Agroecological Practices and Agrobiodiversity: A Case Study on Organic Orange in Southern Italy

 , ,
, ,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
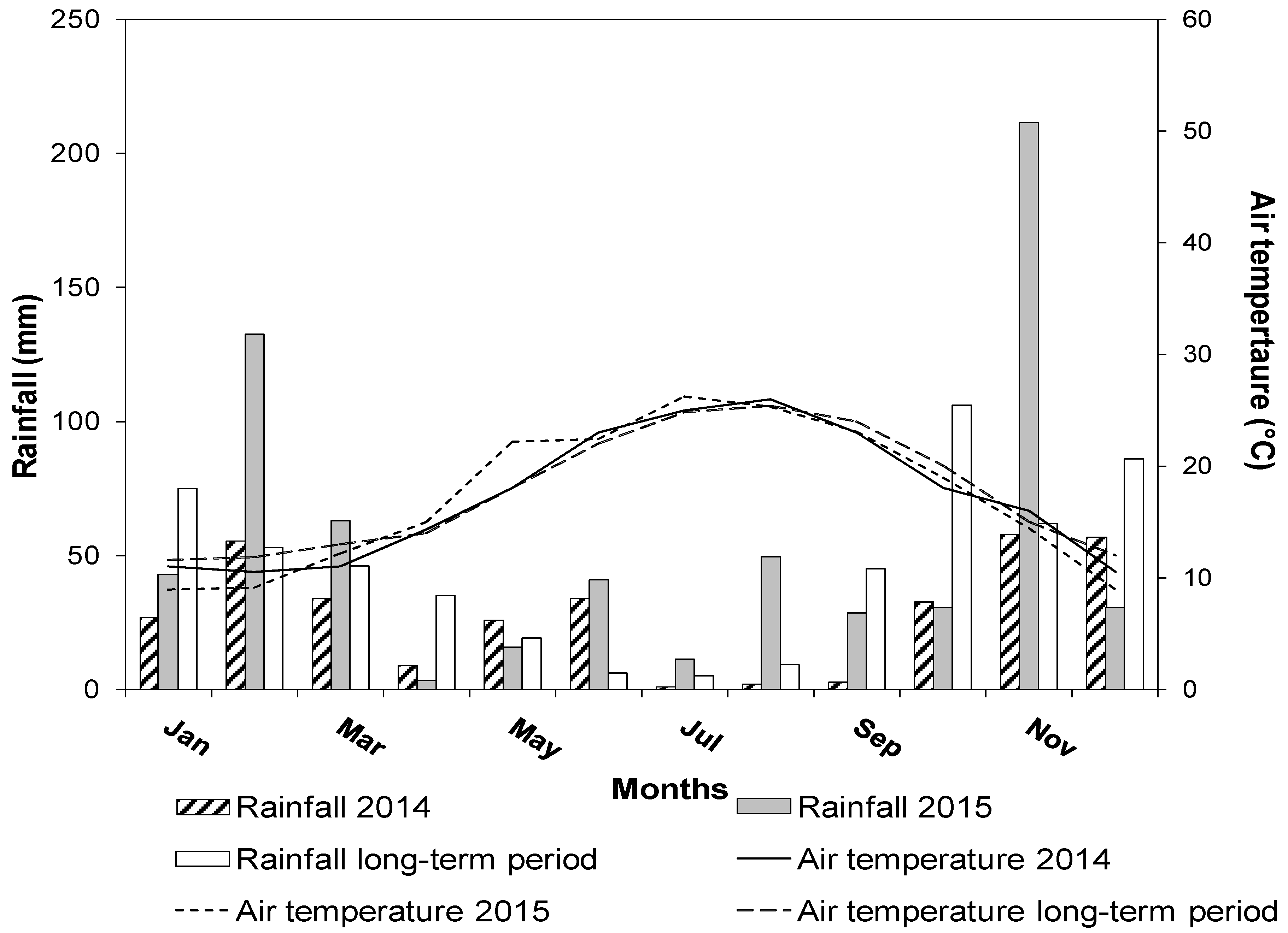
2.1. Site Description and Climatic Data
2.2. System Management and Sampling
2.3. Soil Fertility Attributes
2.4. Fungal and Oomycetal Diversity Assessment
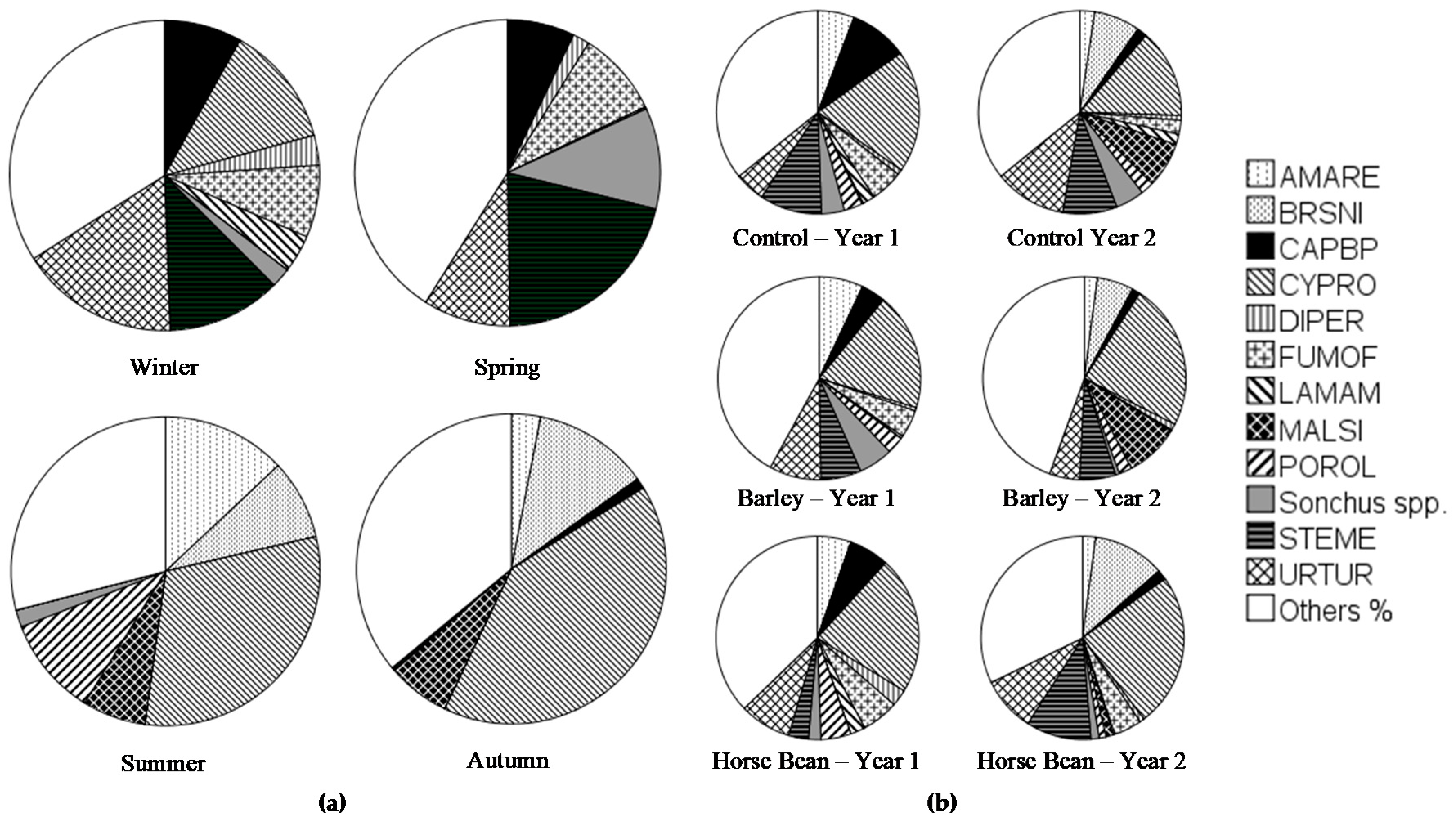
2.5. Weed Diversity Assessment
2.6. Statistical Analysis
3. Results and Discussion
3.1. Principal Component Analysis
3.2. Differences among Compared Systems
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Alexandratos, N.; Bruinsma, J. World Agriculture towards 2030/2050: The 2012 Revision; ESA Working Paper No. 12-03; FAO: Rome, Italy, 2012. [Google Scholar]
- IFOAM. Principles of Organic Agriculture; IFOAM Head Officer: Bonn, Germany, 2005. [Google Scholar]
- Rahmann, G.; Reza Ardakani, M.; Bàrberi, P.; Boehm, H.; Canali, S.; Chander, M.; David, W.; Dengel, L.; Erisman, J.W.; Galvis-Martinez, A.C.; et al. Organic Agriculture 3.0 is innovation with research. Org. Agric. 2017, 7, 169–197. [Google Scholar] [CrossRef]
- Darnhofer, I.; Lindenthal, T.; Bartel-Kratochvil, R.; Zollitsch, W. Conventionalisation of organic farming practices: From structural criteria towards an assessment based on organic principles. Agron. Sustain. Dev. 2010, 30, 67–81. [Google Scholar] [CrossRef]
- Niggli, U. Incorporating Agroecology into Organic Research-An Ongoing Challenge. Sustain. Agric. Res. 2015, 4, 149–157. [Google Scholar] [CrossRef]
- Francis, C.; Lieblein, G.; Gliessman, S.; Breland, T.A.; Creamer, N.; Harwood, R.; Salomonsson, L.; Helenius, J.; Rickerl, D.; Salvador, R.; et al. Agroecology: The ecology of food systems. J. Sustain. Agric. 2003, 22, 99–118. [Google Scholar] [CrossRef]
- Gliessman, S. Transforming food systems with agroecology. Agroecol. Sustain. Food Syst. 2016, 40, 187–189. [Google Scholar] [CrossRef] [Green Version]
- Magdoff, F. Ecological agriculture: Principles, practices, and constraints. Renew. Agric. Food Syst. 2007, 22, 109–117. [Google Scholar] [CrossRef]
