Artificial Seeds (Principle, Aspects and Applications)




Abstract
:1. Artificial Seeds Introduction and Definition
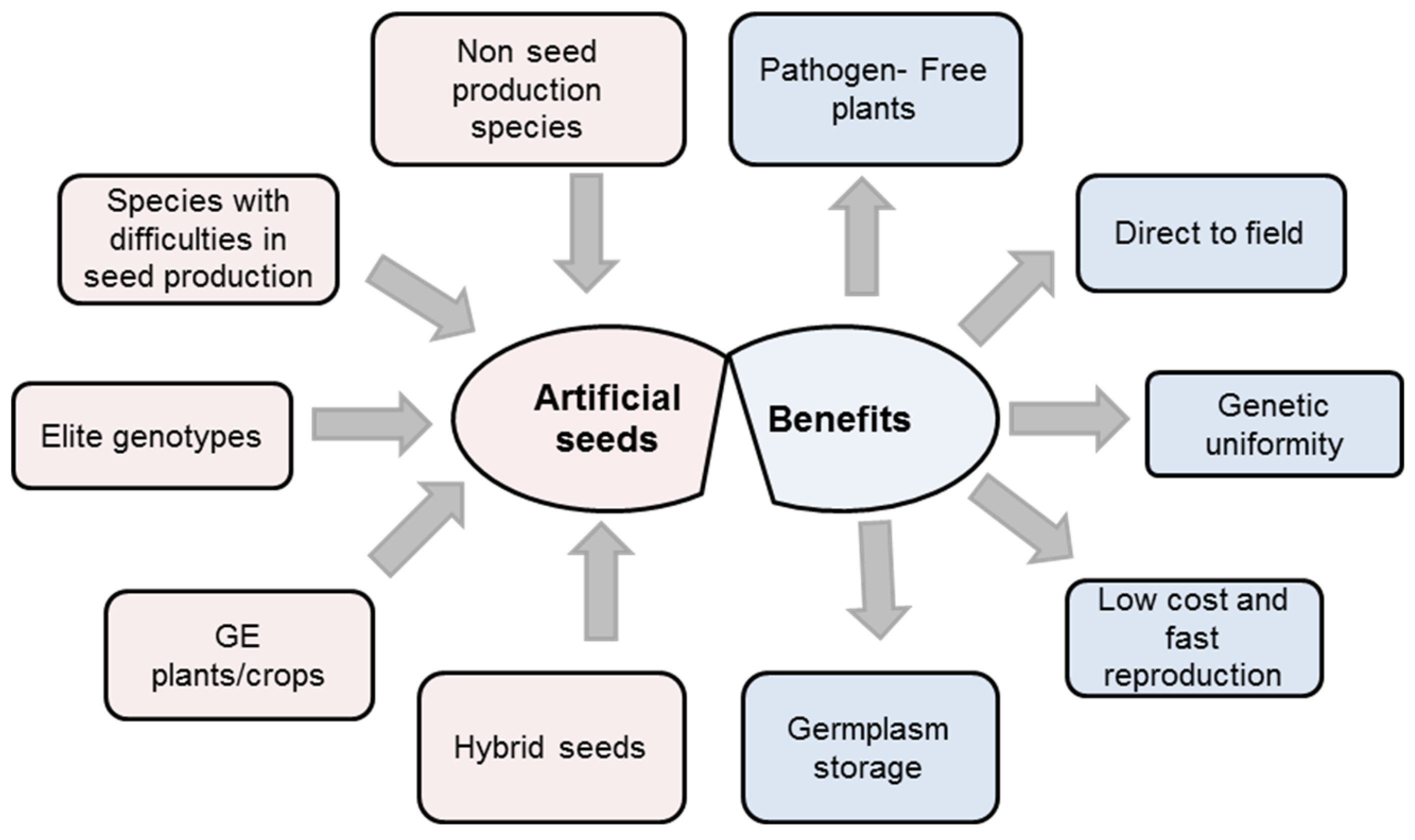
2. The Importance, Uses and Advantages of Artificial Seeds
- Those that have a high quality of somatic embryos, and
- Those with a strong commercial basis [34].
