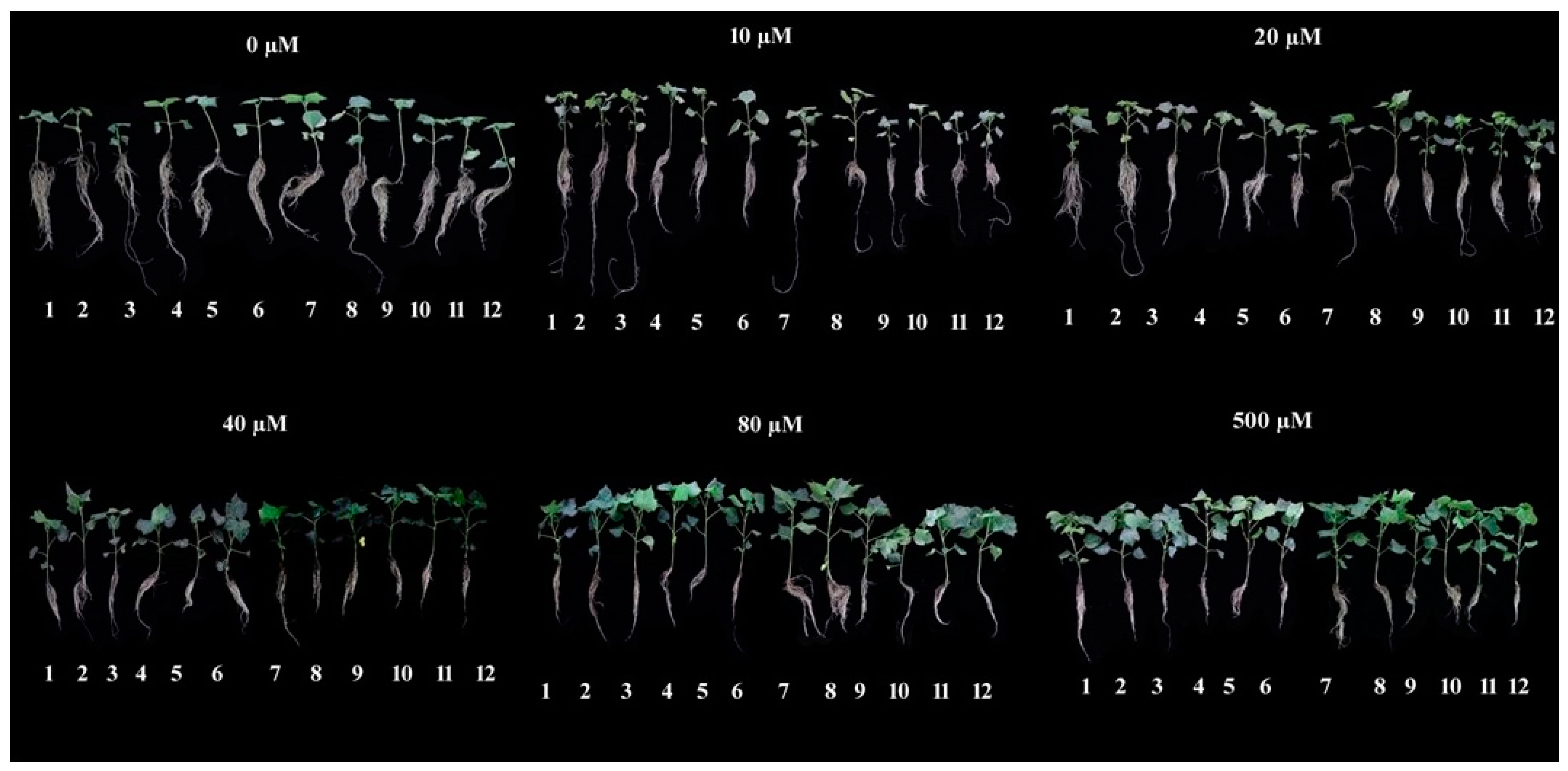
Figure 1.
Representative examples of genotype-specific plant phenotypes of 12 cotton genotypes grown under various P concentrations (0, 10, 20, 40, 80, and 500 µM). Each number from 1 to 12 represents genotypes TM-1, Yumian-21, CCRI-64, CCRI-27, Xinluzhong-17, Xinluzao-35, Xinluzao-48, Xinluzao-49, Xinluzhong-12, Xinluzhong-15, Xinluzao-20, and CCRI-12, respectively.
Figure 1.
Representative examples of genotype-specific plant phenotypes of 12 cotton genotypes grown under various P concentrations (0, 10, 20, 40, 80, and 500 µM). Each number from 1 to 12 represents genotypes TM-1, Yumian-21, CCRI-64, CCRI-27, Xinluzhong-17, Xinluzao-35, Xinluzao-48, Xinluzao-49, Xinluzhong-12, Xinluzhong-15, Xinluzao-20, and CCRI-12, respectively.