- Wezel, A.; Casagrande, M.; Celette, F.; Vian, J.F.; Ferrer, A.; Peigné, J. Agroecological practices for sustainable agriculture. A review. Agron. Sustain. Dev. 2014, 34, 1–20. [Google Scholar] [CrossRef]
- Wood, S.A.; Karp, D.S.; DeClerck, F.; Kremen, C.; Naeem, S.; Palm, C.A. Functional traits in agriculture: Agrobiodiversity and ecosystem services. Trends Ecol. Evol. 2015, 30, 531–539. [Google Scholar] [CrossRef]
- Love, B.; Spaner, D. Agrobiodiversity: Its Value, Measurement, and Conservation in the Context of Sustainable Agriculture. J. Sustain. Agric. 2007, 31, 53–82. [Google Scholar] [CrossRef]
- Tilman, D. The ecological consequences of changes in biodiversity: A search for general principles. Ecology 1999, 80, 1455–1474. [Google Scholar] [CrossRef]
- Bommarco, R.; Kleijn, D.; Potts, S.G. Ecological intensification: Harnessing ecosystem services for food security. Trends Ecol. Evol. 2013, 28, 230–238. [Google Scholar] [CrossRef] [PubMed]
- Canali, S.; Diacono, M.; Campanelli, G.; Montemurro, F. Organic No-Till with Roller Crimpers: Agro-ecosystem Services and Applications in Organic Mediterranean Vegetable Productions. Sustain. Agric. Res. 2015, 4, 70–79. [Google Scholar] [CrossRef]
- Depalo, L.; Burgio, G.; von Fragstein, P.; Kristensen, H.L.; Bavec, M.; Robačer, M.; Campanelli, G.; Canali, S. Impact of living mulch on arthropod fauna: Analysis of pest and beneficial dynamics on organic cauliflower (Brassica oleracea L. var. botrytis) in different European scenarios. Renew. Agric. Food Syst. 2016, 32, 240–247. [Google Scholar] [CrossRef]
- Navarro-Miró, D.; Cabellero-López, B.; Blanco-Moreno, J.M.; Pérez-Ferrer, A.; Depalo, L.; Masetti, A.; Burgio, G.; Canali, S.; Sans, F.X. Agro-ecological Service Crops with roller crimper termination enhance ground-dwelling predator communities and pest regulation. In Proceedings of the Innovative Research for Organic 3.0 Science track at the Organic Word Congress, Delhi, India, 9–11 November 2017; Volume 1, pp. 416–419. [Google Scholar]
- Ciaccia, C.; Kristensen, H.L.; Campanelli, G.; Xie, Y.; Testani, E.; Leteo, F.; Canali, S. Living mulch for weed management in organic vegetable cropping systems under Mediterranean and North European conditions. Renew. Agric. Food Syst. 2016, 32, 248–262. [Google Scholar] [CrossRef]
