
Barley Leaf Area and Leaf Growth Rates Are Maximized during the Pre-Anthesis Phase


Abstract
:1. Introduction
2. Results
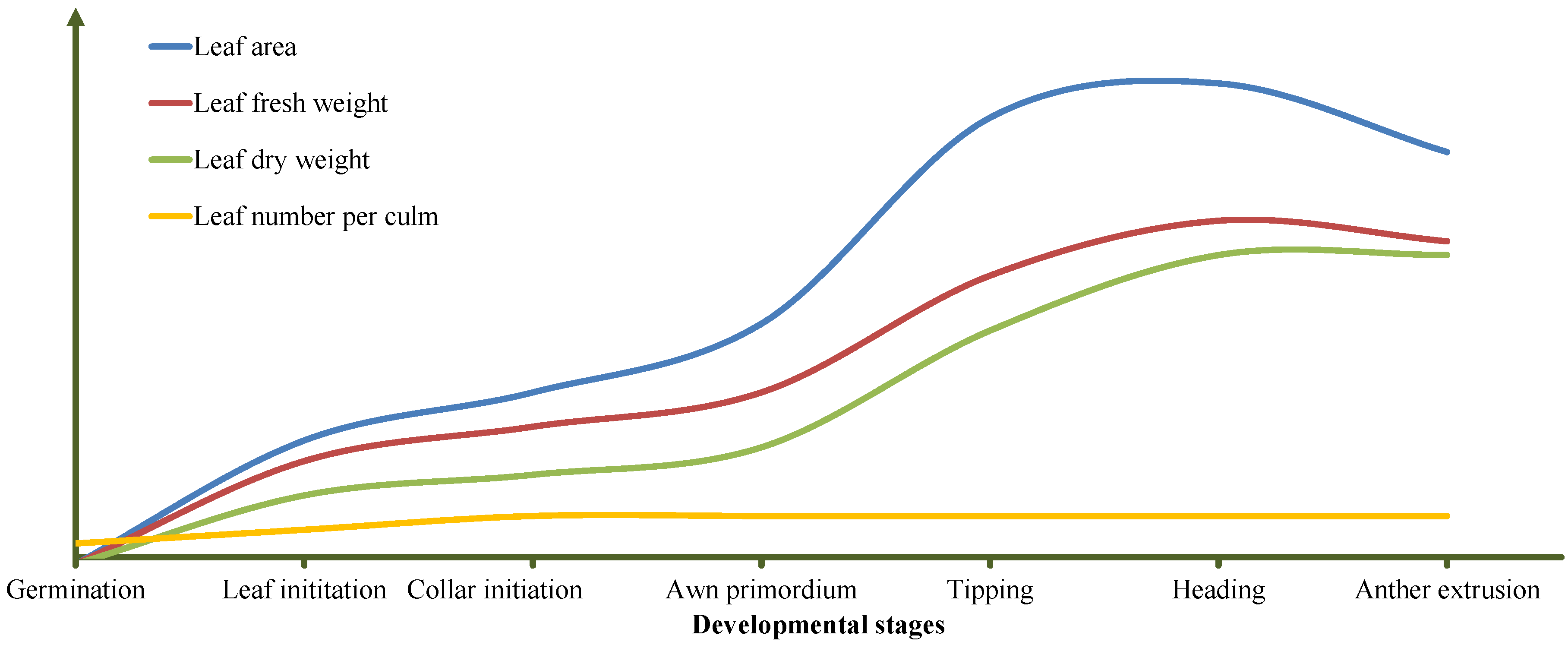
2.1. Correlation Analysis between Thermal Time and Leaf Trait
2.2. Analysis of Phenotypic Leaf Variation

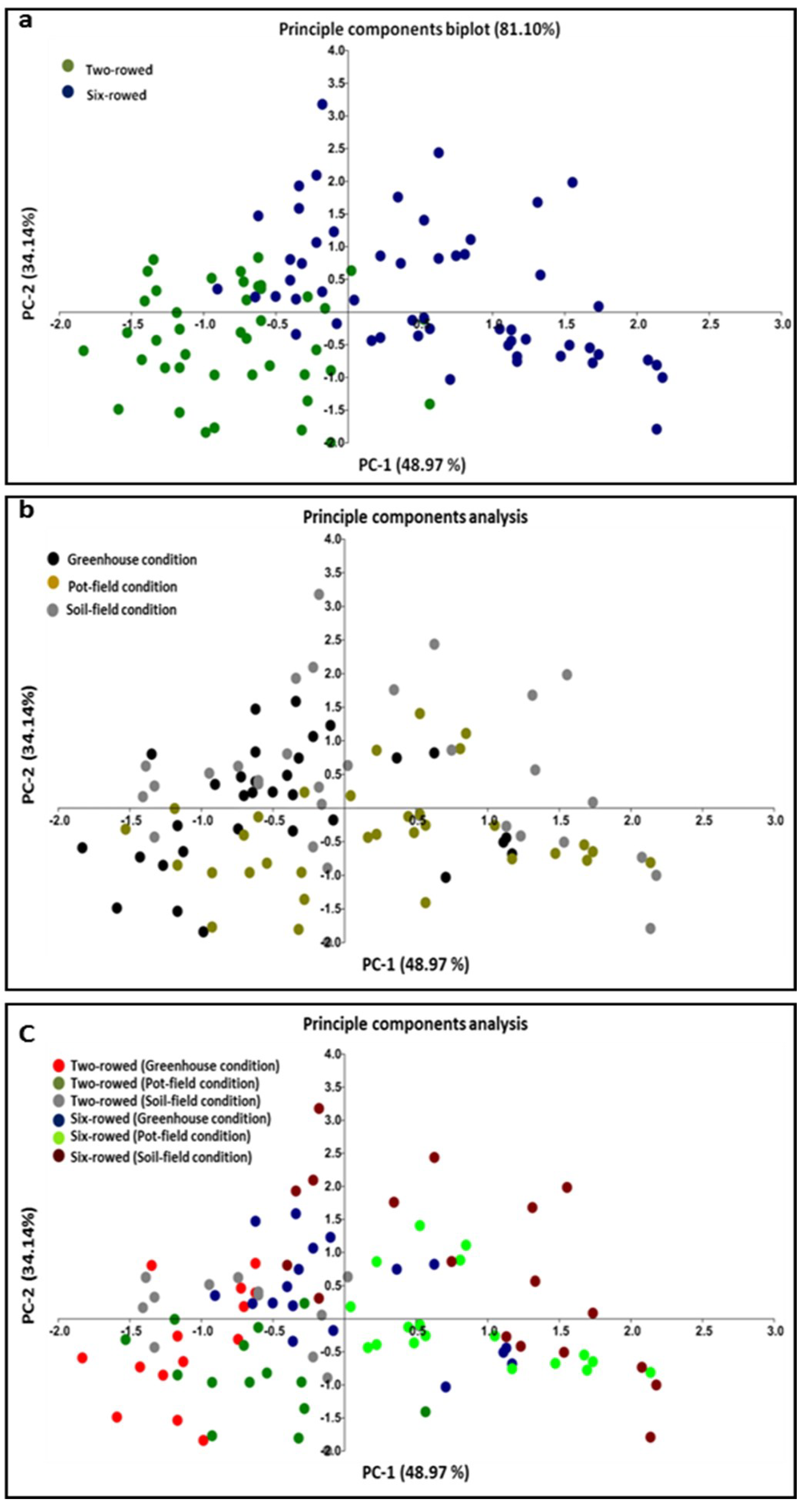{kind=link}
{kind=link}
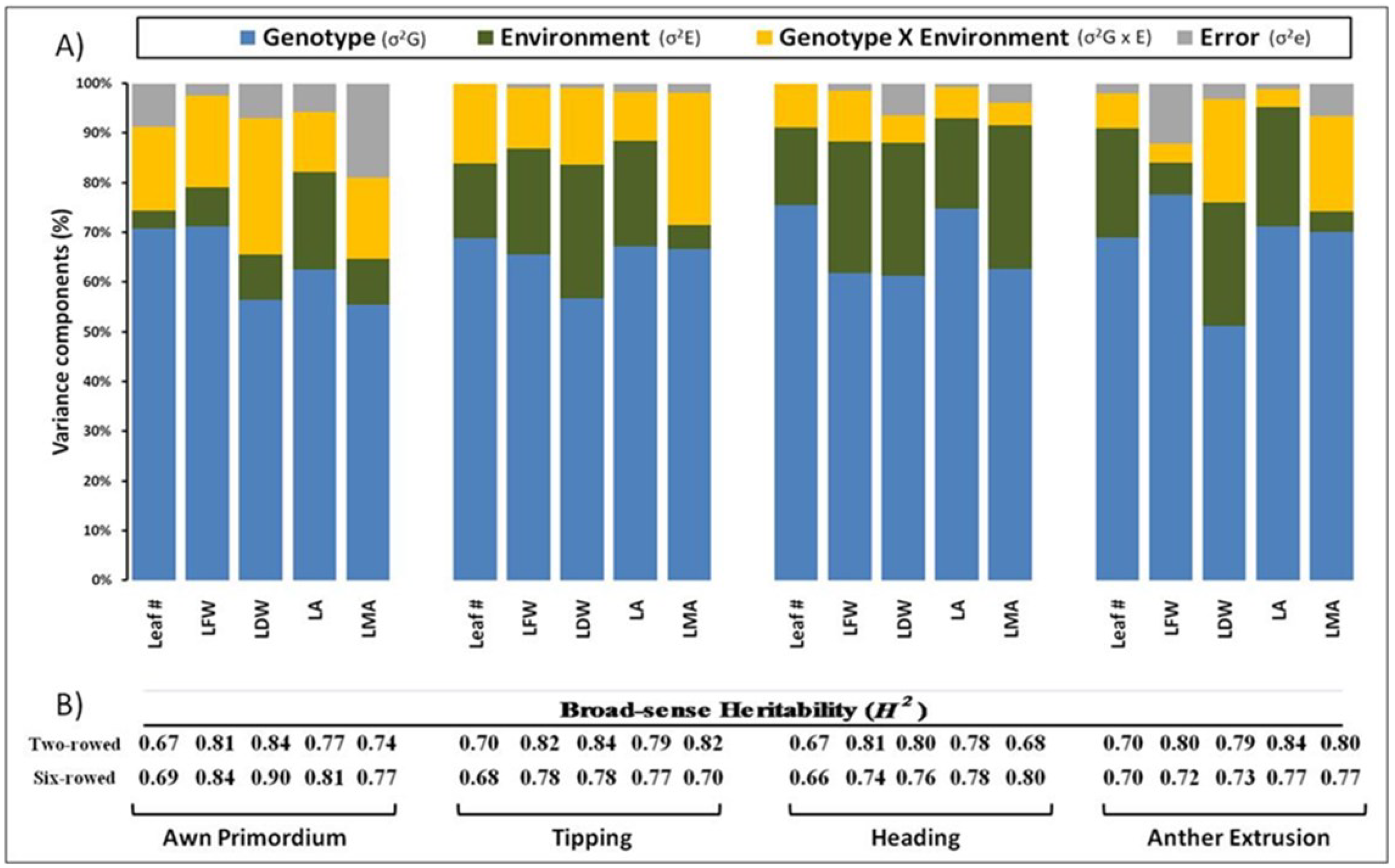
{kind=link}
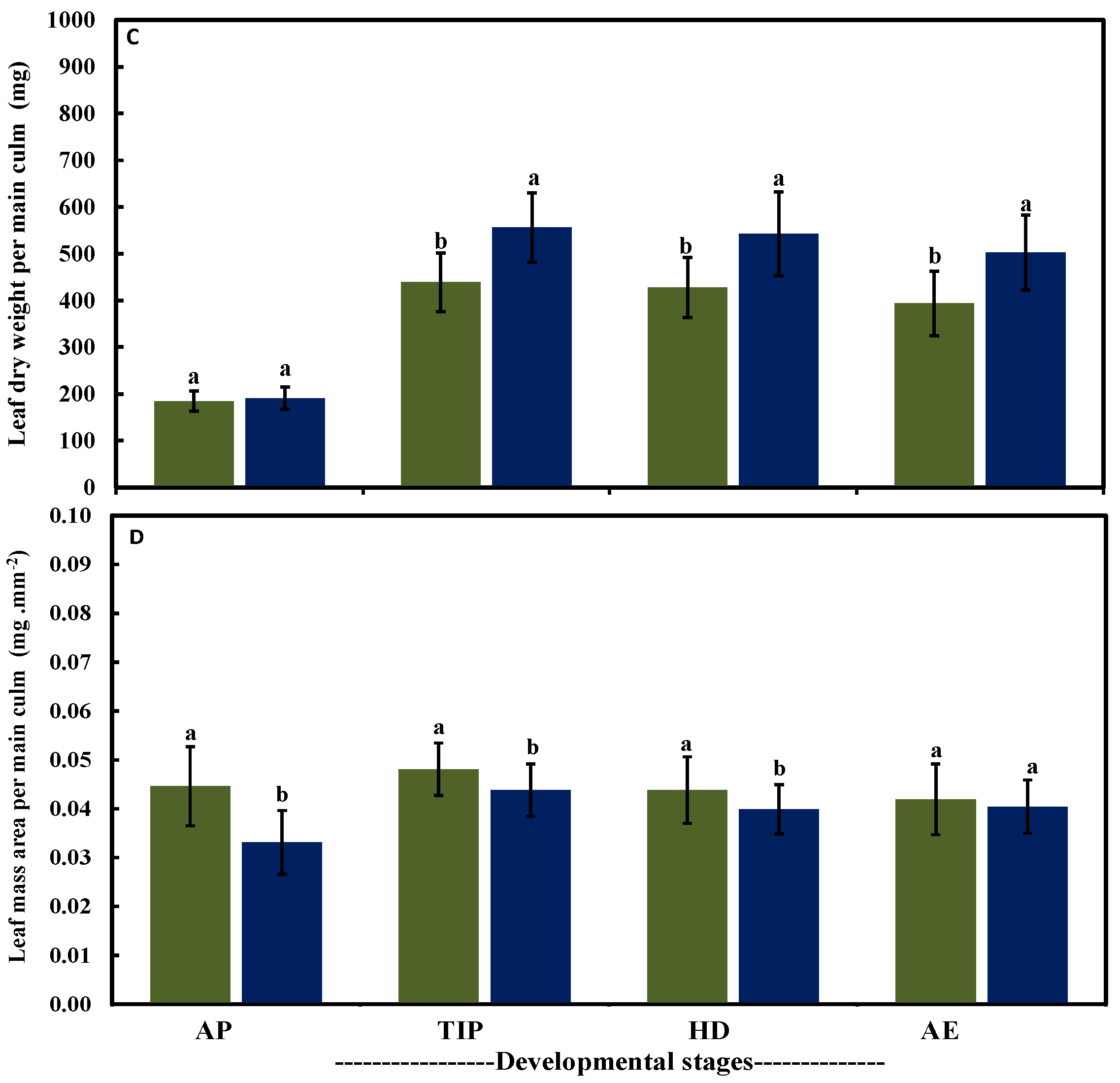{kind=link}
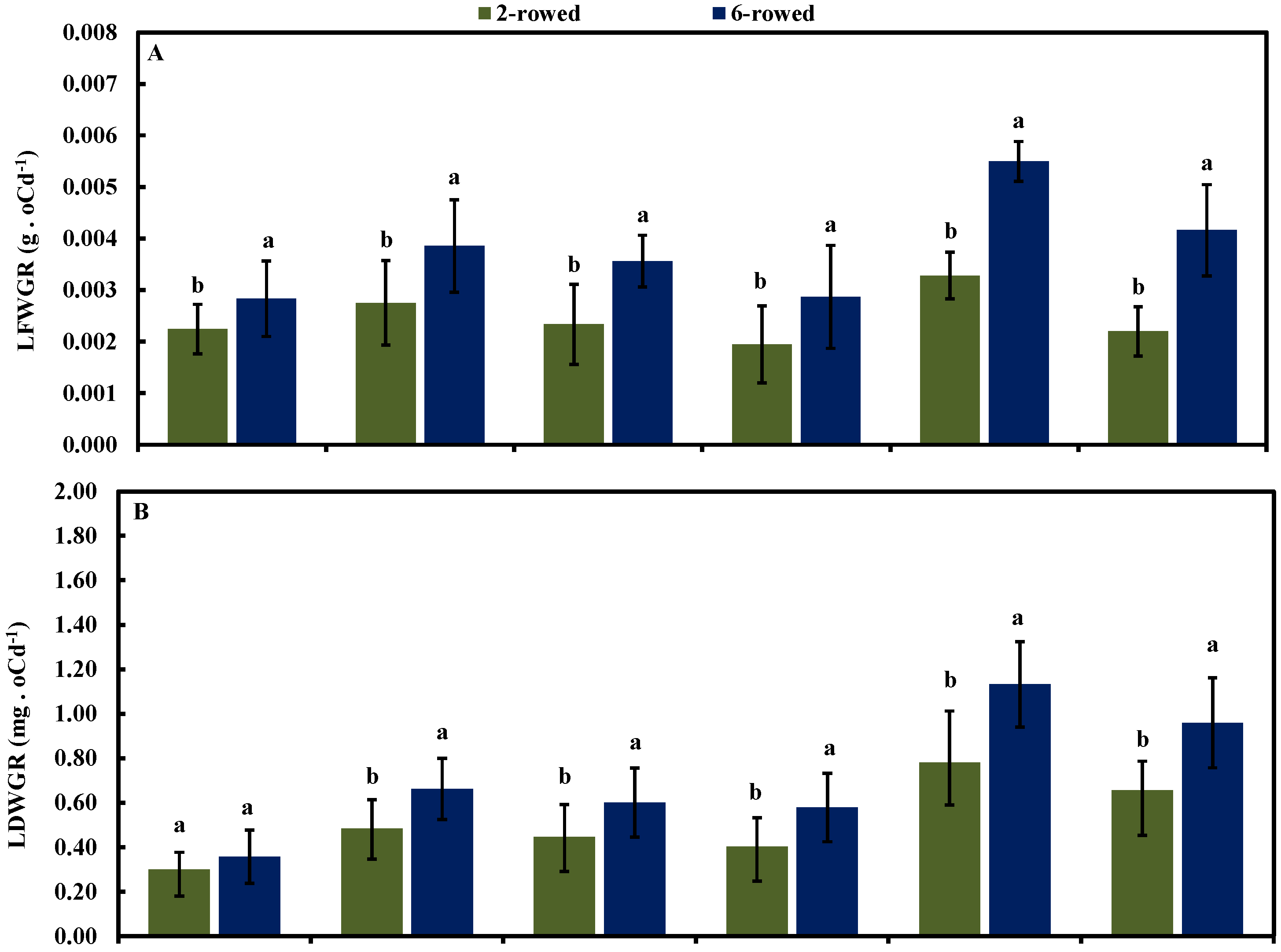{kind=link}
{kind=link}
{kind=link}
| Leaf Traits | Thermal Time/Growing Degree-Day (GDD) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Growing Condition | Awn Primordium | Tipping | Heading | Anther Extrusion | |||||
| Two-Rowed | Six-Rowed | Two-Rowed | Six-Rowed | Two-Rowed | Six-Rowed | Two-Rowed | Six-Rowed | ||
| Leaf number per main culm | GH * | 0.34 | 0.69 | −0.21 | −0.27 | −0.27 | −0.20 | 0.06 | −0.17 |
| Pots | 0.10 | −0.23 | 0.45 | 0.46 | 0.56 | 0.45 | −0.19 | 0.56 | |
| Soil | 0.07 | 0.20 | 0.65 | 0.48 | 0.63 | 0.48 | 0.35 | 0.42 | |
| Leaf fresh weight per main culm (g, LFW) | GH | 0.67 | 0.68 | 0.70 | 0.64 | 0.36 | 0.32 | 0.36 | 0.18 |
| Pots | 0.70 | 0.59 | 0.69 | 0.75 | 0.52 | 0.77 | 0.22 | 0.71 | |
| Soil | 0.40 | 0.47 | 0.49 | 0.74 | 0.43 | 0.24 | 0.32 | 0.23 | |
| Leaf dry weight per main culm (mg, LDW) | GH | 0.52 | 0.65 | 0.68 | 0.55 | 0.68 | 0.48 | 0.65 | 0.46 |
| Pots | 0.76 | 0.81 | 0.75 | 0.62 | 0.55 | 0.77 | 0.30 | 0.76 | |
| Soil | 0.52 | 0.49 | 0.46 | 0.70 | 0.71 | 0.48 | 0.39 | 0.29 | |
| Leaf area per main culm (mm, LA) | GH | 0.40 | 0.37 | 0.23 | −0.22 | 0.30 | 0.20 | 0.42 | −0.09 |
| Pots | 0.59 | 0.60 | 0.67 | 0.79 | 0.40 | 0.84 | 0.43 | 0.75 | |
| Soil | 0.40 | 0.27 | 0.52 | 0.70 | 0.53 | 0.48 | 0.72 | 0.42 | |
| Leaf mass area per main culm (mg·mm−2, LMA) | GH | 0.30 | 0.25 | 0.28 | −0.32 | −0.32 | −0.24 | −0.24 | −0.42 |
| Pots | 0.49 | 0.72 | 0.28 | 0.11 | 0.31 | −0.01 | 0.82 | 0.40 | |
| Soil | 0.46 | 0.50 | 0.05 | −0.22 | −0.11 | 0.20 | −0.47 | 0.02 | |