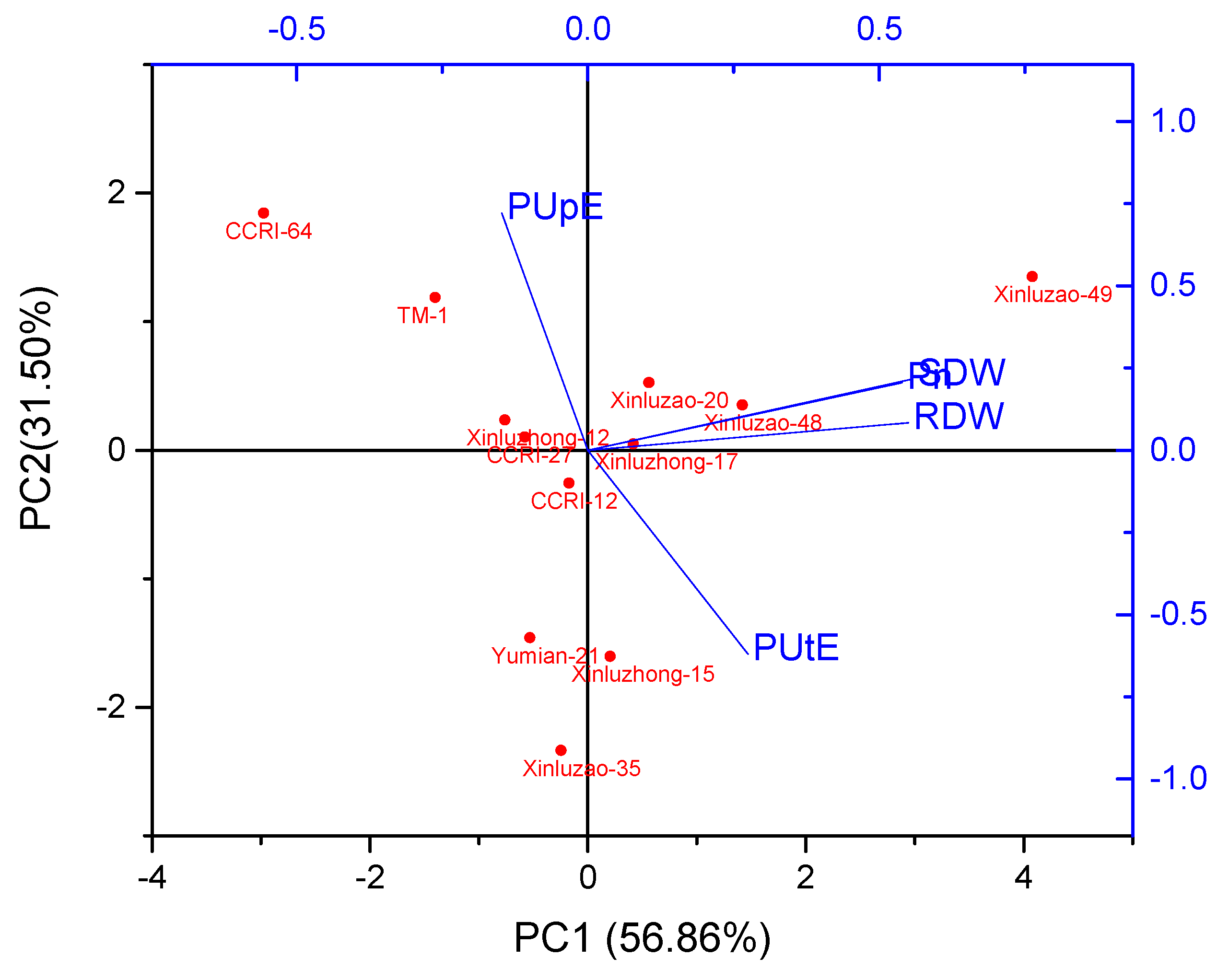
Figure 2.
Principal component analysis (PCA) biplot of morphophysiological traits of 12 cotton genotypes grown under various P concentrations. SDW; shoot dry weight, RDW; root dry weight, Pn: net photosynthesis, PUpE; P uptake efficiency and PUtE; P utilization efficiency.
Figure 2.
Principal component analysis (PCA) biplot of morphophysiological traits of 12 cotton genotypes grown under various P concentrations. SDW; shoot dry weight, RDW; root dry weight, Pn: net photosynthesis, PUpE; P uptake efficiency and PUtE; P utilization efficiency.
