Can Genetic Progress for Drought Tolerance in Popcorn Be Achieved by Indirect Selection?

 ,
, 
 ,
,  , , , ,
, , , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material, Experimental Design and Cultural Treatment
2.2. Traits Evaluated
2.3. Analysis of Variance and Estimates of Genetic Parameters
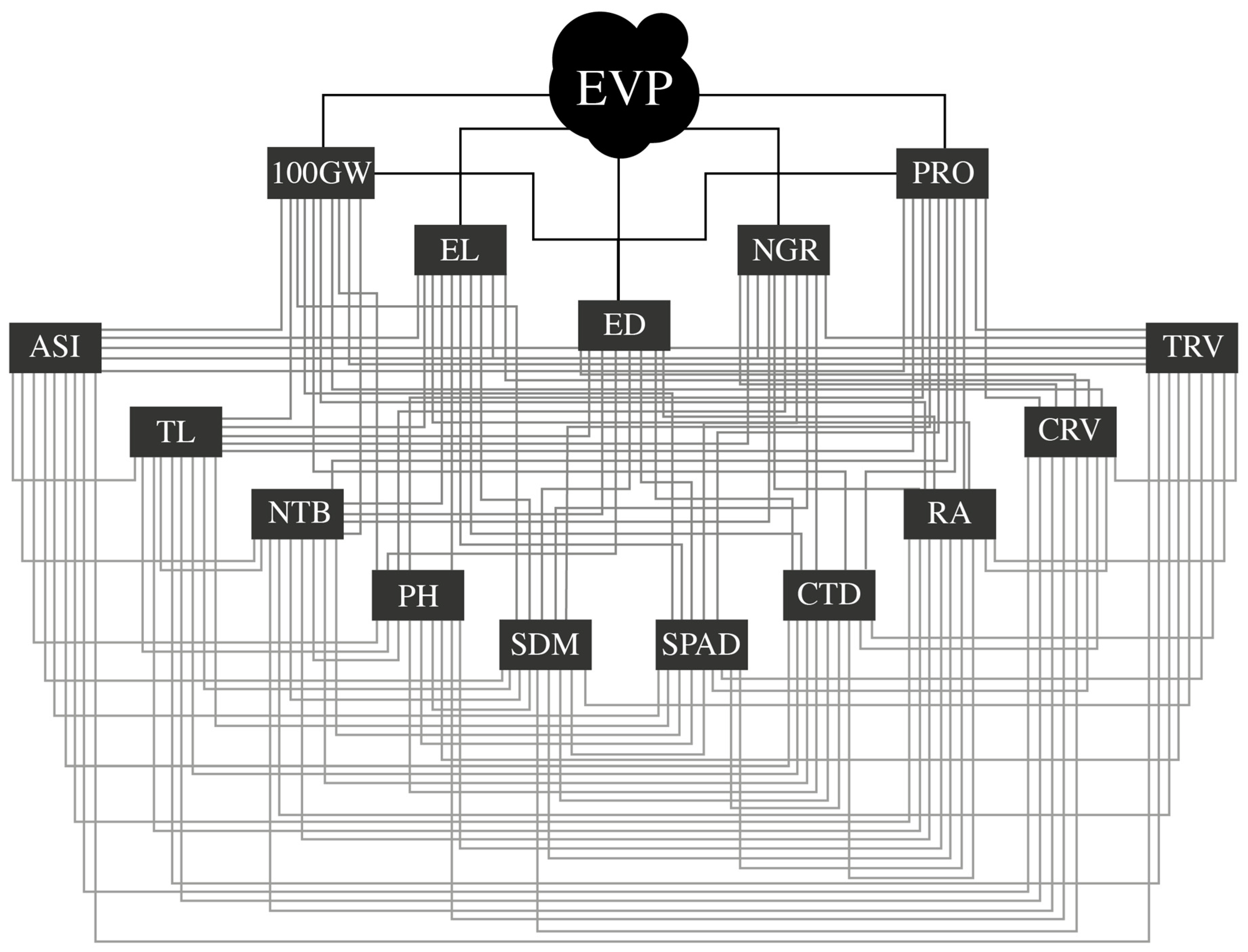
2.4. Path Analysis
3. Results
3.1. Genetic and Environmental Effects of Traits in Full Irrigation (FI) and Drought Stress (DS)
3.2. Direct and Indirect Effects of the First-Order Explanatory Variables on the Variable Expanded Popcorn Volume Per Hectare in FI and DS
3.3. Direct and Indirect Effects of Second-Order Variables on the First-Order Variable Number of Grains per Row (NGR) in FI and DS Environments
3.4. Direct and Indirect Effects of the Second-Order Explanatory Variables on the Main Variable Expanded Popcorn Volume per Hectare (EPV), via NGR, in FI and DS Environments
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Simpson, R.J.; Oberson, A.; Culvenor, R.A.; Ryan, M.H.; Veneklaas, E.J.; Lambers, H.; Lynch, J.P.; Ryan, P.R.; Delhaize, E.; Smith, F.A.; et al. Strategies and agronomic interventions to improve the phosphorus-use efficiency of farming systems. Plant Soil 2011, 349, 89–120. [Google Scholar] [CrossRef]
- Soares, M.O.; Miranda, G.V.; Guimarães, L.J.M.; Marriel, I.E.; Guimarães, C.T. Parâmetros genéticos de uma população de milho em níveis contrastantes de nitrogênio. Rev. Cienc. Agron. 2011, 42, 168–174. [Google Scholar] [CrossRef]
- Adebayo, M.A.; Menkir, A.; Blay, E.; Gracen, V.; Danquah, E.; Hearne, S. Genetic analysis of drought tolerance in adapted × exotic crosses of maize inbred lines under managed stress conditions. Euphytica 2014, 196, 261–270. [Google Scholar] [CrossRef]
- Mendes, F.F.; Guimarães, L.J.M.; Souza, J.C.; Guimarães, P.E.O.; Magalhaes, J.V.; Garcia, A.A.F.; Parentoni, S.N.; Guimarães, C.T. Genetic Architecture of Phosphorus Use Efficiency in Tropical Maize Cultivated in a Low-P Soil. Crop Sci. 2014, 54, 1530–1538. [Google Scholar] [CrossRef]
- Wang, W.; Vinocur, B.; Altman, A. Plant responses to drought, salinity and extreme temperatures: Towards genetic engineering for stress tolerance. Planta 2003, 218, 1–14. [Google Scholar] [CrossRef]
- Durães, F.O.M.; dos Santos, M.X.; Gama, E.; Magalhães, P.C.; Albuquerque, P.E.P.; Guimarães, C.T. Fenotipagem Associada a Tolerância a Seca em Milho para Uso em Melhoramento, Estudos Genômicos e Seleção Assistida por Marcadores. Embrapa Milho e Sorgo-Circular Técnica (INFOTECA-E). 2004, Volume 39, p. 18. Available online: https://www.infoteca.cnptia.embrapa.br/bitstream/doc/487708/1/Circ39.pdf (accessed on 22 November 2019).
- Ghannoum, O. C4 photosynthesis and water stress. Ann. Bot. 2008, 103, 635–644. [Google Scholar] [CrossRef]
- Romano, G.; Zia, S.; Spreer, W.; Sanchez, C.; Cairns, J.; Araus, J.L.; Müller, J. Use of thermography for high throughput phenotyping of tropical maize adaptation in water stress. Comput. Electron. Agric. 2011, 79, 67–74. [Google Scholar] [CrossRef]
- Sanchez, C.; Vargas, M.; Ordóñez, R.A.; Cairns, J.E.; Araus, J.L. Dissecting Maize Productivity: Ideotypes Associated with Grain Yield under Drought Stress and Well-watered Conditions. J. Integr. Plant Biol. 2012, 54, 1007–1020. [Google Scholar]