2.3. Leaf Traits in Both Row-Type Classes under Different Growth Conditions at Four Developmental Stages




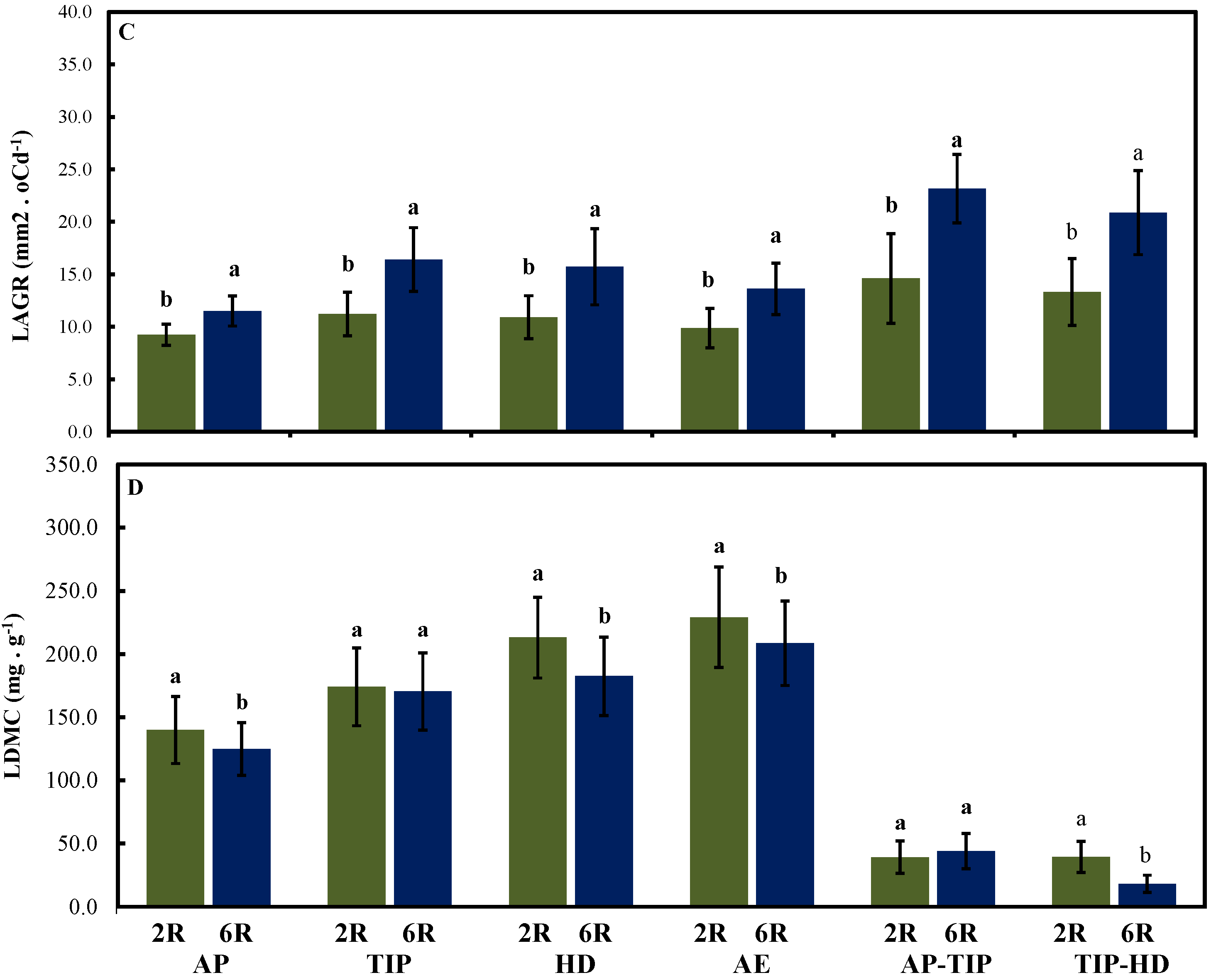
2.4. Leaf Growth Rate in Both Row-Type Classes under Different Growth conditions at Four Developmental Stages
2.5. Correlation Analysis between Leaf Area (LA), Single Plant Yield, and Yield Components
3. Discussion
3.1. The Importance of Leaf Area in Improving Single-Plant Yield
3.1.1. Large Leaf Area
| Yield Components | Leaf Blade Area per Main Culm (mm2) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Growing Condition | Awn Primordium | Tipping | Heading | Anther Extrusion | |||||
| Two-Rowed | Six-Rowed | Two-Rowed | Six-Rowed | Two-Rowed | Six-Rowed | Two-Rowed | Six-Rowed | ||
| Grain number per main culm spike at harvest | GH ‡ | 0.13 | 0.20 | 0.14 | 0.28 | 0.37 * | 0.52 * | 0.43 * | 0.62 * |
| Pots | 0.57 * | 0.55 * | 0.45 * | 0.56 * | 0.53 * | 0.64 * | 0.60 * | 0.74 * | |
| Soil | 0.21 | 0.38 * | 0.51 * | 0.61 * | 0.53 * | 0.62 * | 0.58 * | 0.65 * | |
| Grain weight main culm spike at harvest (g) | GH | 0.12 | 0.13 | 0.26 | 0.30 | 0.23 | 0.40 * | 0.33 | 0.46 * |
| Pots | 0.53 * | 0.40 * | 0.40 * | 0.49 * | 0.35 * | 0.50 * | 0.50 * | 0.55 * | |
| Soil | 0.28 | 0.34 | 0.42 * | 0.62 * | 0.47 * | 0.53 * | 0.55 * | 0.68 * | |
| Main culm spike dry weight at heading (g) | GH | 0.16 | 0.10 | 0.35 * | 0.34 | 0.31 | 0.24 | 0.33 | 0.36 * |
| Pots | 0.10 | 0.36 * | 0.37 * | 0.79 * | 0.43 * | 0.57 * | 0.65 * | 0.83 * | |
| Soil | 0.31 | 0.10 | 0.56 * | 0.69 * | 0.43 * | 0.54 * | 0.74 * | 0.74 * | |
| Tillers per plant | GH | 0.31 | −0.07 | 0.01 | −0.17 | 0.47 * | −0.33 | 0.61 * | −0.36 * |
| Pots | 0.23 | −0.26 | 0.20 | −0.10 | 0.15 | 0.00 | 0.45 * | −0.09 | |
| Soil | 0.03 | −0.17 | 0.28 | −0.24 | 0.39 * | −0.38 * | 0.45 * | −0.31 | |
| Spikes per plant | GH | 0.25 | −0.06 | 0.30 | −0.54 * | 0.43 * | −0.34 | 0.51 * | −0.27 |
| Pots | 0.22 | −0.30 | 0.20 | −0.23 | 0.38 * | −0.22 | 0.47 * | 0.00 | |
| Soil | 0.16 | −0.22 | 0.29 | −0.22 | 0.33 | −0.40 * | 0.40 * | −0.31 | |
