A Dual-Purpose Model for Spring-Sown Oats in Cold Regions of Northern China

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Oat Species
2.3. Experimental Design
2.4. Oat Sowing and Management
2.5. Growth Stage
2.6. Plant Height, Tiller Density, Aboveground Biomass, Seed Yield, and Grain Harvested Index
2.7. Forage Quality
2.8. Data Collected in Field Demonstration
2.9. Data Analysis
3. Results
3.1. Weather
3.2. Agronomic Characteristics and Forage Quality
3.3. Effect of Cutting on Agronomic Characteristics
3.4. Effect of Cutting on Oat Growth and Production across the Whole Growing Season
3.5. Effect of Cutting on Growth Dynamics of Oat Species
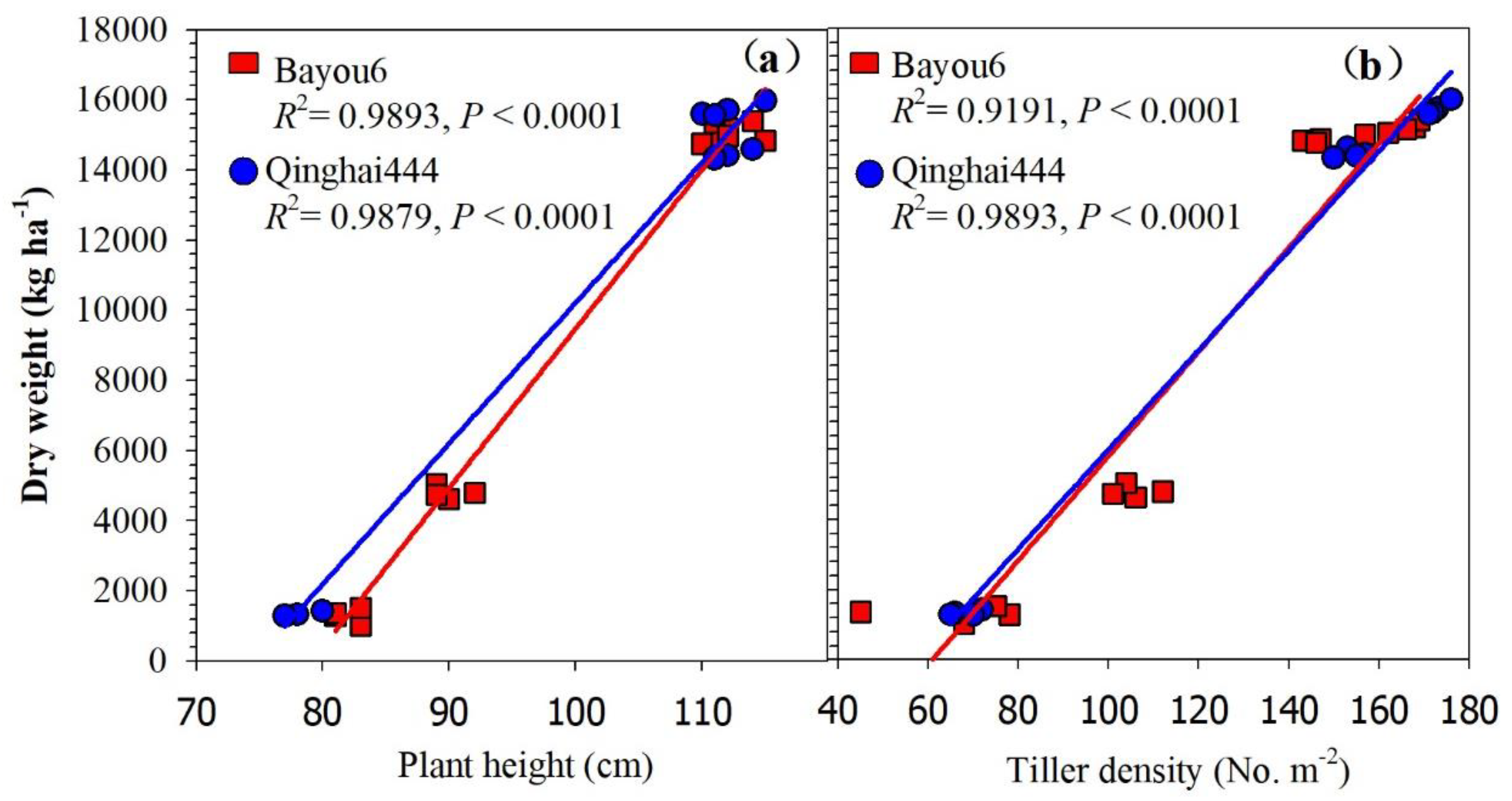
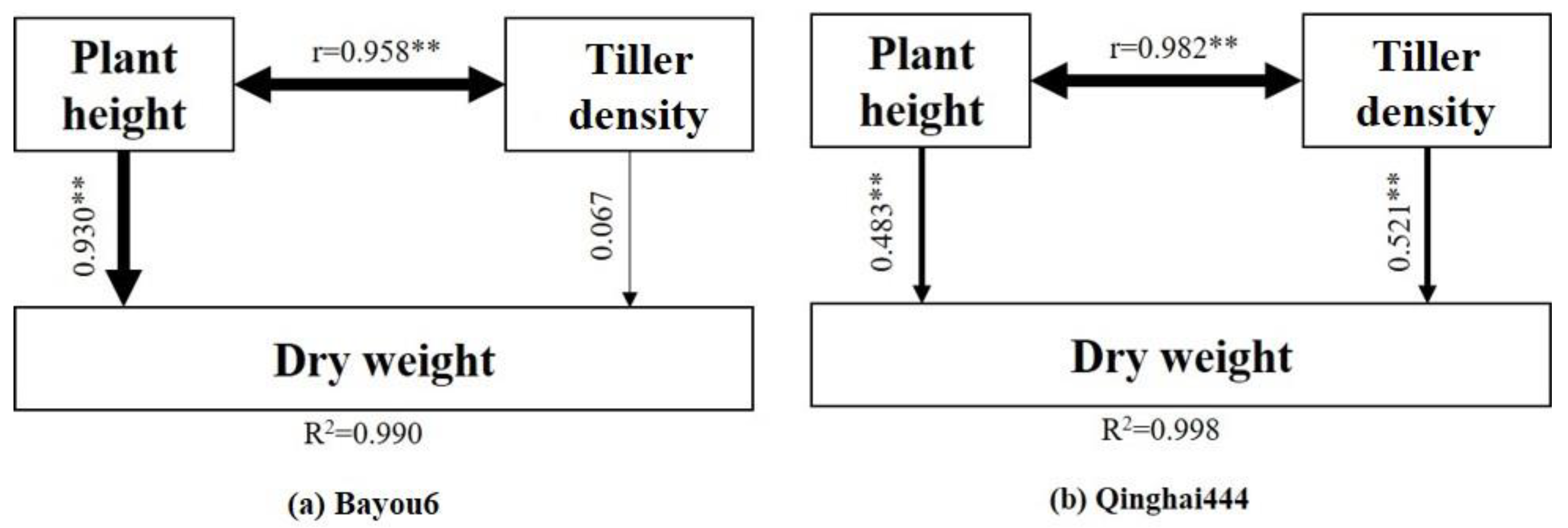
3.6. Dry Weight of Re-Growth and Its Driving Factors
3.7. Effects of Cutting on Grain Harvested Index
3.8. Field Demonstration for the Dual-Purpose Model
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Miller, G.L.; Joost, R.E.; Harrison, S.A. Forage and grain yields of wheat and triticale as affected by forage management-practices. Crop Sci. 1993, 33, 1070–1075. [Google Scholar] [CrossRef]
- Harrison, M.T.; Evans, J.R.; Dove, H.; Moore, A.D. Dual-purpose cereals: Can the relative influences of management and environment on crop recovery and grain yield be dissected? Crop Pasture Sci. 2011, 62, 930–946. [Google Scholar] [CrossRef]
- Lyu, D.; Doce, R.R.; Juskiw, P.; Zhou, G.; Baron, V.S. Diverse grain-filling dynamics affect harvest management of forage barley and triticale cultivars. Agron. J. 2018, 110, 1017–1027. [Google Scholar] [CrossRef]
- Moore, A.D.; Bell, L.W.; Revell, D.K. Feed gaps in mixed-farming systems: Insights from the Grain & Graze program. Anim. Prod. Sci. 2009, 49, 736–748. [Google Scholar]
- Dove, H.; McMullen, K.G. Diet selection, herbage intake and liveweight gain in young sheep grazing dual-purpose wheats and sheep responses to mineral supplements. Anim. Prod. Sci. 2009, 49, 749–758. [Google Scholar] [CrossRef]
- Dove, H.; Kirkegaard, J. Using dual-purpose crops in sheep-grazing systems. J. Sci. Food Agric. 2014, 94, 1276–1283. [Google Scholar] [CrossRef]
- Davidson, J.L.; Jones, D.B.; Christian, K.R. Winter feed production and grain yield in mixtures of spring and winter wheats. Aust. J. Agric. Res. 1990, 41, 1–18. [Google Scholar] [CrossRef]
