Abstract
Winter rapeseed is susceptible to freezing stress during winter, making it difficult to overwinter safely and resulting in a reduction of yield and quality. DNA methylation, the main epigenetic modification, can regulate plant responses to various stresses. However, the regulatory mechanism of DNA methylation in response to freezing stress in winter rapeseed remains unclear. This study investigates how DNA methylation regulates gene expression and protein abundance in response to freezing stress, revealing key regulatory pathways involved in rapeseed cold tolerance. A total of 6776 unique differentially methylated genes (DMGs), 4285 unique differentially expressed genes (DEGs), and 269 unique differentially abundant proteins (DAPs) were identified between the two cultivars under T1 and T2 freezing stress. Function enrichment analysis revealed that these genes were involved in signal transduction, biosynthesis of unsaturated fatty acids, sugar metabolism, peroxidase, peroxisome, photosynthesis, and additional pathways. An integrative analysis of methylome, transcriptome, and proteome showed that only nine genes were shared among all three datasets, and they were closely related to cold tolerance metabolism in rapeseed. The findings provide molecular insights into rapeseed freezing tolerance, which can be applied in breeding programs to enhance cold resistance in oilseed crops.
1. Introduction
Rapeseed is widely distributed in China, with an annual sowing area of 7.3 million square hectares and a total rapeseed production of 14.3 million tons. However, the self-sufficiency rate of vegetable oil in China is seriously insufficient, with about 64.7% relying on foreign imports, and the dependence is increasing [1,2]. The large areas of idle winter fields in northern China have the potential for winter oilseed rape production, which could improve the self-sufficiency of rapeseed [3]. The biggest challenge to the development of the oilseed rape industry in this region is freezing stress during winter that makes it difficult for seedlings to survive until spring, resulting in reduced yields and quality [4]. The lowest temperature in northern China is lower than −25 °C, causing fatal freezing damage to crops, which restricts the growth, development, and geographic distribution of plants [5]. In recent years, significant progress has been made in breeding strong freezing-tolerant rapeseed cultivars such as Ganyou221. These cultivars can survive normally in most northwestern areas with temperatures as low as −26 °C [3,6]. The fundamental reason for this breakthrough is the introduction of cold resistance genes from winter turnip rape through distant crossbreeding. However, limited information is available regarding the mechanism of freezing tolerance in winter rapeseed.
DNA methylation is a major epigenetic modification and an essential regulator of gene imprinting and gene expression, and it does not alter the DNA sequence during plant development and stress defense [7,8,9]. Typically, DNA methylation is referred to as the fifth base and functions at the transcriptional level. Additionally, DNA methylation is associated with a plant’s stress memory, enabling it to respond faster and more accurately to the same stress [10]. Under the cold stress, plant DNA methylation levels are generally down-regulated, activating cold-resistance regulatory pathways or genes and leading to a significant increase in plant cold resistance [11,12]. However, there have been inconsistent reports on the regulatory mode of DNA methylation, which may be related to its location. The patterns of DNA methylation in winter rapeseed were investigated during freezing stress using whole genome bisulfite sequencing (WGBS), revealing a decrease in methylation levels in three contexts after freezing stress [13,14]. Furthermore, some genes and pathways related to rapeseed cold tolerance were found to be regulated by DNA methylation. We hypothesize that DNA methylation plays a critical role in regulating key cold tolerance pathways in rapeseed by modulating gene expression and protein abundance.
Given that winter temperatures in rapeseed-growing regions frequently drop below −10 °C, understanding the molecular basis of freezing tolerance is essential for developing cold-resistant cultivars. In this study, we analyzed the morphological traits, overwintering rate, physiology and biochemistry characteristics, photosynthetic indices, and submicroscopic structure features of a pair of cold-resistant and cold-sensitive rapeseed cultivars after freezing stress. The results showed that the cold-resistant cultivars had stronger activity of antioxidant enzymes, higher content of osmoregulatory substances, and more stable chloroplast and membrane structures under stress compared to the cold-sensitive cultivars, which made them exhibit a stronger overwintering rate. Then, we investigated the transcriptome, proteome, and methylome differences between the two cultivars, using RNA-seq, DIA, and WGBS. At the three omics levels, genes with common differences between both cultivars were screened for functional annotation and enrichment analysis. These findings will further supplement and improve our understanding of the methylation-regulated cold-tolerance mechanism in rapeseed under freezing stress.
2. Materials and Methods
2.1. Plant Samples and Freezing Stress Treatments
Two rapeseed cultivars were used in this research, provided by Gansu Agricultural University. The freezing-resistant cultivar 17NS57 (NS) originates from distant hybridization and cold acclimation between Brassica napus and Brassica rapa, exhibiting strong freezing tolerance in China’s northwestern areas. The freezing-sensitive cultivar NQF24 (NF) is a normal spring rapeseed cultivar with a survival rate of 0% below −10 °C. Full seeds were selected for germination on filter paper in a culture dish at 22/20 °C day/night temperature with a 16 h/8 h light/dark cycle. Seedlings of both cultivars were then transferred to pots (5 L) filled with a mixture of nutritional soil and vermiculite (3:1 ratio) and grown under normal conditions at the same temperature/light cycle as before. The soil used in the pot experiments was a loamy substrate with a pH of 6.5, an organic matter content of 3.2%, and a cation exchange capacity (CEC) of 15 cmol/kg. The experiment was carried out in the greenhouse of Gansu Agricultural University in Lanzhou City, Gansu Province, in 2020. When the seedlings reached the five-leaf stage, they were exposed to two different freezing treatments (−4 °C for 12 h or T1; −4 °C for 24 h or T2), while the control group remained under normal conditions (T0). Leaves from each plant were taken from the same position, immediately frozen in liquid nitrogen, and stored at −80 °C for RNA/DNA/protein extraction, as well as physiological and biochemical characteristic determination.
2.2. Overwintering Rate, Physiological and Biochemical Determination
Two cultivars were sown in the field (Lanzhou, Gansu, 64°10′ N,103°71′ E), with each containing three replicates. The minimum winter temperature is −12 °C, the annual average temperature is 8.9 °C, the annual sunshine duration reaches 2476.4 h, and the frost-free period was 171 days in the field. One hundred plants were planted in each plot, with a row spacing of 25 cm and a plant spacing of 10 cm. The overwintering rate was determined by the ratio of the number of seedlings that successfully greened up after winter to the total number of planted seedlings. After freezing treatment, the third leaf was used to measure physiological and biochemical properties, including antioxidant enzymes [catalase (CAT), peroxidase (POD), and superoxide dismutase (SOD)] activities, malondialdehyde (MDA), and the contents of osmotic regulation substances [free proline content (Pro), soluble sugar (SS), and soluble protein (SP)]. The activities of SOD, POD, and CAT were estimated following Hassan et al. [15]. MDA, SS, SP, and Pro were measured according to Rong et al.’s methods [16]. Relative electrolyte leakage (REL) was determined using a fourth leaf by a digital conductivity meter DDS11A from Leici Instrument Factory, Shanghai, China, according to Wei et al. [5]. Each treatment was performed with three biological replicates, with at least 10 plants per replicate.
2.3. Determination of Photosynthetic Parameters and Observations Using a Transmission Electron Microscope
In repeated experiments, the third set of fresh leaves from both cultivars was used for photosynthesis evaluation. A LI-COR 6400 portable gas analysis system (Lincoln, NE, USA) was used with a built-in light source of 1000 μmol photons m−2·s−1. Additionally, the same fresh leaves from rapeseed plants were used for Transmission Electron Microscope Observations. In addition, the Transmission Electron Microscope (Tokyo, Japan) observations were conducted following the methodology described by Wei et al. [17].
2.4. DNA Extraction, DNA Methylation Sequencing, and Data Analysis
The total genomic DNA was extracted from six samples using a DNase Plant Mini Kit (Beijing, China) according to the manufacturer’s protocol. Subsequently, Illumina’s standard DNA methylation analysis protocol and a DNA Methylation-Gold Kit were employed for library construction. Paired end sequencing was conducted on the high-throughput Illumina HiSeqTM 2500 platform (San Diego, CA, USA), and high-quality clean reads were obtained by removing reads containing more than 10% of unknown nucleotides and more than 40% of low-quality (Q-value ≤ 20) bases from the raw data. The obtained clean reads were then mapped to the rapeseed reference genome using BSMAP software (version 2.90) with default settings. The methylation level was calculated based on the percentage of methylated cytosine in the genome. To identify DMRs between two cultivars in the same region, we followed Zheng et al. [14] standard criteria. A sliding-window approach with a window size of 200-bp sliding at intervals of 100-bp was used for DMR identification. Genes that overlapped with significant DMRs for at least 1 bp in both flanking regions within 2 kb and the gene body were defined as differentially methylated genes (DMGs), which wre utilized in subsequent analyses.
2.5. RNA Extraction, RNA Sequencing, and Data Filtering
The total RNA from treatment and control samples containing three biological replicates were extracted using the TRIzol Reagent (Beijing, China) according to the manufacturer’s instructions. The library construction and sequencing were performed by Gene Denovo Biotechnology Co. (Guangzhou, China) on an Illumina HiSeqTM 2500 platform. After the sequencing, raw sequences for each sample were filtered to generate clean reads for subsequent analysis. The expression level was normalized by calculating the fragments per kilobase of exon model per million mapped fragments (FPKM) value. The differentially expressed genes (DEGs) between both samples were identified using the criteria of a p-value < 0.001 and a value of |log2foldchange| ≥ 2.
2.6. Protein Extraction, Digestion, Mass Spectrometry Analysis, and Data Analysis
The total protein from each sample was extracted using the chilled acetone precipitation method, as described by Mi et al. [18]. Then, all samples were digested according to Illumina’s standard of protein digestion. Digested peptides from each sample were separated on an Ultimate 3000 system (Waltham, MA, USA) with a high pH reversed phase XBridge BEH C18 column. LC–MS/MS analysis was performed on an Orbitrap Fusion Mass Spectrometer (Waltham, MA, USA) coupled with a Nano ACQUITY UPLC system (Milford, MA, USA). For library generation and data acquisition in DDA mode, the Orbitrap Lumos was operated in positive mode. The Raw Data of DDA were processed and analyzed using Spectronaut Pulsar 11.0 (Schlieren, Switzerland). Proteins with p-value < 0.001, peptides ≥ 2, and value of |log2FoldChange| ≥ 1 were regarded as differentially abundant proteins (DAPs) using the edgeR package.
