colloidal nature of the dispersion (Figure 4a). The TEM analysis showed that the pristine sLDH nanoparticles had a hexagonal nanosheet morphology (Figure 4b). The addition of dsRNA to the suspension of sLDH clay nanosheets indeed caused evident aggregation of the nanoparticles (Figure 4c). The sLDH clay nanosheets showed a lateral size in the range of 10–80 nm with a monomodal size distribution and average lateral dimension of 40 nm (n = 150) (Figure 4d).
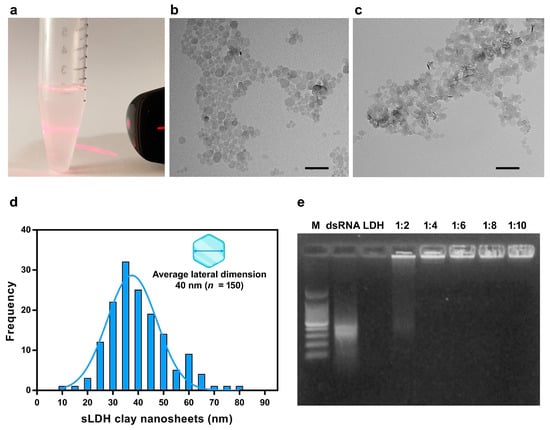
Figure 4.
Characterization of small layered double hydroxide (sLDH) clay nanosheets produced. (a) Tyndall light-scattering effect of sLDH clay nanosheet suspension upon irradiation with a laser beam (λ = 635 nm). (b) Transmission Electron Microscopy (TEM) analysis of the pristine sLDH clay nanosheets’ dispersion confirming their hexagonal morphology. (c) TEM analysis of sLDH clay nanosheet–dsRNA complex showing greater aggregation compared to the pristine sLDH clay nanosheets in (b). (d) Size distribution of sLDH clay nanosheets with an average lateral dimension of 40 nm (n = 150). (e) Retention effect of sLDH clay nanosheet–dsRNA complex in agarose gel where different ratios of sLDH clay nanosheets/dsRNA were evaluated. Scale bars = 200 nm. M = 100 bp DNA ladder.
The ability of sLDH clay nanosheets to complex and release dsRNA was evaluated by the gel retardation assay. In the absence of sLDH clay nanosheets, no complex formation was observed, and dsRNA migrated into the gel (Figure 4e, lane 2). The dsRNA, when complexed to sLDH clay nanosheets is unable to penetrate and migrate through the agarose pores. At ratios of 1:4 to 1:10, no detectable free dsRNA was visualized, indicating that fully complexed dsRNA with the sLDH clay nanosheets remained in the wells (Figure 4e, lanes 5–8). The selected ratio of sLDH clay nanosheets/dsRNA was 1:2 in order to have an aliquot of free naked dsRNA immediately available for gene knockdown (Figure 4e, lane 4).
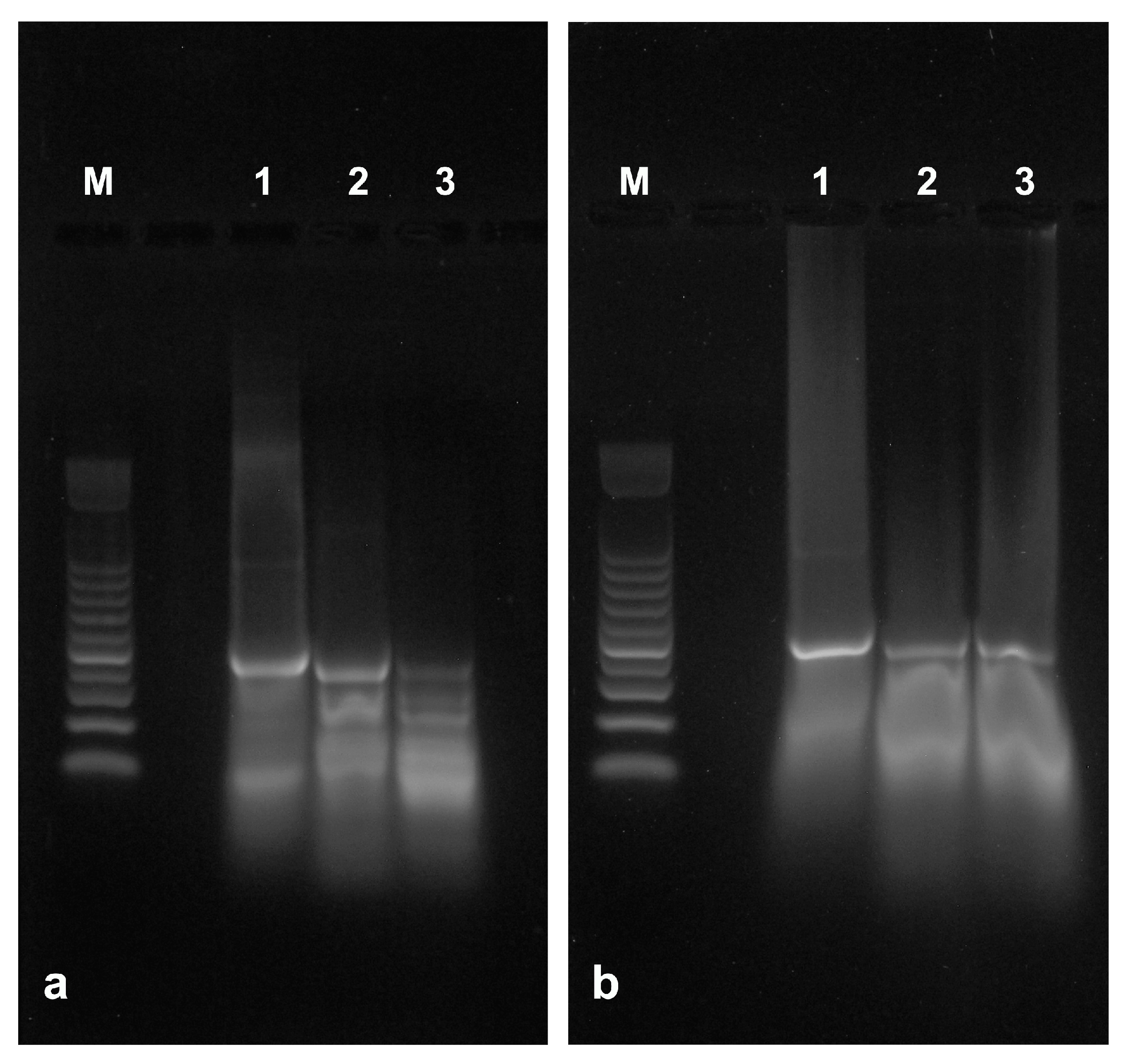
The protective ability of sLDH clay nanosheets was evaluated by exposing naked or complexed control dsRNA to RNase A. Samples contained 0.375 or 0.750 ng of RNase A which gradually degraded naked dsRNA (Figure 5a, lanes 2 and 3) compared to untreated naked dsRNA (Figure 5a, lane 1). The dsRNA complexed with the sLDH clay nanosheets, however, remained intact after acid release (Figure 5b, lanes 2 and 3) compared to untreated complexed dsRNA (Figure 5b, lane 1). This result clearly indicates that sLDH clay nanosheets protect dsRNA from nuclease degradation.
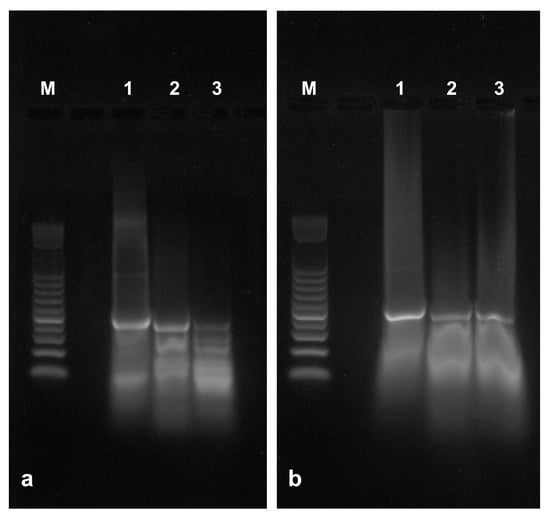
Figure 5.