- Zia, S.; Romano, G.; Spreer, W.; Sanchez, C.; Cairns, J.; Araus, J.L.; Müller, J. Infrared Thermal Imaging as a Rapid Tool for Identifying Water-Stress Tolerant Maize Genotypes of Different Phenology. J. Agron. Crop Sci. 2013, 199, 75–84. [Google Scholar] [CrossRef]
- Das Gracas Dias, K.O.; Gezan, S.A.; Guimaraes, C.T.; Parentoni, S.N.; de Oliveira Guimarães, P.E.; Carneiro, N.P.; Portugal, A.F.; Bastos, E.A.; Cardoso, M.J.; de Oliveira Anoni, C.; et al. Estimating Genotype × Environment Interaction for and Genetic Correlations among Drought Tolerance Traits in Maize via Factor Analytic Multiplicative Mixed Models. Crop Sci. 2018, 58, 72–83. [Google Scholar] [CrossRef]
- Mendes, W.D.S.; Drews, T.A.; Medeiros, J.C.; Rosa, J.D.; Gualberto, A.V.S.; Mielezrski, F. Development and productivity of maize in response to spatial arrangement under semiarid condition of Northeastern Brazil. Aust. J. Crop Sci. 2017, 11, 313–321. [Google Scholar] [CrossRef]
- Vittorazzi, C.; Amaral Júnior, A.T.; Guimarães, A.G.; Silva, F.H.L.; Pena, G.F.; Daher, R.F.; Gerhardt, I.F.S.; Oliveira, G.H.F.; Santos, P.H.A.D.; Souza, Y.P.; et al. Evaluation of genetic variability to form heterotic groups in popcorn. Genet. Mol. Res. 2018, 17. [Google Scholar] [CrossRef]
- Zhuang, Y.; Ren, G.; Yue, G.; Li, Z.; Qu, X.; Hou, G.; Zhu, Y.; Zhang, J. Effects of water-deficit stress on the transcriptomes of developing immature ear and tassel in maize. Plant Cell Rep. 2007, 26, 2137–2147. [Google Scholar] [CrossRef] [PubMed]
- Câmara, T.M.M.; Bento, D.A.V.; Alves, G.F.; Santos, M.F.; Moreira, J.U.V.; de Souza, C.L. Parâmetros genéticos de caracteres relacionados à tolerância à deficiência hídrica em milho tropical. Bragantia 2007, 66, 595–603. [Google Scholar] [CrossRef]
- Teixeira, F.F.; Gomide, R.L.; De Albuquerque, P.E.P.; De Andrade, C.L.T.; Leite, C.E.P.; Parentoni, S.N.; de Oliveira Guimarães, P.E.; Silva, A.R.; Bastos, E.A.; Cardoso, M.J. Evaluation of maize core collection for drought tolerance. Crop Breed. Appl. Biotechnol. 2010, 10, 312–320. [Google Scholar] [CrossRef]
- Costa, E.F.N.; Santos, M.F.; Moro, G.V.; Alves, G.F.; de Souza Júnior, C.L. Herança da senescência retardada em milho. Pesqui. Agropecu. Bras. 2008, 43, 207–213. [Google Scholar] [CrossRef]
- Li, X.H.; Liu, X.D.; Li, M.S.; Zhang, S.H. Identification of quantitative trait loci for anthesis-silking interval and yield components under drought stress in maize. Acta Bot. Sin. 2003, 45, 852–857. [Google Scholar]
- Vaughan, M.M.; Christensen, S.; Schmelz, E.A.; Huffaker, A.; Mcauslane, H.J.; Alborn, H.T.; Romero, M.; Allen, L.H.; Teal, P.E.A. Accumulation of terpenoid phytoalexins in maize roots is associated with drought tolerance. Plant Cell Environ. 2015, 38, 2195–2207. [Google Scholar] [CrossRef]
- Zinselmeier, S.A.; Lauer, M.J.; Boyer, J.S. Reversing Drought-Induced Losses in Grain Yield: Sucrose Maintains Embryo Growth in Maize. Crop Sci. 1995, 35, 1390–1400. [Google Scholar] [CrossRef]
- Cruz, C.D.; Carneiro, P.C.S.; Regazzi, A.J. Modelos Biométricos Aplicados ao Melhoramento Genético—Vol. II; UFV: Vicosa, Brazil, 2014; ISBN 8572691510. [Google Scholar]