2.7. Functional Annotation and Statistical Analysis
The correlation coefficient between the three biological replicates was calculated to evaluate the repeatability of the experimental results between the samples in the transcriptome and proteome. Principal component analysis (PCA) was performed to reveal the structure/relationship of the samples using R package gmodels. Genes and proteins were annotated using the Gene Ontology (GO) database (http://geneontology.org/, accessed on 29 June 2020) and the Kyoto Encyclopedia of Genes and Genomes (KEGG) database (http://www.kegg.jp/, accessed on 29 June 2020) to determine their functions. Enrichment analysis of DEGs and DAPs were conducted on the GO database and KEGG database. Transcriptome, proteome, and methylation data were integrated to identify genes and proteins that are regulated by methylation that are related to cold resistance in rapeseed.
3. Result
3.1. Morphological, Overwintering Rate, Physiological and Biochemical Responses of Rapeseed to Freezing Stress
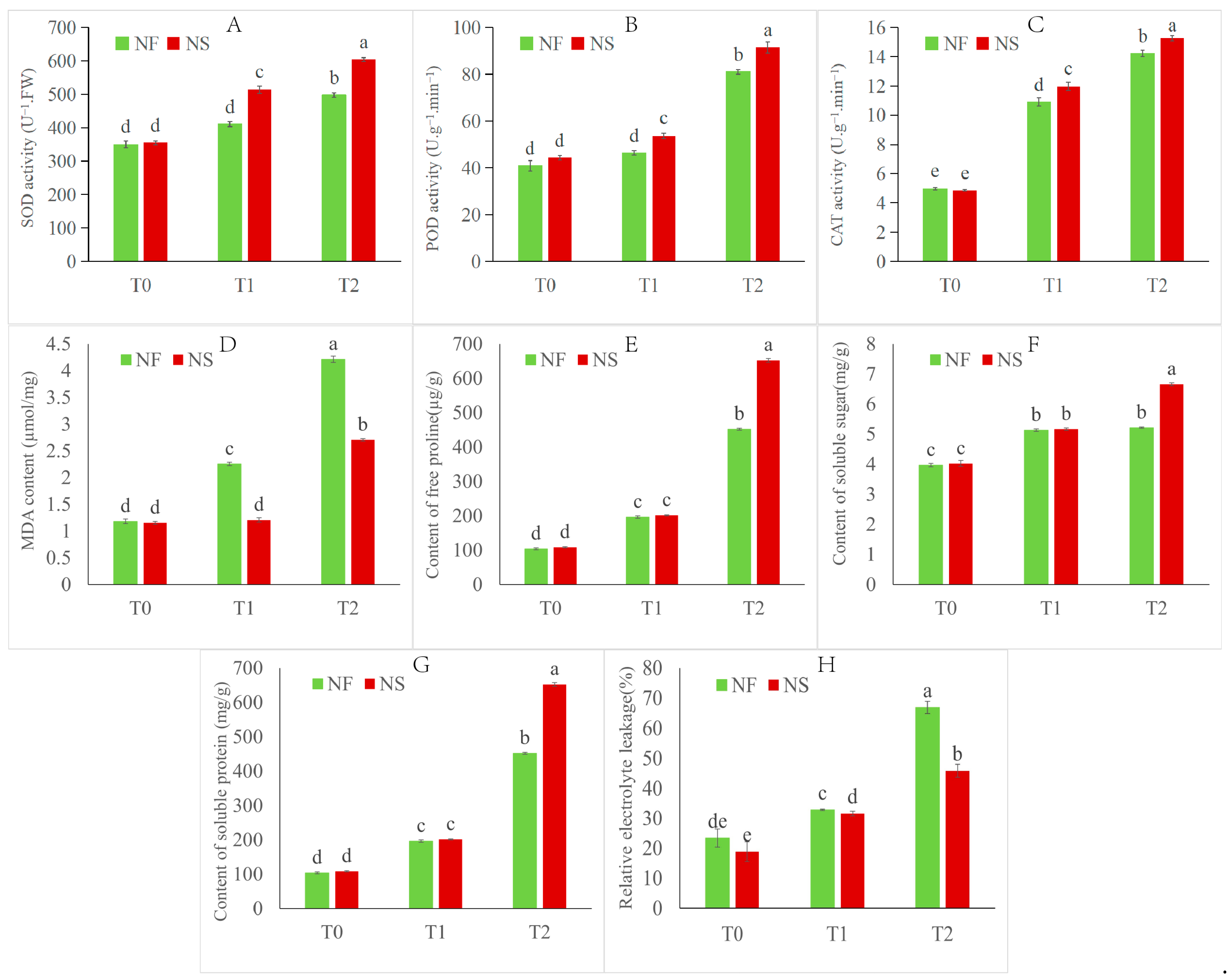
The NS leaf color is darker green than NF under the control treatment (Figure 1). After 12 h (T1) of freezing stress, the leaf color became darker, the leaves became wrinkled, and most leaves sagged, which clearly indicated damage caused by cold stress. After 24 h (T2) of freezing stress, leaf shrinkage became more severe, and some leaves wilted. In summary, NF damage was more obvious than NS during freezing stress treatment, and NS was able to recover after 12 h of freezing stress, while NF could not. Additionally, the overwintering rates of NS were close to 95%, whereas that of NF was close to 0% in the field. The overwintering rate of NS was significantly higher than that of NF (Figure 1). The activities of antioxidant enzymes (SOD, POD, and CAT), the contents of osmotic regulatory substances (SS, SP, Pro), the contents of malondialdehyde (MDA), and relative electrolyte leakage (REL) significantly (p < 0.05) increased under freezing stress for both cultivars (Figure 2). The enzyme activities of antioxidant enzymes and contents of osmotic regulatory substances in cultivar NS were higher compared to those of cultivar NF under freezing stress. Moreover, the MDA and REL in cultivar NF were significantly (p < 0.05) higher than those in cultivar NS, indicating that the NF plasma membrane damage in NF was more severe than in NS.

Figure 1.
Overwintering rate (A) and morphology (B) of winter rapeseed seedling under freezing stress.
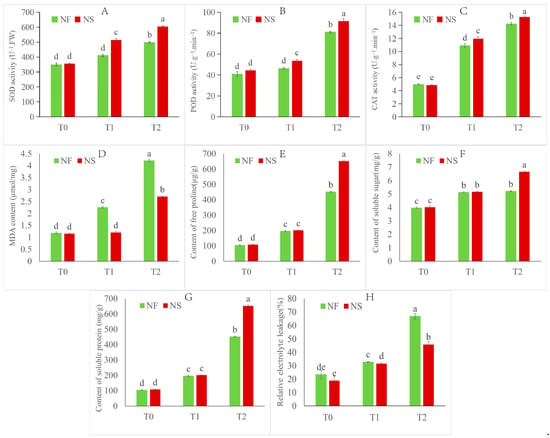
Figure 2.
Physiological and biochemical indexes of NS and NF under freezing stress. (A) SOD activity, (B) POD activity, (C) CAT activity, (D) MDA content, (E) free proline content, (F) soluble sugar, (G) soluble protein, (H) REL. Values of p are means ± SD from three biological replicates (p < 0.05). Different lowercase letters (a–e) in the figure represent significant differences in values.
3.2. Responses of Photosynthetic Parameters, Microstructure of Rapeseed to Freezing Stress
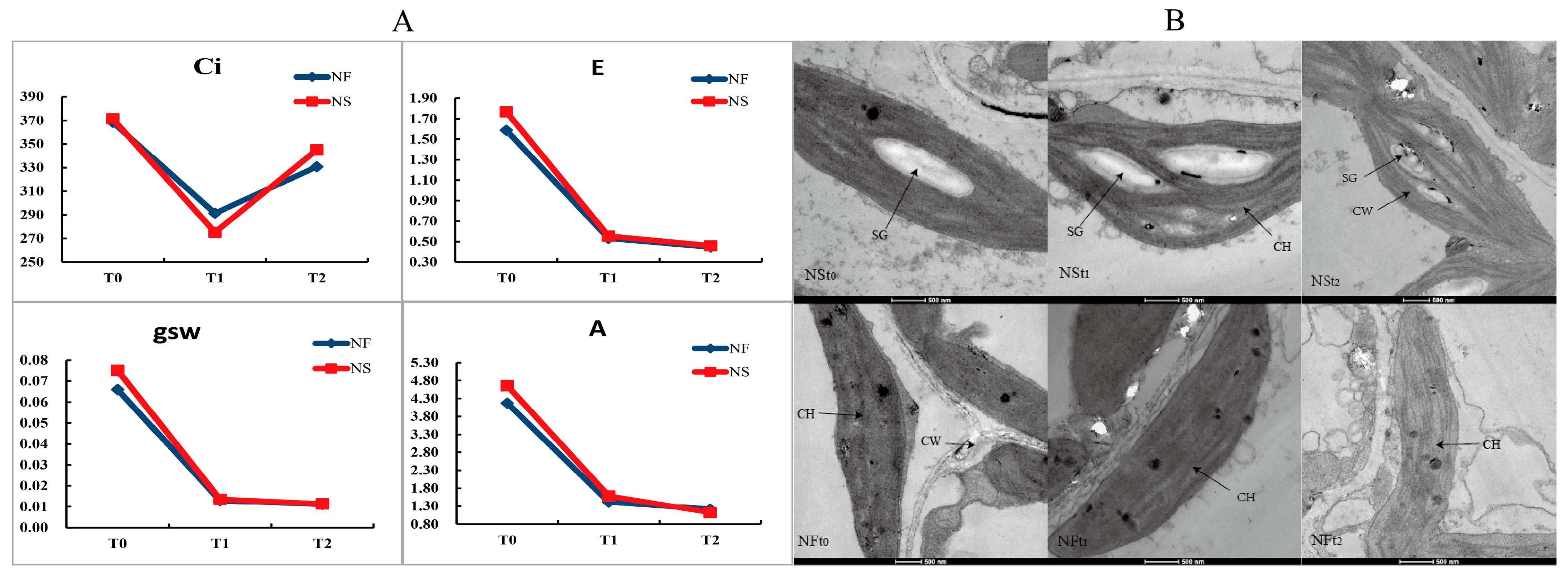
The intercellular carbon dioxide concentration (Ci) first decreased and then increased, while the net photosynthetic rate (A), transpiration rate (E), and stomatal conductance (gsw) decreased under freezing stress (Figure 3). However, there was no obvious difference in these parameters between the cultivars under freezing stress. Under control conditions, chloroplasts are single-sided convex lenses or fusiform shapes and are arranged regularly within the inner part of the cell wall. The chloroplasts contain a small number of elliptic starch grains, and the matrix lamellae are arranged in an orderly manner. After 12 h of freezing stress, the number of starch grains in the chloroplasts of cultivar NS leaves significantly increased, with smaller grain size and a few internal cavities appearing (Figure 3). In NF plants, there was an increase in both the number and size of internal cavities, a reduction in starch grains, and breakage of matrix lamellae within the chloroplasts after 24 h of freezing stress.