- Francia, E.; Pecchioni, N.; Nicosia, O.L.D.; Paoletta, G.; Taibi, L.; Franco, V.; Odoardi, M.; Stanca, A.M.; Delogu, G. Dual-Purpose barley and oat in a Mediterranean environment. Field Crop. Res. 2006, 99, 158–166. [Google Scholar] [CrossRef]
- Ates, S.; Cicek, H.; Gultekin, I.; Yigezu, Y.A.; Keser, M.; Filley, S.J. Bio-Economic analysis of dual-purpose management of winter cereals in high and low input production systems. Field Crop. Res. 2018, 227, 56–66. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, Z.; Zhang, X.; Song, S.; Yang, Z.; Yang, Y. Influence of mowing time on yield and quality of spring and autumn sown oat hay. Acta Pratacult. Sin. 2016, 25, 124–135. [Google Scholar]
- Zhou, Q. Cultivation and Management of Oats in Alpine Region; Zhou, Q., Ed.; Jiangsu Phoenix Science and Technology Press: Nanjing, China, 2014. [Google Scholar]
- Tian, C.; Zhang, B. Practical Technology of Oats; China Agricultural University press: Beijing, China, 2016. [Google Scholar]
- Zhang, W.; Hou, L.; Yang, J.; Song, S.; Mao, X.; Zhang, Q.; Bai, W.; Pan, Q.; Zhou, Q. Establishment and management of alfalfa pasture in cold regions of China. Chin. Sci. Bull. 2018, 63, 1651–1663. [Google Scholar] [CrossRef] [Green Version]
- Fang, J.; Jing, H.; Zhang, W.; Gao, S.; Duan, Z.; Wang, H.; Zhong, J.; Pan, Q.; Zhao, K.; Bai, W.; et al. The concept of Grass-Based Livestock Husbandry and its practice in Hulun Buir, Inner Mongolia. Chin. Sci. Bull. 2018, 63, 1619–1631. [Google Scholar] [CrossRef]
- Zhou, Q.; Gou, X.; Tian, L.; Chen, Y.; Gao, S.; Bai, W.; Zhang, W. Performances of early and late maturing oat varieties in cold regions. Chin. Sci. Bull. 2018, 63, 1722–1730. [Google Scholar] [CrossRef] [Green Version]
- Tao, J.; Hou, L.; Yang, J.; Song, S.; Bai, W.; Pan, Q.; Yin, S.; Hao, J.; Zhang, W. Comparative studies on seed germination characteristics and indicators of oat varieties under salt stress. Pratacult. Sci. 2018, 35, 2414–2421. [Google Scholar]
- Wu, Y.; Zhang, B.; Wang, X.; Ge, J.; Li, Y.; Zuo, W.; Dong, Z.; Tian, C. Evaluation on productivity performance and genetic diversity of 20 major oat cultivars in north China. Southwest China J. Agric. Sci. 2018, 31, 448–456. [Google Scholar]