3. Artificial Seed Concept
4. Essential Requirements for the Production of Artificial Seeds
4.1. Explant Material
4.2. Somatic Embryos
5. Artificial Seed Gelling Agents and Adjuvant Materials
6. Artificial Endosperm Structure and Their Effects in the Artificial Seed Characters
7. Artificial Seeds Storage Ability
8. Limitation of Artificial Seeds
9. Conclusions
Author Contributions
Conflicts of Interest
Abbreviations
| SA | sodium alginate |
| WPM | woody plant medium |
| BAP | N6–benzylaminopurine |
| BA | 6-Benzylaminopurine |
| MS | Murashige and Skoog medium |
| NAA | naphathaleneacetic acid |
| GD | Gresshoff and Doy medium |
| IAA | indolacetic acid |
| LN | liquid nitrogen |
References
- Murashige, T. Plant cell and organ cultures as horticultural practices. In Proceedings of the Symposium on Tissue Culture for Horticultural Purposes, Ghent, Belgium, 6–9 September 1977. [Google Scholar]
- Gray, D.J.; Purohit, A.; Triglano, R.N. Somatic embryogenesis and development of synthetic seed technology. Crit. Rev. Plant Sci. 1991, 10, 33–61. [Google Scholar] [CrossRef]
- Redenbaugh, K.; Paasch, B.D.; Nichol, J.W.; Kossler, M.E.; Viss, P.R.; Walker, K.A. Somatic seeds-encapsulation of asexual plant embryos. Nat. Biotechnol. 1986, 4, 797–801. [Google Scholar] [CrossRef]
- Redenbaugh, K.; Fujii, J.A.; Slade, D. Encapsulated plant embryos. In Advances in Biotechnological Processes; Mizrahi, A., Ed.; Alan R. Liss Inc.: New York, NY, USA, 1988; pp. 225–248. [Google Scholar]
- Bapat, V.A.; Mhatre, M.; Rao, P.S. Propagation of Morus indica L. (Mulberry) by encapsulated shoot buds. Plant Cell Rep. 1987, 6, 393–395. [Google Scholar] [CrossRef] [PubMed]
- Ara, H.; Jaiswal, U.; Jaiswal, V. Synthetic seed: Prospects and limitation. Curr. Sci. 2000, 78, 1438–1444. [Google Scholar]
- Daud, M.; Taha, M.Z.; Hasbullah, A.Z. Artificial seed production from encapsulated micro shoots of Sainpaulia ionantha Wendl. (African Violet). J. Appl. Sci. 2008, 8, 4662–4667. [Google Scholar]
- Saiprasad, G. Artificial seeds and their applications. Resonance 2001, 6, 39–47. [Google Scholar] [CrossRef]
- Onishi, N.; Sakamoto, Y.; Hirosawa, T. Synthetic seeds as an applicationof mass-production of somatic embryos. Plant Cell Tissue Organ Cult. 1994, 39, 137–145. [Google Scholar] [CrossRef]
- Sharma, S.; Shahzad, A. Encapsulation technology for short-term storage and conservation of a woody climber, Decalepis hamiltonii Wight and Arn. Plant Cell Tissue Organ Cult. 2012, 111, 191–198. [Google Scholar] [CrossRef]
- Rihan, H.Z.; Al-Issawi, M.; Burchett, S.; Fuller, M.P. Encapsulation of cauliflower (Brassica oleracea var botrytis) microshoots as artificial seeds and their conversion and growth in commercial substrates. Plant Cell Tissue Organ Cult. 2011, 107, 243–250. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, N.; Anis, M. Direct plant regeneration from encapsulated nodal segments of Vitex negundo. Biol. Plant. 2010, 54, 748–752. [Google Scholar] [CrossRef]
- Rai, M.K.; Jaiswal, V.S.; Jaiswal, U. Encapsulation of shoot tips of guava (Psidium guajava L.) for short-term storage and germplasm exchange. Sci. Hortic. 2008, 118, 33–38. [Google Scholar] [CrossRef]
- Danso, K.E.; Ford-Lloyd, B.V. Encapsulation of nodal cuttings and shoot tips for storage and exchange of cassava germplasm. Plant Cell Rep. 2003, 21, 718–725. [Google Scholar] [PubMed]
- Bapat, V.A.; Rao, P.S. Sandalwood plantlets from synthetic seeds. Plant Cell Rep. 1988, 7, 434–436. [Google Scholar] [PubMed]
- Mathur, J.; Ahuja, P.S.; Lal, N.; Mathur, A.K. Propagation of Valeriana wallichii DC. using encapsulated apical and axial shoot buds. Plant Sci. 1989, 60, 111–116. [Google Scholar] [CrossRef]