- De Carvalho, C.G.P.; Oliveira, V.R.; Cruz, C.D.; Casali, V.W.D. Análise de trilha sob multicolinearidade em pimentão. Pesqui. Agropecu. Bras. 1999, 34, 603–613. [Google Scholar] [CrossRef]
- Wright, S. Correlation and causation. J. Agric. Res. 1921, 20, 557–580. [Google Scholar]
- Do Amaral Júnior, A.T.; Dos Santos, A.; Gerhardt, I.F.S.; Kurosawa, R.N.F.; Moreira, N.F.; Pereira, M.G.; Gravina, G.; Silva, F. Proposal of a super trait for the optimum selection of popcorn progenies based on path analysis. Genet. Mol. Res. 2016, 15. [Google Scholar] [CrossRef] [PubMed]
- Kamphorst, S.H.; de Lima, V.J.; do Amaral Júnior, A.T.; Schmitt, K.F.M.; Leite, J.T.; Carvalho, C.M.; Silva, R.M.R.; Xavier, K.B.; Fereira, F.R.A.; Santos, P.H.A.D.; et al. Popcorn breeding for water-stress tolerance or for agronomic water-use efficiency? Genet. Mol. Res. 2018, 17. [Google Scholar] [CrossRef]
- Jákli, B.; Hauer-Jákli, M.; Böttcher, F.; Meyer zur Müdehorst, J.; Senbayram, M.; Dittert, K. Leaf, canopy and agronomic water-use efficiency of field-grown sugar beet in response to potassium fertilization. J. Agron. Crop Sci. 2018, 204, 99–110. [Google Scholar] [CrossRef]
- Trachsel, S.; Kaeppler, S.M.; Brown, K.M.; Lynch, J.P. Shovelomics: High throughput phenotyping of maize (Zea mays L.) root architecture in the field. Plant Soil 2011, 341, 75–87. [Google Scholar] [CrossRef]
- Christensen, H.K.; Montgomery, C.A. Corporate economic performance: Diversification strategy versus market structure. Strat. Manag. J. 1981, 2, 327–343. [Google Scholar] [CrossRef]
- Cruz, C.D. Genes Software—Extended and integrated with the R, Matlab and Selegen. Acta Sci. Agron. 2016, 38, 547–552. [Google Scholar] [CrossRef]
- Da Silva, P.P.; Soares, L.; Da Costa, J.G.; Viana, L.D.S.; De Andrade, J.C.F.; Gonçalves, E.R.; Dos Santos, J.M.; de Souza Barbosa, G.V.; Nascimento, V.X.; Todaro, A.R.; et al. Path analysis for selection of drought tolerant sugarcane genotypes through physiological components. Ind. Crop Prod. 2012, 37, 11–19. [Google Scholar] [CrossRef]
- Edmeades, G.O.; Bolanos, J.; Chapman, S.C.; Lafitte, H.R.; Banziger, M. Selection Improves Drought Tolerance in Tropical Maize Populations. Crop Sci. 1999, 39, 1306–1315. [Google Scholar] [CrossRef]
- Bolaños, J.; Edmeades, G.O. The importance of the anthesis-silking interval in breeding for drought tolerance in tropical maize. Field Crop Res. 1996, 48, 65–80. [Google Scholar] [CrossRef]
- Maazou, A.R.S.; Tu, J.; Qiu, J.; Liu, Z. Breeding for Drought Tolerance in Maize (Zea mays L.). Am. J. Plant Sci. 2016, 7, 1858–1870. [Google Scholar] [CrossRef]
- Lu, Y.; Hao, Z.; Xie, C.; Crossa, J.; Araus, J.; Gao, S.; Vivek, B.S.; Magorokosho, C.; Mugo, S.; Makumbi, D.; et al. Field Crops Research Large-scale screening for maize drought resistance using multiple selection criteria evaluated under water-stressed and well-watered environments. Field Crop Res. 2011, 124, 37–45. [Google Scholar] [CrossRef]
- Carena, M.J.; Hallauer, A.R.; Filho, J.M. Quantitative Genetics in Maize Breeding; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2010. [Google Scholar]