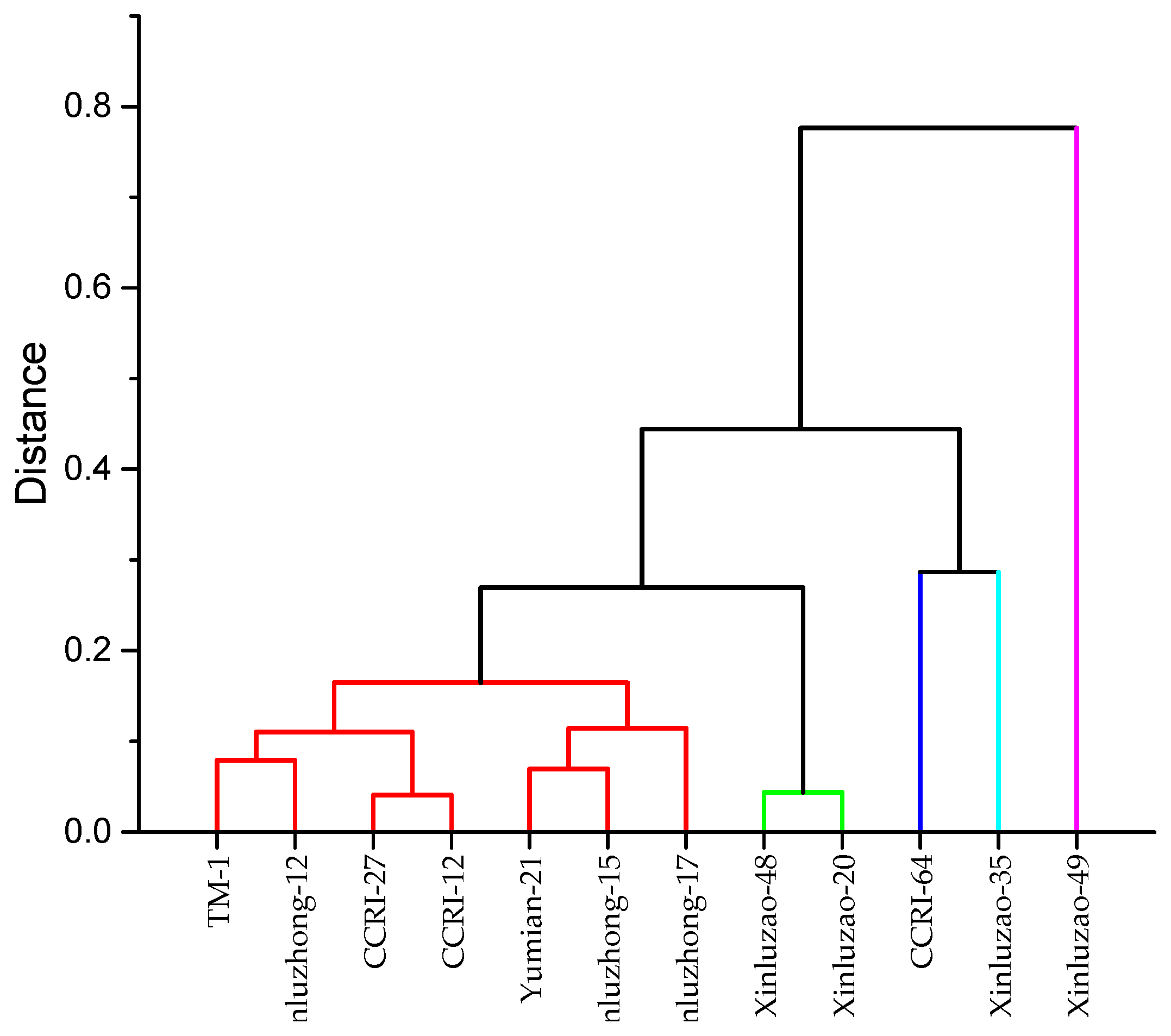
Figure 3.
Dendrogram hierarchical cluster analysis of cotton genotypes based on morphophysiological traits grown under various P concentrations (0, 10, 20, 40, 80, and 500 µM).
Figure 3.
Dendrogram hierarchical cluster analysis of cotton genotypes based on morphophysiological traits grown under various P concentrations (0, 10, 20, 40, 80, and 500 µM).
Table 1.
Shoot length (SL, cm), root dry weight (RDW, g), shoot dry weight (SDW, g), total plant dry weight (TDW, g), number of leaves (NOL), and leaf area (LA, cm2) of 12 cotton genotypes grown at different P concentrations in hydroponic culture.
Table 1.
