
Synergistic Effects of Exogenous Lactobacillus plantarum and Fibrolytic Enzymes on Fermentation Quality, Fiber Degradation, and In Vitro Digestibility of Napiergrass (Pennisetum purpureum) Silage




Abstract
1. Introduction
2. Materials and Methods
2.1. Ensiling Material and Silage Making
2.2. Chemical Analysis
2.3. Measurement of IVDMD
2.4. Statistical Analyses
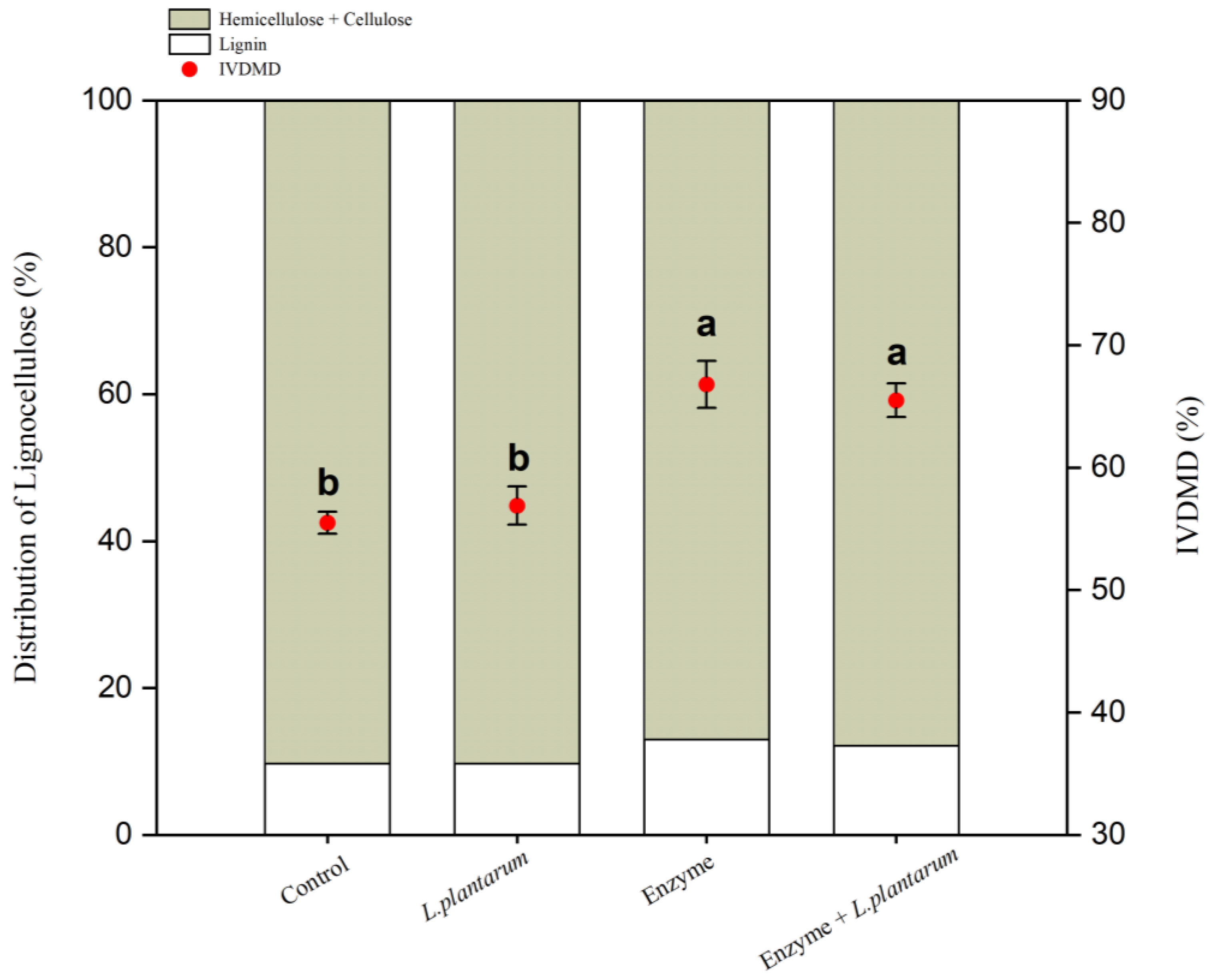
3. Results
3.1. Chemical and Microbial Composition of Napiergrass
3.2. Dynamics Changes in Fermentation Profiles and Structural Carbohydrates Compositions During Ensiling
3.3. Correlation of Fermentative Profiles and Parameters in Silages
4. Discussion
4.1. Fermentation Profile and Microbial Counts
4.2. Structural Carbohydrates Degradation and Digestibility
4.3. Correlation Between Treatments and Fermentative Parameters
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zailan, M.Z.; Yaakub, H.; Jusoh, S. Yield and nutritive quality of napier (Pennisetum purpureum) cultivars as fresh and ensiled fodder. J. Anim. Plant Sci. 2018, 28, 63–72. [Google Scholar]
- Jaipolsaen, N.; Sangsritavong, S.; Uengwetwanit, T.; Angthong, P.; Plengvidhya, V.; Rungrassamee, W.; Yammuenart, S. Comparison of the Effects of Microbial Inoculants on Fermentation Quality and Microbiota in Napier grass (Pennisetum purpureum) and Corn (Zea mays L.) Silage. Front. Microbiol. 2022, 12, 784535. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Dong, Z.H.; Li, J.F.; Chen, L.; Bai, Y.F.; Jia, Y.S.; Shao, T. Ensiling as pretreatment of rice straw: The effect of hemicellulase and Lactobacillus plantarum on hemicellulose degradation and cellulose conversion. Bioresour. Technol. 2018, 266, 158–165. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.R.; Guo, G.; Li, J.F.; Chen, L.; Dong, Z.H.; Shao, T. Improvement of fermentation profile and structural carbohydrate compositions in mixed silages ensiled with fibrolytic enzymes, molasses and Lactobacillus plantarum MTD-1. Ital. J. Anim. Sci. 2019, 18, 328–335. [Google Scholar] [CrossRef]
- Yang, C.M.J.; Huang, S.C.; Chang, T.; Cheng, Y.H.; Chang, C.T. Fermentation acids, aerobic fungal growth, and intake of napiergrass ensiled with nonfiber carbohydrates. J. Dairy Sci. 2004, 87, 630–636. [Google Scholar] [CrossRef]