- Ganapathi, T.R.; Suprasanna, P.; Bapat, V.A.; Rao, P.S. Propagation of banana through encapsulated shoot tips. Plant Cell Rep. 1992, 11, 571–575. [Google Scholar] [CrossRef] [PubMed]
- Corrie, S.; Tandon, P. Propagation of Cymbidium giganteum wall through high frequency conversion of encapsulated protocorms under in vivo and in vitro conditions. Indian J. Exp. Biol. 1993, 31, 61–64. [Google Scholar]
- Sharma, T.R.; Singh, B.M.; Chauhan, R.S. Production of disease free encapsulated buds of Zingiber officinale Rosc. Plant Cell Rep. 1994, 13, 300–302. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, D.; Naik, P.S. Synseeds in potato: An investigation using nutrient-encapsulated in vitro nodal segments. Sci. Hortic. 1998, 7, 179–184. [Google Scholar] [CrossRef]
- Mandal, J.; Pattnaik, S.; Chand, P. Alginate encapsulation of axillary buds of Ocimum americanum L. (hoary basil), Basilicum L. (sweet basil), Gratissimum L. (shrubby basil), and Sanctum L. In Vitro Cell. Dev. Biol. Plant 2000, 36, 287–292. [Google Scholar] [CrossRef]
- Nyende, A.B.; Schittenhelm, S.; Mix-Wagner, G.; Greef, J.M. Production, storability, and regeneration of shoot tips of potato (Solanum tuberosum L.) encapsulated in calcuim alginate hollow beads. In Vitro Cell. Dev. Biol. Plant 2003, 39, 540–544. [Google Scholar] [CrossRef]
- Chand, S.; Singh, A.K. Plant regeneration from encapsulated nodal segments of Dalbergia sissoo Roxb, a timber-yielding leguminous tree species. J. Plant Physiol. 2004, 161, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Naik, S.K.; Chand, P.K. Nutrient-alginate encapsulation of in vitro nodal segments of pomegranate (Punica granatum L.) for germplasm distribution and exchange. Sci. Hortic. 2006, 108, 247–252. [Google Scholar] [CrossRef]
- Singh, A.K.; Varshney, R.; Sharma, M.; Agarwal, S.S.; Bansal, K.C. Regeneration of plants from alginate-encapsulated shoot tips of Withania somnifera (L.) Dunal, a medicinally important plant species. J. Plant Physiol. 2006, 163, 220–223. [Google Scholar] [CrossRef] [PubMed]
- Faisal, M.; Anis, M. Regeneration of plants from alginate-encapsulated shoots of Tylophora indica (Burm. f.) Merrill, an endangered medicinal. J. Hortic. Sci. Biotechnol. 2007, 82, 351–354. [Google Scholar] [CrossRef]
- Micheli, M.; Hafiz, I.A.; Standardi, A. Encapsulation of in vitro-derived explants of olive (Olea europaea L. cv. Moraiolo): II. Effects of storage on capsule and derived shoots performance. Sci. Hortic. 2007, 113, 286–292. [Google Scholar] [CrossRef]
- Rai, M.K.; Jaiswa, V.S. Alginate-encapsulation of nodal segments of guava (Psidium guajava L.) for germplasm exchange and distribution. J. Hortic. Sci. Biotechnol. 2008, 83, 569–573. [Google Scholar] [CrossRef]
- Rai, M.K.; Asthana, P.; Singh, S.K.; Jaiswal, V.S.; Jaiswal, U. The encapsulation technology in fruit plants—A review. Biotechnol. Adv. 2009, 27, 671–679. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.K.; Rai, M.K.; Asthana, P.; Pandey, S.; Jaiswal, V.S.; Jaiswal, U. Plant regeneration from alginate-encapsulated shoot tips of Spilanthes acmella L. Murr., a medicinally important and herbal pesticidal plant species. Acta Physiol. Plant. 2009, 31, 649–653. [Google Scholar] [CrossRef]
- Sharma, S.; Shahzad, A.; da Silva, J.A.T. Synseed technology—A complete synthesis. Biotechnol. Adv. 2013, 31, 186–207. [Google Scholar] [CrossRef] [PubMed]
- Pattnaik, S.K.; Sahoo, Y.; Chand, P.K. Efficient plant retrieval from alginate-encapsulated vegetative buds of mature mulberry trees. Sci. Hortic. 1995, 61, 227–239. [Google Scholar] [CrossRef]