- Rohweder, D.A.; Barnes, R.F.; Jorgensen, N. Proposed hay grading standards based on laboratory analyses for evaluating quality. J. Anim. Sci. 1978, 47, 747–759. [Google Scholar] [CrossRef]
- Silva, A.B.; Lira, M.A.; Dubeux, J.C.B.; Figueiredo, M.D.B.; Vicentin, R.P. Soil litter stock and fertility after planting leguminous shrubs and forage trees on degraded signal grass pasture. Revista Brasileira De Ciência Do Solo 2013, 37, 502–511. [Google Scholar] [CrossRef]
- Nie, Z.; Tremblay, G.F.; Belanger, G.; Berthiaume, R.; Castonguay, Y.; Bertrand, A.; Michaud, R.; Allard, G.; Han, J. Near-infrared reflectance spectroscopy prediction of neutral detergent-soluble carbohydrates in timothy and alfalfa. J. Dairy Sci. 2009, 92, 1702–1711. [Google Scholar] [CrossRef]
- Zhang, G.; Ma, H.; Shao, X.; Wang, J.; Shen, Z.; Fu, G. A comparative study of yield and nutritive value of nine imported oat varieties in the valley region of Tibet, China. Acta Pratacult. Sin. 2019, 28, 121–131. [Google Scholar]
- Guo, X.; Tian, Z.; Liang, D.; Fu, B.; Lan, J. Path-Coefficient analysis of seed yield with main agronomic aharacters in 11 oat varieties. Acta Agrestia Sin. 2017, 25, 142–147. [Google Scholar]
- Wu, Y.; Li, Z. Analysis of forage oat at nutrient content dynamic in different growth stages. China Dairy Cattle 2015, 288, 60–63. [Google Scholar]
- Liu, J.; Shi, Y.; Wu, X.; Guo, P.; Fang, Z.; Jia, H.; Zhang, Y.; Chi, H. Growth dynamics and optimum harvest time of forage grass oats in northern Shanxi. Chin. Agric. Sci. Bull. 2019, 35, 107–111. [Google Scholar]
- Nan, M.; Ma, N.; Liu, Y.; Ren, S.; Bian, F. Genetic diversity analysis on agronomic characteristics of oat germplasms. Agric. Res. Arid Areas 2015, 33, 262–267. [Google Scholar]
- Zhou, P.; Zhao, J.; Yan, H.; Lan, X.; Peng, Y. Effects of sowing date, sowing rate and fertilizer rate on grain yield and agronomic characteristics of Avena sativa. Pratacult. Sci. 2015, 32, 433–441. [Google Scholar]
- Bao, G.; Zhou, Q.; Han, Z. Effects of nitrogen and potassium fertilizer on yield and quality of oat. Pratacult. Sci. 2008, 25, 48–53. [Google Scholar]