Shoot length (SL, cm), root dry weight (RDW, g), shoot dry weight (SDW, g), total plant dry weight (TDW, g), number of leaves (NOL), and leaf area (LA, cm2) of 12 cotton genotypes grown at different P concentrations in hydroponic culture.
| Genotypes | SL | RDW | SDW | TDW | NOL | LA |
|---|
| TM-1 | 11.6 cd | 0.28 d | 0.64 b | 0.92 cd | 5.8 abc | 51.94 b |
| Yumian-21 | 12.3 bcd | 0.27 de | 0.64 b | 0.91 cd | 5.8 abc | 50.83 b |
| CCRI-64 | 9.10 d | 0.26 e | 0.63 b | 0.89 d | 5.2 cd | 44.09 b |
| CCRI-27 | 14.2 bc | 0.30 c | 0.65 b | 0.95 bc | 5.7 bcd | 50.09 b |
| Xinluzhong-17 | 16.6 b | 0.31 bc | 0.66 b | 0.97 bc | 5.3 cd | 55.05 ab |
| Xinluzao-35 | 14.0 bc | 0.30 c | 0.65 b | 0.95 bc | 5.0 d | 41.63 b |
| Xinluzao-48 | 17.0 b | 0.34 b | 0.67 b | 1.01 b | 6.2 ab | 54.93 ab |
| Xinluzao-49 | 19.8 a | 0.36 a | 0.73 a | 1.09 a | 6.7 a | 76.84 a |
| Xinluzhong-12 | 13.3 bc | 0.29 bc | 0.64 b | 0.93 bc | 5.0 d | 43.79 b |
| Xinluzhong-15 | 14.2 bc | 0.30 bc | 0.65 b | 0.95 bc | 5.5 cd | 50.07 b |
| Xinluzao-20 | 14.9 bc | 0.32 b | 0.66 b | 0.98 b | 6.3 ab | 43.42 b |
| CCRI-12 | 14.7 bc | 0.31 bc | 0.65 b | 0.96 bc | 6.2 ab | 54.34 ab |
| P rates (µM) | | | | | | |
| 0 | 6.3 e | 0.25 e | 0.40 e | 0.64 e | 2.8 f | 15.15 d |
| 10 | 10.4 d | 0.29 d | 0.51 d | 0.80 d | 4.9 e | 29.75 c |
| 20 | 11.4 d | 0.30 d | 0.52 d | 0.82 d | 5.4 d | 40.65 c |
| 40 | 14.2 c | 0.32 c | 0.69 c | 1.01 c | 5.9 c | 61.37 b |
| 80 | 20.1 b | 0.33 b | 0.82 b | 1.15 b | 7.1 b | 79.75 a |
| 500 | 23.5 a | 0.35 a | 0.99 a | 1.33 a | 8.2 a | 81.83 a |
| Coefficient of variation (%) | 19.27 | 4.09 | 5.85 | 4.88 | 10.19 | 26.86 |
Table 2.
Root shoot ratio (RSR), root length (RL, m), projected root area (PRA, cm2), root surface area (RSA, cm2), root diameter (RD, mm), and root volume (RV, cm3) of 12 cotton genotypes grown at different P concentrations in hydroponic culture.
Table 2.
Root shoot ratio (RSR), root length (RL, m), projected root area (PRA, cm2), root surface area (RSA, cm2), root diameter (RD, mm), and root volume (RV, cm3) of 12 cotton genotypes grown at different P concentrations in hydroponic culture.
| Genotypes | RSR | RL | PRA | RSA | RD | RV |
|---|
| TM-1 | 0.46 d | 14.0 ab | 34.8 bc | 226 cd | 0.50 ab | 3.3 b |
| Yumian-21 | 0.45 de | 13.5 b | 34.4 bc | 234 c | 0.48 b | 2.8 b |
| CCRI-64 | 0.44 e | 13.9 b | 35.4 b | 248 b | 0.57 ab | 3.8 b |
| CCRI-27 | 0.50 bc | 14.3 ab | 35.3 b | 253 b | 0.62 a | 3.2 b |
| Xinluzhong-17 | 0.50 c | 11.0 c | 33.0 c | 213 d | 0.45 b | 2.3 b |
| Xinluzao-35 | 0.50 c | 12.5 c | 34.5 bc | 212 d | 0.51 ab | 3.2 b |
| Xinluzao-48 | 0.54 a | 11.7 c | 33.7 c | 221 cd | 0.54 ab | 2.9 b |
| Xinluzao-49 | 0.54 a | 15.1 a | 37.1 a | 281 a | 0.68 a | 4.2 a |
| Xinluzhong-12 | 0.48 c | 12.4 c | 34.4 bc | 219 d | 0.55 ab | 3.3 b |
| Xinluzhong-15 | 0.49 c | 11.5 c | 33.5 c | 217 d | 0.59 ab | 3.4 b |
| Xinluzao-20 | 0.52 b | 11.5 c | 33.5 c | 224 cd | 0.45 b | 2.6 b |
| CCRI-12 | 0.51 bc | 14.3 ab | 35.4 b | 244 b | 0.55 ab | 3.0 b |
| P rates (µM) | | | | | | |
| 0 | 0.61 a | 14.1 b | 35.8 b | 159 c | 0.42 b | 2.2 c |
| 10 | 0.57 b | 17.5 a | 39.5 a | 250 b | 0.45 b | 3.6 ab |
| 20 | 0.57 b | 14.5 b | 36.0 b | 273 a | 0.57 a | 3.8 a |
| 40 | 0.46 c | 12.3 c | 33.7 c | 247 b | 0.57 a | 3.4 ab |
| 80 | 0.40 d | 10.1 d | 31.45 d | 241 b | 0.6 a | 3.3 ab |
| 500 | 0.35 e | 9.5 d | 31.5 d | 225 b | 0.61 a | 2.7 bc |
| Coefficient of variation (%) | 3.44 | 13.4 | 5.4 | 12 | 20.17 | 20.3 |
Table 3.