- Ciaccia, C.; Canali, S.; Campanelli, G.; Testani, E.; Montemurro, F.; Leteo, F.; Delate, K. Effect of roller-crimper technology on weed management in organic zucchini production in a Mediterranean climate zone. Renew. Agric. Food Syst. 2016, 31, 111–121. [Google Scholar] [CrossRef]
- Trinchera, A.; Testani, E.; Ciaccia, C.; Campanelli, G.; Leteo, F.; Canali, S. Effects induced by living mulch on rhizosphere interactions in organic artichoke: The cultivar’s adaptive strategy. Renew. Agric. Food Syst. 2016, 32, 214–223. [Google Scholar] [CrossRef]
- Yue, X.; Kristensen, H.L. Overwintering grass-clover as intercrop and moderately reduced nitrogen fertilization maintain yield and reduce the risk of nitrate leaching in an organic cauliflower (Brassica oleracea L. var. botrytis) agroecosystem. Sci. Hort. 2016, 206, 71–79. [Google Scholar]
- Callaway, R.M.; Mahall, B.E.; Wicks, C.; Pankey, J.; Zabinski, C. Soil fungi and the effects of an invasive forb on grasses: Neighbor identity matters. Ecology 2003, 84, 129–135. [Google Scholar] [CrossRef]
- IUSS Working Group. WRB World Reference Base for Soil Resources 2014, Update 2015 International Soil Classification System for Naming Soils and Creating Legends for Soil Maps; World Soil Resources Reports No. 106; FAO: Rome, Italy, 2015. [Google Scholar]
- Peel, M.C.; Finlayson, B.L.; McMahon, T.A. Updated world map of the Köppen-Geiger climate classification. Hydrol. Earth Syst. Sci. 2007, 11, 1633–1644. [Google Scholar] [CrossRef]
- Canali, S.; Trinchera, A.; Intrigliolo, F.; Pompili, L.; Nisini, L.; Mocali, S.; Torrisi, B. Effect of long term addition of composts and poultry manure on soil quality of citrus orchards in Southern Italy. Biol. Fertil. Soils 2004, 40, 206–210. [Google Scholar] [CrossRef]
- Krom, M.D. Spectrophotometric determination of ammonia: A study of a modified Berthelot reaction using salicylate and dichloroisocyanurate. Analyst 1980, 105, 305–316. [Google Scholar] [CrossRef]
- Henriksen, A.; Selmer-Olsen, A.R. Automatic methods for determining nitrate and nitrite in water and soil extracts. Analyst 1970, 95, 514–518. [Google Scholar] [CrossRef]