- Chen, H.; Wang, H.; Du, G. Impacts of clipping time, clipping intensity and fertilization on plant compensation of Avena sativa. Acta Bot. Boreali-Occident. Sin. 2003, 23, 969–975. [Google Scholar]
- Steve, F. Drought Advisory Series: Oat Hay and Straw; Washington State University Extension: Washington, DC, USA, 2005; p. EM4918E. [Google Scholar]
- Belsky, A.J.; Carson, W.P.; Jensen, C.L.; Fox, G.A. Overcompensation by plants - herbivore optimization or red herring. Evol. Ecol. 1993, 7, 109–121. [Google Scholar] [CrossRef]
- Kelman, W.M.; Dove, H. Growth and phenology of winter wheat and oats in a dual-purpose management system. Crop Pasture Sci. 2009, 60, 921–932. [Google Scholar] [CrossRef]
- Wallace, L.L.; Mcnaughton, S.J.; Coughenour, M.B. Compensatory photosynthetic responses of three African graminoids to different fertilization, watering, and clipping regimes. Bot. Gaz. 1984, 145, 151–156. [Google Scholar] [CrossRef]
- Richards, J.H.; Caldwell, M.M. Soluble carbohydrates, concurrent photosynthesis and efficiency in regrowth following defoliation, a field-study with Agropyron species. J. Appl. Ecol. 1985, 22, 907–920. [Google Scholar] [CrossRef]
- Jones, G.B.; Alpuerto, J.B.; Tracy, B.F.; Fukao, T. Physiological effect of cutting height and high temperature on regrowth vigor in Orchardgrass. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Erice, G.; Sanz-Saez, A.; Aranjuelo, I.; Irigoyen, J.J.; Aguirreolea, J.; Avice, J.C.; Sanchez-Diaz, M. Photosynthesis, N2 fixation and taproot reserves during the cutting regrowth cycle of alfalfa under elevated CO2 and temperature. J. Plant Physiol. 2011, 168, 2007–2014. [Google Scholar] [CrossRef] [PubMed]
- Meuriot, F.; Avice, J.C.; Simon, J.C.; Laine, P.; Decau, M.L.; Ourry, A. Influence of initial organic N reserves and residual leaf area on growth, N uptake, N partitioning and N storage in Alfalfa (Medicago sativa) during post-cutting regrowth. Ann. Bot. 2004, 94, 311–321. [Google Scholar] [CrossRef] [PubMed]
- Pumphrey, F.V. Semidwarf winter wheat response to early spring clipping and grazing. Agron. J. 1970, 62, 641–643. [Google Scholar] [CrossRef]
- Lyon, D.J.; Baltensperger, D.D.; Siles, M. Wheat grain and forage yields are affected by planting and harvest dates in the central Great Plains. Crop Sci. 2001, 41, 488–492. [Google Scholar] [CrossRef]