The ability of sLDH clay nanosheets to protect dsRNA from RNase A degradation was evaluated using in vitro transcribed control dsRNA (500 bp). (a) Naked dsRNA untreated (lane 1) or treated with RNase A; samples (10 µL) contained 0.375 ng (lane 2) or 0.750 ng (lane 3) of enzyme. (b) dsRNA complexed with sLDH clay nanosheets (ratio 1:10) untreated (lane 1) or treated with RNase A; samples (10 µL) contained 0.375 ng (lane 2) or 0.750 ng (lane 3) of enzyme. dsRNA was released by acidic treatment from sLDH clay nanosheets immediately before agarose gel electrophoresis. M = 100 bp DNA ladder.
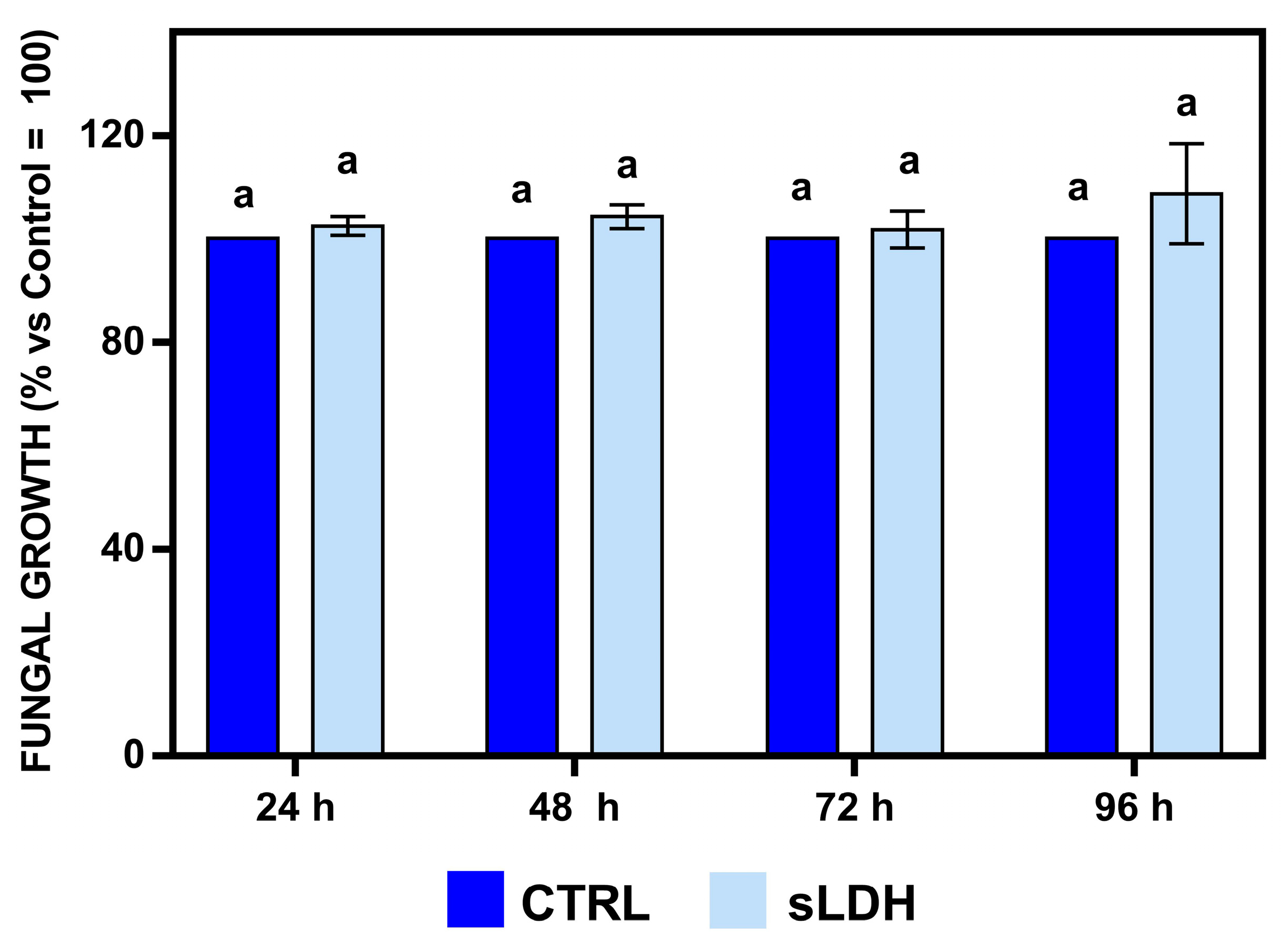
The in vitro-produced BcBmp1-, BcBmp3-, and BcPls1-dsRNAs were loaded to sLDH clay nanosheets and used to challenge B. cinerea in vivo during spray application experiments on whole lettuce plants. Before that, to explore the possible effects of sLDH clay nanosheets alone on fungal growth, an in vitro assay was performed, and the optical density of fungal mycelium at different times was measured. No effect of sLDH clay nanosheets was detected on fungal growth during this assay for any time tested (Figure 6).
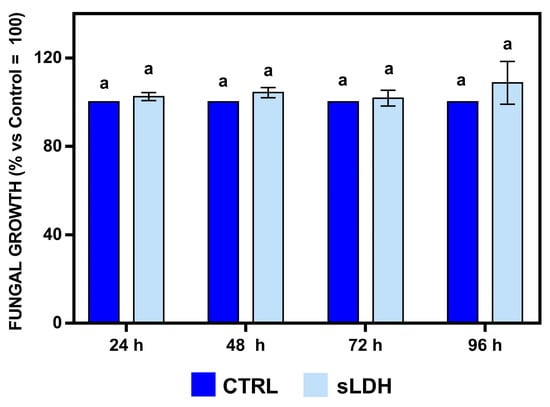
Figure 6.
In vitro effects of sLDH clay nanosheets on the growth of B. cinerea were evaluated by measuring the optical density (OD, 595 nm) at 24, 48, 72, and 96 h in 96-well microtiter plates. In each well, there were aliquots of a conidial suspension (5 × 102 spores) of B. cinerea in SMB medium and sLDH clay nanosheets (light blue) or TE buffer (CTRL, blue). The amount of sLDH clay nanosheets corresponded to the ratio 1:2 used in the sLDH clay nanosheet–dsRNA complexes. The graph shows the mean values (±SE) of two independent experiments with eight biological replicates (n = 8). Same letters above the bars indicate no significant differences from each other (ANOVA) according to Tukey’s HSD test (p ≤ 0.05). Statistical analysis was conducted separately for 24, 48, 72, and 96 h.
3.3. Spray-Induced Gene Silencing (SIGS): Application of sLDH Clay Nanosheet–dsRNA Complexes by Pressure Spraying Prolongs Protection Against Botrytis cinerea on Lettuce Plants
Even in the case of an in vivo spray application of sLDH clay nanosheet–dsRNA complexes, the efficacy of the dsRNA treatment was evaluated starting from an empirical qualitative ordinal scale, as described before (see Section 3.1). All data presented below are in percentage (%) form.