- Olsen, S.R.; Sommers, L.E. Phosphorus. In Methods of Soil Analysis; Page, A.L., Ed.; American Society of Agronomy, Soil Science Society of America: Madison, WI, USA, 1982; Part 2; pp. 403–430. [Google Scholar]
- LECO Corporation. Instruction Manual—CHN—600 Elemental Analyzer for Macro-Samples System; LECO Corporation: St. Joseph, MI, USA, 1987; p. 49085. [Google Scholar]
- Waksman, S.A. A method for counting the number of fungi in the soil. J. Bacteriol. 1922, 7, 339–341. [Google Scholar] [PubMed]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communicatio; University of Illinois Press: Urbana, IL, USA, 1948. [Google Scholar]
- Pignatti, S. Geobotanica. In Trattato di Botanica; Cappelletti, C., Ed.; UTET: Turin, Italy, 1976; pp. 801–997. [Google Scholar]
- Wikum, D.A.; Shanholtzer, G. Application of the Braun-Blanquet cover-abundance scale for vegetation analysis in land development studies. Environ. Manag. 1978, 2, 323–329. [Google Scholar] [CrossRef]
- Magurran, A.E. Ecological Diversity and Its Measurement; Springer Science & Business Media: Dordrecht, The Netherlands, 2013. [Google Scholar]
- Gotelli, N.J.; Ellison, A.M. A Primer of Ecological Statistics, 2nd ed.; Sinauer Associates Inc.: Sunderland, MA, USA, 2004. [Google Scholar]
- Pili, S.; Grigoriadis, E.; Carlucci, M.; Clemente, M.; Salvati, L. Towards Sustainable Growth? A Multi-criteria Assessment of (Changing) Urban Forms. Ecol. Indic. 2017, 76, 71–80. [Google Scholar] [CrossRef]
- Duvernoy, I.; Zambon, I.; Sateriano, A.; Salvati, L. Pictures from the Other Side of the Fringe: Urban Growth and Peri-urban Agriculture in a Post-industrial City (Toulouse, France). J. Rural Stud. 2018, 57, 25–35. [Google Scholar] [CrossRef]
- Bangarwa, S.K.; Norsworthy, J.K.; Jha, P.; Malik, M. Purple Nutsedge (Cyperus rotundus) Management in an Organic Production System. Weed Sci. 2008, 56, 606–613. [Google Scholar] [CrossRef]
- Joshi, I.J.; Chauhan, R.K.S. Soil fungal ecology of cultivated areas of Chambal ravines. Proc. Indian Natl. Sci. Acad. 1981, 47, 248–254. [Google Scholar]