- Yunus, M.; Ohba, N.; Tobisa, M.; Nakano, Y.; Shimojo, M.; Furuse, M.; Masuda, Y. Improving fermentation and nutritive quality of napiergrass silage by mixing with phasey bean. Asian-Australas. J. Anim. Sci. 2001, 14, 947–950. [Google Scholar] [CrossRef]
- Lunsin, R.; Pilajun, R.; Cherdthong, A.; Wanapat, M.; Duanyai, S.; Sombatsri, P. Influence of fibrolytic enzymes in total mixed ration containing urea-molasses-treated sugarcane bagasse on the performance of lactating Holstein-Friesian crossbred cows. Anim. Sci. J. 2021, 92, e13652. [Google Scholar] [CrossRef]
- Sorensen, A.; Lubeck, M.; Lubeck, P.S.; Ahring, B.K. Fungal Beta-glucosidases: A bottleneck in industrial use of lignocellulosic materials. Biomolecules 2013, 3, 612–631. [Google Scholar] [CrossRef]
- Khota, W.; Pholsen, S.; Higgs, D.; Cai, Y.M. Comparative analysis of silage fermentation and in vitro digestibility of tropical grass prepared with Acremonium and Tricoderma species producing cellulases. Asian-Australas. J. Anim. Sci. 2018, 31, 1913–1922. [Google Scholar] [CrossRef]
- Ren, H.W.; Wang, L.; Sun, Y.A.; Zhao, Q.L.; Sun, Y.M.; Li, J.P.; Zhang, B.Y. Enhancing the Co-ensiling Performance of Corn Stover and Cabbage Waste via the Addition of Cellulase. Bioresources 2021, 16, 6342–6362. [Google Scholar] [CrossRef]
- Ding, D.Y.; Zhou, X.; Ji, Z.; You, T.T.; Xu, F. How Does Hemicelluloses Removal Alter Plant Cell Wall Nanoscale Architecture and Correlate with Enzymatic Digestibility? Bioenergy Res. 2016, 9, 601–609. [Google Scholar] [CrossRef]
- Campana, M.; de Morais, J.P.G.; Capucho, E.; Garcia, T.M.; Pedrini, C.A.; Gandra, J.R.; Del Valle, T.N. Fibrolytic enzymes increases fermentation losses and reduces fiber content of sorghum silage. Ann. Anim. Sci. 2023, 23, 165–172. [Google Scholar] [CrossRef]
- Li, G.H.; Sun, Y.X.; Guo, W.J.; Yuan, L. Comparison of various pretreatment strategies and their effect on chemistry and structure of sugar beet pulp. J. Clean. Prod. 2018, 181, 217–223. [Google Scholar] [CrossRef]
- Shah, A.A.; Wu, J.Z.; Qian, C.; Liu, Z.W.; Mobashar, M.; Tao, Z.J.; Zhang, X.M.; Zhong, X.X. Ensiling of whole-plant hybrid pennisetum with natamycin and Lactobacillus plantarum impacts on fermentation characteristics and meta-genomic microbial community at low temperature. J. Sci. Food Agric. 2020, 100, 3378–3385. [Google Scholar] [CrossRef] [PubMed]