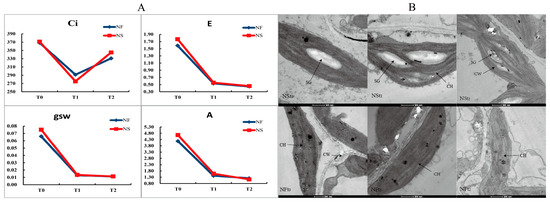
Figure 3.
Photosynthetic parameter (A) and microstructure (B) of winter rapeseed seedling under freezing stress. CH, chloroplast; CW, cell wall; SG, starch grain. This picture was the image under 500 nm of Transmission Electron Microscope.
3.3. Quality Control Analysis of the Rapeseed Methylome, Transcriptome, and Proteome
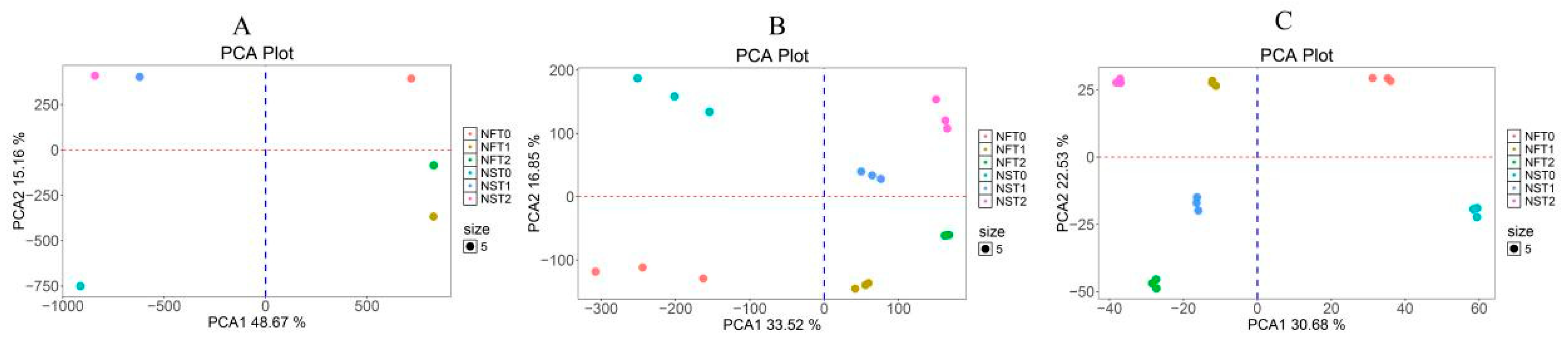
After filtering, a total of 39.72–59.44 million high-quality clean reads were generated per library, over 77% of the reads were uniquely mapped to the rapeseed genome, and the correlation coefficient between the three replicates was close to 99.9% (Table S2). A total of 85,338 genes were detected across all samples of both cultivars, including 6842 new genes and 78,496 known genes (Table S3). For the proteome analysis, 31,042 peptides from 38,804 precursors corresponded to 14,002 proteins (Table S4). The PCA indicated that three biological replicates of the transcriptome and proteome had good conformity, with a correlation coefficient exceeding 0.9 (Figure 4 and Table S5). The main source of variation in the first principal component of both the transcriptome and proteome data is the difference between different treatments, while the second principal component mainly represents the difference between cultivars. Additionally, the first principal component of the methylome data is mainly the difference between cultivars. These data indicate that both inter-cultivar differences and variations between treatments are sources of experimental variability, which should be taken into account in the subsequent analysis.

Figure 4.
Principal component analysis (PCA) of (A) detected sites in methylation sequencing, (B) detected genes in transcript sequencing, and (C) detected proteins in proteomics sequencing.
3.4. The DMGs, DEGs, and DAPs Were Identified Between Freezing-Resistant and Freezing-Sensitive Cultivars
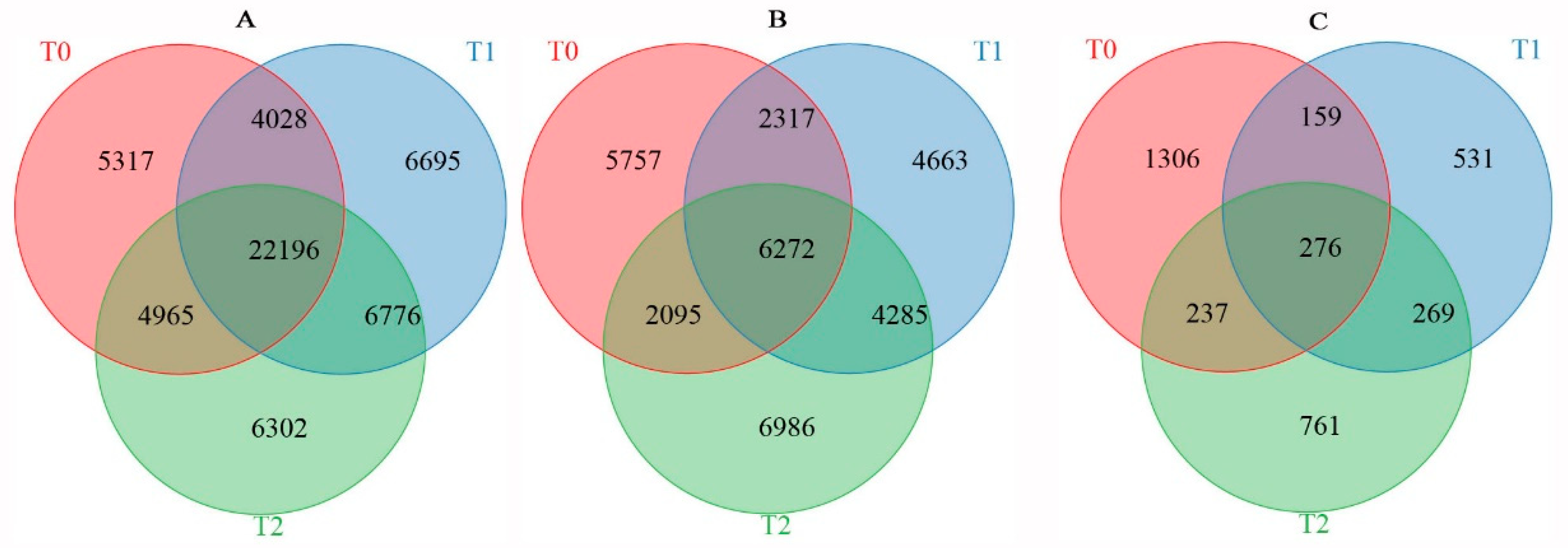
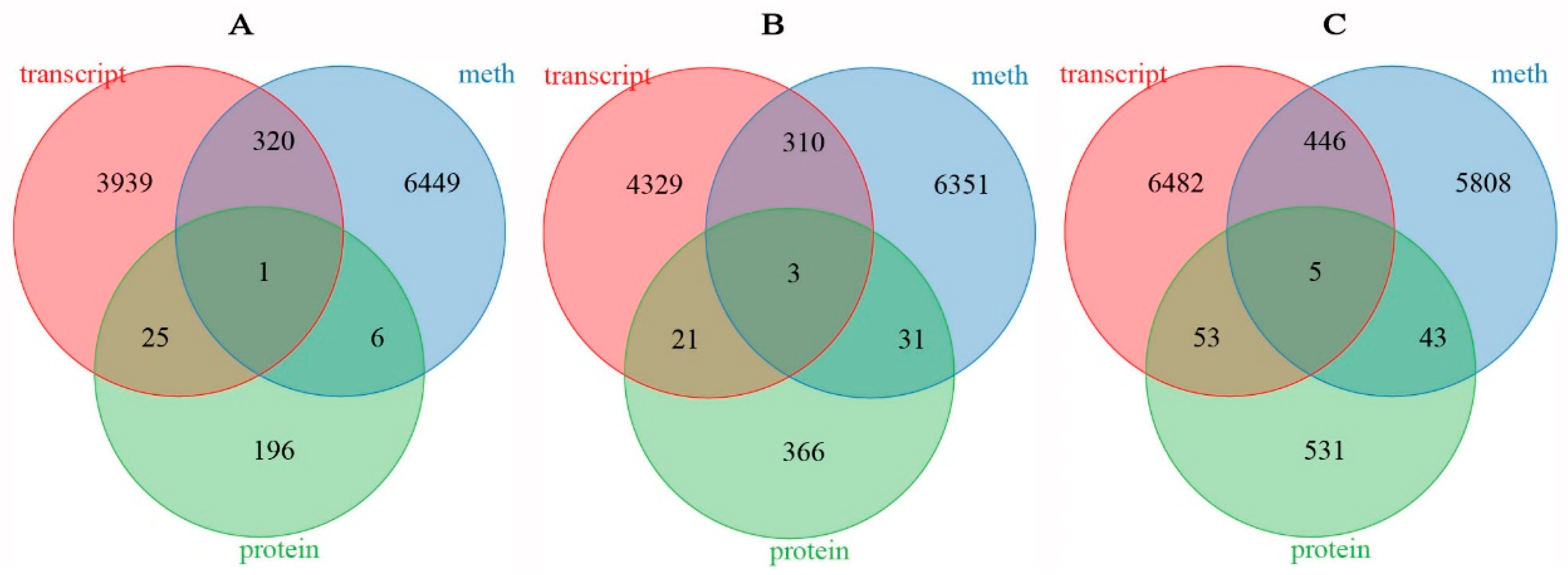
The average methylation level exhibited a downward trend in rapeseed seedings under freezing stress, and the methylation level of NF was lower than that of NS after freezing treatment. DNA methylation in winter rapeseed mainly occurred in CG, CHG, and CHH genomic sequence contexts. A total of 36,506, 39,695, and 40,239 genes between two cultivars showed significant differences in methylation levels under T0, T1, and T2 freezing stress. Among these genes, there were 6776 DMGs commonly identified in both T1 and T2 freezing stress, while 6695 DMGs and 6302 DMGs were specifically found in T1 and T2 freezing stress conditions, respectively. Correspondingly, a total of 164,41, 17,537, and 19,638 DEGs were identified in NS compared to NF under T0, T1, and T2 freezing stress. However, a total of 4285 DEGs were common to both T1 and T2, while 4663 DEGs and 6986 DEGs were uniquely identified in T1 and T2 freezing stress (Figure 5). Simultaneously, a total of 1978, 1235, and 1543 DAPs between the two cultivars were filtrated in T0, T1, and T2 freezing stress, respectively. Among them, 269 DAPs were commonly filtrated out in both T1 and T2, while 531 and 761 DAPs were only identified in T1 and T2.