- Hart, S.C.; Stark, J.M. Nitrogen limitation of the microbial biomass in an old-growth forest soil. EcoScience 1997, 4, 91–98. [Google Scholar] [CrossRef]
- Zhang, Q.H.; Zak, J.C. Effects of water and nitrogen amendment on soil microbial biomass and fine root production in a semi-arid environment in west Texas. Soil Biol. Biochem. 1998, 30, 39–45. [Google Scholar] [CrossRef]
- Ladygina, N.; Hedlund, K. Plant species influence microbial diversity and carbon allocation in the rhizosphere. Soil Biol. Biochem. 2010, 42, 162–168. [Google Scholar] [CrossRef]
- Rottstock, T.; Joshi, J.; Kummer, V.; Fischer, M. Higher plant diversity promotes higher diversity of fungal pathogens, while it decreases pathogen infection per plant. Ecology 2014, 95, 1907–1917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LeBlanc, N.; Kinkel, L.; Kistler, H.C. Soil fungal communities respond to grassland plant community richness and soil edaphics. Microb. Ecol. 2015, 70, 188–195. [Google Scholar] [CrossRef] [PubMed]
- Frąc, M.; Hannula, S.E.; Bełka, M.; Jędryczka, M. Fungal biodiversity and their role in soil health. Front. Microbiol. 2018, 9, 707. [Google Scholar] [CrossRef] [PubMed]
- Gonnella, M.; Charfeddine, M.; Conversa, G.; Santamaria, P. Purslane: A review of its potential for health and agricultural aspects. Eur. J. Plant Sci. Biotechnol. 2010, 4, 131–136. [Google Scholar]
- Abbasi, M.K.; Tahir, M.M.; Sabir, N.; Khurshid, M. Impact of the addition of different plant residues on nitrogen mineralization–immobilization turnover and carbon content of a soil incubated under laboratory conditions. Solid Earth 2015, 6, 197–205. [Google Scholar] [CrossRef] [Green Version]
- Pignatti, S. Valori di bioindicazione delle piante vascolari della flora d’Italia. BraunBlanquetia 2005, 39, 2–97. [Google Scholar]
- Brullo, S.; Giusso Del Galdo, G.; Guarino, R.; Minissale, P. A survey of the weedy communities of Sicily. Annali di Botanica 2007, 7, 127–161. [Google Scholar] [CrossRef]
- Halde, C.; Entz, M.H. Plant species and mulch application rate affected decomposition of cover crop mulches used in organic rotational no-till systems. Can. J. Plant Sci 2016, 96, 59–71. [Google Scholar] [CrossRef] [Green Version]