- AOAC. Official Analytical Chemists, 15th ed.; Association of Official Analytical Chemist: Arlington, VA, USA, 1990. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for Dietary Fiber, Neutral Detergent Fiber, and Nonstarch Polysaccharides in Relation To Animal Nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef] [PubMed]
- Tucker, W.B.; Hogue, J.F.; Aslam, M.; Lema, M.; Martin, M.; Owens, F.N.; Shin, I.S.; Leruyet, P.; Adams, G.D. A Buffer Value Index To Evaluate Effects of Buffers on Ruminal Milieu in Cows Fed High or Low Concentrate, Silage, or Hay Diets. J. Dairy Sci. 1992, 75, 811–819. [Google Scholar] [CrossRef]
- Dong, D.; Xu, G.F.; Dai, T.T.; Zong, C.; Yin, X.J.; Bao, Y.H.; Shao, T. Effect of molasses on fermentation quality of wheat straw ensiled with perennial ryegrass. Anim. Prod. Sci. 2022, 62, 1471–1479. [Google Scholar] [CrossRef]
- Kung, L.M.; Shaver, R.D.; Grant, R.J.; Schmidt, R.J. Silage review: Interpretation of chemical, microbial, and organoleptic components of silages. J. Dairy Sci. 2018, 101, 4020–4033. [Google Scholar] [CrossRef]
- Bao, X.Y.; Feng, H.R.; Guo, G.; Huo, W.J.; Li, Q.H.; Xu, Q.F.; Liu, Q.; Wang, C.; Chen, L. Effects of laccase and lactic acid bacteria on the fermentation quality, nutrient composition, enzymatic hydrolysis, and bacterial community of alfalfa silage. Front. Microbiol. 2022, 13, 1035942. [Google Scholar] [CrossRef]
- Du, Z.M.; Yamasaki, S.; Oya, T.; Cai, Y.M. Cellulase-lactic acid bacteria synergy action regulates silage fermentation of woody plant. Biotechnol. Biofuels Bioprod. 2023, 16, 125. [Google Scholar] [CrossRef]
- Liu, J.Y.; Zhao, M.Q.; Hao, J.F.; Yan, X.Q.; Fu, Z.H.; Zhu, N.; Jia, Y.S.; Wang, Z.J.; Ge, G.T. Effects of temperature and lactic acid Bacteria additives on the quality and microbial community of wilted alfalfa silage. BMC Plant Biol. 2024, 24, 844. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.Q.; Wu, H.; Li, L.; He, J.J.; Hu, Z.C.; Yang, X.J.; Xie, X.X. Effects of applying cellulase and starch on the fermentation characteristics and microbial communities of Napier grass (Pennisetum purpureum Schum.) silage. J. Anim. Sci. Technol. 2021, 63, 1301–1313. [Google Scholar] [CrossRef] [PubMed]
- Huisden, C.M.; Adesogan, A.T.; Kim, S.C.; Ososanya, T. Effect of applying molasses or inoculants containing homofermentative or heterofermentative bacteria at two rates on the fermentation and aerobic stability of corn silage. J. Dairy Sci. 2009, 92, 690–697. [Google Scholar] [CrossRef] [PubMed]