Root length ratio (RLR), root mass ratio (RMR), root thickness (RT), and root density (RD) of 12 cotton genotypes grown at different P concentrations in hydroponic culture.
Table 3.
Root length ratio (RLR), root mass ratio (RMR), root thickness (RT), and root density (RD) of 12 cotton genotypes grown at different P concentrations in hydroponic culture.
| Genotypes | RLR | RMR | RT | RD |
|---|
| TM-1 | 16.92 a | 0.314 d | 4.79 ab | 0.090 ab |
| Yumian-21 | 16.39 a | 0.308 de | 5.46 a | 0.107 ab |
| CCRI-64 | 17.24 a | 0.303 e | 3.78 b | 0.071 b |
| CCRI-27 | 16.60 a | 0.331 bc | 4.72 ab | 0.099 ab |
| Xinluzhong-17 | 12.26 c | 0.329 c | 5.42 a | 0.154 a |
| Xinluzao-35 | 15.04 ab | 0.329 c | 4.08 ab | 0.099 ab |
| Xinluzao-48 | 12.67 bc | 0.347 a | 4.14 ab | 0.126 a |
| Xinluzao-49 | 15.27 ab | 0.345 a | 3.69 b | 0.088 b |
| Xinluzhong-12 | 14.63 bc | 0.323 c | 4.39 ab | 0.101ab |
| Xinluzhong-15 | 13.48 bc | 0.328 c | 3.51 b | 0.096 ab |
| Xinluzao-20 | 12.47 c | 0.339 ab | 4.54 ab | 0.138 a |
| CCRI-12 | 16.27 a | 0.334 bc | 4.89 ab | 0.108 ab |
| P rates (µM) | | | | |
| 0 | 21.97 a | 0.379 a | 6.91 a | 0.125 ab |
| 10 | 21.77 a | 0.364 b | 4.90 b | 0.082 bc |
| 20 | 17.73 b | 0.362 b | 4.01 bc | 0.084 c |
| 40 | 12.20 c | 0.313 c | 3.96 bc | 0.104 bc |
| 80 | 8.79 d | 0.285 d | 3.14 c | 0.104 bc |
| 500 | 7.16 e | 0.261 e | 3.78 c | 0.142 a |
| Coefficient of variation (%) | 12.53 | 3.22 | 18.19 | 20.51 |
Table 4.
Photosynthetic rate (Pn, µmol CO2 m−2 s−1), stomatal conductance (gs, mmol H2O m−2 s−1), transpiration rate (E, mmol m−2 s−1), intercellular CO2 concentration (Ci, µmol CO2 mol−1 air), and SPAD value of 12 cotton genotypes grown at different P concentrations in hydroponic culture.
Table 4.
Photosynthetic rate (Pn, µmol CO2 m−2 s−1), stomatal conductance (gs, mmol H2O m−2 s−1), transpiration rate (E, mmol m−2 s−1), intercellular CO2 concentration (Ci, µmol CO2 mol−1 air), and SPAD value of 12 cotton genotypes grown at different P concentrations in hydroponic culture.
| Genotypes | Pn | Gs | E | Ci | SPAD |
|---|
| TM-1 | 5.5 bcd | 0.291 c | 2.59 b | 312 a | 34.4 b |
| Yumian-21 | 5.6 bc | 0.288 d | 2.59 b | 310 ab | 34.4 b |
| CCRI-64 | 5.2 d | 0.288 d | 2.59 b | 313 a | 34.3 b |
| CCRI-27 | 5.5 bcd | 0.290 cd | 2.59 b | 309 ab | 34.4 b |
| Xinluzhong-17 | 5.7 bc | 0.292 bc | 2.60 a | 298 c | 34.3 b |
| Xinluzao-35 | 5.3 cd | 0.291 c | 2.59 b | 315 a | 34.8 b |
| Xinluzao-48 | 5.8 ab | 0.294 b | 2.60 a | 303 c | 35.9 b |
| Xinluzao-49 | 6.3 a | 0.299 a | 2.60 a | 291 c | 36.6 a |
| Xinluzhong-12 | 5.6 bc | 0.292 bc | 2.59 b | 311 ab | 34.9 b |
| Xinluzhong-15 | 5.6 bc | 0.292 bc | 2.59 b | 306 b | 34.6 b |
| Xinluzao-20 | 5.8 ab | 0.294 b | 2.59 b | 296 c | 34.5 b |
| CCRI-12 | 5.5 bcd | 0.292 bc | 2.59 b | 316 a | 34.7 b |
| P rates (µM) | | | | | |
| 0 | 3.5 f | 0.281 f | 2.58 c | 337 a | 30.5 f |
| 10 | 4.7 e | 0.285 e | 2.59 b | 324 b | 33.6 e |
| 20 | 5.1 d | 0.288 d | 2.59 b | 315 c | 34.8 d |
| 40 | 6.1 c | 0.294 c | 2.60 a | 301 d | 35.8 c |
| 80 | 6.9 b | 0.298 b | 2.60 a | 286 e | 36.7 b |
| 500 | 7.3 a | 0.301 a | 2.60 a | 281 e | 37.4 a |
| Coefficient of variation (%) | 6.6 | 8.6 | 4.16 | 6 | 5.6 |
Table 5.