- De Castro, F.A.; Campostrini, E.; Netto, A.T.; De Menezes De Assis Gomes, M.; Ferraz, T.M.; Glenn, D.M. Portable chlorophyll meter (PCM-502) values are related to total chlorophyll concentration and photosynthetic capacity in papaya (Carica papaya L.). Theor. Exp. Plant Physiol. 2014, 26, 201–210. [Google Scholar] [CrossRef]
- Liu, Y.; Subhash, C.; Yan, J.; Song, C.; Zhao, J.; Li, J. Maize leaf temperature responses to drought: Thermal imaging and quantitative trait loci (QTL) mapping. Environ. Exp. Bot. 2011, 71, 158–165. [Google Scholar] [CrossRef]
- Kaman, H.; Kirda, C.; Sesveren, S. Genotypic differences of maize in grain yield response to deficit irrigation. Agric. Water Manag. 2011, 98, 801–807. [Google Scholar] [CrossRef]
- Li, M.; Liu, S.; Zhang, S.; Hao, Z.; Li, X.; Xie, C.; Weng, J.; Zhang, D.; Liang, X.; Liu, L. Identification of Functional Genetic Variations Underlying Drought Tolerance in Maize Using SNP Markers. J. Integr. Plant Biol. 2011, 53, 641–652. [Google Scholar]
- Chairi, F.; ElAzab, A.; Sanchez-Bragado, R.; Araus, J.L.; Serret, M.D. Heterosis for water status in maize seedlings. Agric. Water Manag. 2016, 164, 100–109. [Google Scholar] [CrossRef]
- Cabral, P.D.S.; do Amaral Júnior, A.T.; de Freitas, I.L.; Ribeiro, R.M.; da Silva, T.R. Cause and effect of quantitative characteristics on grain expansion capacity in popcorn. Rev. Cienc. Agron. 2016, 47, 108–117. [Google Scholar] [CrossRef]
- Pacheco, C.A.P.; dos Santos, M.X.; Cruz, C.D.; Parentoni, S.N.; de Oliveira Guimarães, P.E.; Gama, E.E.G.E.; de Carvalho, H.W.L.; Vieira Júnior, P.A. Inbreeding depression of 28 maize elite open pollinated varieties. Genet. Mol. Biol. 2002, 25, 441–448. [Google Scholar] [CrossRef]
- Araus, J.L.; Kefauver, S.C.; Zaman-Allah, M.; Olsen, M.S.; Cairns, J.E. Translating High-Throughput Phenotyping into Genetic Gain. Trends Plant Sci. 2018, 23, 451–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hawkesford, M.J.; Araus, J.L.; Park, R.; Calderini, D.; Miralles, D.; Shen, T.; Zhang, J.; Parry, M.A.J. Prospects of doubling global wheat yields. Food Energy Secur. 2013, 2, 34–48. [Google Scholar] [CrossRef]
- Chen, D.; Neumann, K.; Friedel, S.; Kilian, B.; Chen, M.; Altmann, T.; Klukas, C. Dissecting the phenotypic components of crop plant growth and drought responses based on high-throughput image analysis. Plant Cell 2014, 26, 4636–4655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elsayed, S.; Elhoweity, M.; Ibrahim, H.H.; Dewir, Y.H.; Migdadi, H.M.; Schmidhalter, U. Thermal imaging and passive reflectance sensing to estimate the water status and grain yield of wheat under different irrigation regimes. Agric. Water Manag. 2017, 189, 98–110. [Google Scholar] [CrossRef]
- Petrov, P.; Petrova, A.; Dimitrov, I.; Tashev, T.; Olsovska, K.; Brestic, M.; Misheva, S. Relationships between leaf morpho-anatomy, water status and cell membrane stability in leaves of wheat seedlings subjected to severe soil drought. J. Agron. Crop Sci. 2018, 204, 219–227. [Google Scholar] [CrossRef]
- Gao, Y.; Lynch, J.P. Reduced crown root number improves water acquisition under water deficit stress in maize (Zea mays L.). J. Exp. Bot. 2016, 67, 4545–4557. [Google Scholar] [CrossRef] [Green Version]