- Murray, J.; Dunnett, C.; Moore-Colyer, M.J.S.; Longland, A.C. In vitro assessment of three fibrolytic enzyme preparations as potential feed additives in equine diets. Anim. Feed Sci. Technol. 2012, 171, 192–204. [Google Scholar] [CrossRef]
- Saparrat, M.C.N.; Arambarri, A.M.; Balatti, P.A. Growth response and extracellular enzyme activity of Ulocladium botrytis LPSC 813 cultured on carboxy-methylcellulose under a pH range. Biol. Fertil. Soils 2007, 44, 383–386. [Google Scholar] [CrossRef]
- Khalil, A.I. Production and characterization of cellulolytic and xylanolytic enzymes from the ligninolytic white-rot fungus Phanerochaete chrysosporium grown on sugarcane bagasse. World J. Microbiol. Biotechnol. 2002, 18, 753–759. [Google Scholar] [CrossRef]
- Jung, H.G.; Smith, R.R.; Endres, C.S. Cell-wall composition and degradability of stem tissue from lucerne divergently selected for lignin and in-vitro dry-matter disappearance. Grass Forage Sci. 1994, 49, 295–304. [Google Scholar] [CrossRef]
- de Melo, N.N.; Carvalho-Estrada, P.D.; Tavares, Q.G.; Pereira, L.D.; Vigne, G.L.D.; Rezende, D.M.L.C.; Schmidt, P. The Effects of Short-Time Delayed Sealing on Fermentation, Aerobic Stability and Chemical Composition on Maize Silages. Agronomy 2023, 13, 223. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Items | Value |
|---|---|
| Chemical composition | |
| DM (g kg−1 FW) | 378 |
| WSC (g kg−1 DM) | 90.2 |
| CP (g kg−1 DM) | 55.1 |
| EE (g kg−1 DM) | 42.6 |
| Ash (g kg−1 DM) | 68.8 |
| Buffering capacity (mEq kg−1 DM) | 102 |
| NDF (g kg−1 DM) | 541 |
| ADF (g kg−1 DM) | 389 |
| ADL (g kg−1 DM) | 114 |
| Hemicellulose (g kg−1 DM) | 152 |
| Cellulose (g kg−1 DM) | 275 |
| Lignin (g kg−1 DM) | 46.5 |
| microbial composition | |
| Lactic acid bacteria (log10 CFU g−1 FW) | 3.17 |
| Aerobic bacteria (log10 CFU g−1 FW) | 7.54 |
| Yeast (log10 CFU g−1 FW) | 5.60 |
| Items | Treatments | Days of Ensiling | Mean | SEM | p-Value | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 3 | 7 | 30 | T | D | T × D | ||||
| DM (g kg−1 FW) | Control | 373 | 360 | 354 | 346 | 358AB | 2.31 | <0.001 | <0.001 | 0.078 |
| L | 371 | 364 | 360 | 357 | 363A | |||||
| E | 378 | 363 | 353 | 341 | 355B | |||||
| EL | 376 | 370 | 367 | 361 | 369A | |||||
| Mean | 375a | 364b | 359bc | 351c | ||||||
| pH | Control | 5.22 | 4.78 | 4.71 | 4.56 | 4.82A | 0.106 | <0.001 | <0.001 | 0.306 |
| L | 4.94 | 4.79 | 4.63 | 4.14 | 4.63A | |||||