- Chen, X.; Zhao, B.; Ji, C.; Zhu, B.; Wang, R. Effect of defoliation timing on the yield and quality of six-row barley forage and grain. J. Triticeae Crop. 2017, 37, 409–413. [Google Scholar]
- Virgona, J.M.; Gummer, F.A.J.; Angus, J.F. Effects of grazing on wheat growth, yield, development, water use, and nitrogen use. Aust. J. Agric. Res. 2006, 57, 1307–1319. [Google Scholar] [CrossRef]
- Kelman, W.M.; Dove, H. Effects of a spring-sown brassica crop on lamb performance and on subsequent establishment and grain yield of dual-purpose winter wheat and oat crops. Aust. J. Exp. Agric. 2007, 47, 815–824. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Xieertala |
|---|---|
| Frost-free period (d) | 110–125 |
| Growing season (Month-Month) | May–August |
| Accumulated temperature ≥0 °C | 2279–2647 |
| Accumulated temperature ≥5 °C | 2159–2562 |
| Accumulated temperature ≥10 °C | 1856–2274 |
| Mean annual temperature (°C) | −1.3 |
| Mean annual precipitation (mm) | 348.4 |
| Soil type | Chestnut soil |
| Oat Species | 1000-Grain Weight g | Germination Rate % |
|---|---|---|
| Avena nuda L. cv. Bayou6 | 22.27 ± 0.10 | 98.5 ± 0.8 |
| Avena sativa L. cv. Qinghai444 | 33.15 ± 0.13 | 86.3 ± 2.0 |
| Parameters | Year | Apr. | May | Jun. | Jul. | Aug. | Sep. | Average/Sum |
|---|---|---|---|---|---|---|---|---|
| Monthly Precipitation (mm) | 2017 | 4.8 | 6.7 | 15.3 | 36.4 | 37.3 | 17.1 | 117.6 |
| 2018 | 1.3 | 32.9 | 75.9 | 109.2 | 59.5 | 47.4 | 326.2 | |
| 30-year mean | 13.7 | 25.4 | 55.6 | 94.6 | 84.9 | 36.4 | 310.6 | |
| Mean monthly Temperature (°C) | 2017 | 2.4 | 9.6 | 19.3 | 23.4 | 19.4 | 9.5 | 13.9 |
| 2018 | 3.3 | 12.4 | 16.6 | 21.5 | 17.9 | 11.2 | 13.8 | |
| 30-year mean | 1.9 | 10.9 | 18.1 | 20 | 17.7 | 10.3 | 13.2 |
| Oat Bayou6 | Oat Qinghai444 | |||||
|---|---|---|---|---|---|---|
| Treatments | Vegetative Growth (days) | Reproductive Growth (days) | Whole Growth Period (days) | Vegetative Growth (days) | Reproductive Growth (days) | Whole Growth Period (days) |
| No cutting (CK) | 40 ± 1d | 30 ± 2a | 70 ± 2d | 40 ± 2d | 30 ± 2a | 70 ± 2d |
| 30 days cutting | 68 ± 1c | 32 ± 1a | 100 ± 1c | 67 ± 2c | 33 ± 2a | 100 ± 2c |
| 40 days cutting | 77 ± 2b | 33 ± 2a | 110 ± 2b | 76 ± 1b | 33 ± 1a | 109 ± 1b |