- Banerjee, S.; Singh, S.; Pandey, H.; Pandey, P.; ur Rahman, L. Conservation and storage of Curcuma amada Roxb. synseeds on Luffa sponge matrix and RAPD analysis of the converted plantlets. Ind. Crop. Prod. 2012, 36, 383–388. [Google Scholar] [CrossRef]
- Redenbaugh, K.; Slade, D.; Viss, P.; Fujii, J.A. Encapsulation of somatic embryos in synthetic seed coats. HortScience 1987, 22, 803–809. [Google Scholar]
- Cartes, P.; Castellanos, H.; Ríos, D.; Sáez, K.; Spierccolli, S.; Sánchez, M. Encapsulated somatic embryos and zygotic embryos for obtaining artificial seeds of rauli-beech (Nothofagus alpina (Poepp. & Endl.) oerst.). Chil. J. Agric. Res. 2009, 69, 112–118. [Google Scholar]
- Standardi, A.; Micheli, M. Encapsulation of in vitro-derived explants: An innovative tool for nurseries. Methods Mol. Biol. 2013, 11013, 397–418. [Google Scholar] [PubMed]
- Nyende, A.B.; Schittenhelm, S.; Mix-Wagner, G.; Greef, J.M. Yield and canopy development of field grown potato plants derived from synthetic seeds. Eur. J. Agron. 2005, 22, 175–184. [Google Scholar] [CrossRef]
- Lata, H.; Chandra, S.; Khan, I.A.; ElSohly, M.A. Propagation through alginate encapsulation of axillary buds of Cannabis sativa L.—An important medicinal plant. Physiol. Mol. Biol. Plants 2009, 15, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Pennycooke, J.C.; Towill, L.E. Medium alterations improve regrowth of sweet potato (Ipomoea batatas (L.) Lam.) shoot tips cryopreserved by vitrification and encapsulation-dehydration. Cryoletters 2001, 22, 381–389. [Google Scholar] [PubMed]
- Wang, Q.; Batuman, Ö.; Li, P.; Bar-Joseph, M.; Gafny, R. A simple and efficient cryopreservation of in vitro-grown shoot tips of ‘Troyer’ citrange [Poncirus trifoliata (L.) Raf. × Citrus sinensis (L.) Osbeck.] by encapsulation-vitrification. Euphytica 2002, 128, 135–142. [Google Scholar] [CrossRef]
- Mohamed, M. Application of Synthetic Seeds—Present and the Future. Available online: http://www.scribd.com/doc/22585470/Application-of-Synthetic-Seeds-Present-and-the-Future (accessed on 28 December 2009).
- Leroy, X.J.; Leon, K.; Charles, G.; Branchard, M. Cauliflower somatic embryogenesis and analysis of regenerant stability by ISSRs. Plant Cell Rep. 2000, 19, 1102–1107. [Google Scholar] [CrossRef]
- Deng, W. A novel method for induction of plant regeneration via somatic embryogenesis. Plant Sci. 2009, 177, 43–48. [Google Scholar] [CrossRef]
- Kitto, S.L.; Janick, J. Polyox as an artificial seed coat for a sexual embryos. HortScience 1982, 17, 488. [Google Scholar]
- Gupta, P.K.; Durzan, D.J. Somatic embryos from protoplasts of Loblolly-pine proembryonal cells. Nat. Biotechnol. 1987, 5, 710–712. [Google Scholar] [CrossRef]
- Onay, A.; Jeffree, C.E.; Yeoman, M.M. Plant regeneration from encapsulated embryoids and an embryogenic mass of pistachio, Pistacia vera L. Plant Cell Rep. 1996, 15, 723–726. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, B.; Sen, S. Plant-regeneration from alginate encapsulated somatic embryos of Asparagus-cooperi-baker. Plant Cell Rep. 1994, 13, 381–385. [Google Scholar] [CrossRef] [PubMed]
- Ara, H.; Jaiswal, U.; Jaiswal, V.S. Germination and plantlet regeneration from encapsulated somatic embryos of mango (Mangifera indica L.). Plant Cell Rep. 1999, 19, 166–170. [Google Scholar] [CrossRef]
- Antonietta, G.M.; Emanuele, P.; Alvaro, S. Effects ofencapsulation on Citrus reticulata Blanco somatic embryo conversion. Plant Cell Tissue Organ Cult. 1999, 55, 235–237. [Google Scholar] [CrossRef]
- Sunilkumar, K.K.; Sudhakara, K.; Vijaykumar, N.K. An attempt to improve storage life of Hopea parviflora seeds through synthetic seed production. Seed Res. 2000, 28, 126–130. [Google Scholar]