| Biological yield (g) | GH | 0.00 | −0.38 * | 0.27 | −0.32 | 0.29 | −0.21 | 0.39 * | −0.11 |
| Pots | 0.46 * | −0.08 | 0.47 * | −0.07 | 0.36 * | −0.13 | 0.47 * | −0.31 | |
| Soil | 0.20 | −0.29 | 0.42 * | −0.29 | 0.52 * | −0.27 | 0.65 * | −0.52 * | |
| Harvest Index (%) | GH | −0.26 | 0.38 * | −0.14 | 0.37 * | −0.29 | 0.12 | −0.17 | 0.20 |
| Pots | −0.35 * | 0.29 | −0.22 | 0.16 | −0.39 * | 0.21 | −0.17 | 0.28 | |
| Soil | −0.22 | −0.06 | −0.12 | 0.22 | −0.31 | 0.54 * | −0.30 | 0.41 * | |
3.1.2. Small Leaf Area
3.2. Maximized Leaf Growth Rate in the Two Row-Type Classes

3.3. Genetic Background and Variance Analysis
4. Conclusions
5. Materials and Methods
5.1. Plant Material
5.2. Growth Environment and Experimental Procedure
| Month | Field | Greenhouse ‡ | ||||
|---|---|---|---|---|---|---|
| Temperature °C | Rainfall (Mm) | Relative Humidity (%) | Global Solar Radiation (W/M2) | Temperature °C | Relative Humidity % | |
| April | 8.8 | 17.4 | 75.8 | 157 | 9.3 | 72.9 |
| May | 15.0 | 48.7 | 73.0 | 161 | 14.3 | 71.7 |
| Jun | 15.6 | 72.4 | 80.3 | 175 | 18.8 | 77.2 |
| July | 18.1 | 93.4 | 78.7 | 194 | 19.7 | 75.9 |
| August | 18.7 | 38.1 | 75.3 | 194 | 20.0 | 72.6 |
| No. | Name | Germplasm Status | Origin | Name | Germplasm Status | Origin |
|---|---|---|---|---|---|---|
| Six-Rowed | Two-Rowed | |||||
| 1 | BCC1453 | Cultivar | Finland | BCC1497 | Landrace | Kyrgyzstan |
| 2 | HOR2835 | Landrace | Iran | BCC1541 | Cultivar | Yugoslavia |
| 3 | BCC1494 | Landrace | Kazakhstan | BCC869 | Cultivar | Mexico |
| 4 | BCC579 | Cultivar | India | HOR8006 | Landrace | Turkey |
| 5 | BCC219 | Landrace | Tajikistan | Barke | Cultivar | Germany |
| 6 | BCC447 | Cultivar | China | BCC1566 | Landrace | Greece |
| 7 | BCC719 | Cultivar | Korea | BCC1589 | Landrace | Italy |
| 8 | Morex | Cultivar | USA | Triumph | Cultivar | Germany |
| 9 | BCC814 | Breeder line | USA | BCC801 | Cultivar | Canada |
| 10 | BCC818 | Cultivar | USA | Proctor | Cultivar | UK |