| 50 days cutting | 83 ± 2a | 31 ± 2a | 114 ± 2a | 84 ± 2a | 33 ± 3a | 117 ± 2a |
| Species | Year | The First Forage Harvest | The Second Harvest | Overall Harvest | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Emergence Days | Plant Height | Tiller Density | Dry Weight | Plant Height | Tiller Density | Dry Weight | Dry Weight | Seed Yield | ||
| days | cm | No. m−2 | kg ha−1 | cm | No. m-2 | kg ha−1 | kg ha−1 | kg ha−1 | ||
| Bayou6 | 2017 | 40 | 82.4 ± 1.9 a | 738 ± 18 a | 4819 ± 158 a | 85.3 ± 1.8 c | 678 ± 12 c | 9967 ± 558 bc | 14,786 ± 784 a | 4436 ± 493 bc |
| Un-cut (CK) | ─ | ─ | ─ | 98.2 ± 1.7 ab | 738 ± 18 ab | 12,151 ± 449 a | 12,151 ± 498 b | 5724 ± 372 a | ||
| 2018 | 30 | 71.1 ± 1.5 b | 726 ± 12 a | 4011 ± 147 b | 95.3 ± 1.3 b | 756 ± 18 a | 10,677 ± 347 b | 14,688 ± 487 a | 5848 ± 419 a | |
| Un-cut (CK) | ─ | ─ | ─ | 101.3 ± 1.8 a | 726 ± 18 ab | 11,798 ± 445 a | 11,798 ± 698 b | 5249 ± 568 ab | ||
| Qinghai444 | 2017 | 40 | 79.3 ± 1.8 a | 708 ± 18 a | 4725 ± 189 a | 83.3 ± 1.6 c | 672 ± 12 c | 9435 ± 434 c | 14,160 ± 454 a | 3635 ± 434 c |
| Un-cut (CK) | ─ | ─ | ─ | 97.4 ± 2.1 ab | 708 ± 18 b | 10,231 ± 567 bc | 10,231 ± 675 c | 4846 ± 469 b | ||
| 2018 | 30 | 72.1 ± 1.3 b | 720 ± 12 a | 4034 ± 166 b | 94.2 ± 1.4 b | 726 ± 18 ab | 9887 ± 466 bc | 13,930 ± 587 a | 4935 ± 387 b | |
| Un-cut (CK) | ─ | ─ | ─ | 98.3 ± 1.8 ab | 696 ± 12 bc | 10,583 ± 498 b | 10,583 ± 693 c | 4338 ± 438 bc | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, J.; Hou, L.; Bai, W.; Yan, J.; Hao, J.; Tao, J.; Luo, Y.; Zhang, J.; Zhang, W.-H. A Dual-Purpose Model for Spring-Sown Oats in Cold Regions of Northern China. Agronomy 2019, 9, 721. https://doi.org/10.3390/agronomy9110721
Yang J, Hou L, Bai W, Yan J, Hao J, Tao J, Luo Y, Zhang J, Zhang W-H. A Dual-Purpose Model for Spring-Sown Oats in Cold Regions of Northern China. Agronomy. 2019; 9(11):721. https://doi.org/10.3390/agronomy9110721
Chicago/Turabian StyleYang, Jie, Longyu Hou, Wenming Bai, Jingyun Yan, Jianxi Hao, Jin Tao, Yingluo Luo, Jianmin Zhang, and Wen-Hao Zhang. 2019. "A Dual-Purpose Model for Spring-Sown Oats in Cold Regions of Northern China" Agronomy 9, no. 11: 721. https://doi.org/10.3390/agronomy9110721