| E | 5.12 | 4.85 | 4.57 | 4.43 | 4.74A | |||||
| EL | 4.51 | 4.23 | 3.81 | 3.76 | 4.08B | |||||
| Mean | 4.95a | 4.66b | 4.43 | 4.22c | ||||||
| Lactic acid (g kg−1 DM) | Control | 18.5Bb | 20.0Bb | 22.4Ca | 24.1Ca | 21.3 | 2.60 | <0.001 | <0.001 | <0.001 |
| L | 21.3Ac | 25.1Bbc | 28.2Bb | 33.1Ba | 26.9 | |||||
| E | 17.4Bb | 18.9Cb | 24.1BCa | 25.9Ca | 21.6 | |||||
| EL | 21.1Ad | 33.5Ac | 42.0Ab | 58.2Aa | 38.7 | |||||
| Mean | 19.6 | 24.4 | 29.2 | 35.3 | ||||||
| WSC (g kg−1 DM) | Control | 78.6ABa | 68.1Ab | 52.5Ac | 33.9Ad | 58.3 | 5.22 | <0.001 | <0.001 | 0.001 |
| L | 74.4Ba | 54.5Bb | 40.7Bc | 24.6Bd | 48.6 | |||||
| E | 87.6Aa | 70.3Ab | 54.9Ac | 31.2Ad | 61.0 | |||||
| EL | 84.2Aa | 64.2ABb | 42.0Bc | 22.3Bd | 53.2 | |||||
| Mean | 81.2 | 64.3 | 42.0 | 22.3 | ||||||
| NH3-N (g kg−1 TN) | Control | 26.7d | 42.1Ac | 55.4Ab | 75.2Aa | 49.9 | 4.08 | <0.001 | <0.001 | <0.001 |
| L | 28.3c | 34.6ABbc | 40.1Bb | 62.5Ba | 41.4 | |||||
| E | 27.8d | 37.9ABc | 45.0Bb | 76.3Aa | 46.8 | |||||
| EL | 26.7c | 31.2Bc | 39.4Bb | 57.2Ba | 38.6 | |||||
| Mean | 27.4 | 36.5 | 45.0 | 67.8 | ||||||
| Items | Treatments | Days of Ensiling | Mean | SEM | p-Value | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 3 | 7 | 30 | T | D | T × D | ||||
| Acetic acid (g kg−1 DM) | Control | 7.44c | 10.8b | 12.3ab | 14.4Ba | 11.2 | 0.855 | 0.011 | <0.001 | <0.001 |
| L | 6.98c | 8.95bc | 10.2b | 13.2Ba | 9.83 | |||||
| E | 7.58c | 9.83bc | 11.8b | 15.2Ba | 11.1 | |||||
| EL | 6.25c | 8.14bc | 9.88b | 18.6Aa | 10.7 | |||||
| Mean | 7.06 | 9.43 | 11.0 | 15.4 | ||||||
| LA/AA | Control | 2.49 | 1.85 | 1.82 | 1.67 | 1.96C | 0.220 | <0.001 | 0.005 | 0.080 |
| L | 3.05 | 2.80 | 2.76 | 2.51 | 2.78B | |||||
| E | 2.30 | 1.92 | 2.04 | 1.70 | 1.99C | |||||
| EL | 3.38 | 4.12 | 4.25 | 3.13 | 3.72A | |||||
| Mean | 2.81a | 2.67a | 2.72a | 2.25b | ||||||
| Propionic acid (g kg−1 DM) | Control | ND | 0.39Bb | 0.59b | 0.88Ba | 0.62 | 0.098 | <0.001 | <0.001 | <0.001 |
| L | ND | 0.20C | 0.33 | 0.35C | 0.29 | |||||
| E | ND | 0.66Ab | 0.70b | 1.29Aa | 0.88 | |||||
| EL | ND | ND | ND | ND | ||||||
| Mean | 0.42 | 0.54 | 0.84 | |||||||
| Butyric acid (g kg−1 DM) | Control | ND | ND | 0.22ABb | 0.68a | 0.45 | 0.070 | <0.001 | <0.001 | <0.001 |
| L | ND | ND | 0.08B | 0.19 | 0.14 | |||||
| E | ND | ND | 0.26Ab | 0.94a | 0.60 | |||||
| EL | ND | ND | ND | ND | ||||||
| Mean | 0.19 | 0.60 | ||||||||