| 11 | BCC718 | Cultivar | Korea | BCC1370 | Cultivar | France |
| 12 | BCC551 | Cultivar | India | BCC1371 | Cultivar | France |
| 13 | BCC577 | Cultivar | India | BCC903 | Landrace | Afghanistan |
| 14 | BCC888 | Cultivar | Canada | Weeah | Cultivar | Australia |
| 15 | BCC942 | Cultivar | USA | |||
| 16 | BCC875 | Cultivar | USA | |||
| 17 | BCC921 | Cultivar | Colombia | |||
| 18 | BCC868 | Breeder line | Mexico | |||
5.3. Data Recording and Experimental Design
5.4. Data Analyses
Acknowledgments
Author Contributions
Abbreviations
| AP | Awn Primordium |
| AE | Anther Extrusion |
| BY | Biological Yield |
| CV | Coefficients of Variation |
| GDD or °C * D | Growing Degree Days or Thermal Time |
| GH | Greenhouse |
| HI | Harvest Index |
| HD | Heading |
| H2 | Broad-sense Heritability |
| LA | Leaf Area |
| LAGR | Leaf Area Growth Rate |
| LDMC | Leaf Dry Matter Content |
| LDW | Leaf Dry Weight |
| LDWGR | Leaf Dry Weight Growth Rate |
| LFWGR | Leaf Fresh Weight Growth Rate |
| LMA | Leaf Mass Area |
| MSDW | Main Spike Dry Weight |
| PCA | Principle component analysis |
| TIP | Tipping |
Conflicts of Interest
References
- Aase, J.K. Relationship between leaf area and dry-matter in winter-wheat. Agron. J. 1978, 70, 563–565. [Google Scholar] [CrossRef]
- Witkowski, E.T.F.; Lamont, B.B. Leaf specific mass confounds leaf density and thickness. Oecologia 1991, 88, 486–493. [Google Scholar] [CrossRef]
- Poorter, H.; Niinemets, U.; Poorter, L.; Wright, I.J.; Villar, R. Causes and consequences of variation in leaf mass per area (LMA): A meta-analysis. New Phytol. 2009, 182, 565–588. [Google Scholar] [CrossRef] [PubMed]
- Lambers, H.; Poorter, H. Inherent variation in growth-rate between higher-plants—A search for physiological causes and ecological consequences. Adv. Ecol. Res. 1992, 23, 187–261. [Google Scholar]
- Westoby, M.; Falster, D.S.; Moles, A.T.; Vesk, P.A.; Wright, I.J. Plant ecological strategies: Some leading dimensions of variation between species. Annu. Rev. Ecol. Syst. 2002, 33, 125–159. [Google Scholar] [CrossRef]