Shoot nitrogen (SN%), root nitrogen (RN%), total nitrogen (TN%), shoot phosphorus (SP%), root phosphorus (RP%), and total phosphorus (TP%) of 12 cotton genotypes grown at different P concentrations in hydroponic culture.
Table 5.
Shoot nitrogen (SN%), root nitrogen (RN%), total nitrogen (TN%), shoot phosphorus (SP%), root phosphorus (RP%), and total phosphorus (TP%) of 12 cotton genotypes grown at different P concentrations in hydroponic culture.
| Genotypes | SN% | RN% | TN% | SP% | RP% | TP% |
|---|
| TM-1 | 2.5 b | 1.4 ab | 3.9 b | 0.51 b | 0.33 b | 0.85 b |
| Yumian-21 | 2.5 b | 1.4 ab | 3.9 b | 0.48 b | 0.31 b | 0.78 b |
| CCRI-64 | 2.6 b | 1.4 ab | 4.0 b | 0.55 b | 0.38 b | 0.93 ab |
| CCRI-27 | 2.5 b | 1.4 ab | 4.0 b | 0.62 a | 0.45 a | 1.08 a |
| Xinluzhong-17 | 2.4 b | 1.2 b | 3.7 b | 0.39 b | 0.22 c | 0.61 b |
| Xinluzao-35 | 2.5 b | 1.3 b | 3.8 b | 0.51 b | 0.36 b | 0.87 b |
| Xinluzao-48 | 2.7 b | 1.3 b | 4.0 b | 0.63 a | 0.46 a | 1.08 a |
| Xinluzao-49 | 3.2 a | 1.5 a | 4.7 a | 0.69 a | 0.49 a | 1.18 a |
| Xinluzhong-12 | 2.6 b | 1.4 ab | 4.0 b | 0.66 a | 0.47 a | 1.13 a |
| Xinluzhong-15 | 2.7 b | 1.6 a | 4.2 b | 0.62 a | 0.45 a | 1.07 a |
| Xinluzao-20 | 2.6 b | 1.3 b | 3.9 b | 0.50 b | 0.33 b | 0.83 b |
| CCRI-12 | 2.6 b | 1.4 ab | 4.0 b | 0.63 a | 0.46 a | 1.08 a |
| P rates (µM) | | | | | | |
| 0 | 1.4 e | 0.7 e | 2.1 e | 0.35 d | 0.18 d | 0.53 d |
| 10 | 2.1 d | 1.0 d | 3.1 d | 0.48 c | 0.31 c | 0.79 c |
| 20 | 2.6 c | 1.3 c | 3.9 c | 0.53 bc | 0.36 bc | 0.90 bc |
| 40 | 3.0 d | 1.5 b | 4.5 b | 0.65 ab | 0.48 ab | 1.12 ab |
| 80 | 3.3 a | 1.9 a | 5.2 a | 0.68 a | 0.51 a | 1.18 a |
| 500 | 3.3a | 1.9 a | 5.2 a | 0.70 a | 0.53 a | 1.24 a |
| Coefficient of variation (%) | 7.1 | 11.7 | 7.6 | 16.91 | 14.7 | 15.64 |
Table 6.
Root phosphorus accumulation (RPA), shoot phosphorus accumulation (SPA), total plant phosphorus accumulation (TPA), phosphorus utilization efficiency (PUtE, g DW mg−1 P), phosphorus uptake efficiency (PUpE, mg P g−1 RDW), and physiological phosphorus-use efficiency (PPUE, g2 DW mg−1 P) of 12 cotton genotypes grown at different P concentrations in hydroponic culture.
Table 6.