- Ipekei, Z.; Gozukirmizi, N. Direct somatic embryogenesis and synthetic seed production from Paulownia elongata. Plant Cell Rep. 2003, 22, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Nieves, N.; Zambrano, Y.; Tapia, R.; Cid, M.; Pina, D.; Castillo, R. Field performance of artificial seed derived sugarcane plants. Plant Cell Tissue Organ Cult. 2003, 75, 279–282. [Google Scholar] [CrossRef]
- Kumar, M.B.A.; Vakeswaran, V.; Krishnasamy, V. Enhancement of synthetic seed conversion to seedlings in hybrid rice. Plant Cell Tissue Organ Cult. 2005, 81, 97–100. [Google Scholar] [CrossRef]
- Chithra, M.; Martin, K.P.; Sunandakumari, C.; Madhusoodanan, P.V. Somatic embryogenesis, encapsulation, and plant regeneration of Rotula aquatica lour, a rare rhoeophytic woody medicinal plant. In Vitro Cell. Dev. Biol. Plant 2005, 41, 28–31. [Google Scholar] [CrossRef]
- Aquea, F.; Poupin, M.J.; Matus, J.T.; Gebauer, M.; Medina, C.; Arce-Johnson, P. Synthetic seed production from somatic embryos of Pinus radiata. Biotechnol. Lett. 2008, 30, 1847–1852. [Google Scholar] [CrossRef] [PubMed]
- Maqsood, M.; Mujib, A.; Siddiqui, Z.H. Synthetic seed development and conversion to plantlet in Catharanthus roseus (L.) G. Don. Biotechnology 2012, 11, 37–43. [Google Scholar]
- Attree, S.M.; Pomeroy, M.K.; Fowke, L.C. Production of vigoous, dessication tolerant white spruce (Picea-glauca (moench) voss) synthetic seeds in a bioreactor. Plant Cell Rep. 1994, 13, 601–606. [Google Scholar] [CrossRef] [PubMed]
- Lulsdorf, M.M.; Tautorus, T.E.; Kikcio, S.I.; Bethune, T.D.; Dunstan, D.I. Germenation of encapsulated embryos of interior spruce (Picea-glauca-engelmanni complex) and black spruce (Picea-mariqana Mill). Plant Cell Rep. 1993, 12, 385–389. [Google Scholar] [PubMed]
- Ganapathi, T.R.; Srinivas, L.; Suprasanna, P.; Bapat, V.A. Regeneration of plants from alginate-encapsulated somatic embryos of banana cv. Rasthali (Musa SPP. AAB Group). In Vitro Cell. Dev. Biol. Plant 2001, 37, 178–181. [Google Scholar] [CrossRef]
- Malabadi, R.B.; van Staden, J. Storability and germination of sodium alginate encapsulated somatic embryos derived from the vegetative shoot apices of mature Pinus patula trees. Plant Cell Tissue Organ Cult. 2005, 82, 259–265. [Google Scholar] [CrossRef]
- Manjkhola, S.; Dhar, U.; Joshi, M. Organogenesis, embryogenesis, and synthetic seed production in Arnebia euchroma—A critically endangered medicinal plant of the Himalaya. In Vitro Cell. Dev. Biol. Plant 2005, 41, 244–248. [Google Scholar] [CrossRef]
- West, T.P.; Ravindra, M.B.; Preece, J.E. Encapsulation, cold storage, and growth of Hibiscus moscheutos nodal segments. Plant Cell Tissue Organ Cult. 2006, 87, 223–231. [Google Scholar] [CrossRef]
- Gomez, R. Embriogénesis somática. In Propagación y Mejora Genética de Plantas por Biotecnología; Pérez, J.N., Ed.; Instituto de Biotecnología de las Plantas: Santa Clara, Cuba, 1998; pp. 57–79. [Google Scholar]
- Rodríguez, N.N.; Lorenzo, F.; Blanco, J.M.A.; González, O.; Tapia, T. Composicion de un endospermo artificial para embriones de mandarina ‘Cleopatra’ (Citrus reshni Hort es Tan). Rev. Fac. Agron. 2001, 27, 11–15. [Google Scholar]
- Nair, R.R.; Gupta, S.D. In vitro plant regeneration from encapsulated somatic embryos of black pepper (Piper nigrum L.). J. Plant Sci. 2007, 2, 283–292. [Google Scholar]
- Tapia, R.; Castillo, R.; Nieves, N.; Blanco, M.A.; González, J.; Sánchez, M.; Rodríguez, Y. Inducción, Maduración Y Encapsulación De Embriones Somáticos De Caña De Azucar (Saccharum sp.) var CP 5243. Biotecnol. Apl. 1999, 16, 20–23. [Google Scholar]