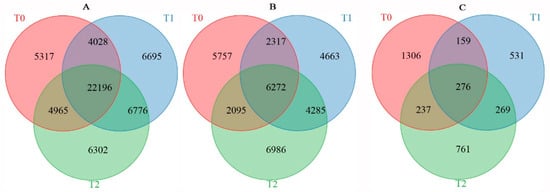
Figure 5.
Venn diagram of DMGs (A), DEGs (B), and DAPs (C) identified between resistant and sensitive freezing cultivars of winter rapeseed under freezing stress.
3.5. KEGG Analysis of DMGs, DEGs, and DAPs Under Freezing Stress
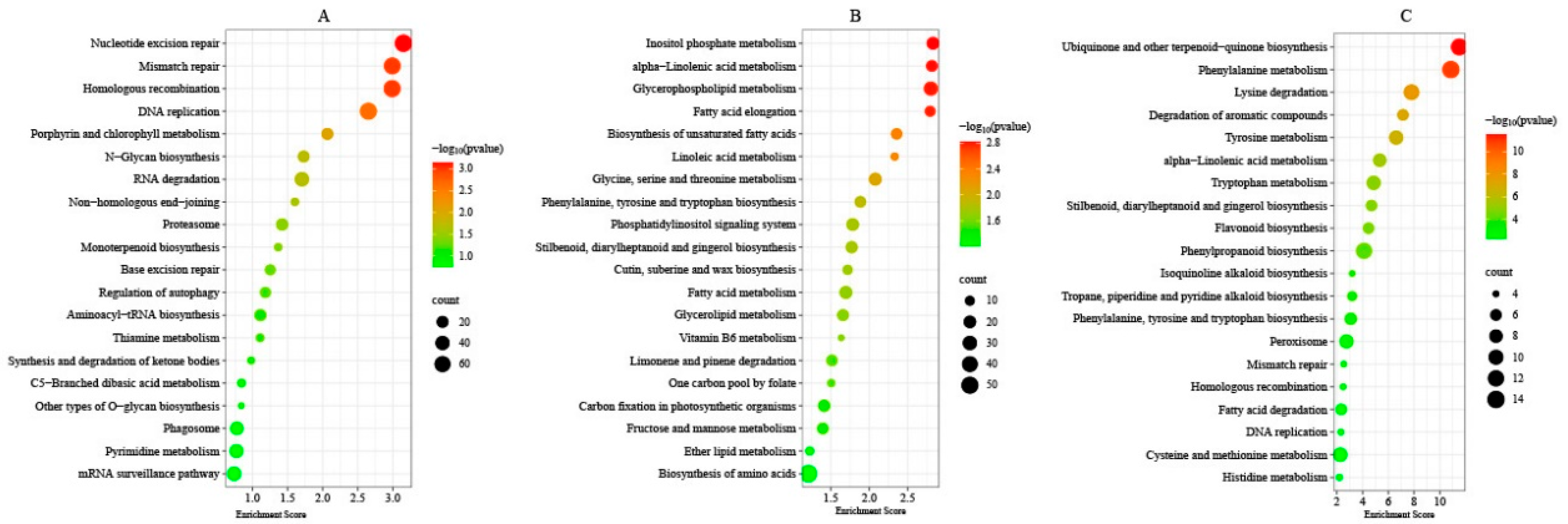
The specific DMGs, DEGs, and DAPs under T1 and T2 freezing stress were subjected to KEGG enrichment analysis in order to understand the cold-tolerance mechanism of winter rapeseed seedings under freezing stress. In total, 122, 120, and 74 KEGG pathways were successfully altered (Tables S6–S8). The KEGG enrichment pathways of DMGs showed that Nucleotide excision repair (ko03420), Mismatch repair (ko03430), Homologous recombination (ko03440), DNA replication (ko03030), and Porphyrin and chlorophyll metabolism (ko00860) were significantly (q < 0.05) enriched (Figure 6). The enrichment pathways of the DEGs exhibited genes mainly enriched in Inositol phosphate metabolism (ko00562), alpha-Linolenic acid metabolism (ko00592), Glycerophospholipid metabolism (ko00564), Fatty acid elongation (ko00062), and the Biosynthesis of unsaturated fatty acids (ko01040) (Figure 6). The result of DAPs showed that Ubiquinone and other terpenoid–quinone biosynthesis (ko00130), Phenylalanine metabolism (ko00360), Lysine degradation (ko00310), Degradation of aromatic compounds (ko01220), Tyrosine metabolism (ko00350), alpha-Linolenic acid metabolism (ko00592), Tryptophan metabolism (ko00380), and Flavonoid biosynthesis (ko00941) were the most enriched (Figure 6). Intriguingly, 69 pathways were enriched in both DEGs and DAPs, of which 52 pathways overlapped with DMGs’ enrichment pathways.

Figure 6.
KEGG pathway enrichment. Top 20 KEGG pathways enriched by unique differentially methylated genes (A), differentially expressed genes (B), and differentially abundant proteins (C) between both rapeseed cultivars at T1 and T2 freezing stress.
3.6. Identification and Analysis of TFs for Core DMGs, DEGs, and DAPs
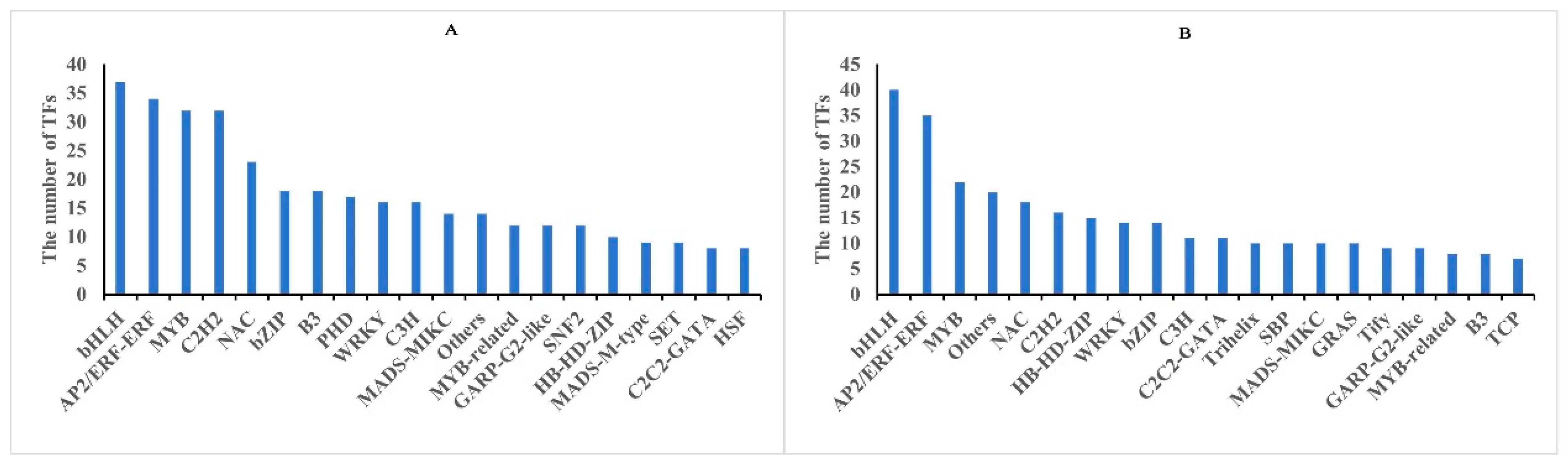
To date, a considerable number of TFs associated with cold stress have been isolated and confirmed in various plants. The result showed that 76 and 70 TFs were respectively identified from core DMGs and DEGs, while only one transcriptional regulatory factor (SET) was identified in the core DAPs (Tables S9 and S10). Among them, bHLH, AP2/ERF-ERF, MYB, NAC, and C2H2 were the most enriched TFs in the core DMGs and DEGs (Figure 7). Transcription factors such as bHLH and AP2/ERF exhibited differential expression, suggesting their involvement in cold stress signaling. These findings indicated that DNA methylation can regulate TFs under freezing stress.
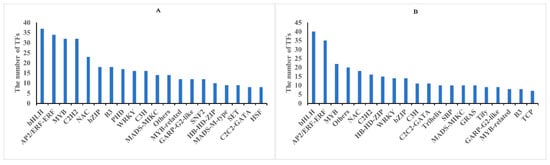
Figure 7.
Classification of differentially expressed transcription factors (TFs) in rapeseed seedings. TFs identified in methylome (A), and transcriptome (B) in between both rapeseed cultivars at T1 and T2 freezing stress, respectively.
3.7. Integrative Analysis of Methylome, Transcriptome, and Proteome
To understand the role of DNA methylation in the regulation of gene and protein expression under freezing stress, crossover analysis was used to integrate DEGs, DMGs, and genes encoding DAPs by their gene IDs. It should be noted that some proteins were encoded by a single gene. After removing redundancies, 228 DAPs were obtained for both T1 and T2 freezing stress. Additionally, 421 DAPs were obtained for T1 only and 632 DAPs were obtained for T2 only. However, only one gene showed significantly different transcription, methylation, and protein abundance levels in both T1 and T2 freezing stress, while three genes were identified only at T1 and five genes identified only at T2 (Figure 8). The result is challenging to attribute solely to the direct effect of DNA methylation on gene expression and protein abundance; it may involve more indirect mechanciem or be limited to the level of gene expression. Function enrichment showed that these genes encoded trypsin inhibitor-like (TIL), UDP-glucuronic acid decarboxylase 2 (UXS), gamma-glutamyl hydrolase (GGH), oxygen-evolving enhancer protein 2 (OEE2), Iron–sulfur cluster biosynthesis family protein (SUFB), subtilisin-like protease (SLP), Germin-like protein subfamily 2 member 4 (GLP2-4), beta-glucosidase (β-glu), and BnaC02g26290D, which are closely related to plant resistance to abiotic stress (Table S6). The TIL transcription and protein abundance level of NS were higher than NF in T1 treatment, and the methylation levels of the 2 kb upstream region under CHH context were up-regulated. TIL protein abundance was significantly decreased in T2 treatment and hypomethylated in the downstream. The UXS participated in amino sugar and nucleotide sugar metabolism (ko00520) and starch and sucrose metabolism (ko00500), which were up-regulated in transcription and protein abundance level in NS compared to NF, while it was hypomethylated in the gene body. The expression and protein abundance levels of GGH were also up-regulated in NS compared to NF, which was hypermethylated in the downstream. The other gene was down-regulated in transcription and protein abundance level in NS compared to NF, and it was hypermethylated in the gene body and downstream. Meanwhile, OEE2 was enriched in the Photosynthesis (ko00195) pathway, while BnaC01g40610D participated in starch and sucrose metabolism (ko00500) and cyanoamino acid metabolism (ko00460). Additionally, three other genes did not show enrichment in any of the analyzed pathway (Tables S6 and S7). The transcription and protein abundance level of OEE2 were down-regulated in NS compared to NF, and OEE2 was hypomethylated upstream. The BnaC01g40610D protein corresponded to beta-glucosidase (β-Glu), which showed down-regulation in transcription and protein abundance levels and was hypermethylated in the gene body. The SUFB exhibited up-regulation at the transcript level in NS compared to NF, along with hypermethylation in the gene body. SBT1.7 demonstrated up-regulation at both transcription and protein abundance levels, along with hypermethylation upstream. GLP2-4 displayed up-regulated expression but down-regulated protein abundance in this study, accompanied by an increase in methylation level downstream.