Root phosphorus accumulation (RPA), shoot phosphorus accumulation (SPA), total plant phosphorus accumulation (TPA), phosphorus utilization efficiency (PUtE, g DW mg−1 P), phosphorus uptake efficiency (PUpE, mg P g−1 RDW), and physiological phosphorus-use efficiency (PPUE, g2 DW mg−1 P) of 12 cotton genotypes grown at different P concentrations in hydroponic culture.
| Genotypes | RPA | SPA | TPA | PUtE | PUpE | PPUE |
|---|
| TM-1 | 0.08 de | 0.31 bc | 0.39 bc | 1.2 b | 1.4 a | 1.3 c |
| Yumian-21 | 0.07 e | 0.29 c | 0.36 c | 1.3 a | 1.3 b | 1.5 a |
| CCRI-64 | 0.07 e | 0.30 bc | 0.38 bc | 1.2 c | 1.4 a | 1.3 c |
| CCRI-27 | 0.09 c | 0.32 bc | 0.41 bc | 1.2 b | 1.3 b | 1.3 c |
| Xinluzhong-17 | 0.09 c | 0.32 bc | 0.42 bc | 1.3 a | 1.3 b | 1.4 b |
| Xinluzao-35 | 0.08 de | 0.30 bc | 0.38 bc | 1.3 a | 1.2 c | 1.4 b |
| Xinluzao-48 | 0.11 b | 0.34 b | 0.45 b | 1.2 b | 1.3 b | 1.3 c |
| Xinluzao-49 | 0.12 a | 0.38 a | 0.49 a | 1.3 a | 1.4 a | 1.5 a |
| Xinluzhong-12 | 0.09 c | 0.31 bc | 0.39 bc | 1.2 b | 1.3 b | 1.4 b |
| Xinluzhong-15 | 0.08 de | 0.30 bc | 0.39 bc | 1.3 a | 1.2 c | 1.4 b |
| Xinluzao-20 | 0.10 b | 0.33 b | 0.43 b | 1.2 b | 1.3 b | 1.3 c |
| CCRI-12 | 0.09 c | 0.32 bc | 0.41 b | 1.2 b | 1.3 b | 1.4 b |
| P rates (µM) | | | | | |
| 0 | 0.04 f | 0.14 e | 0.18 e | 1.2 d | 0.8 e | 1.1 d |
| 10 | 0.08 e | 0.23 d | 0.31 d | 1.1 d | 1.1 d | 1.1 d |
| 20 | 0.09 d | 0.24 d | 0.33 d | 1.1 d | 1.1 d | 1.2 d |
| 40 | 0.10 c | 0.34 c | 0.44 c | 1.3 c | 1.4 c | 1.4 c |
| 80 | 0.11 b | 0.42 b | 0.53 b | 1.4 b | 1.6 b | 1.6 b |
| 500 | 0.12 a | 0.53 a | 0.66 a | 1.5 a | 1.9 a | 1.8 a |
| Coefficient of variation (%) | 7.6 | 8.26 | 7.59 | 15.10 | 14.57 | 15.24 |
Table 7.
Relationship between shoot dry weight (SDW), root dry weight (RDW), photosynthesis (Pn), phosphorus utilization efficiency (PUtE), and phosphorus uptake efficiency (PUpE).
Table 7.
Relationship between shoot dry weight (SDW), root dry weight (RDW), photosynthesis (Pn), phosphorus utilization efficiency (PUtE), and phosphorus uptake efficiency (PUpE).
| | SDW | RDW | Pn | PUtE | PUpE |
|---|
| SDW | 1 | 0.78 * | 0.92 * | 0.73 * | 0.97 * |
| RDW | 0.78 * | 1 | 0.80 * | 0.42 * | 0.71 * |
| Pn | 0.92 * | 0.80 * | 1 | 0.58 * | 0.91 * |
| PUtE | 0.73 * | 0.42 * | 0.58 * | 1 | 0.58 * |
| PUpE | 0.97 * | 0.71 * | 0.91 * | 0.58 * | 1 |
Table 8.
Classification of cotton genotypes (grown at P starvation; 0 μM KH2PO4) based on their index scores of various parameters into efficient (E), medium (M), and inefficient (I) scoring genotypes.
Table 8.
Classification of cotton genotypes (grown at P starvation; 0 μM KH2PO4) based on their index scores of various parameters into efficient (E), medium (M), and inefficient (I) scoring genotypes.
| Genotypes | SDW | RDW | Pn | PUtE | PUpE | Total Score |
|---|
| TM-1 | M | M | M | M | M | 10 |
| Yumian-21 | M | I | M | E | I | 9 |
| CCRI-64 | I | I | I | M | M | 7 |
| CCRI-27 | M | M | M | M | M | 10 |
| Xinluzhong-17 | M | M | E | M | M | 11 |
| Xinluzao-35 | M | M | M | E | I | 10 |
| Xinluzao-48 | E | E | M | I | E | 12 |
| Xinluzao-49 | E | E | E | M | M | 13 |
| Xinluzhong-12 | M | M | M | M | M | 10 |
| Xinluzhong-15 | M | M | M | E | I | 10 |
| Xinluzao-20 | M | M | M | I | E | 10 |
| CCRI-12 | M | M | M | I | E | 10 |
Table 9.