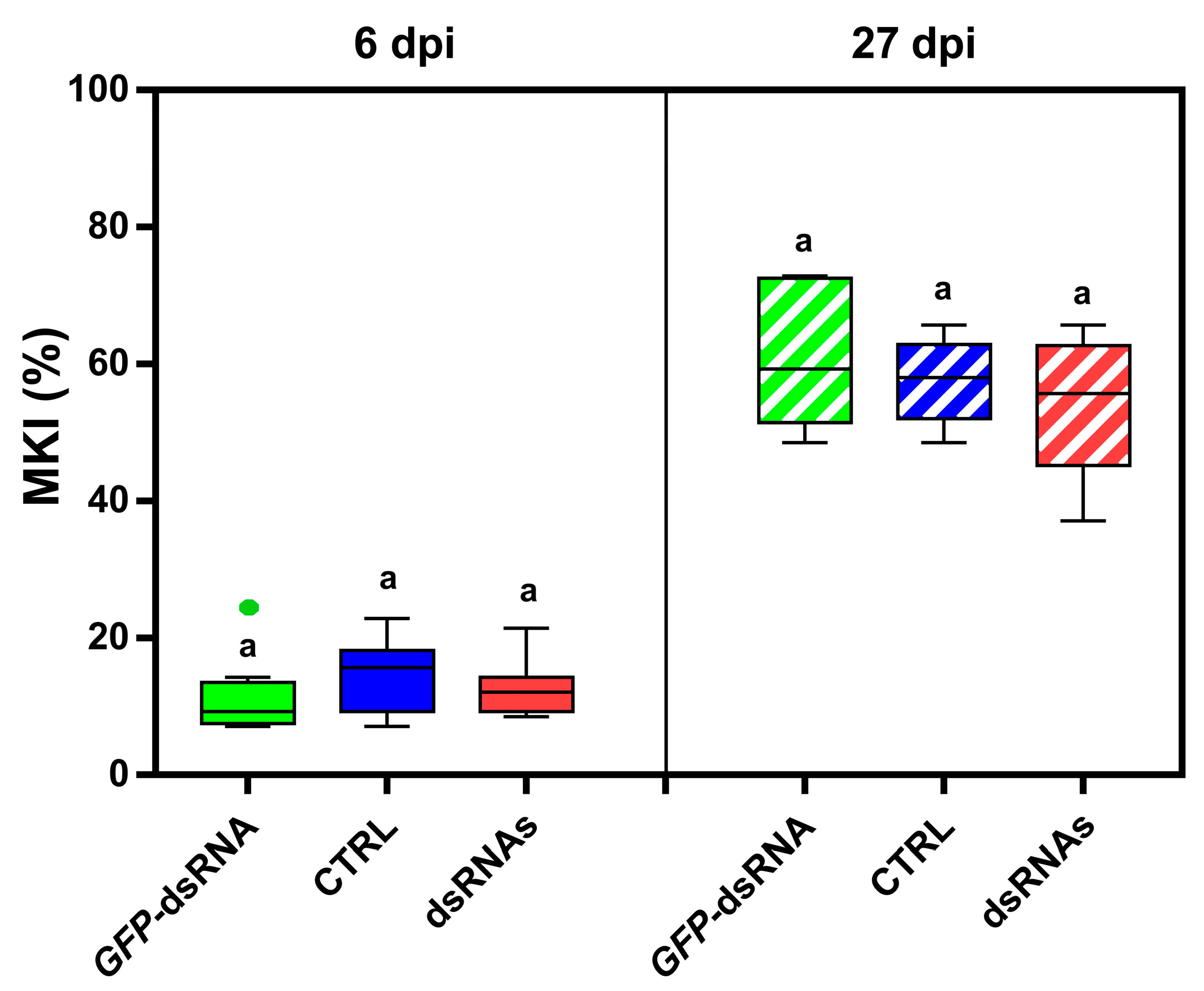
MKI analysis at 6 dpi showed that the sprayed treatments were all effective in reducing the disease symptoms caused by B. cinerea (Figure 7a). In particular, for BcBmp1-dsRNA/sLDH clay nanosheets the MKI was 19.6 ± 1.1, for BcBmp3-dsRNA/sLDH clay nanosheets the MKI was 11.2 ± 1.2, and in the case of BcPls1-dsRNA/sLDH clay nanosheets the MKI was 23.4 ± 3.2 compared to controls (MKI CTRL = 48.9 ± 5.2, MKIGFP-dsRNA = 51.8 ± 5.3 and MKIsLDH = 47.3 ± 5.1) (Figure 7a).
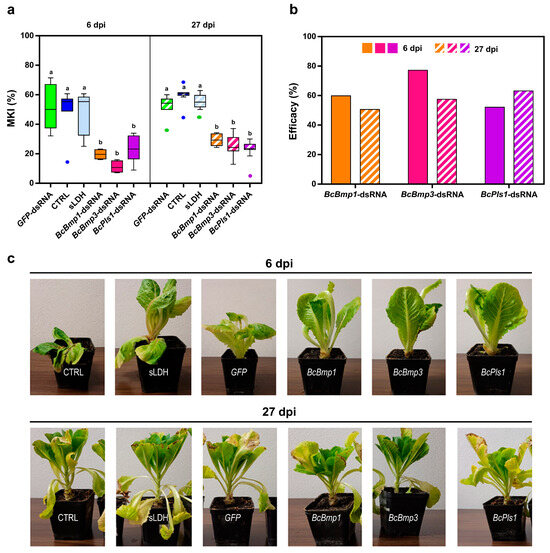
Figure 7.
The assessment of disease symptoms caused by B. cinerea and the relative effectiveness of sLDH clay nanosheet–dsRNA complexes treatment are shown. (a) The McKinney index (MKI) was used to evaluate disease symptoms after spray applications of in vitro-produced dsRNA complexed to sLDH clay nanosheets BcBmp1-dsRNA/sLDH, BcBmp3-dsRNA/sLDH, and BcPls1-dsRNA/sLDH complexes (BcBmp1, BcBmp3, and BcPls1) (10 µg per plant) and controls (CTRL, sLDH and GFP-dsRNA) at 6 dpi and 27 dpi. Graphs and box plots show data from sixteen biological replicates (n = 16) of the experiment. The circles on box plots indicate 5th and 95th percentile outliers. Same letters above the bars indicate no significant differences from each other (ANOVA) according to Tukey’s test (p ≤ 0.001). (b) The efficacy of spray applications of in vitro-produced dsRNA complexed to sLDH clay nanosheets BcBmp1-dsRNA/sLDH, BcBmp3-dsRNA/sLDH, and BcPls1-dsRNA/sLDH complexes (BcBmp1, BcBmp3, and BcPls1) was assessed at 6 dpi and 27 dpi by calculating the Abbott Index (AI) from the McKinney index (MKI). (c) Representative images of potted lettuce plants showing symptoms caused by B. cinerea when sampled at 6 dpi and 27 dpi. Symptoms of gray mold disease were analyzed in controls [CTRL, sLDH, and GFP-dsRNA (GFP)] and in lettuce plants treated with BcBmp1-dsRNA/sLDH, BcBmp3-dsRNA/sLDH, and BcPls1-dsRNA/sLDH complexes (BcBmp1, BcBmp3, and BcPls1).
At 6 dpi, in all three spray treatments, disease symptoms were reduced by half, or even more than half, compared to controls when lettuce plants were treated with sLDH clay nanosheet–dsRNA complexes (Figure 7b,c).
At 27 dpi, MKI analysis showed that all the sprayed dsRNA/LDH complexes were still effective in reducing disease symptoms caused by B. cinerea. In particular, for BcBmp1-dsRNA the MKI was 29.4 ± 1.4, for BcBmp3-dsRNA the MKI was 25.3 ± 2.6, and in the case of BcPls1-dsRNA the MKI was 21.9 ± 2.7 compared to controls (MKICTRL = 59.5 ± 2.4, MKIGFP-dsRNA = 52.1 ± 2.6, and MKIsLDH = 55.7 ± 1.6) (Figure 7a).