- Castellanos, H.; Sánchez-Olate, M.; Ríos, Y.D. Embriogénesis somática recurrente en raulí (Nothofagus alpina (Poepp. et Endl.) Oerst.). In Segundo Congreso Chileno de Ciencias Forestales, Valdivia, Chile. 10–12 de Noviembre; Universidad Austral de Chile: Valdivia, Chile, 2004; p. 36. [Google Scholar]
- Bapat, V.A.; Mhatre, M. Bioencapsulation of Somatic Embryos in Woody Plants. In Protocol for Somatic Embryogenesis in Woody Plants; Springer: Dordrecht, The Netherlands, 2005; pp. 539–552. [Google Scholar]
- Sundararaj, S.G.; Agrawal, A.; Tyagi, R.K. Encapsulation for in vitro short-term storage and exchange of ginger (Zingiber officinale Rosc.) germplasm. Sci. Hortic. 2010, 125, 761–766. [Google Scholar] [CrossRef]
- Pond, S.; Cameron, S. Tissue Culture\Artificial Seed. In Encyclopedia of Applied Plant Sciences; Thomas, B., Ed.; Elsevier: Oxford, UK, 2003; pp. 1379–1388. [Google Scholar]
- Redenbaugh, K. Synseeds: Applications of Synthetic Seeds to Crop Improvement; CRC Press: Boca Raton, FL, USA, 1993; pp. 38–46. [Google Scholar]
- Latif, Z.; Nasir, I.A.; Riazuddin, S. Indigenous production of synthetic seeds in Daucus carota. Pak. J. Bot. 2007, 39, 849–855. [Google Scholar]
- Bapat, V.A.; Rao, P.S. In vivo growth of encapsulated axillary buds of mulberry (Morus-indical L.). Plant Cell Tissue Organ Cult. 1990, 20, 69–70. [Google Scholar] [CrossRef]
- Piccioni, E.; Standardi, A. Encapsulation of micropropagated buds of 6 woody species. Plant Cell Tissue Organ Cult. 1995, 42, 221–226. [Google Scholar] [CrossRef]
- Capuano, G.; Piccioni, E.; Standardi, A. Effect of different treatments on the conversion of M.26 apple rootstock synthetic seeds obtained from encapsulated apical and axillary micropropagated buds. J. Hortic. Sci. Biotechnol. 1998, 73, 299–305. [Google Scholar] [CrossRef]
- Rihan, H.Z.; Al-Issawi, M.; Al-Swedi, F.; Fuller, M.P. The effect of using PPM (plant preservative mixture) on the development of cauliflower microshoots and the quality of artificial seed produced. Sci. Hortic. 2012, 141, 47–52. [Google Scholar] [CrossRef]
- Datta, K.B.; Kanjilal, B.; de Sarker, D. Artificial seed technology: Development of a protocol in Geodorum densiflorum (Lam) Schltr.—An endangered orchid. Curr. Sci. 1999, 76, 1142–1145. [Google Scholar]
- Redenbaugh, K.; Nichol, J.; Kossler, M.E.; Paasch, B. Encapsulation of Somatic Embryos for Artificial Seed Production. In Vitro J. Tissue Cult. Assoc. 1984, 20, 256–257. [Google Scholar]
- Redenbaugh, K.; Fujii, J.A.; Slade, D. Hydrated coating for synthetic seeds. In Synseeds: Application of the Synthetic Seeds to Crop Improvement; CRC Press: Boca Raton, FL, USA, 1993; pp. 305–327. [Google Scholar]
- Ahuja, M.R. Micropropagation of woody plants. In Synthetic Seeds in Forest Trees; Gupta, P., Kreitinger, M., Eds.; Kluwer Academic Publisher: Dordrecht, The Netherlands, 1993; p. 107. [Google Scholar]
- Castillo, B.; Smith, M.A.L.; Yadava, U.L. Plant regeneration from encapsulated somatic embryos of Carica papaya L. Plant Cell Rep. 1998, 17, 172–176. [Google Scholar] [CrossRef]
- Senaratna, T. Artificial seeds. Biotechnol. Adv. 1992, 10, 379–392. [Google Scholar] [CrossRef]
- Khor, E.; Loh, C. Artificial seeds. In Applications of Cell Immobilization Biotechnology; Nedovic, V., Willaert, R., Eds.; Springer: Berlin/Heidelberg, Germany, 2005; pp. 527–537. [Google Scholar]
- Gray, D.J. Synthetic seed for clonal production of crop plants. In Recent Advances in the Development and Germination of Seeds; Springer: Boston, MA, USA, 1989. [Google Scholar]
- Nieves, N.; Lorenzo, J.C.; Blanco, M.A. Artificial endosperm of Cleopatra tangerine zygotic embryos: A model for somatic embryo encapsulation. Plant Cell Tissue Organ Cult. 1998, 54, 77–83. [Google Scholar] [CrossRef]