- Brestic, M.; Živčák, M.; Hauptvogel, P.; Misheva, S.; Kocheva, K.; Yang, X.; Li, X.; Allakhverdiev, S.I. Wheat plant selection for high yields entailed improvement of leaf anatomical and biochemical traits including tolerance to non-optimal temperature conditions. Photosynth. Res. 2018, 136, 245–255. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Traits | Water Condition | F Test | GE | Mean ± SD | CVe% | CVg% | H2 |
|---|---|---|---|---|---|---|---|
| EPV | DS | 9.52 * | 1.92 * | 23.82 ± 5.75 | 23.63 | 39.83 | 89.5 |
| FI | 3.59 # | 74.79 ± 19.94 | 26.94 | 25.07 | 72.21 | ||
| 100GW | DS | 10.46 # | 1.52 ns | 9.69 ± 0.73 | 7.50 | 13.32 | 90.44 |
| FI | 3.20 # | 12.67 ± 1.31 | 10.36 | 8.87 | 68.76 | ||
| EL | DS | 5.40 # | 0.80 ns | 12.63 ± 1.07 | 8.47 | 10.26 | 81.49 |
| FI | 6.82 # | 12.72 ± 0.90 | 7.10 | 9.9 | 85.34 | ||
| ED | DS | 7.28 # | 1.34 ns | 27.94 ± 1.66 | 5.94 | 8.6 | 86.28 |
| FI | 7.38 # | 29.12 ± 1.55 | 5.32 | 7.76 | 86.47 | ||
| NGR | DS | 11.16 # | 0.88 ns | 21.44 ± 3.29 | 15.24 | 28.06 | 91.05 |
| FI | 10.10 * | 26.4 ± 3.03 | 11.49 | 20.02 | 90.1 | ||
| PRO | DS | 2.16 # | 4.82 # | 0.86 ± 0.12 | 14.00 | 8.72 | 53.79 |
| FI | 3.75 # | 1.02 ± 0.12 | 12.38 | 11.87 | 73.37 | ||
| ASI | DS | 16.31 # | 0.76 ns | –1.77 ± 0.58 | 32.65 | 73.76 | 93.87 |
| FI | 11.56 # | –1.41 ± 0.61 | 43.57 | 81.77 | 91.35 | ||
| TL | DS | 8.34 # | 1.43 ns | 31.60 ± 2.48 | 7.86 | 12.3 | 88.01 |
| FI | 9.63 # | 32.75 ± 2.37 | 7.24 | 12.28 | 89.63 | ||
| NTB | DS | 9.28 # | 1.42 ns | 14.18 ± 1.82 | 12.85 | 21.36 | 89.24 |
| FI | 11.61 # | 15.25 ± 2.19 | 14.34 | 26.97 | 91.39 | ||
| PH | DS | 18.55 # | 1.15 ns | 160.31 ± 8.06 | 5.03 | 12.17 | 94.61 |
| FI | 24.78 # | 174.14 ± 9.07 | 5.21 | 14.66 | 95.96 | ||
| SDM | DS | 3.76 # | 1.19 ns | 313.95 ± 42.52 | 13.54 | 13.01 | 73.46 |
| FI | 4.04 # | 368.31 ± 50.58 | 13.73 | 13.84 | 75.31 | ||
| SPAD | DS | 4.40 # | 1.49 ns | 32.51 ± 4.75 | 14.6 | 15.65 | 77.52 |
| FI | 6.18 # | 45.99 ± 3.11 | 6.77 | 8.9 | 83.82 | ||
| CTD | DS | 9.00 # | 5.57 # | –0.37 ± 0.33 | 88.36 | 144.32 | 88.89 |
| FI | 3.27 # | –2.88 ± 0.45 | 16.06 | 15.92 | 74.67 | ||
| RA | DS | 9.22 # | 3.84 # | 53.09 ± 3.01 | 5.68 | 24.91 | 89.16 |
| FI | 14.98 # | 42.63 ± 3.33 | 7.81 | 51.74 | 93.33 | ||
| CRV | DS | 22.12 # | 4.81 # | 91.35 ± 8.78 | 9.63 | 25.55 | 95.48 |
| FI | 6.43 # | 63.05 ± 9.94 | 15.76 | 21.21 | 84.45 | ||
| TRV | DS | 18.77 # | 4.20 # | 54.40 ± 5.26 | 9.68 | 23.55 | 94.67 |
| FI | 7.52 # | 46.40 ± 6.96 | 15.00 | 22.11 | 86.7 |
| Variables/Effect | 100GW | EL | ED | NGR | PRO | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| DS | FI | DS | FI | DS | FI | DS | FI | DS | FI | |
| Direct on EPV | 0.57 | 0.87 | −1.16 | 0.02 | −1.00 | 0.00 | 2.65 | 1.06 | −0.32 | −0.70 |
| Indirect via 100GW | - | - | −0.02 | −0.24 | 0.09 | 0.40 | −0.15 | −0.38 | −0.43 | 0.55 |
| Indirect via EL | 0.05 | 0.00 | - | - | 0.05 | 0.00 | −0.88 | 0.01 | −0.56 | 0.00 |
| Indirect via ED | −0.17 | 0.00 | 0.05 | 0.00 | - | - | −0.48 | 0.00 | 0.26 | 0.00 |
| Indirect via NGR | −0.71 | −0.44 | 2.08 | 0.86 | 1.24 | 0.35 | - | - | 1.71 | −0.28 |
| Indirect via PRO | 0.24 | −0.46 | −0.15 | −0.10 | 0.08 | −0.28 | −0.21 | 0.18 | - | - |
| Total effects | −0.01 | −0.05 | 0.71 # | 0.53 * | 0.45 * | 0.47 * | 0.92 # | 0.88 # | 0.65 # | −0.42 |
| DS | FI | |||||||||
| R2 | 0.88 # | 1.21 # | ||||||||
| Residual effect | 0.34 | 0.00 | ||||||||