- Van Oosterom, E.J.; Acevedo, E. Leaf area and crop growth in relation to phenology of barley in mediterranean environments. Plant Soil 1993, 148, 223–237. [Google Scholar] [CrossRef]
- Tesarová, J.; Nátr, L. Phyllochron and winter barley leaf growth rate. Biol. Plant 1990, 32, 450–459. [Google Scholar] [CrossRef]
- Zhao, D.L.; Atlin, G.N.; Bastiaans, L.; Spiertz, J.H.J. Cultivar weed-competitiveness in aerobic rice: Heritability, correlated traits, and the potential for indirect selection in weed-free environments. Crop Sci. 2006, 46, 372–380. [Google Scholar] [CrossRef]
- Coleman, R.K.; Gill, G.S.; Rebetzke, G.J. Identification of quantitative trait loci for traits conferring weed competitiveness in wheat (Triticum aestivum L.). Aust. J. Agric. Res. 2001, 52, 1235–1246. [Google Scholar] [CrossRef]
- Jennings, P.R. Plant type as a rice breeding objective. Crop Sci. 1964, 4, 13–15. [Google Scholar] [CrossRef]
- Donald, C.M. The breeding of crop ideotypes. Euphytica 1968, 17, 385–403. [Google Scholar] [CrossRef]
- Peng, S.B.; Khush, G.S.; Virk, P.; Tang, Q.Y.; Zou, Y.B. Progress in ideotype breeding to increase rice yield potential. Field Crops Res. 2008, 108, 32–38. [Google Scholar] [CrossRef]
- Rasmusson, D.C. An evaluation of ideotype breeding. Crop Sci. 1987, 27, 1140–1146. [Google Scholar] [CrossRef]
- Yoshida, S. Physiological aspects of grain yield. Annu. Rev. Plant Physiol. 1972, 23, 437–464. [Google Scholar] [CrossRef]
- Fenta, B.A.; Beebe, S.E.; Kunert, K.J.; Burridge, J.D.; Barlow, K.M.; Lynch, J.P.; Foyer, C.H. Field phenotyping of soybean roots for drought stress tolerance. Agronomy 2014, 4, 418–435. [Google Scholar] [CrossRef]
- Bertholdsson, N.-O. Screening for barley waterlogging tolerance in nordic barley cultivars (Hordeum vulgare L.) using chlorophyll fluorescence on hydroponically-grown plants. Agronomy 2013, 3, 376–390. [Google Scholar] [CrossRef]