The efficacy results (Abbott index) of SIGS treatments using naked dsRNAs or complexed to sLDH clay nanosheets at both 6 dpi and 27 dpi were compared statistically (ANOVA) (Table 1). At 6 dpi, only the treatment with BcBmp3-dsRNA complexed to sLDH clay nanosheets was more effective than naked dsRNA. However, at 27 dpi all treatments with dsRNA (Bmp1-, Bmp3-, and Pls1-dsRNAs) complexed to sLDH clay nanosheets were more effective than naked dsRNA.

Table 1.
Comparison of the efficacy (AI %) of dsRNA treatments against B. cinerea target genes at 6 dpi and 27 dpi using naked dsRNA or dsRNA complexed to sLDH clay nanosheets. Asterisks indicate significant differences from each other (ANOVA) according to Tukey’s test (* p ≤ 0.05; *** p ≤ 0.001). Standard errors are in parenthesis. Statistical analysis was conducted separately for each gene.
3.4. Effectiveness of Fungicide Treatment by Pressure Spraying Against Botrytis cinerea on Lettuce Plants
The efficacy of the fungicide based on cyprodinil + fludioxonil (Switch® 62.5 WG) was evaluated using the same methodology (single treatment) used for the application of dsRNAs. In this case, its effectiveness was assessed only at 6 dpi as it is known that at 7 days in lettuce the residues of the two active ingredients are very low or not detected [46].
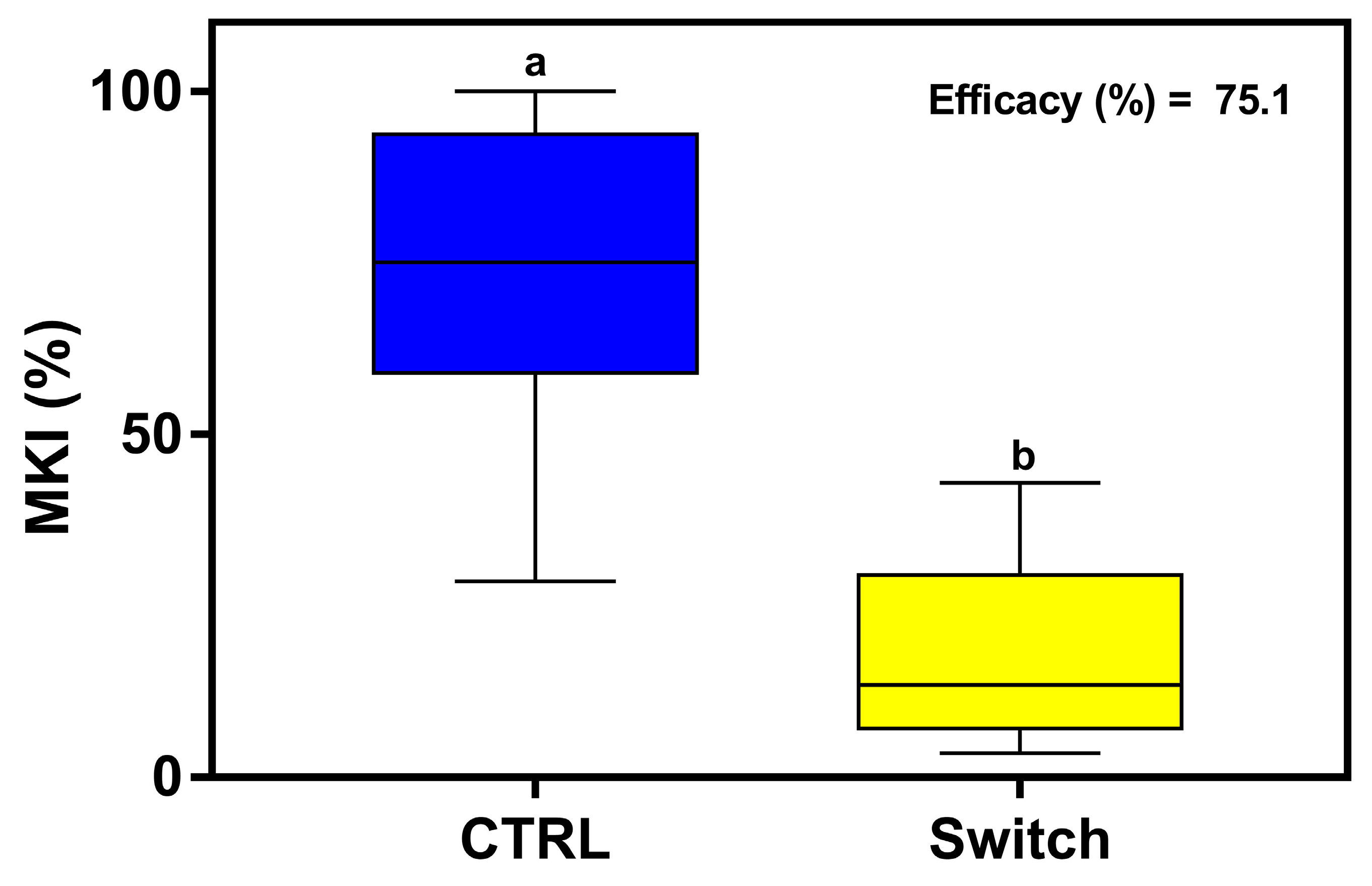
MKI analysis at 6 dpi showed that the sprayed fungicide was very effective in reducing disease symptoms caused by B. cinerea, with an AI of 75.1% (Figure 8).
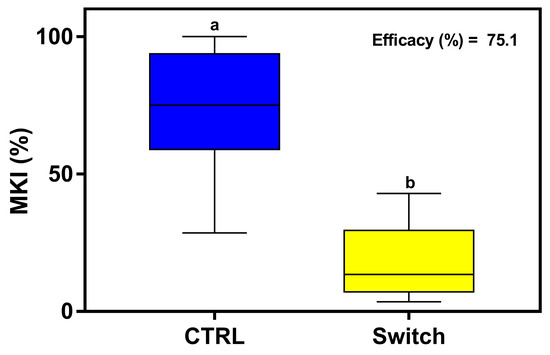
Figure 8.
The assessment of disease symptoms caused by B. cinerea and the relative effectiveness of fungicide treatment are shown. The McKinney index (MKI) was used to evaluate disease symptoms after spray applications of the fungicide based on cyprodinil + fludioxonil (Switch® 62.5 WG) at the recommended dose (0.7 g/L) for lettuce. Graphs and box plots show data from eight biological replicates (n = 8) of the experiment. Different letters above the bars indicate significant differences from each other (ANOVA) according to Tukey’s test (p ≤ 0.001). The efficacy of the fungicide treatment, evaluated at 6 dpi by calculating the Abbott Index (AI) from the McKinney index (MKI), is shown in the graph.
4. Discussion
Topical application of BcBmp1-, BcBmp3-, and BcPls1-dsRNAs was shown to trigger both in vitro and in vivo knockdown of their related genes, with a reduction in the disease symptoms on lettuce leaves as assessed by a detached leaf assay [38,39].
In this research, the efficacy of BcBmp1, BcBmp3, and BcPls1 genes as novel targets for SIGS against B. cinerea was demonstrated on whole lettuce plants. Indeed, with a view to developing an effective large-scale treatment, such as for use in greenhouses or fields, it was essential to evaluate the effectiveness of the treatments directly on plants and not just on detached organs.
Driven by this goal, it seemed crucial to consider not only the feasibility of the spray delivery technique but also to focus attention on the properties of dsRNA treatment. In particular, the improvement in the stability of the previously selected dsRNAs was evaluated.
In recent years, the use of topical application of dsRNA has been investigated for its applications in plant protection. In particular, the dsRNA spraying technique has proven to be a valid delivery method, as demonstrated in several studies concerning RNAi-based crop protection strategies [12,13,16,21,47,48].