| Collinearity | 73.37 | 50.36 | ||||||||
| Variables/Effect | NGR | Variables/Effect | NGR | ||||
|---|---|---|---|---|---|---|---|
| DS | FI | DS | FI | ||||
| ASI | Direct | −0.35 | −0.65 | SPAD | Direct | 0.70 | 0.94 |
| Indirect via TL | 0.00 | 0.18 | Indirect via ASI | −0.10 | −0.17 | ||
| Indirect via NTB | 0.36 | 0.19 | Indirect via TL | −0.01 | −0.21 | ||
| Indirect via PH | 0.07 | 0.05 | Indirect via NTB | −0.03 | 0.07 | ||
| Indirect via SDM | 0.00 | −0.07 | Indirect via PH | 0.26 | 0.14 | ||
| Indirect via SPAD | 0.20 | 0.25 | Indirect via SDM | 0.00 | −0.73 | ||
| Indirect via CTD | 0.01 | −0.04 | Indirect via CTD | −0.13 | −0.02 | ||
| Indirect via RA | −0.18 | 0.20 | Indirect via RA | −0.25 | 0.19 | ||
| Indirect via CRV | 0.09 | 0.00 | Indirect via CRV | 0.10 | 0.00 | ||
| Indirect via TRV | −0.20 | −0.01 | Indirect via TRV | −0.11 | −0.03 | ||
| Total effects | 0.00 | 0.10 | Total effects | 0.44 * | 0.18 | ||
| TL | Direct | 0.55 | 1.17 | CTD | Direct | −0.39 | −0.09 |
| Indirect via ASI | 0.00 | −0.10 | Indirect via ASI | 0.01 | −0.30 | ||
| Indirect via NTB | −0.26 | −0.19 | Indirect via TL | −0.12 | 0.31 | ||
| Indirect via PH | 0.37 | 0.16 | Indirect via NTB | 0.05 | −0.01 | ||
| Indirect via SDM | 0.00 | 0.03 | Indirect via PH | 0.20 | 0.25 | ||
| Indirect via SPAD | −0.01 | −0.17 | Indirect via SDM | 0.00 | 0.05 | ||
| Indirect via CTD | 0.09 | −0.02 | Indirect via SPAD | 0.24 | 0.21 | ||
| Indirect via RA | 0.02 | −0.05 | Indirect via RA | −0.24 | 0.17 | ||
| Indirect via CRV | 0.14 | −0.02 | Indirect via CRV | 0.07 | 0.00 | ||
| Indirect via TRV | −0.22 | 0.02 | Indirect via TRV | 0.30 | 0.00 | ||
| Total effects | 0.67 # | 0.79 # | Total effects | 0.12 | 0.44 * | ||
| NTB | Direct | −0.77 | −0.55 | RA | Direct | 0.44 | −0.32 |
| Indirect via ASI | 0.16 | 0.23 | Indirect via ASI | 0.15 | 0.41 | ||
| Indirect via TL | 0.18 | 0.39 | Indirect via TL | 0.03 | 0.19 | ||
| Indirect via PH | 0.31 | 0.17 | Indirect via NTB | −0.11 | −0.09 | ||
| Indirect via SDM | 0.00 | 0.08 | Indirect via PH | −0.37 | −0.12 | ||
| Indirect via SPAD | 0.03 | −0.13 | Indirect via SDM | 0.00 | 0.35 | ||
| Indirect via CTD | 0.02 | −0.02 | Indirect via SPAD | −0.40 | −0.56 | ||
| Indirect via RA | 0.06 | −0.05 | Indirect via CTD | 0.21 | 0.05 | ||
| Indirect via CRV | 0.03 | −0.01 | Indirect via CRV | −0.06 | 0.01 | ||
| Indirect via TRV | 0.08 | 0.00 | Indirect via TRV | 0.19 | 0.01 | ||
| Total effects | 0.14 | 0.10 | Total effects | 0.07 | −0.05 | ||
| PH | Direct | 0.69 | 0.37 | CRV | Direct | −0.56 | 0.05 |
| Indirect via ASI | −0.03 | −0.09 | Indirect via ASI | 0.05 | 0.00 | ||
| Indirect via TL | 0.29 | 0.48 | Indirect via TL | −0.13 | −0.40 | ||
| Indirect via NTB | −0.35 | −0.25 | Indirect via NTB | 0.04 | 0.19 | ||
| Indirect via SDM | 0.00 | −0.31 | Indirect via PH | −0.13 | −0.07 | ||
| Indirect via SPAD | 0.27 | 0.37 | Indirect via SDM | 0.00 | −0.10 | ||
| Indirect via CTD | −0.11 | −0.06 | Indirect via SPAD | −0.12 | −0.12 | ||
| Indirect via RA | −0.24 | 0.10 | Indirect via CTD | 0.04 | 0.00 | ||
| Indirect via CRV | 0.10 | −0.01 | Indirect via RA | 0.04 | −0.05 | ||
| Indirect via TRV | −0.14 | 0.01 | Indirect via TRV | 0.48 | 0.03 | ||