- Lakshmana Rao, P.V.; Singh, B. Plantlet regeneration from encapsulated somatic embryos of hybrid Solanum melongena L. Plant Cell Rep. 1991, 10, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Rai, M.K.; Jaiswal, V.S.; Jaiswal, U. Effect of ABA and sucrose on germination of encapsulated somatic embryos of guava (Psidium guajava L.). Sci. Hortic. 2008, 117, 302–305. [Google Scholar] [CrossRef]
- Ray, A.; Bhattacharya, S. Storage and plant regeneration from encapsulated shoot tips of Rauvolfia serpentine—An effective way of conservation and mass propagation. S. Afr. J. Bot. 2008, 74, 776–779. [Google Scholar] [CrossRef]
- Fabre, J.; Dereuddre, J. Encapsulation dehydration-a new approach to cryopreservation of Solanium shoot-tips. Cryo-Letters 1990, 11, 413–426. [Google Scholar]
- Tsvetkov, I.; Hausman, J.-F. In vitro regeneration from alginate-encapsulated microcuttings of Quercus sp. Sci. Hortic. 2005, 103, 503–507. [Google Scholar] [CrossRef]
- Gresshof, P.M.; Doy, C.H. Development and differentiation of haploid lycopersicon-esculentum (tomato). Planta 1972, 107, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Verleysen, H.; van Bockstaele, E.; Debergh, P. An encapsulation-dehydration protocol for cryopreservation of the azalea cultivar ‘Nordlicht’ (Rhododendron simsii Planch.). Sci. Hortic. 2005, 106, 402–414. [Google Scholar] [CrossRef]
- Faisal, M.; Ahmad, N.; Anis, M. In vitro plant regeneration from alginate-encapsulated microcuttings of Rauvolfia tetraphylla L. Am. Eurasian J. 2006, 1, 1–6. [Google Scholar]
- Wang, W.G.; Wang, S.H.; Wu, X.A.; Jin, X.Y.; Chen, F. High frequency plantlet regeneration from callus and artificial seed production of rock plant Pogonatherum paniceum (Lam.) Hack. (Poaceae). Sci. Hortic. 2007, 113, 196–201. [Google Scholar] [CrossRef]
- Sujatha, G.; Kumari, B.D.R. Micropropagation, encapsulation and growth of Artemisia vulgaris node explants for germplasm preservation. S. Afr. J. Bot. 2008, 74, 93–100. [Google Scholar] [CrossRef]
- Rihan, H.Z.; Al-Shamari, M.; Al-Swedi, F.; Burchett, S.; Fuller, M.P. The effect of sugar type, source and concentration on Brassica oleraceae var botrytis microproshoot production. Acta Hortic. 2013. under review. [Google Scholar]
- Rihan, H.Z.; Al-Issawi, M.; Burchett, S.; Fuller, M.P. Artificial seed production from encapsulated microshoots of cauliflower (Brassica oleraceae var botrytis). Acta Hortic. 2012, 961, 419–425. [Google Scholar] [CrossRef]
- Rihan, H.Z.; Shamari, M.; Fuller, M.P. The production of cauliflower microshoots using curd meristematic tissues and hypocotyl-derived callus. Acta Hortic. 2012, 961, 427–434. [Google Scholar] [CrossRef]
- Reddy, M.C.; Murthy, K.S.R.; Pullaiah, T. Synthetic seeds: A review in agriculture and forestry. Afr. J. Biotechnol. 2012, 11, 14254–14275. [Google Scholar]
- Makowczyńska, J.; Andrzejewska-Golec, E. Somatic seeds of Plantago asiatica L. Acta Soc. Bot. Pol. 2006, 75, 17–21. [Google Scholar]
- Hung, C.; Trueman, S. Alginate encapsulation of shoot tips and nodal segments for short-term storage and distribution of the eucalypt Corymbia torelliana × C. citriodora. Acta Physiol. Plant 2012, 34, 117–128. [Google Scholar] [CrossRef]
- Jung, S.J.; Yoon, E.S.; Jeong, J.H.; Choi, Y.E. Enhanced post-germinative growth of encapsulated somatic embryos of Siberian ginseng by carbohydrate addition to the encapsulation matrix. Plant Cell Rep. 2004, 23, 365–370. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Reference | Species and Explant Used | Pre-Encapsulation Culture | Capsule Structure | Additional Materials | Storage Conditions | Post Storage Treatment or Culture | Storage Duration | Experiment Assessment |