In Fusarium asiaticum, dsRNA was successfully used to reduce resistance to the fungicide phenamacril by spraying 0.1 pM of Myo5-dsRNA plus 10 ng/L phenamacril on wheat coleoptiles, resulting in reduced growth of F. asiaticum along with an enhanced sensibility to the fungicide [47]. Spray experiments were also conducted on wounded leaves and whole rice plants using 300 nM of dsRNA targeting MoDES1, an innate defense suppressor of Magnaporthe oryzae leading to impaired biotrophic growth and virulence, resulting in an outstanding resistance against the blast disease [48]. In a study on Fusarium graminearum, detached barley leaves were locally sprayed with 20 ng/μL of in vitro-synthesized dsRNAs targeting the CYP51A, CYP51B, and CYP51C genes, which led to impaired development of the pathogen on barley, with a reduction in disease symptoms at the site of inoculation [12]. Similarly, bell peppers, cherries, mangoes, and grapes were sprayed with 15 ng/μL of dsRNA targeting three transcripts involved in B. cinerea ergosterol biosynthesis. The dsRNA treatment has been shown to decrease the development of fruit decay caused by B. cinerea [21].
As demonstrated in previous research, also in this study, the spray technology proved to be efficient for dsRNA delivery, leading to the control of B. cinerea. In vitro-synthesized BcBmp1-, BcBmp3-, and BcPls1-dsRNAs were sprayed onto lettuce plants, lately inoculated with B. cinerea. All sprayed dsRNAs were effective in reducing disease symptoms, with a remarkable reduction compared to controls, confirming the potential of this technology as alternative tool in gray mold disease management. In particular, dsRNA was used at the concentration of 20 ng/μL, which seemed an optimal dose of dsRNA to use for activating the RNAi mechanism of the target pathogen, as also seen in previous studies [12,49].
Conversely, when in vitro-produced BcBmp1-, BcBmp3-, and BcPls1-dsRNAs were sprayed together, the efficacy of dsRNA treatment in protecting lettuce from B. cinerea was lost, and this could be explained by an excessive amount of dsRNA used (30 µg per plant, 10 µg for each dsRNA). This is probably due to competitive inhibition of RNAi which had no effects in reducing disease symptoms. In fact, as also demonstrated in other studies, the dose of dsRNA used is crucial for the efficacy of the treatment since a high dose can be counterproductive because it could saturate the RNAi silencing machinery [49,50,51]. Moreover, when a mixture of dsRNA is used, it can lead to a competition between the different dsRNAs for components of the RNAi machinery resulting in a reduction in RNAi effect due to inhibition of the process [52,53].
The dose of dsRNA to be applied in SIGS experiments can depend on many factors including the target gene, the fungal pathogen, the type of plant and/or fruit, the phenological phase of the plant, and the dsRNA produced in vitro or in vivo. For example, in SIGS experiments against B. cinerea, dsRNA concentrations ranged from 15 to 300 ng/μL [20,36,37].
Recently, the possibility of improving the dsRNA stability over time by loading it to nanoparticles (or in nanovesicles) has been demonstrated [16,27,29,36,37,54,55].
In a SIGS study on Phytophthora infestans, dsRNA targeting different genes of the pathogen (SDH, EF−1α, GPI-HAM344, PLD-3, and HSP-90) reduced its growth, resulting in a decrease in disease symptoms. The treatment was applied by a high-pressure topical spray and Kaolinite nano clay particles to enhance delivery [54].
Nanoparticle-mediated dsRNA delivery was also successfully evaluated to protect rice from Rhizoctonia solani, where six different nanocarriers were used: chitosan, polyethyleneimine, protamine, polyamidoamine, carbon quantum dot, and chitosan/SPc (star polymer complex). In particular, the last two showed the best ability to load dsRNA, improving its stability and uptake by the target fungal pathogen, prolonging plant protection over time, up to 20 days [56].
Mitter et al. [29] showed that dsRNA complexed to layered double hydroxide (LDH) clay nanosheets conferred prolonged plant protection against viruses up to 20 dpi after spraying, while naked dsRNA protected plants for up to 5 days.
These types of highly efficient nanocarriers have a lamellar structure with a well-distributed array of positive charges that allows them to deliver negatively charged biomolecules such as dsRNA. When biomolecules are intercalated between LDH layers, they are protected from degradation. In addition to this important physical property, LDHs exhibit low cytotoxicity, pH sensitivity, gradual biodegradability under atmospheric CO2 and moisture conditions, and intracellular delivery efficacy, especially when their size is ≤50 nm [29,43,57].
In our experiments, sLDH clay nanosheets typically possessed a narrow particle size distribution with an average lateral dimension of 40 nm. The heat treatment was skipped by going straight to purification and dispersion of sLDH clay nanosheets in water because it had limited impact on size. Washing the sLDH slurry with methanol and water are the key steps that significantly affect the particle size and purity of the final product [43].
LDH clay nanosheets strongly adhere to the leaf surface through hydrogen bonds and electrostatic attraction and enhance the stability of dsRNA. They protect dsRNA from external degradation and promote the slow release of dsRNA over time [29,37,58].
The sLDH clay nanosheet–dsRNA complexes produced in this work proved resistant to RNase A treatment, thus showing the ability to protect dsRNA from environmental degradation in agreement with the results of other works [29,37].
In this study, in vitro-synthesized BcBmp1-, BcBmp3-, and BcPls1-dsRNAs were loaded to sLDH clay nanosheets and sprayed at a final concentration of 20 ng/μL on lettuce plants lately inoculated with B. cinerea. All sprayed sLDH clay nanosheet–dsRNA complexes showed better plant protection over time compared to the use of naked dsRNA. This was particularly evident at 27 dpi, where the use of sLDH clay nanosheet–dsRNA complexes achieved an efficiency of approximately 50% or even higher, in agreement with recent research studies where the dsRNA/LDH complex was also used for the RNAi-based control of B. cinerea. According to these studies, LDH clay nanosheets prolonged the efficiency of dsRNA treatment for six weeks on fruits [37] and up to 3–4 weeks on tomato leaves and chickpea plants [36]. On the contrary, the same naked dsRNA showed a reduced ability to control gray mold disease over time [36,37].
The fungicide Switch® (a mixture of the two active ingredients cyprodinil and fludioxonil) is considered a robust marketed solution for the control of B. cinerea, but no more than three applications per season with intervals of 10–12 days are recommended. Fungicide application reduced disease incidence and severity at 6 dpi compared to untreated plants, with an efficacy of 75.1%. Similar results were observed by other authors with the efficacy (AI %) of the applied fungicide ranging from 70.9 to 86.5% [59].
If we consider the dsRNA-based treatments carried out in this work at 6 dpi, they have an efficacy between 50.5 and 52.1% for naked dsRNAs and an efficacy between 59.8 and 77.2% for the dsRNAs complexed to the sLDH clay nanosheets. The effectiveness of SIGS using BcBmp1-, BcBmp3-, and BcPls1-dsRNAs in reducing disease caused by B. cinerea is high overall and increases significantly with the use of sLDH clay nanosheets.
Therefore, in our opinion, it should be noted that the efficacy of dsRNA treatments obtained in our experiments, especially when sLDH clay nanosheet–dsRNA complexes were used, turned out to be not so far from that achieved with the more traditional use of fungicides.
According to Duanis-Assaf et al. [37], LDHs themselves produce a physical barrier that protects fruit from fungal decay. Furthermore, it was remarked that the ready availability of free dsRNA is crucial in the early phase of B. cinerea infection since it enhances protection against the pathogen compared to dsRNA that is not immediately available when it is still linked in the dsRNA/LDH complex. Indeed, when B. cinerea infection was conducted at 7 days after treatment, naked dsRNA showed better results than the LDH-dsRNA complex [29,37]. This is in agreement with our results; in fact, we used a 1:2 ratio between dsRNA and sLDH clay nanosheets to have a quantity of dsRNA immediately available, as also suggested by Mitter et al. [29]. In our case, at 6 dpi, the efficiency of dsRNA/LDH treatment is lower than at 27 dpi, probably due to a smaller amount of free dsRNA. Conversely, at 27 dpi, the dsRNA released over time allowed a prolonged protection of lettuce plants.