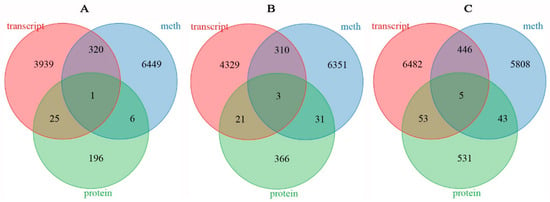
Figure 8.
Venn diagram of common genes at methylome, transcriptome, and proteome in both T1 and T2 (A), unique T1 (B), and unique T2 (C).
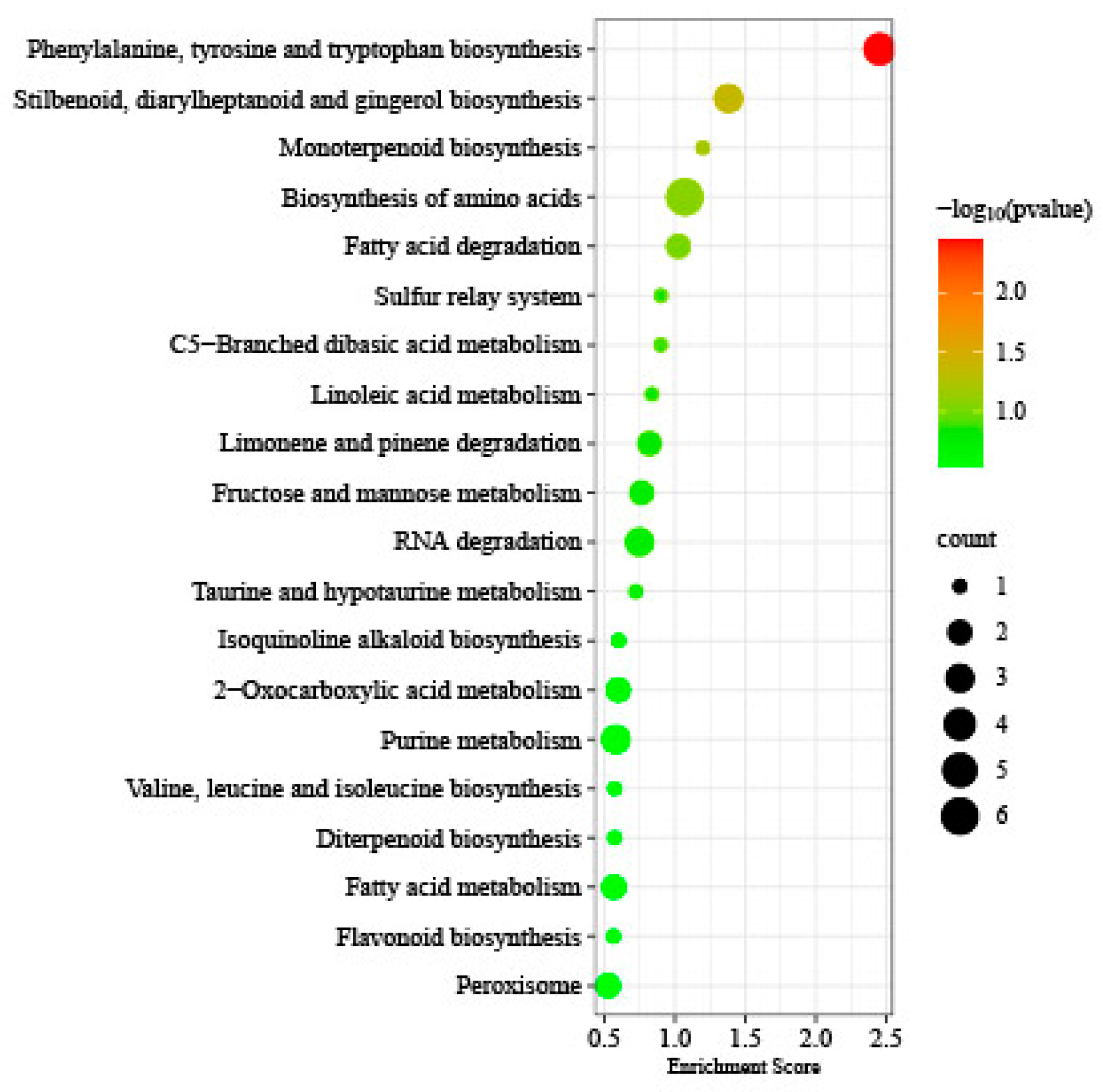
In addition, 320 genes were identified between DMGs and DEGs under T1 and T2 freezing stress. The KEGG enrichment analysis showed that Phenylalanine, tyrosine, and tryptophan biosynthesis (ko00400), Stilbenoid, diarylheptanoid, and gingerol biosynthesis (ko00945), Monoterpenoid biosynthesis (ko00902), Biosynthesis of amino acids (ko012300, Fatty acid degradation (ko00071), etc., were enriched. Furthermore, six genes were found to be present in both DMGs and DAPs, with only one of them encoding pyruvate dehydrogenase E1 component subunit alpha-2, which is reflected in carbohydrate metabolism (Figure 9).
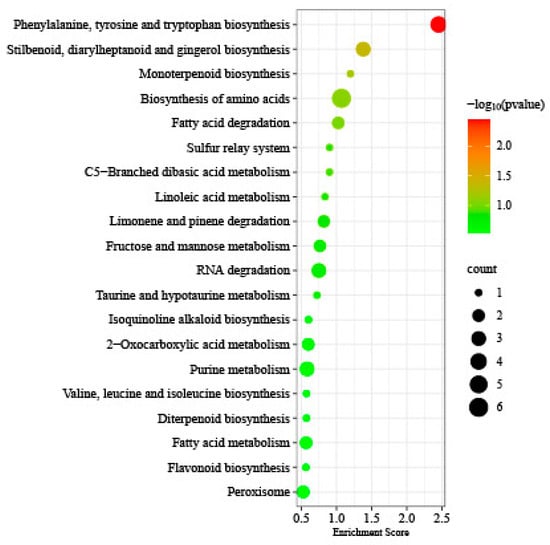
Figure 9.
KEGG pathway enrichment in between DMGs and DEGs under T1 and T2 freezing stress.
4. Discussion
Freezing stress is the most serious cause of yield decline and geographical distribution for winter rapeseed [19]. It has always been the goal of researchers to breed freezing-resistant cultivars and dissect the mechanism of freezing resistance in winter rapeseed, but no cultivar has been able to grow north of 35 degrees latitude, and our understanding of these mechanisms remains limited. However, in recent years, our laboratory has made a breakthrough in the breeding of strong cold-resistant cultivars by crossing distant varieties with winter turnip rape (Brassica rapa) to introduce its cold-resistant traits into winter oilseed rape, enabling these cultivars to overwinter in northern China [3,5]. The plant’s morphology, overwintering rate, physiology and biochemistry, photosynthetic index, and submicroscopic structure can all reflect its cold-resistant characteristics [18]. For example, the leaf color of the freezing-resistant NS cultivar is darker green than that of the freezing-sensitive NF cultivar, and the leaf of the freezing-sensitive cultivar NF was more damaged by cold stress. After 12 h of freezing stress, the freezing-resistant cultivar can recover, but the freezing-sensitive cultivar cannot. The observed shrinkage of chloroplasts in the freezing-sensitive cultivar suggests impaired photosynthetic efficiency under freezing stress. In the field, the overwintering rate of the freezing-resistant cultivar was significantly higher than that of the freezing-sensitive cultivar. These results indicate that although responding to stress of different degrees, the freezing-resistant NS cultivar possesses strong cold-resistant characteristics in both field and plot experiments, which can be utilized in rapeseed production in northern China. DNA methylation, as a primary epigenetic modification, can regulate plant responses to various stresses. Therefore, understanding the genetic mechanism of DNA methylation in winter rapeseed coping with freezing stress will help elucidate the mechanism of freezing resistance [7]. Thus, by jointly analyzing the transcriptome, proteome, and methylome, we can further supplement and improve our understanding of the cold-resistant mechanism regulated by methylation.
4.1. Cold Signal Transduction in Rapeseed Under Freezing Stress
Cold signal transduction plays an important role in plant cold tolerance by regulating the expression of cold tolerance genes, and TFs have been reported to participate in conveying cold stress signals in many plants [20,21]. In response to cold stress, the ICE-CBF-COR signaling cascade is a core pathway that alleviates cold stress in plants and has been isolated and verified [22]. In addition, a large number of TFs have been identified to regulate plant freezing tolerance, such as AP2/ERF, MYB, bHLH, WRKY, bZIP, C3H, NAC, etc. [23]. In this study, we found that bHLH, AP2/ERF-ERF, MYB, NAC, and C2H2 were the most enriched TFs in the core DMGs and DEGs under freezing stress. However, one transcriptional regulatory factor (SET) was identified in the core DAPs after freezing stress. The result indicated that DNA methylation may be involved in gene transcription regulation.