| Total effects | 0.48 * | 0.61 # | Total effects | −0.28 | −0.46 * | ||
| SDM | Direct | 0.00 | −0.78 | TRV | Direct | 0.77 | 0.07 |
| Indirect via ASI | 0.01 | −0.06 | Indirect via ASI | 0.09 | 0.15 | ||
| Indirect via TL | 0.14 | −0.05 | Indirect via TL | −0.16 | 0.38 | ||
| Indirect via NTB | −0.27 | 0.06 | Indirect via NTB | −0.08 | −0.03 | ||
| Indirect via PH | 0.57 | 0.15 | Indirect via PH | −0.13 | 0.06 | ||
| Indirect via SPAD | 0.43 | 0.88 | Indirect via SDM | 0.00 | −0.14 | ||
| Indirect via CTD | −0.27 | 0.00 | Indirect via SPAD | −0.10 | −0.37 | ||
| Indirect via RA | −0.27 | 0.14 | Indirect via CTD | −0.15 | 0.00 | ||
| Indirect via CRV | 0.08 | 0.00 | Indirect via RA | 0.11 | −0.07 | ||
| Indirect via TRV | −0.01 | 0.01 | Indirect via CRV | −0.35 | 0.02 | ||
| Total effects | 0.42 | 0.37 | Total effects | −0.01 | 0.07 | ||
| DS | FI | ||||||
| R2 | 1.03 # | 0.85 # | |||||
| Residual effect | 0.00 | 0.38 | |||||
| Variables | Effect | Residual effect | Total | ||||
|---|---|---|---|---|---|---|---|
| DS | FI | DS | FI | DS | FI | ||
| TL | Direct EPV via NGR | 1.47 | 1.22 | 0.49 | −0.59 | 0.18 | 0.86 |
| Indirect via ASI and via NGR | 0.99 | 0.17 | 0.72 | −0.27 | 0.99 | −0.09 | |
| Indirect via NTB and via NGR | 0.01 | 0.04 | −0.64 | −0.02 | −0.22 | 0.01 | |
| Indirect via PH and via NGR | −0.05 | −0.19 | −0.01 | −0.03 | −0.03 | −0.22 | |
| Indirect via SDM and via NGR | 0.24 | −0.03 | 0.30 | 0.17 | 0.41 | 0.29 | |
| Indirect via SPAD and via NGR | 0.07 | −0.06 | −0.14 | 0.05 | −0.03 | 0.06 | |
| Indirect via CTD and via NGR | 0.37 | 0.03 | 0.32 | 0.19 | 0.40 | 0.05 | |
| Indirect via RA and via NGR | −0.60 | −0.02 | −0.62 | 0.18 | −0.66 | 0.04 | |
| Indirect via CRV and via NGR | −0.69 | −0.20 | −0.30 | 0.00 | −0.47 | −0.32 | |
| Indirect via TRV and via NGR | −0.01 | −0.11 | −0.01 | 0.01 | −0.01 | −0.13 | |
| Total effects | 1.80 # | 0.85 # | 0.12 | −0.33 | 0.57 # | 0.56 # | |
| PH | Direct EPV via NGR | 1.85 | 0.40 | 1.35 | −0.65 | 1.86 | −0.22 |
| Indirect via ASI and via NGR | 0.02 | −0.33 | −1.99 | 0.19 | −0.69 | −0.13 | |
| Indirect via TL and via NGR | 0.72 | 0.40 | 0.15 | 0.07 | 0.38 | 0.46 | |
| Indirect via NTB and via NGR | −0.31 | −0.07 | −0.38 | 0.41 | −0.52 | 0.72 | |
| Indirect via SDM and via NGR | −0.64 | 0.11 | 1.27 | −0.10 | 0.25 | −0.12 | |
| Indirect via SPAD and via NGR | 0.28 | 0.01 | 0.25 | 0.10 | 0.30 | 0.03 | |
| Indirect via CTD and via NGR | −0.39 | −0.01 | −0.40 | 0.09 | −0.43 | 0.02 | |
| Indirect via RA and via NGR | 0.78 | 0.52 | 0.26 | −0.25 | 0.10 | 0.37 | |
| Indirect via CRV and via NGR | −0.93 | −0.28 | −0.41 | 0.00 | −0.63 | −0.43 | |
| Indirect via TRV and via NGR | −0.10 | −0.10 | −0.16 | 0.01 | −0.10 | −0.11 | |
| Total effects | 1.29 # | 0.66 # | −0.06 | −0.13 | 0.51 * | 0.59 # | |
| SPAD | Direct EPV via NGR | 1.88 | 1.01 | 0.40 | 0.18 | 1.00 | 1.18 |
| Indirect via ASI and via NGR | 0.71 | 0.16 | 0.52 | −0.25 | 0.71 | −0.09 | |
| Indirect via TL and via NGR | 0.02 | −0.78 | −1.49 | 0.45 | −0.52 | −0.30 | |
| Indirect via NTB and via NGR | −0.37 | −0.02 | −0.45 | 0.14 | −0.62 | 0.25 | |
| Indirect via PH and via NGR | −0.68 | 0.20 | 1.36 | −0.19 | 0.27 | −0.22 | |
| Indirect via SDM and via NGR | 0.27 | −0.03 | 0.24 | −0.22 | 0.29 | −0.06 | |