Another important aspect considered in our study is that the dsRNA spray treatments were applied directly on whole host plants to be as close as possible to real natural conditions found in the field or greenhouse. Our work therefore contributes to adding information on the effectiveness of SIGS in the control of B. cinerea on whole plants, joining the few experiments conducted in this regard [16,20,36].
In a study on the early stages of B. cinerea–tomato interaction, Qin et al. [60] reported that SIGS offers an interesting perspective for future plant protection strategies. In any case, the success of this method does not necessarily imply natural cross-kingdom RNA interference, i.e., sRNAs trafficking between the host and the pathogen that promotes silencing, during the interaction between B. cinerea and its host [13,61].
To date, studies using naked dsRNA or that complexed with nanomaterials to reduce the impact of phytopathogenic fungi have not yet been sufficiently validated, and therefore our results should be considered with the aim of contributing to filling this gap.
5. Conclusions
RNAi-based biocontrol strategies could be an excellent alternative to chemical fungicide since they are species-specific, low-toxicity, and environmentally friendly, and several RNAi fungicides are expected to be approved in the near future. However, RNAi applications must face several challenges that need to be overcome before reaching the market [62].
First, when designing dsRNA, the selection of key genes of the target pathogen is a crucial step for the effectiveness of the treatment. Contextually, dsRNAs should be rigorously designed, through the help of in silico analysis, to avoid non-target effects in non-target organisms, including the host plant. The physiological, nutritional, and organoleptic characteristics of the crop must remain intact following the treatment. Moreover, degradation of dsRNA can occur due to the action of ribonucleases and/or UV radiation present in the environment. To overcome this problem, new nanomaterials or nanovesicles are coming to the forefront that extend the duration of dsRNA treatments and sometimes improve dsRNA delivery. Considering large-scale application, reducing production costs is a priority. For this reason, some engineered bacteria can be used as a biofactory and help to increase the amount of dsRNA produced with a huge reduction in costs compared to in vitro synthesis. In the end, it is critical to find and establish a consensual regulatory framework for the commercialization of RNAi fungicides, based on risk assessment, as with any new technology. We must also ask how RNAi technology is still relevant in terms of the exogenous application of dsRNA in this new era of CRISPR/Cas [63]. Indeed, it is necessary to consider how RNAi and genome editing technologies can intersect to be used jointly in improving plant disease resistance against insects, fungi, and nematodes.
Author Contributions
Conceptualization, M.S., C.P., M.F. and S.P.; methodology, M.S, C.P., M.F. and S.P.; investigation, M.S., C.P., M.F., D.P., A.C. and S.P.; writing—original draft preparation, M.S. and C.P.; writing—review and editing, C.P., M.F. and S.P.; supervision, C.P., M.F. and S.P.; project administration, C.P., M.F. and S.P.; funding acquisition, M.S., C.P., M.F. and S.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was partially supported by the University of Pisa within the framework of a PhD research project entitled “Botrytis cinerea-Lactuca sativa L. pathosystem: nanoparticles-mediated delivery of dsRNA in Spray-Induced Gene Silencing (SIGS) for plant disease control”.
Data Availability Statement
All data generated or analyzed during this study are included in this publication.
Acknowledgments
We acknowledge Grazia Puntoni (University of Pisa, Italy) for the technical and administrative support. We extend our special thanks to Federico Tasca and Michele Persico (University of Pisa, Italy) for their valuable assistance during the evaluation of the stability of dsRNA complexed to sLDH clay nanosheets.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Williamson, B.; Tudzynski, B.; Tudzynski, P.; Van Kan, J.A. Botrytis cinerea: The cause of grey mould disease. Mol. Plant Pathol. 2007, 8, 561–580. [Google Scholar] [CrossRef]
- Dean, R.; Van Kan, J.A.L.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef] [PubMed]
- Romanazzi, G.; Feliziani, E. Botrytis cinerea. In Postharvest Decay: Control Strategies; Bautista-Baños, S., Ed.; Elsevier: London, UK, 2014; pp. 131–146. [Google Scholar]
- Fillinger, S.; Walker, A.S. Chemical control and resistance management of Botrytis diseases. In Botrytis—The Fungus, the Pathogen and Its Management in Agricultural Systems; Fillinger, S., Elad, Y., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 189–216. [Google Scholar]
- Sofianos, G.; Samaras, A.; Karaoglanidis, G. Multiple and multidrug resistance in Botrytis cinerea: Molecular mechanisms of MLR/MDR strains in Greece and effects of co-existence of different resistance mechanisms on fungicide sensitivity. Front. Plant Sci. 2023, 14, 1273193. [Google Scholar] [CrossRef]
- Medina-Lozano, I.; Bertolín, J.R.; Díaz, A. Nutritional value of commercial and traditional lettuce (Lactuca sativa L.) and wild relatives: Vitamin C and anthocyanin content. Food Chem. 2021, 359, 129864. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Su, W.; Tao, R.; Zhang, W.; Chen, J.; Wu, P.; Yan, C.; Jia, Y.; Larkin, R.M.; Lavelle, D.; et al. RNA sequencing provides insights into the evolution of lettuce and the regulation of flavonoid biosynthesis. Nat. Commun. 2017, 8, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Raid, R.N. Lettuce diseases and their management. In Diseases of Fruits and Vegetables; Naqvi, S.A.M.H., Ed.; Kluwer Academic: Dordrecht, The Netherlands, 2004; Volume 2, pp. 121–148. [Google Scholar] [CrossRef]
- Horská, T.; Kocourek, F.; Stará, J.; Holý, K.; Mráz, P.; Krátký, F.; Kocourek, V.; Hajšlová, J. Evaluation of pesticide residue dynamics in lettuce, onion, leek, carrot and parsley. Foods 2020, 9, 680. [Google Scholar] [CrossRef]
- Shim, C.K.; Kim, M.J.; Kim, Y.K.; Jee, H.J. Evaluation of lettuce germplasm resistance to gray mold disease for organic cultivations. Plant Pathol. J. 2014, 30, 90–95. [Google Scholar] [CrossRef]
- Chakraborty, S.; Newton, A.C. Climate change, plant diseases and food security: An overview. Plant Pathol. 2011, 60, 2–14. [Google Scholar] [CrossRef]