| Items | Treatments | Days of Ensiling | Mean | SEM | p-Value | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 3 | 7 | 30 | T | D | T × D | ||||
| NDF (g kg−1 DM) | Control | 539Aa | 534Aa | 530Aab | 518Ab | 530 | 11.9 | <0.001 | <0.001 | <0.001 |
| L | 536Aa | 533Aa | 530Aa | 501Ab | 521 | |||||
| E | 510Ba | 486Bab | 461Bb | 382Bc | 461 | |||||
| EL | 512Ba | 497Bab | 477Bb | 395Bc | 470 | |||||
| Mean | 524 | 513 | 500 | 449 | ||||||
| ADF (g kg−1 DM) | Control | 386 | 382 | 375A | 369A | 379 | 6.22 | <0.001 | <0.001 | <0.001 |
| L | 384 | 383 | 377A | 358A | 373 | |||||
| E | 379a | 368ab | 351Bb | 305Bc | 351 | |||||
| EL | 376a | 370a | 356Ba | 312Bb | 354 | |||||
| Mean | 381 | 376 | 365 | 336 | ||||||
| ADL (g kg−1 DM) | Control | 114 | 113 | 114 | 110 | 113A | 0.931 | <0.001 | 0.001 | 0.163 |
| L | 113 | 115 | 112 | 104 | 111AB | |||||
| E | 109 | 112 | 109 | 106 | 109B | |||||
| EL | 111 | 110 | 108 | 105 | 109B | |||||
| Mean | 112a | 113a | 111a | 106b | ||||||
| Ash (g kg−1 DM) | Control | 68.3 | 69.6 | 69.1 | 66.2 | 68.3 | 0.759 | 0.267 | 0.101 | 0.573 |
| L | 65.9 | 65.9 | 67.8 | 62.5 | 65.5 | |||||
| E | 66.7 | 69.4 | 67.4 | 64.8 | 67.1 | |||||
| EL | 65.5 | 68.5 | 65.4 | 64.8 | 66.1 | |||||
| Mean | 66.6 | 68.4 | 67.4 | 64.6 | ||||||
| Items | Treatments | Days of Ensiling | Mean | SEM | p-Value | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 3 | 7 | 30 | T | D | T × D | ||||
| Hemicellulose (g kg−1 DM) | Control | 153 | 152 | 155A | 148A | 152 | 6.49 | <0.001 | <0.001 | 0.006 |
| L | 152 | 149 | 153A | 141A | 149 | |||||
| E | 131a | 118a | 110Ba | 76.5Bb | 109 | |||||
| EL | 136a | 128a | 121Ba | 83.5Bb | 117 | |||||
| Mean | 143 | 137 | 135 | 112 | ||||||
| Cellulose (g kg−1 DM) | Control | 272 | 269 | 261AB | 259A | 265 | 5.76 | <0.001 | <0.001 | <0.001 |
| L | 273 | 269 | 265A | 259A | 266 | |||||
| E | 270a | 256ab | 242Cb | 199Bc | 242 | |||||
| EL | 265a | 260a | 248BCa | 206Bb | 245 | |||||
| Mean | 270 | 264 | 254 | 231 | ||||||
| Lignin (g kg−1 DM) | Control | 46.0 | 43.4 | 44.9 | 43.9 | 44.6 | 0.870 | 0.074 | 0.357 | 0.729 |
| L | 45.4 | 49.0 | 44.4 | 43.4 | 45.6 | |||||
| E | 42.4 | 42.7 | 42.1 | 41.6 | 42.2 | |||||
| EL | 45.8 | 41.6 | 42.6 | 40.8 | 42.7 | |||||
| Mean | 44.9 | 44.2 | 43.5 | 42.4 | ||||||
| Items | Treatments | Days of Ensiling | Mean | SEM | p-Value | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 3 | 7 | 30 | T | D | T × D | ||||