Classification of cotton genotypes (grown at P deficient; 10 μM KH2PO4) based on their index scores of various parameters into efficient (E), medium (M), and inefficient (I) scoring genotypes.
Table 9.
Classification of cotton genotypes (grown at P deficient; 10 μM KH2PO4) based on their index scores of various parameters into efficient (E), medium (M), and inefficient (I) scoring genotypes.
| Genotypes | SDW | RDW | Pn | PUtE | PUpE | Total Score |
|---|
| TM-1 | M | M | M | M | M | 10 |
| Yumian-21 | M | I | I | E | I | 8 |
| CCRI-64 | I | I | I | I | M | 6 |
| CCRI-27 | M | M | M | M | M | 10 |
| Xinluzhong-17 | M | M | E | M | M | 11 |
| Xinluzao-35 | M | M | M | M | I | 9 |
| Xinluzao-48 | E | E | E | I | E | 13 |
| Xinluzao-49 | E | E | E | M | M | 13 |
| Xinluzhong-12 | M | M | M | I | E | 10 |
| Xinluzhong-15 | M | M | M | M | I | 9 |
| Xinluzao-20 | M | M | M | I | E | 10 |
| CCRI-12 | M | M | M | I | E | 10 |
Table 10.
Classification of cotton genotypes (grown at P sufficient; 500 μM KH2PO4) based on their index scores of various parameters into efficient (E), medium (M), and inefficient (I) scoring genotypes.
Table 10.
Classification of cotton genotypes (grown at P sufficient; 500 μM KH2PO4) based on their index scores of various parameters into efficient (E), medium (M), and inefficient (I) scoring genotypes.
| Genotypes | SDW | RDW | Pn | PUtE | PUpE | Total Score |
|---|
| TM-1 | M | M | M | I | E | 10 |
| Yumian-21 | M | I | M | M | M | 9 |
| CCRI-64 | M | I | I | M | E | 9 |
| CCRI-27 | M | M | M | M | M | 10 |
| Xinluzhong-17 | M | M | M | M | M | 10 |
| Xinluzao-35 | M | M | M | E | I | 10 |
| Xinluzao-48 | M | E | M | M | I | 10 |
| Xinluzao-49 | E | E | E | E | E | 15 |
| Xinluzhong-12 | M | M | M | M | M | 10 |
| Xinluzhong-15 | M | M | E | E | M | 12 |
| Xinluzao-20 | M | M | M | M | M | 10 |
| CCRI-12 | M | M | M | M | M | 10 |
Table 11.
Scoring of genotypes grown at P starvation (P; 0 μM KH2PO4), P deficient (P; 10 μM KH2PO4) and P sufficient (P; 500 μM KH2PO4) concentration under hydroponic conditions.
Table 11.
Scoring of genotypes grown at P starvation (P; 0 μM KH2PO4), P deficient (P; 10 μM KH2PO4) and P sufficient (P; 500 μM KH2PO4) concentration under hydroponic conditions.
| Genotypes | Score at 0 μM/15 | Score at 10 μM/15 | Score at 500 μM/15 | Total Score/45 |
|---|
| TM-1 | 10 | 10 | 10 | 30 |
| Yumian-21 | 9 | 8 | 9 | 26 |
| CCRI-64 | 7 | 6 | 9 | 22 |
| CCRI-27 | 10 | 10 | 10 | 30 |
| Xinluzhong-17 | 11 | 11 | 10 | 32 |
| Xinluzao-35 | 10 | 9 | 10 | 29 |
| Xinluzao-48 | 12 | 13 | 10 | 35 |
| Xinluzao-49 | 13 | 13 | 15 | 41 |
| Xinluzhong-12 | 10 | 10 | 10 | 30 |
| Xinluzhong-15 | 10 | 9 | 12 | 31 |
| Xinluzao-20 | 10 | 10 | 10 | 30 |
| CCRI-12 | 10 | 10 | 10 | 30 |
Table 12.
Principal component analysis of morphophysiological traits under high and low nitrogen either from ammonium nitrate or nitrate.
Table 12.
Principal component analysis of morphophysiological traits under high and low nitrogen either from ammonium nitrate or nitrate.
| Traits | PC1 | PC2 |
|---|
| Shoot dry weight | 0.56 | 0.22 |
| Root dry weight | 0.55 | 0.08 |
| Photosynthesis | 0.54 | 0.21 |
| P utilization efficiency | 0.27 | −0.62 |
| P uptake efficiency | −0.15 | 0.72 |
| Eigen value | 2.84 | 1.58 |
| Variance contribution rate | 56.9% | 31.5% |
| Cumulative percentage | 56.9% | 88.4% |






{kind=link}
{kind=link}
{kind=link}