|---|---|---|---|---|---|---|---|---|
| [90] | Apical segments of Turkey oak (Quercus cerris L.) | Modified GD medium [91] 0.88 BAP + 7.5 g·L−1 agar | 4% (w/v) SA immersed in 1.4% (w/v) CaCl2 for 10 min | GD modified medium + 0.88mM BAP | 4 °C | - | 6 weeks | 85% regeneration and 95% viability |
| [92] | Shoot tips of azalea (Rhododendorn simsii Planch.) | Modified (WPM) medium + (0.3, 0.45, 0.6 M) sucrose (24) h | drops SA immersed in CaCl2 for 15 min | 1.62 M glycerol + 0.32 M sucrose (cryoprotectants) | Dehydration to WC (38.6)% + transfer to LN | Detoxification in (1) M sucrose for (2) h | 24 h | 40% survived after thawing |
| [24] | Nodal segments of promegranate (Punica granatum L.) | - | 3% Sa + 100 mM CaCl2 for 30 min | MS medium + 4.44 mM (BA) + 0.54 mM (NAA) | 4 °C | (MS) medium + 4.44 mM (BA) + 0.54 mM (NAA) | 30 days | The nodals were capable of sprouting. |
| [93] | Nodal segments of Rauvolfia tetraphylla L. | MS medium + 10 mM BA + 0.5 mM NAA + 3% (w/v) sucrose + 0.8% (w/v) agar | 3% SA + 100 mM for 30 min | MS medium | 4 °C | MS medium + 10 mM BA + 0.5 NAA + 3% sucrose + 0.8% agar | 4 weeks | Shoot proliferation 86.7% |
| [94] | Shoot buds were derived from lam (Progonatherum paniceum) calli | MS medium supplemented with 2.0 mg·L−1 BAP and 0.2 mg·L−1 NAA. | 3.0% SA + 2% CaCl2 for 20 min | 1% activate carbon (w/v) + 0.3% bavistin (w/v) + MS medium containing 3% sucrose | - | substrata containing 8 g·L−1 agar with half-strength MS medium and 2% sucrose | - | 61.58% conversion rate |
| [95] | Node explants of mugwort (Artemisia vulgaris) | Nodal segments were excised from poliferating microshoots | 2% SA + 50 mM CaCl2 for 15–25 min | - | 5 °C | MS + 3% (w/v) sucrose | 60 weeks | 85% of encapsulated nodal segments survived on poliferation mendium |
| [88] | Shoot tips of Rauvolfia serpentina | MS medium + 3% sucrose+ 0.8% agar | 3% SA + 100mM CaCl2 for 45 min | Encapsulated shoot tips kept in vessels containing MS + 3% sucrose during storage period | 4 °C | MS + 3% scrose | 14 weeks | (68.5–100%) regrowth rate during storage period (several tests in the normal culture conditions) |
| [87] | Somatic embryos of guava (Psidium guajava L.) | - | 2% SA + 100 mM CaCl2 for 20–30 min | Encapsulated somatic embryos + MS + 9% sucrose or + MS+ 1 mg·L−1 ABA | 25 ± 2° | MS + 3% sucrose | 60 days | 28.8% germination level and 37.5% germination level For (sucrose and ABA respectively) |
| [35] | Somatic embryos from mature seeds of rauli-beech (Nothofagus alpine) | - | 3% SA + 5.5 g·L−1 CaCl2 for 30 min | MS + 0.5 mg·L−1 IAA + 0.5 mg·L−1 NAA + 2 mg·L−1 BAP + 30 sucrose | - | culture MS + 30 g·L−1 sucrose + 7 g·L−1 agar | - | The germination rate was 45% after 4 weeks |
| [38] | Axillary buds of Cannabis sativa L. | MS medium + 3% (w/v) sucrose + 0.8% agar + 0.5 mM (TDZ) | 5% SA + CaCl2 for 30 min | MS + 0.5 mM (TDZ) + 2.5 mM (IBA). A + fungicide + 5% (PPM) | - | MS medium + 0.5 TDZ + 0.075% PPM | - | 100% artificial seed conversion rate |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rihan, H.Z.; Kareem, F.; El-Mahrouk, M.E.; Fuller, M.P. Artificial Seeds (Principle, Aspects and Applications). Agronomy 2017, 7, 71. https://doi.org/10.3390/agronomy7040071
Rihan HZ, Kareem F, El-Mahrouk ME, Fuller MP. Artificial Seeds (Principle, Aspects and Applications). Agronomy. 2017; 7(4):71. https://doi.org/10.3390/agronomy7040071
Chicago/Turabian StyleRihan, Hail Z., Fakhriya Kareem, Mohammed E. El-Mahrouk, and Michael P. Fuller. 2017. "Artificial Seeds (Principle, Aspects and Applications)" Agronomy 7, no. 4: 71. https://doi.org/10.3390/agronomy7040071