- Koch, A.; Biedenkopf, D.; Furch, A.; Weber, L.; Rossbach, O.; Abdellatef, E.; Linicus, L.; Johannsmeler, J.; Jelonek, L.; Goesmann, A.; et al. An RNAi-based control of Fusarium graminearum infections through spraying of long dsRNAs involves a plant passage and is controlled by the fungal silencing machinery. PLoS Pathog. 2016, 12, e1005901. [Google Scholar] [CrossRef]
- Wang, M.; Weiberg, A.; Lin, F.-M.; Thomma, B.P.H.J.; Huang, H.-D.; Jin, H. Bidirectional cross-kingdom RNAi and fungal uptake of external RNAs confer plant protection. Nat. Plants 2016, 2, 1651. [Google Scholar] [CrossRef] [PubMed]
- Mezzetti, B.; Smagghe, G.; Arpaia, S.; Christiaens, O.; Dietz-Pfeilstetter, A.; Jones, H.; Kostov, K.; Sabbadini, S.; Opsahl-Sorteberg, H.G.; Ventura, V.; et al. RNAi: What is its position in agriculture? J. Pest Sci. 2020, 93, 1125–1130. [Google Scholar] [CrossRef]
- Rodrigues, T.B.; Petrick, J.S. Safety considerations for humans and other vertebrates regarding agricultural uses of externally applied RNA molecules. Front. Plant Sci. 2020, 11, 407. [Google Scholar] [CrossRef]
- Spada, M.; Pugliesi, C.; Fambrini, M.; Pecchia, S. Challenges and opportunities arising from host-Botrytis cinerea interactions to outline novel and sustainable control strategies: The key role of RNA interference. Int. J. Mol. Sci. 2024, 25, 6798. [Google Scholar] [CrossRef]
- Islam, M.T.; Sherif, S.M. RNAi-based biofungicides as a promising next-generation strategy for controlling devastating gray mold diseases. Int. J. Mol. Sci. 2020, 21, 2072. [Google Scholar] [CrossRef]
- Fletcher, S.J.; Reeves, P.T.; Hoang, B.T.; Mitter, N. A perspective on RNAi-based biopesticides. Front. Plant Sci. 2020, 11, 51. [Google Scholar] [CrossRef]
- Dalakouras, A.; Jarausch, W.; Buchholz, G.; Bassler, A.; Braun, M.; Manthey, T.; Krczal, G.; Wassenegger, M. Delivery of hairpin RNAs and small RNAs into woody and herbaceous plants by trunk injection and petiole absorption. Front. Plant Sci. 2018, 9, 1253. [Google Scholar] [CrossRef]
- Nerva, L.; Sandrini, M.; Gambino, G.; Chitarra, W. Double-stranded RNAs (dsRNAs) as a sustainable tool against gray mold (Botrytis cinerea) in grapevine: Effectiveness of different application methods in an open-air environment. Biomolecules 2020, 10, 200. [Google Scholar] [CrossRef] [PubMed]
- Duanis-Assaf, D.; Galsurker, O.; Davydov, O.; Maurer, D.; Feygenberg, O.; Sagi, M.; Poverenov, E.; Fluhr, R.; Alkan, N. Double-stranded RNA targeting fungal ergosterol biosynthesis pathway controls Botrytis cinerea and postharvest grey mould. Plant Biotechnol. J. 2022, 20, 226–237. [Google Scholar] [CrossRef]
- Hernández-Soto, A.; Chacón-Cerdas, R. RNAi crop protection advances. Int. J. Mol. Sci. 2021, 22, 12148. [Google Scholar] [CrossRef]
- Álvarez-Sánchez, A.R.; Romo-Quinones, C.; Rosas-Quijano, R.; Reyes, A.G.; Barraza, A.; Magallón-Barajas, F.; Angulo, C.; Mejía-Ruíz, C.H. Production of specific dsRNA against white spot syndrome virus in the yeast Yarrowia lipolytica. Aquacult. Res. 2018, 49, 480–491. [Google Scholar] [CrossRef]
- Taning, C.N.T.; Arpaia, S.; Christiaens, O.; Dietz-Pfeilstetter, A.; Jones, H.; Mezzetti, B.; Sabbadini, S.; Sorteberg, H.-G.; Sweet, J.; Ventura, V.; et al. RNA-based biocontrol compounds: Current status and perspectives to reach the market. Pest Manag. Sci. 2020, 76, 841–845. [Google Scholar] [CrossRef]
- Delgado-Martín, J.; Velasco, L. An efficient dsRNA constitutive expression system in Escherichia coli. Appl. Microbiol. Biotechnol. 2021, 105, 6381–6393. [Google Scholar] [CrossRef]
- Verdonckt, T.-W.; Vanden Broeck, J. Methods for the cost-effective production of bacteria-derived double-stranded RNA for in vitro knockdown studies. Front. Physiol. 2022, 13, 836106. [Google Scholar] [CrossRef] [PubMed]
- Scarpin, D.; Nerva, L.; Chitarra, W.; Moffa, L.; D’Este, F.; Vuerich, M.; Filippi, A.; Braidot, E.; Petrussa, E. Characterisation and functionalisation of chitosan nanoparticles as carriers for double-stranded RNA (dsRNA) molecules towards sustainable crop protection. Biosci. Rep. 2023, 43, BSR20230817. [Google Scholar] [CrossRef]
- Niño-Sánchez, J.; Chen, L.H.; de Souza, J.T.; Mosquera, S.; Stergiopoulos, I. Targeted delivery of gene silencing in fungi using genetically engineered bacteria. J. Fungi 2021, 7, 125. [Google Scholar] [CrossRef] [PubMed]
- Mitter, N.; Worrall, E.A.; Robinson, K.E.; Li, P.; Jain, R.G.; Taochy, C.; Fletcher, S.J.; Carroll, B.J.; Lu, G.; Xu, Z.P. Clay nanosheets for topical delivery of RNAi for sustained protection against plant viruses. Nat. Plants 2017, 3, 16207. [Google Scholar] [CrossRef] [PubMed]
- Demirer, G.S.; Zhang, H.; Goh, N.S.; Pinals, R.L.; Chang, R.; Landry, M.P. Carbon nanocarriers deliver siRNA to intact plant cells for efficient gene knockdown. Sci. Adv. 2020, 6, eaaz0495. [Google Scholar] [CrossRef]
- Schwartz, S.H.; Hendrix, B.; Hoffer, P.; Sanders, R.A.; Zheng, W. Carbon dots for efficient small interfering RNA delivery and gene silencing in plants. Plant Physiol. 2020, 184, 647–657. [Google Scholar] [CrossRef]
- Chen, A.; Halilovi, L.; Shay, J.-H.; Koch, A.; Mitter, N.; Jin, H. Improving RNA-based crop protection through nanotechnology and insights from cross-kingdom RNA trafficking. Curr. Opin. Plant Biol. 2023, 76, 102441. [Google Scholar] [CrossRef]
- Mann, C.W.G.; Sawyer, A.; Gardiner, D.M.; Mitter, N.; Carroll, B.J.; Eamens, A.L. RNA-based control of fungal pathogens in plants. Int. J. Mol. Sci. 2023, 24, 12391. [Google Scholar] [CrossRef]
- Christiaens, O.; Tardajos, M.G.; Martinez Reyna, Z.L.; Dash, M.; Dubruel, P.; Smagghe, G. Increased RNAi efficacy in Spodoptera exigua via the formulation of dsRNA with guanylated polymers. Front. Physiol. 2018, 9, 316. [Google Scholar] [CrossRef]
- Worrall, E.A.; Bravo-Cazar, A.; Nilon, A.T.; Fletcher, S.J.; Robinson, K.E.; Carr, J.P.; Mitter, N. Exogenous application of RNAi-inducing double-stranded RNA inhibits aphid-mediated transmission of a plant virus. Front. Plant Sci. 2019, 10, 265. [Google Scholar] [CrossRef]
- Niño-Sánchez, J.; Sambasivam, P.T.; Sawyer, A.; Hamby, R.; Chen, A.; Czislowski, E.; Li, P.; Manzie, N.; Gardiner, D.M.; Ford, R.; et al. BioClay™ prolongs RNA interference-mediated crop protection against Botrytis cinerea. J. Integr. Plant Biol. 2022, 64, 2187–2198. [Google Scholar] [CrossRef]
- Duanis-Assaf, D.; Shlar, I.; Galsurker, O.; Davydov, O.; Maurer, D.; Feygenberg, O.; Poverenov, E.; Fluhr, R.; Alkan, N. Nano-clay, layered-double hydroxide (LDH), improves the efficacy of double-stranded RNA in controlling postharvest decay. Postharvest Biol. Technol. 2022, 193, 112051. [Google Scholar] [CrossRef]