Plant hormones were also critical to environmental signal transduction [24]. In many instances, plants respond to environmental stresses by producing amounts of ABA, BR, ETH, JA, and IAA, and interact with ROS and TFs to orchestrates the plant response to abiotic stress [13]. The study identified 70 and 63 genes from DMGs and DEGs, respectively, which were enriched in plant hormone signal transduction (ko04075). Among them, three genes were shared between DMGs and DEGs, encoding two-component response regulator ARR18 (ARR18), ethylene receptor 2 (ETR2), and two-component response regulator ARR12 isoform X2 (ARR12). Under T1 and T2 freezing stress, ARR18 and ERT2 were downregulated, while ARR2 was upregulated across the cultivars. The result of methylation showed the methylation regulation patterns of homologous genes or genes’ different regions were inconsistent. Previous studies have shown that the phosphatidylinositol signaling system is an important signal transduction pathway in response to drought stress, and it may interact with plant hormones [25]. The research indicated that 17 DMGs, 21 DEGs, and 2 DAPs were identified in the phosphatidylinositol signaling system. Phospholipase C (PLC), a lipolytic enzymes hydrolyzing the ester bond between glycerol and the phosphate group and forming a diacylgcerol (DAG) molecule and an I (1, 4, 5) P3, is a key enzyme in phosphoinositide metabolism, which is activated by cold stress [26]. In our study, four genes were identified to encode PLC in the core DEGs, and half of the PLC (PLC1 and PLC4) were upregulated in T1 and T2. The methylation data showed that PLC1 was hypermethylated in the downstream. The result indicated that the phosphatidylinositol signaling system also plays an essential role in the response to freezing stress of rapeseed. Calcium/calmodulin-like protein (CML), calmodulin (CaM), and calcium-dependent protein kinase (CDPK) are master players in the Ca2+ signaling pathway, which plays an important role in plant stress response [27,28]. The study showed that 13 CML, 8 CDPK, and 2 CAM were identified in T1 and T2 stress from core DEGs. It is noteworthy that the expression of these genes was upregulated after freezing stress. Meanwhile, 12 CDPK, 7CML, and 2 CaM were obtained in the core DMGs, and the methylation level changed significantly in the up/downstream or gene body of genes.
4.2. Osmotic Balance Plays an Essential Role in Rapeseed Under Freezing Stress
Soluble sugars play a crucial role as an osmotic regulator in maintaining the osmotic balance of plant cells under abiotic stress [29,30]. Previous studies have demonstrated that the soluble sugar content of freezing-resistant cultivars is higher than that of freezing-sensitive cultivars [18]. The study revealed a significant increase in soluble sugar content under freezing stress, with NS exhibiting higher levels than NF. Furthermore, metabolic pathways associated with soluble sugars, such as amino sugar and nucleotide sugar metabolism and starch and sucrose metabolism, were found to be enriched in the core DMGs, DEGs, and DAPs. However, no shared genes were identified among these three omics analyses. The research showed that 42 genes of core DEGs are significantly related to the starch and sucrose metabolism, and other 29 genes show enrichment in the amino sugar and nucleotide sugar metabolism. Notably, approximately 80% and 90% of these genes exhibited upregulation under T1 and T2 stress conditions, respectively. Meanwhile, three and one DAPs were found to be involved in amino sugar and nucleotide sugar metabolism and starch and sucrose metabolism, exhibiting upregulation under T2 stress. Methylation analysis identified of 46 DMGs associated with starch and sucrose metabolism, while another 30 DMGs was enriched in amino sugar and nucleotide sugar metabolism. In response to T1 freezing stress, the one gene encoding UDP-glucuronic acid decarboxylase (UXS), which is enriched in both amino sugar and nucleotide sugar metabolism, as well as starch and sucrose metabolism pathways, was identified. UXS functioned as an enzyme catalyzing the chemical reaction of UDP-D-glucuronate into UDP-D-xylose + CO2. This reaction represents the conversion of glucose into xylose during polysaccharide synthesis from monosaccharides. The OsUXS3 positively regulates Na+ ion toxicity tolerance under salt stress in rice [31]. In this study, we observed an up-regulation of the UXS gene and protein expression levels in NS compared to NF after T1 treatment, accompanied by hypomethylation within the gene body. Meanwhile, the BnaC01g40610D protein corresponds to β-Glu, which is enriched in starch and sucrose metabolism, as well as cyanoamino acid metabolism. The β-Glu is a rate-limiting enzyme that catalyzes the production of D-glucose from beta-D-glucoside, Cellobiose, and Cellodextrin in starch and sucrose metabolism [32]. Previous studies have shown that β-Glu can influence cell wall structure and beta-glucosidases hydrolyze inert precursors to release antioxidant substances under various abiotic stresses [33]. RNA-seq analysis revolved around a close association between β-Glu and cold resistance in poplars [34]. In kiwifruit, cold stress enhances the activities of β-Glu, with the freezing-tolerant genotype exhibiting higher expression levels of the β-Glu gene compared to the freezing-sensitive genotype [35]. In our study, both cultivars exhibited differential gene and protein abundances of β-Glu during freezing stress; however, after T2 treatment, these levels were down-regulated in NS compared to NF. Apart from SS, the SP and Pro are important osmoregulatory substances involved in maintaining osmotic balance under abiotic stress [36]. Consistent with previous findings on strong cold-resistant varieties during cold stress, we observed significantly increased SP and Pro content in NS compared to NF during freezing stress.
4.3. Redox and Folate Homeostasis Were Critical Under Freezing Stress in Rapeseed
Under the freezing stress, the balance of reactive oxygen species (ROS) is broken, leading to the activation of metabolism processes associated with plant oxidative stress, thereby enhancing plant resistance and mitigating the damage caused by freezing stress [37]. In plants, numerous enzymes form the enzymatic antioxidant defense system, which plays a crucial role in scavenging excess ROS and maintaining normal cellular redox homeostasis [38]. In this study, the activities of antioxidant enzymes (SOD, POD, and CAT) were significantly elevated in both cultivars under freezing stress, with the activity in NS being higher than that in NF. These findings align with those reported by Wei et al. [5], suggesting that the freezing-resistant cultivar exhibits robust antioxidant enzyme activity under freezing stress. Seventeen DEGs encode peroxidase/L-ascorbate peroxidase, and most of these genes are upregulated following T1 and T2 stress. Sixteen DMGs are enriched in peroxidase/glutathione peroxidase, and the methylation level in NS differs significantly from that in NF under freezing stress. Peroxisomes, specialized organelles with oxidoreductase functions, play a critical role in the removal of excess ROS [39]. In this study, 14 DEGs were enriched in the peroxisome pathway, with nearly half of these genes being upregulated after freezing stress. Additionally, methylation data revealed that 27 genes were annotated in the peroxisome pathway. GLPs are ubiquitous plant proteins that play a vital role in plant responses to various abiotic and biotic stresses [40]. In rice, the expression of the OsGLP gene was significantly increased under salt and drought stress [41]. The knockout of GLP 8–14 (OsGLP1) in the CRISPR/Cas9 system demonstrated the involvement of OsGLP1 in acclimation to UV-B radiation [42]. In transgenic potato plants, StGLP participates in hydrogen peroxide (H2O2) production and activates ROS scavenging signaling pathways, with regulated antioxidant and heat stress-responsive genes, thereby conferring heat stress tolerance [43]. Overexpression of ZmGLP1 in Arabidopsis enhanced resistance to pathogens [44]. Research showed that GLP2-4 gene expression was upregulated, while protein abundance was downregulated, and downstream methylation levels were increased.
GGH plays a crucial role in plant folate homeostasis, and folate is involved in various aspects of plant metabolism and development [45]. Previous research demonstrated that GGH protein activity increased in soybean plants under different abiotic stresses [46]. In soybean root, GGH protein expression was induced during the early response to compatible symbiotic bacteria [47]. Additionally, GGH expression was upregulated during induced systemic resistance in the presence of beneficial bacteria. Our findings indicate that the expression of the GGH gene and the abundance of the GHH protein were upregulated in T1, and the downstream methylation level was higher in NS compared to NF.
4.4. The Stability of the Membrane Structure and the Integrity of the Photosynthetic System Are Crucial for the Freezing-Resistant Stress in Rapeseed
Maintaining the stability of photosynthesis and the integrity of the photosynthetic system under stress can enhance a plant’s stress resistance [48]. The results indicated that although there was no significant difference in photosynthetic parameters between the two cultivars, the chloroplast structure of NS exhibited greater stability than that of NF during freezing stress. Additionally, 20, 21, and 3 core DEGs, DMGs, and DAPs were enriched in carbon fixation in photosynthetic organisms, but no shared genes were identified across the three omics. Furthermore, 11, 8, 2 core DEGs, DMGs, and DAPs were annotated in photosynthesis. Ferredoxin (Fd) is a water-soluble, low-molecular-weight, acidic electron transport protein that plays a crucial role in plant photosynthesis and is involved in the ROS scavenging systems [49,50]. The study demonstrated that the expression of the Fd gene was upregulated under the T1 and T2 stress conditions, while other genes were downregulated. Oxygen-evolving enhancer protein 2 (OEE2) belongs to the oxygen-evolving complex in photosystem II and is responsible for catalyzing the splitting of water [51]. Its mutation leads to the incomplete assembly of the PSII complex. OEE was upregulated in response to severe drought stress in wheatgrass and downregulated during salt stress in wheat [52,53]. In response to cold stress, the OEE was reduced in wucai, and it is speculated that this is related to wucai being a cold-resistant vegetable [54]. In this study, we found that OEE2 was also downregulated in NS compared to NF during freezing stress, and its upstream was hypomethylated. This result indicated OEE2 may be one of the genes involved in the response of PSII to cold, and its expression may be regulated by methylation. Meanwhile, the iron–sulfur cluster biosynthesis protein SUFB is essential for chlorophyll synthesis and plays pivotal roles in various metabolic processes, including photosynthesis and redox metabolism [55]. Our study revealed that the transcription level of SUFB was upregulated in NS compared to NF, while the gene body was hypermethylated. Here, the protein abundance of SUFB was down-regulated under T2 freezing stress. Membrane lipid metabolism and remodeling are crucial strategies for plants to cope with cold stress. Cold tolerance in plants is often associated with increased unsaturated fatty acid content in membranes, which enhances fluidity and prevents damage. Strong cold-tolerant cultivars exhibit higher membrane unsaturation and greater membrane stability [56,57]. In this study, 15, 9, and 4 core DEGs, DMGs, and DAPs, respectively, were enriched in the biosynthesis of unsaturated fatty acids. More than half of the 15 DEGs showed significantly higher expression in NS compared to NF under freezing stress. The identified cold-responsive genes or proteins suggest potential targets for breeding rapeseed varieties with improved cold resistance. Malondialdehyde (MDA), a final product of membrane lipid peroxidation, is used to measure the degree of oxidation damage to biological membranes, while relative electrolyte leakage (REL) measures the extent of electrolyte exosmosis [58]. Under freezing stress, the integrity of the membrane structure is positively correlated with freezing tolerance. Cold acclimation may change the membrane lipids, and the winter-hardy genes of cold-tolerant cultivars linked with unsaturated fatty acids in the wheat. The result showed that MDA and REL levels in the freezing-sensitive NF cultivar were significantly higher than those in the freezing-resistant NS cultivar. Meanwhile, the freezing-tolerant cultivars have a stronger ability to maintain the integrity of chloroplast structure under freezing stress. These findings indicate that the cell membranes of cold-tolerant cultivars have a higher degree of unsaturation and higher stability under freezing stress. At the same time, it is speculated that this characteristic is related to the long-term cold acclimation of winter rapeseed, and DNA methylation may be involved in the cold acclimation of rapeseed.