| LAB (log10 CFU g−1 FW) | Control | 3.51Bc | 3.89Bbc | 4.22Bb | 5.25Ca | 4.22 | 0.364 | <0.001 | <0.001 | <0.001 |
| L | 5.68Ac | 6.62Ab | 7.37Aa | 6.27Bbc | 6.49 | |||||
| E | 3.32Bc | 4.05Bb | 4.59Bb | 5.44Ca | 4.35 | |||||
| EL | 5.81Ac | 6.74Ab | 7.55Aa | 7.84Aa | 6.99 | |||||
| Mean | 4.58 | 5.33 | 5.93 | 6.20 | ||||||
| Aerobic bacteria (log10 CFU g−1 FW) | Control | 7.42a | 5.08Ab | 4.70Ab | 4.52Ab | 5.43 | 0.373 | <0.001 | <0.001 | 0.003 |
| L | 7.01a | 4.66ABb | 4.03ABbc | 3.63Bc | 4.83 | |||||
| E | 7.29a | 5.23Ab | 4.41Ac | 4.12ABc | 5.26 | |||||
| EL | 7.33a | 4.01Bb | 3.42Bb | 2.65Cc | 4.35 | |||||
| Mean | 7.26 | 4.75 | 4.14 | 3.73 | ||||||
| Yeasts (log10 CFU g−1 FW) | Control | 5.52 | 3.08 | 2.41 | 2.13 | 3.29AB | 0.352 | <0.001 | <0.001 | 0.118 |
| L | 5.40 | 2.56 | 2.00 | 2.00 | 2.99B | |||||
| E | 5.61 | 3.37 | 2.48 | 2.39 | 3.46A | |||||
| EL | 5.36 | 2.33 | 2.00 | 2.00 | 2.92B | |||||
| Mean | 5.47a | 2.84b | 2.22bc | 2.13c | ||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, D.; Zhang, L.; Zhao, J.; Dong, Z.; Li, J.; Shao, T. Synergistic Effects of Exogenous Lactobacillus plantarum and Fibrolytic Enzymes on Fermentation Quality, Fiber Degradation, and In Vitro Digestibility of Napiergrass (Pennisetum purpureum) Silage. Agronomy 2025, 15, 340. https://doi.org/10.3390/agronomy15020340
Dong D, Zhang L, Zhao J, Dong Z, Li J, Shao T. Synergistic Effects of Exogenous Lactobacillus plantarum and Fibrolytic Enzymes on Fermentation Quality, Fiber Degradation, and In Vitro Digestibility of Napiergrass (Pennisetum purpureum) Silage. Agronomy. 2025; 15(2):340. https://doi.org/10.3390/agronomy15020340
Chicago/Turabian StyleDong, Dong, Lei Zhang, Jie Zhao, Zhihao Dong, Junfeng Li, and Tao Shao. 2025. "Synergistic Effects of Exogenous Lactobacillus plantarum and Fibrolytic Enzymes on Fermentation Quality, Fiber Degradation, and In Vitro Digestibility of Napiergrass (Pennisetum purpureum) Silage" Agronomy 15, no. 2: 340. https://doi.org/10.3390/agronomy15020340
APA StyleDong, D., Zhang, L., Zhao, J., Dong, Z., Li, J., & Shao, T. (2025). Synergistic Effects of Exogenous Lactobacillus plantarum and Fibrolytic Enzymes on Fermentation Quality, Fiber Degradation, and In Vitro Digestibility of Napiergrass (Pennisetum purpureum) Silage. Agronomy, 15(2), 340. https://doi.org/10.3390/agronomy15020340