- Fujita, D.; Trijatmiko, K.R.; Tagle, A.G.; Sapasap, M.V.; Koide, Y.; Sasaki, K.; Tsakirpaloglou, N.; Gannaban, R.B.; Nishimura, T.; Yanagihara, S.; et al. Nal1 allele from a rice landrace greatly increases yield in modern indica cultivars. Proc. Natl. Acad. Sci. USA 2013, 110, 20431–20436. [Google Scholar]
- Khush, G. Breaking the yield frontier of rice. GeoJournal 1995, 35, 329–332. [Google Scholar] [CrossRef]
- Berdahl, J.D.; Rasmusson, D.C.; Moss, D.N. Effects of leaf area on photosynthetic rate, light penetration, and grain yield in barley. Crop Sci. 1972, 12, 177–180. [Google Scholar] [CrossRef]
- Gallagher, J.N.; Biscoe, P.V. Radiation absorption, growth and yield of cereals. J. Agric. Sci. 1978, 91, 47–60. [Google Scholar] [CrossRef]
- Jiang, D.; Fang, J.; Lou, L.; Zhao, J.; Yuan, S.; Yin, L.; Sun, W.; Peng, L.; Guo, B.; Li, X. Characterization of a null allelic mutant of the rice nal1 gene reveals its role in regulating cell division. PLoS ONE 2015, 10, e0118169. [Google Scholar] [CrossRef] [PubMed]
- Driever, S.M.; Lawson, T.; Andralojc, P.J.; Raines, C.A.; Parry, M.A.J. Natural variation in photosynthetic capacity, growth, and yield in 64 field-grown wheat genotypes. J. Exp. Bot. 2014, 65, 4959–4973. [Google Scholar] [CrossRef] [PubMed]
- Richards, R.A. Manipulation of leaf-area and its effect on grain-yield in droughted wheat. Aust. J. Agric. Res. 1983, 34, 23–31. [Google Scholar] [CrossRef]
- Jamieson, P.D.; Martin, R.J.; Francis, G.S.; Wilson, D.R. Drought effects on biomass production and radiation-use efficiency in barley. Field Crops Res. 1995, 43, 77–86. [Google Scholar] [CrossRef]
- Miralles, D.; Slafer, G. Radiation interception and radiation use efficiency of near-isogenic wheat lines with different height. Euphytica 1997, 97, 201–208. [Google Scholar] [CrossRef]
- Morgan, J.A.; LeCain, D.R.; Wells, R. Semidwarfing genes concentrate photosynthetic machinery and affect leaf gas exchange of wheat. Crop Sci. 1990, 30, 602–608. [Google Scholar]
- Reynolds, M.P.; van Ginkel, M.; Ribaut, J.M. Avenues for genetic modification of radiation use efficiency in wheat. J. Exp. Bot. 2000, 51, 459–473. [Google Scholar] [CrossRef] [PubMed]