- Spada, M.; Pugliesi, C.; Fambrini, M.; Pecchia, S. Silencing of the Slt2-type MAP kinase Bmp3 in Botrytis cinerea by application of exogenous dsRNA affects fungal growth and virulence on Lactuca sativa. Int. J. Mol. Sci. 2021, 22, 5362. [Google Scholar] [CrossRef]
- Spada, M.; Pugliesi, C.; Fambrini, M.; Palpacelli, D.; Pecchia, S. Knockdown of Bmp1 and Pls1 virulence genes by exogenous application of RNAi-inducing dsRNA in Botrytis cinerea. Int. J. Mol. Sci. 2023, 24, 4869. [Google Scholar] [CrossRef] [PubMed]
- Bock, C.H.; Pethybridge, S.J.; Barbedo, J.G.A.; Esker, P.D.; Mahlein, A.K.; Del Ponte, E.M. A phytopathometry glossary for the twenty-first century: Towards consistency and precision in intra- and inter-disciplinary dialogues. Trop. Plant Pathol. 2022, 47, 14–24. [Google Scholar] [CrossRef]
- Büttner, P.; Koch, F.; Voigt, K.; Quidde, T.; Risch, S.; Blaich, R.; Brückner, B.; Tudzynski, P. Variations in ploidy among isolates of Botrytis cinerea: Implications for genetic and molecular analyses. Curr. Genet. 1994, 25, 445–450. [Google Scholar] [CrossRef] [PubMed]
- Quidde, T.; Osbourn, A.E.; Tudzynski, P. Detoxification of alpha-tomatine by Botrytis cinerea. Physiol. Mol. Plant Pathol. 1998, 52, 151–165. [Google Scholar] [CrossRef]
- Dong, H.; Chen, M.; Rahman, S.; Parekh, H.S.; Cooper, H.M.; Xu, Z.P. Engineering small MgAl-layered double hydroxide nanoparticles for enhanced gene delivery. Appl. Clay Sci. 2014, 100, 66–75. [Google Scholar] [CrossRef]
- McKinney, H.H. Influence of soil temperature and moisture on infection of wheat seedlings by Helminthosporium sativum. J. Agric. Res. 1923, 26, 195–218. [Google Scholar]
- Abbott, W.S. A method of computing the effectiveness of an insecticide. J. Econom. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Marín, A.; Oliva, J.; Garcia, C.; Navarro, S.; Barba, A. Dissipation rates of cyprodinil and fludioxonil in lettuce and table grape in the field and under cold storage conditions. J. Agric. Food Chem. 2003, 51, 4708–4711. [Google Scholar] [CrossRef]
- Song, X.-S.; Gu, K.-X.; Duan, X.-X.; Xiao, X.-M.; Hou, Y.-P.; Duan, Y.-B.; Wang, J.-X.; Zhou, M.-G. A myosin5 dsRNA that reduces the fungicide resistance and pathogenicity of Fusarium asiaticum. Pestic. Biochem. Physiol. 2018, 150, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, A.; Roy-Barman, S. Spray-induced silencing of pathogenicity gene MoDES1 via exogenous double-stranded RNA can confer partial resistance against fungal blast in rice. Front. Plant Sci. 2021, 12, 733129. [Google Scholar] [CrossRef]
- McLoughlin, A.G.; Wytinck, N.; Walker, P.L.; Girard, I.J.; Rashid, K.Y.; de Kievit, T.; Fernando, W.G.D.; Whyard, S.; Belmonte, M.F. Identification and application of exogenous dsRNA confers plant protection against Sclerotinia sclerotiorum and Botrytis cinerea. Sci. Rep. 2018, 8, 7320. [Google Scholar] [CrossRef] [PubMed]
- Grimm, D.; Streetz, K.L.; Jopling, C.L.; Storm, T.A.; Pandey, K.; Davis, C.R.; Marion, P.; Salazar, F.; Kay, M.A. Fatality in mice due to oversaturation of cellular microRNA/short hairpin RNA pathways. Nature 2006, 441, 537–541. [Google Scholar] [CrossRef]
- Bensoussan, N.; Dixit, S.; Tabara, M.; Letwin, D.; Milojevic, M.; Antonacci, M.; Pengyu, J.P.; Arai, Y.; Bruinsma, K.; Suzuki, T.; et al. Environmental RNA interference in two-spotted spider mite, Tetranychus urticae, reveals dsRNA processing requirements for efficient RNAi response. Sci. Rep. 2020, 10, 19126. [Google Scholar] [CrossRef] [PubMed]
- Parrish, S.; Fleenor, J.; Xu, S.; Mello, C.; Fire, A. Functional anatomy of a dsRNA trigger: Differential requirement for the two trigger strands in RNA interference. Mol. Cell 2000, 6, 1077–1087. [Google Scholar] [CrossRef] [PubMed]
- Barik, S. RNAi in moderation. Nat. Biotechnol. 2006, 24, 796–797. [Google Scholar] [CrossRef]
- Sundaresha, S.; Sharma, S.; Bairwa, A.; Tomar, M.; Kumar, R.; Bhardwaj, V.; Jeevalatha, A.; Bakade, R.; Salaria, N.; Thakur, K.; et al. Spraying of dsRNA molecules derived from Phytophthora infestans, along with nanoclay carriers as a proof of concept for developing novel protection strategy for potato late blight. Pest Manag. Sci. 2022, 78, 3183–3192. [Google Scholar] [CrossRef]
- Qiao, L.; Niño-Sánchez, J.; Hamby, R.; Capriotti, L.; Chen, A.; Mezzetti, B.; Jin, H. Artificial nanovesicles for dsRNA delivery in spray-induced gene silencing for crop protection. Plant Biotech. J. 2023, 21, 854–865. [Google Scholar] [CrossRef]
- Wang, Y.; Yan, Q.; Lan, C.; Tang, T.; Wang, K.; Shen, J.; Niu, D. Nanoparticle carriers enhance RNA stability and uptake efficiency and prolong the protection against Rhizoctonia solani. Phytopathol. Res. 2023, 5, 2. [Google Scholar] [CrossRef]
- Chen, M.; Cooper, H.M.; Zhou, J.Z.; Bartlett, P.F.; Xu, Z.P. Reduction in the size of layered double hydroxide nanoparticles enhances the efficiency of siRNA delivery. J. Colloid Interface Sci. 2013, 390, 275–281. [Google Scholar] [CrossRef]
- Zhi, H.; Chen, H.; Yu, M.; Wang, C.; Cui, B.; Zhao, X.; Wang, Y.; Cui, H.; Zhang, B.; Zeng, Z. Layered double hydroxide nanosheets improve the adhesion of fungicides to leaves and the antifungal performance. ACS Appl. Nano Mater. 2022, 5, 5316–5325. [Google Scholar] [CrossRef]
- Vuković, S.; Brzaković, N.; Lazić, S.; Šunjka, D.; Žunić, A.; Bošković, D. Control of gray rot (Botrytis cinerea Pers.: Fr.) on lettuce. Biljni Lekar (Plant Doctor) 2019, 47, 147–156. [Google Scholar]
- Qin, S.; Veloso, J.; Baak, M.; Boogmans, B.; Bosman, T.; Puccetti, G.; Shi-Kunne, X.; Smit, S.; Grant-Downton, R.; Leisen, T.; et al. Molecular characterization reveals no functional evidence for naturally occurring cross-kingdom RNA interference in the early stages of Botrytis cinerea–tomato interaction. Mol. Plant Pathol. 2022, 24, 3–15. [Google Scholar] [CrossRef]
- Weiberg, A.; Wang, M.; Lin, F.M.; Zhao, H.; Zhang, Z.; Kaloshian, I.; Huang, H.D.; Jin, H. Fungal small RNAs suppress plant immunity by hijacking host RNA interference pathways. Science 2013, 342, 118–123. [Google Scholar] [CrossRef]
- De Schutter, K.; Taning, C.N.T.; Van Daele, L.; Van Damme, E.J.M.; Dubruel, P.; Smagghe, G. RNAi-based biocontrol products: Market status, regulatory aspects, and risk assessment. Front. Insect Sci. 2022, 1, 818037. [Google Scholar] [CrossRef] [PubMed]
- Halder, K.; Chaudhuri, A.; Abdin, M.Z.; Majee, M.; Datta, A. RNA interference for improving disease resistance in plants and its relevance in this clustered regularly interspaced short palindromic repeats-dominated era in terms of dsRNA-based biopesticides. Front. Plant Sci. 2022, 13, 885128. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).