5. Conclusions
Under freezing stress, the freezing-resistant cultivar exhibited distinct characteristics in phenotype, overwintering rate, physiology and biochemistry parameters, photosynthetic indices, and submicroscopic structure features. Subsequently, we integrated methylome, transcriptome, and proteome analyses to uncover novel mechanisms underlying the response of rapeseed to freezing stress. The findings provide valuable insights for breeding programs aiming to enhance the cold tolerance of winter rapeseed by targeting key epigenetic regulators.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/agronomy15030739/s1. Table S1: Whole genome DNA bisulfite sequencing data in two rapeseed cultivars; Table S2: The RNA-seq data of two rapeseed cultivars under freezing stress; Table S3: Coefficient correlation between samples of transcriptome; Table S4: Statistical table of protein and peptide identification results; Table S5: Coefficient correlation between samples of proteome; Table S6: KEGG pathway enrichment of core DMGs; Table S7: KEGG pathway enrichment of core DEGs; Table S8: KEGG pathway enrichment of core DAPs; Table S9: Transcription factor identified within the core DMGs; Table S10: Transcription factor identified within the core DEGs.
Author Contributions
Conceptualization, Z.L. and G.Z.; Methodology, J.W. (Jiaping Wei) and H.T.; Software, Z.W. and H.T.; Validation, J.C. and X.D.; Formal Analysis, H.L.; Investigation, J.W. (Jinxiong Wang) and Y.W.; Resources, Z.L. and H.L.; Data Curation, G.Z. and X.D; Writing—Original Draft Preparation, G.Z. and J.W. (Jinxiong Wang); Writing—Review and Editing, J.W. (Jiaping Wei), Z.W. and J.C.; Visualization, H.L., Y.W. and Z.W.; Supervision, H.T.; Project Administration, J.W. (Jiaping Wei) and X.D; Funding Acquisition, Z.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Scientific Research Start-up Funds for the Gansu Province Science and Technology Major Project [22ZD6NA009], the Natural Science Foundation of China [32360520], the Natural Science Foundation of Gansu Province [23JRRA1408], the scientific and technological projects of Gansu Province [23CXJA0001], the Key Talent Projects of Gansu Province [2023RCXM33], the scientific and technological projects of Linzhi City [LZZX-04], and the Innovator’s Star project [2025CXZX].
Data Availability Statement
The original contributions presented in the study are publicly available. The sequenced transcriptome raw data and Whole-genome bisulfite sequencing raw data have been deposited to the SRA at NCBI with the accession number PRJNA685002. The mass spectrometry data have been submitted into a public iProX database with the accession number of IPX0002682000.
Acknowledgments
We are grateful to Guangzhou Genedenovo Biotechnology Co., Ltd. for assisting in sequencing and bioinformatics analysis.
Conflicts of Interest
The authors declare no conflicts interest.
References
- Wang, H. New-demand oriented oilseed rape industry developing strategy. Oil Crop Sci. 2018, 40, 613–617. [Google Scholar]
- Cheng, L.; Jie, H.; Bofeng, L.; Zhongchao, F.; Junpeng, L. Current situation, development difficulties and suggestions of chinese rape industry. J. China Agric. Univ. 2017, 22, 203–210. [Google Scholar]
- Liu, Z.; Dong, X.; Cao, X.; Xu, C.; Wei, J.; Zheng, G.; Wang, J.; Li, H.; Fang, X.; Wang, Y.; et al. QTL mapping for cold tolerance and higher overwintering survival rate in winter rapeseed (Brassica napus). J. Plant Physiol. 2022, 275, 153735. [Google Scholar] [CrossRef]
- Ding, Y.; Shi, Y.; Yang, S. Advances and challenges in uncovering cold tolerance regulatory mechanisms in plants. New Phytol. 2019, 222, 1690–1704. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Zheng, G.; Yu, X.; Liu, S.; Dong, X.; Cao, X.; Fang, X.; Li, H.; Jin, J.; Mi, W.; et al. Comparative Transcriptomics and Proteomics Analyses of Leaves Reveals a Freezing Stress-Responsive Molecular Network in Winter Rapeseed (Brassica rapa L.). Front. Plant Sci. 2021, 12, 664311. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Liu, Z.; Mi, W.; Xu, C.; Zou, Y.; Xu, M.; Zheng, G.; Fang, X.; Cui, X.; Dong, X.; et al. Analysis on the Adaptability of Northwand Planting of Brassica napus. Sci. Agric. Sin. 2020, 53, 4164–4176. [Google Scholar]
- Chang, Y.; Zhu, C.; Jiang, J.; Zhang, H.; Zhu, J.; Duan, C. Epigenetic regulation in plant abiotic stress responses. J. Integr. Plant Biol. 2020, 62, 563–580. [Google Scholar] [CrossRef]
- Zhang, M.; Zhang, X.; Guo, L.; Qi, T.; Liu, G.; Feng, J.; Shahzad, K.; Zhang, B.; Li, X.; Wang, H.; et al. Single-base resolution methylome of cotton cytoplasmic male sterility system reveals epigenomic changes in response to high-temperature stress during anther development. J. Exp. Bot. 2020, 71, 951–969. [Google Scholar] [CrossRef]
- Seymour, D.K.; Becker, C. The causes and consequences of DNA methylome variation in plants. Curr. Opin. Plant Biol. 2017, 36, 56–63. [Google Scholar] [CrossRef]
- Lämke, J.; Bäurle, I. Epigenetic and chromatin-based mechanisms in environmental stress adaptation and stress memory in plants. Genome Biol. 2017, 18, 124. [Google Scholar] [CrossRef]
- Ahmad, F.; Farman, K.; Waseem, M.; Rana, R.M.; Nawaz, M.A.; Rehman, H.M.; Abbas, T.; Baloch, F.S.; Akrem, A.; Huang, J.; et al. Genome-wide identification, classification, expression profiling and DNA methylation (5mC) analysis of stress-responsive ZFP transcription factors in rice (Oryza sativa L.). Gene 2019, 718, 144018. [Google Scholar] [CrossRef] [PubMed]
- Wei, T.; Li, R.; Huang, J.; Zhao, H.; Ge, R.; Wu, Q.; Mallano, A.I.; Wang, Y.; Li, F.; Deng, Y.; et al. Divergent DNA methylation contributes to duplicated gene evolution and chilling response in tea plants. Plant J. 2021, 106, 1312–1327. [Google Scholar] [CrossRef]
- Zheng, G.; Dong, X.; Wei, J.; Liu, Z.; Aslam, A.; Cui, J.; Li, H.; Wang, Y.; Tian, H.; Cao, X. Integrated methylome and transcriptome analysis unravel the cold tolerance mechanism in winter rapeseed (Brassica napus L.). BMC Plant Biol. 2022, 22, 414. [Google Scholar] [CrossRef]
- Wei, J.; Shen, Y.; Dong, X.; Zhu, Y.; Cui, J.; Li, H.; Zheng, G.; Tian, H.; Wang, Y.; Liu, Z. DNA methylation affects freezing tolerance in winter rapeseed by mediating the expression of genes related to JA and CK pathways. Front. Genet. 2022, 13, 968494. [Google Scholar] [CrossRef]
- Hassan, S.A.-Z.; Hesham, F.A.; Shah, F. Antioxidative Defense System, Hormones, and Metabolite Accumulation in Different Plant Parts of Two Contrasting Rice Cultivars as Influenced by Plant Growth Regulators Under Heat Stress. Front. Plant Sci. 2022, 13, 911846. [Google Scholar] [CrossRef]
- Rongmiao, H.; Lizhi, Y.; Tana, W.; Shiyao, C.; Lu, Z. Genes related to osmoregulation and antioxidation play important roles in the response of Trollius chinensis seedlings to saline-alkali stress. Front. Plant Sci. 2023, 14, 1080504. [Google Scholar] [CrossRef]
- Wei, J.; Zheng, G.; Dong, X.; Li, H.; Liu, S.; Wang, Y.; Liu, Z. Integration of transcriptome and proteome analysis reveals the mechanism of freezing tolerance in winter rapeseed. Plant Growth Regul. 2022, 96, 103–118. [Google Scholar] [CrossRef]
- Mi, W.; Liu, Z.; Jin, J.; Dong, X.; Xu, C.; Zou, Y.; Xu, M.; Zheng, G.; Cao, X.; Fang, X.; et al. Comparative proteomics analysis reveals the molecular mechanism of enhanced cold tolerance through ROS scavenging in winter rapeseed (Brassica napus L.). PLoS ONE 2021, 16, e0243292. [Google Scholar] [CrossRef]
- Xu, Y.; Zeng, X.; Wu, J.; Zhang, F.; Li, C.; Jiang, J.; Wang, Y.; Sun, W. iTRAQ-Based Quantitative Proteome Revealed Metabolic Changes in Winter Turnip Rape (Brassica rapa L.) under Cold Stress. Int. J. Mol. Sci. 2018, 19, 3346. [Google Scholar] [CrossRef]
- Gusain, S.; Joshi, S.; Joshi, R. Sensing, signalling, and regulatory mechanism of cold-stress tolerance in plants. Plant Physiol. Biochem. PPB 2023, 197, 107646. [Google Scholar] [CrossRef]
- Zhu, J. Abiotic Stress Signaling and Responses in Plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef]
- Hwarari, D.; Guan, Y.; Ahmad, B.; Movahedi, A.; Tian, M.; Hao, Z.; Lu, Y.; Chen, J.; Yang, L. ICE-CBF-COR Signaling Cascade and Its Regulation in Plants Responding to Cold Stress. Int. J. Mol. Sci. 2022, 23, 1549. [Google Scholar] [CrossRef]
- Waititu, J.K.; Cai, Q.; Sun, Y.; Sun, Y.; Li, C.; Zhang, C.; Liu, J.; Wang, H. Transcriptome Profiling of Maize (Zea mays L.) Leaves Reveals Key Cold-Responsive Genes, Transcription Factors, and Metabolic Pathways Regulating Cold Stress Tolerance at the Seedling Stage. Genes 2021, 12, 1638. [Google Scholar] [CrossRef]