- Kemanian, A.R.; Stöckle, C.O.; Huggins, D.R. Variability of barley radiation-use efficiency. Crop Sci. 2004, 44, 1662–1672. [Google Scholar] [CrossRef]
- Forster, B.P.; Franckowiak, J.D.; Lundqvist, U.; Lyon, J.; Pitkethly, I.; Thomas, W.T.B. The barley phytomer. Ann. Bot. 2007, 100, 725–733. [Google Scholar] [CrossRef] [PubMed]
- Alqudah, A.M.; Schnurbusch, T. Awn primordium to tipping is the most decisive developmental phase for spikelet survival in barley. Funct. Plant Biol. 2014, 41, 424–436. [Google Scholar] [CrossRef]
- Badr, A.; Muller, K.; Schafer-Pregl, R.; El Rabey, H.; Effgen, S.; Ibrahim, H.H.; Pozzi, C.; Rohde, W.; Salamini, F. On the origin and domestication history of barley (Hordeum vulgare L.). Mol. Biol. Evol. 2000, 17, 499–510. [Google Scholar] [CrossRef] [PubMed]
- Zohary, D.; Hopf, M. Domestication of Plants in the Old World; Oxford University Press: Oxford, UK, 2000. [Google Scholar]
- Komatsuda, T.; Pourkheirandish, M.; He, C.; Azhaguvel, P.; Kanamori, H.; Perovic, D.; Stein, N.; Graner, A.; Wicker, T.; Tagiri, A.; et al. Six-rowed barley originated from a mutation in a homeodomain-leucine zipper I-class homeobox gene. Proc. Natl. Acad. Sci. USA 2007, 104, 1424–1429. [Google Scholar] [CrossRef] [PubMed]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A decimal code for the growth stages of cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Curtis, E.M.; Leigh, A.; Rayburg, S. Relationships among leaf traits of Australian arid zone plants: Alternative modes of thermal protection. Aust. J. Bot. 2012, 60, 471–483. [Google Scholar] [CrossRef]
- Alqudah, A.M.; Sharma, R.; Pasam, R.K.; Graner, A.; Kilian, B.; Schnurbusch, T. Genetic dissection of photoperiod response based on gwas of pre-anthesis phase duration in spring barley. PLoS ONE 2014, 9, e113120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadras, V.O.; Denison, R.F. Do plant parts compete for resources? An evolutionary viewpoint. New Phytol. 2009, 183, 565–574. [Google Scholar] [CrossRef] [PubMed]