| Indirect via CTD and via NGR | −0.30 | −0.01 | −0.30 | 0.06 | −0.33 | 0.02 | |
| Indirect via RA and via NGR | −0.04 | −0.23 | −0.01 | 0.11 | 0.00 | −0.16 | |
| Indirect via CRV and via NGR | −0.10 | 0.08 | −0.04 | 0.00 | −0.07 | 0.13 | |
| Indirect via TRV and via NGR | −0.27 | −0.18 | −0.45 | 0.01 | −0.29 | −0.22 | |
| Total effects | 1.13 # | 0.20 | −0.24 | 0.29 | 0.45 * | 0.53 * | |
| CTD | Direct EPV via NGR | −1.04 | −0.11 | −1.29 | 0.61 | −1.76 | 1.06 |
| Indirect via ASI and via NGR | 0.55 | 0.27 | 0.40 | −0.44 | 0.55 | −0.15 | |
| Indirect via TL and via NGR | 0.02 | 0.06 | −1.68 | −0.04 | −0.59 | 0.02 | |
| Indirect via NTB and via NGR | 0.66 | 0.23 | 0.14 | 0.04 | 0.35 | 0.27 | |
| Indirect via PH and via NGR | −0.66 | 0.19 | 1.31 | −0.18 | 0.26 | −0.21 | |
| Indirect via SDM and via NGR | 0.19 | 0.01 | 0.16 | 0.04 | 0.20 | 0.01 | |
| Indirect via SPAD and via NGR | 0.81 | −0.01 | 0.83 | 0.05 | 0.89 | 0.01 | |
| Indirect via RA and via NGR | −0.34 | 0.33 | −0.11 | −0.16 | −0.04 | 0.23 | |
| Indirect via CRV and via NGR | 0.13 | −0.18 | 0.06 | 0.00 | 0.09 | −0.27 | |
| Indirect via TRV and via NGR | 0.03 | −0.33 | 0.05 | 0.02 | 0.03 | −0.39 | |
| Total effects | 0.34 | 0.48 * | −0.14 | −0.05 | −0.02 | 0.60 # | |
| CRV | Direct EPV via NGR | 2.06 | 0.06 | 2.11 | −0.50 | 2.25 | −0.12 |
| Indirect via ASI and via NGR | −0.35 | −0.08 | −0.26 | 0.12 | −0.35 | 0.04 | |
| Indirect via TL and via NGR | 0.00 | −0.11 | 0.04 | 0.06 | 0.01 | −0.04 | |
| Indirect via NTB and via NGR | −0.27 | −0.13 | −0.06 | −0.02 | −0.14 | −0.15 | |
| Indirect via PH and via NGR | −0.41 | 0.01 | −0.51 | −0.06 | −0.69 | −0.11 | |
| Indirect via SDM and via NGR | 0.29 | −0.06 | −0.58 | 0.06 | −0.12 | 0.07 | |
| Indirect via SPAD and via NGR | −0.94 | 0.04 | −0.82 | 0.24 | −1.01 | 0.06 | |
| Indirect via CTD and via NGR | −0.43 | −0.43 | −0.14 | 0.21 | −0.05 | −0.30 | |
| Indirect via RA and via NGR | −0.22 | 0.20 | −0.10 | 0.00 | −0.15 | 0.31 | |
| Indirect via TRV and via NGR | 0.25 | 0.00 | 0.41 | 0.00 | 0.26 | 0.01 | |
| Total effects | −0.03 | −0.49 * | 0.09 | 0.11 | 0.00 | −0.24 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kamphorst, S.H.; Amaral Júnior, A.T.d.; Lima, V.J.d.; Guimarães, L.J.M.; Schmitt, K.F.M.; Leite, J.T.; Santos, P.H.A.D.; Chaves, M.M.; Mafra, G.S.; Santos Junior, D.R.d.; et al. Can Genetic Progress for Drought Tolerance in Popcorn Be Achieved by Indirect Selection? Agronomy 2019, 9, 792. https://doi.org/10.3390/agronomy9120792
Kamphorst SH, Amaral Júnior ATd, Lima VJd, Guimarães LJM, Schmitt KFM, Leite JT, Santos PHAD, Chaves MM, Mafra GS, Santos Junior DRd, et al. Can Genetic Progress for Drought Tolerance in Popcorn Be Achieved by Indirect Selection? Agronomy. 2019; 9(12):792. https://doi.org/10.3390/agronomy9120792
Chicago/Turabian StyleKamphorst, Samuel Henrique, Antônio Teixeira do Amaral Júnior, Valter Jário de Lima, Lauro José Moreira Guimarães, Kátia Fabiane Medeiros Schmitt, Jhean Torres Leite, Pedro Henrique Araújo Diniz Santos, Marcelo Moura Chaves, Gabrielle Sousa Mafra, Divino Rosa dos Santos Junior, and et al. 2019. "Can Genetic Progress for Drought Tolerance in Popcorn Be Achieved by Indirect Selection?" Agronomy 9, no. 12: 792. https://doi.org/10.3390/agronomy9120792