- Waadt, R.; Seller, C.A.; Hsu, P.K.; Takahashi, Y.; Munemasa, S.; Schroeder, J.I. Plant hormone regulation of abiotic stress responses. Nat. Rev. Mol. Cell Biol. 2022, 23, 680–694. [Google Scholar] [CrossRef]
- Wang, X.; Deng, Y.; Gao, L.; Kong, F.; Shen, G.; Duan, B.; Wang, Z.; Mei, D.; Han, Z. Series-temporal transcriptome profiling of cotton reveals the response mechanism of phosphatidylinositol signaling system in the early stage of drought stress. Genomics 2022, 114, 110465. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Gu, J.; Cui, X.; Fu, H.; Wang, F.; Qi, M.; Sun, Z.; Li, T.; Liu, Y. Genome-wide investigation of the phospholipase C gene family in Solanum lycopersicum and abiotic stress analysis. Environ. Exp. Bot. 2023, 210, 105336. [Google Scholar] [CrossRef]
- Yuan, P.; Yang, T.; Poovaiah, B.W. Calcium Signaling-Mediated Plant Response to Cold Stress. Int. J. Mol. Sci. 2018, 19, 3896. [Google Scholar] [CrossRef] [PubMed]
- Zeng, H.; Zhu, Q.; Yuan, P.; Yan, Y.; Yi, K.; Du, L. Calmodulin and calmodulin-like protein-mediated plant responses to biotic stresses. Plant Cell Environ. 2023, 46, 3680–3703. [Google Scholar] [CrossRef]
- Tarkowski, Ł.P.; Van den Ende, W. Cold tolerance triggered by soluble sugars: A multifaceted countermeasure. Front. Plant Sci. 2015, 6, 203. [Google Scholar] [CrossRef]
- Li, C.; Wan, Y.; Shang, X.; Fang, S. Integration of transcriptomic and metabolomic analysis unveils the response mechanism of sugar metabolism in Cyclocarya paliurus seedlings subjected to PEG-induced drought stress. Plant Physiol. Biochem. PPB 2023, 201, 107856. [Google Scholar] [CrossRef]
- Shen, T.; Li, K.; Yan, R.; Xu, F.; Ni, L.; Jiang, M. The UDP-glucuronic acid decarboxylase OsUXS3 regulates Na + ion toxicity tolerance under salt stress by interacting with OsCATs in rice. Plant Physiol. Biochem. PPB 2023, 196, 850–858. [Google Scholar] [CrossRef] [PubMed]
- Khairudin, N.B.A.; Mazlan, N.S.F. Molecular docking study of Beta-glucosidase with cellobiose, cellotetraose and cellotetriose. Bioinformation 2013, 9, 813–817. [Google Scholar] [CrossRef]
- James, R.K.C.; Karunambigai, A.; Jong-Seong, J.; Jisnuson, S. Functions of rice beta-glucosidases and transglucosidases. Scienceasia 2023, 49, 635. [Google Scholar] [CrossRef]
- Yang, C.; Li, X.; Zhang, Y.; Jiang, H. Transcriptome analysis of Populus × canadensis ’Zhongliao1’ in response to low temperature stress. BMC Genom. 2023, 24, 77. [Google Scholar] [CrossRef]
- Sun, S.; Lin, M.; Qi, X.; Chen, J.; Gu, H.; Zhong, Y.; Sun, L.; Abid, M.; Bai, D.; Hu, C.; et al. Full-length transcriptome profiling reveals insight into the cold response of two kiwifruit genotypes (A. arguta) with contrasting freezing tolerances. BMC Plant Biol. 2021, 21, 365. [Google Scholar] [CrossRef]
- María Elena, A.; Arnould, S.; László, S. Proline metabolism as regulatory hub. Trends Plant Sci. 2022, 27, 39–55. [Google Scholar] [CrossRef]
- Baxter, A.J.; Mittler, R.; Suzuki, N. ROS as key players in plant stress signalling. J. Exp. Bot. 2014, 65, 1229–1240. [Google Scholar] [CrossRef]
- Xu, D.; Hu, M.; Wang, Y.; Cui, Y. Antioxidant Activities of Quercetin and Its Complexes for Medicinal Application. Molecules 2019, 24, 1123. [Google Scholar] [CrossRef]
- Wei, M.; Xu, L.; Du, Z.; Wang, F.; Zhang, R.; Song, X.; Lam, S.; Shui, G.; Li, Y.; Chye, M.-L. RICE ACYL-COA-BINDING PROTEIN6 Affects Acyl-CoA Homeostasis and Growth in Rice. Rice 2020, 13, 75. [Google Scholar] [CrossRef]
- Zaynab, M.; Peng, J.; Sharif, Y.; Fatima, M.; Albaqami, M.; Al-Yahyai, R.; Raza, A.; Khan, K.A.; Alotaibi, S.S.; Alaraidh, L.A.; et al. Genome-Wide Identification and Expression Profiling of Germin-Like Proteins Reveal Their Role in Regulating Abiotic Stress Response in Potato. Front. Plant Sci. 2021, 12, 831140. [Google Scholar] [CrossRef]
- Anum, J.; O’Shea, C.; Zeeshan Hyder, M.; Farrukh, S.; Skriver, K.; Malik, S.I.; Yasmin, T. Germin like protein genes exhibit modular expression during salt and drought stress in elite rice cultivars. Mol. Biol. Rep. 2022, 49, 293–302. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Tao, M.; Leung, D.W.M.; Yan, X.; Chen, L.; Peng, X.; Liu, E.-E. The rice germin-like protein OsGLP1 participates in acclimation to UV-B radiation. Plant Physiol. 2021, 186, 1254–1268. [Google Scholar] [CrossRef]
- Gangadhar, B.H.; Mishra, R.K.; Kappachery, S.; Venkidasamy, B.; Venkatesh, J.; Nookaraju, A.; Thiruvengadam, M. Enhanced thermo-tolerance in transgenic potato (Solanum tuberosum L.) overexpressing hydrogen peroxide-producing germin-like protein (GLP). Genomics 2021, 113, 3224–3234. [Google Scholar] [CrossRef] [PubMed]
- Mao, L.; Ge, L.; Ye, X.; Xu, L.; Si, W.; Ding, T. ZmGLP1, a Germin-like Protein from Maize, Plays an Important Role in the Regulation of Pathogen Resistance. Int. J. Mol. Sci. 2022, 23, 14316. [Google Scholar] [CrossRef] [PubMed]
- Tariq, A.A.; Giuseppe, O.; Payam, M.; Aurora, L.N.; Malcolm, J.B.; Jesse, F.G.; Andrew, D.H. A central role for gamma-glutamyl hydrolases in plant folate homeostasis. Plant J. 2010, 64, 256–266. [Google Scholar] [CrossRef]
- Vaishnav, A.; Kumari, S.; Jain, S.; Varma, A.; Choudhary, D.K. Putative bacterial volatile-mediated growth in soybean (Glycine max L. Merrill) and expression of induced proteins under salt stress. J. Appl. Microbiol. 2015, 119, 539–551. [Google Scholar] [CrossRef]
- Salavati, A.; Bushehri, A.A.; Taleei, A.; Hiraga, S.; Komatsu, S. A comparative proteomic analysis of the early response to compatible symbiotic bacteria in the roots of a supernodulating soybean variety. J. Proteom. 2012, 75, 819–832. [Google Scholar] [CrossRef]
- Wei, X.; Tana, W.; Chen, J.; Yu, S.; Zhang, X.; Zhang, L. Responses of Trollius chinensis to drought stress and rehydration: From photosynthetic physiology to gene expression. Plant Physiol. Biochem. PPB 2023, 201, 107841. [Google Scholar] [CrossRef]
- Grabsztunowicz, M.; Rantala, M.; Ivanauskaite, A.; Blomster, T.; Koskela, M.M.; Vuorinen, K.; Tyystjärvi, E.; Burow, M.; Overmyer, K.; Mähönen, A.P.; et al. Root-type ferredoxin-NADP + oxidoreductase isoforms in Arabidopsis thaliana: Expression patterns, location and stress responses. Plant Cell Environ. 2021, 44, 548–558. [Google Scholar] [CrossRef]
- Lin, Y.; Huang, L.; Hase, T.; Huang, H.-T.D.; Feng, T. Expression of plant ferredoxin-like protein (PFLP) enhances tolerance to heat stress in Arabidopsis thaliana. New Biotechnol. 2015, 32, 235–242. [Google Scholar] [CrossRef]
- Shuai, W.; Qiuping, L.; Jianfeng, W.; Yan, Y.; Guoliang, Z.; Huifei, Z.; Jiajie, W.; Feng, C.; Xiaojie, W.; Zhensheng, K.; et al. YR36/WKS1-Mediated Phosphorylation of PsbO, an Extrinsic Member of Photosystem II, Inhibits Photosynthesis and Confers Stripe Rust Resistance in Wheat. Mol. Plant 2019, 12, 1639–1650. [Google Scholar] [CrossRef]
- Gao, L.; Xing, Y.; Li, X.; Guo, G.; Hu, Y.; Ma, W.; Yan, Y. Proteome analysis of wheat leaf under salt stress by two-dimensional difference gel electrophoresis (2D-DIGE). Phytochemistry 2011, 72, 1180–1191. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Bian, B.; Zhang, M.; Wang, C.; Li, C.; Liao, W. The role and proteomic analysis of ethylene in hydrogen gas-induced adventitious rooting development in cucumber (Cucumis sativus L.) explants. PeerJ 2020, 8, e8896. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Zhang, L.; Wu, Y.; Zheng, Y.; Nie, L.; Zhang, S.; Lan, T.; Zhao, Y.; Zhu, S.; Hou, J.; et al. Comparative transcriptome analysis reveals that chlorophyll metabolism contributes to leaf color changes in wucai (Brassica campestris L.) in response to cold. BMC Plant Biol. 2021, 21, 438. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Page, M.T.; Sumida, A.; Tanaka, A.; Terry, M.J.; Tanaka, R. The iron-sulfur cluster biosynthesis protein SUFB is required for chlorophyll synthesis, but not phytochrome signaling. Plant J. 2017, 89, 1184–1194. [Google Scholar] [CrossRef]
- Zhao, X.; Wei, Y.; Zhang, J.; Yang, L.; Liu, X.; Zhang, H.; Shao, W.; He, L.; Li, Z.; Zhang, Y.; et al. Membrane Lipids’ Metabolism and Transcriptional Regulation in Maize Roots Under Cold Stress. Front. Plant Sci. 2021, 12, 639132. [Google Scholar] [CrossRef]
- Mendoza, D.D. Temperature sensing by membranes. Annu. Rev. Microbiol. 2014, 68, 101–116. [Google Scholar] [CrossRef]
- Takahashi, D.; Uemura, M.; Kawamura, Y. Freezing Tolerance of Plant Cells: From the Aspect of Plasma Membrane and Microdomain. Adv. Exp. Med. Biol. 2018, 1081, 61–79. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).