- Rebetzke, G.J.; Botwright, T.L.; Moore, C.S.; Richards, R.A.; Condon, A.G. Genotypic variation in specific leaf area for genetic improvement of early vigour in wheat. Field Crops Res. 2004, 88, 179–189. [Google Scholar] [CrossRef]
- Fasoula, D.A.; Fasoula, V.A. Competitive ability and plant breeding. In Plant Breeding Reviews; Janick, J., Ed.; John Wiley & Sons, Inc.: Oxford, UK, 1996; pp. 90–138. [Google Scholar]
- Baldissera, T.C.; Frak, E.; Carvalho, P.C.; Louarn, G. Plant development controls leaf area expansion in alfalfa plants competing for light. Ann. Bot. 2014, 113, 145–157. [Google Scholar] [CrossRef] [PubMed]
- Perez-Harguindeguy, N.; Diaz, S.; Garnier, E.; Lavorel, S.; Poorter, H.; Jaureguiberry, P.; Bret-Harte, M.S.; Cornwell, W.K.; Craine, J.M.; Gurvich, D.E.; et al. New handbook for standardised measurement of plant functional traits worldwide. Aust. J. Bot. 2013, 61, 167–234. [Google Scholar] [CrossRef]
- SAS. The Statistical Analysis Software (SAS) for Windows; Version 9.3; SAS Institute Inc: Cary, NC, USA, 2013. [Google Scholar]
- Snedecor, G.W.; Cochran, W.G. Statistical Methods, 7th ed.; Iowa State University Press: Ames, IA, USA, 1980. [Google Scholar]
- GenStat. Genstat for Windows; Version 16; VSN International: Hemel Hempstead, UK, 2013. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alqudah, A.M.; Schnurbusch, T. Barley Leaf Area and Leaf Growth Rates Are Maximized during the Pre-Anthesis Phase. Agronomy 2015, 5, 107-129. https://doi.org/10.3390/agronomy5020107
Alqudah AM, Schnurbusch T. Barley Leaf Area and Leaf Growth Rates Are Maximized during the Pre-Anthesis Phase. Agronomy. 2015; 5(2):107-129. https://doi.org/10.3390/agronomy5020107
Chicago/Turabian StyleAlqudah, Ahmad M., and Thorsten Schnurbusch. 2015. "Barley Leaf Area and Leaf Growth Rates Are Maximized during the Pre-Anthesis Phase" Agronomy 5, no. 2: 107-129. https://doi.org/10.3390/agronomy5020107