- Hill, E.C.; Renner, K.A.; Sprague, C.L.; Davis, A.S. Cover crop impact on weed dynamics in an organic dry bean system. Weed Sci. 2016, 64, 261–275. [Google Scholar] [CrossRef] [Green Version]
- Calderon, F.J.; Nielsen, D.; Acosta-Martinez, V.; Vigil, M.F.; Drew, L.Y. Cover crop and irrigation effects on soil microbial communities and enzymes in semiarid agroecosystems of the central Great Plains of North America. Pedosphere 2016, 26, 192–205. [Google Scholar] [CrossRef]
- Finney, D.M.; Buyer, J.S.; Kaye, J.P. Living cover crops have immediate impacts on soil microbial community structure and function. J. Soil Water Constr. 2017, 72, 361–373. [Google Scholar] [CrossRef] [Green Version]
- Diacono, M.; Montemurro, F. Long-term effects of organic amendments on soil fertility. Sustain. Agric. 2011, 2, 761–786. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Parameter | Unit | Assessment |
|---|---|---|
| Total Nitrogen (TN) | % | Soil fertility |
| Total Organic Carbon (TOC) | % | |
| Soil available N (Nitrate, N-NO3−) | kg ha−1 | |
| Soil available N (Ammonium, N-NH4+) | kg ha−1 | |
| Available Phosphorus (Olsen, avP) | mg kg−1 | |
| Amaranthus retroflexus L. (AMARE) | % | Weed community |
| Brassica nigra (L.) Koch (BRSNI) | % | |
| Capsella borsa-pastoris (L.) Medik. (CAPBP) | % | |
| Calendula arvensis L. (CLDAR) | % | |
| Convolvulus arvensis L. (CONAR) | % | |
| Cyperus rotondus L. (CYPRO) | % | |
| Diplotaxis erucoides (L.) DC. (DIPER) | % | |
| Conyza canadensis L. (ERICA) | % | |
| Fumaria officinalis L. (FUMOF) | % | |
| Lamium amplexicaule L. (LAMAM) | % | |
| Malva sylvestris L. (MALSI) | % | |
| Oxalis pes-caprae L. (OXAPC) | % | |
| Portulaca oleracea L. (POROL) | % | |
| S. arvensis, S. asper, S. oleraceus L. (Sonchus spp.) | % | |
| Stellaria media (L.) Vill. (STEME) | % | |
| Urtica urens L. (URTUR) | % | |
| Broad leaves species (27 species) | % | |
| Grass species (8 species) | % | |
| Weed density | - | |
| Weed Evenness (Weed e) | - | |
| Weed Richness (Weed R) | n m−2 | |
| Alternaria spp. | % | Fungi and oomycetes community |
| Aspergillus spp. | % | |
| Cladosporium spp. | % | |
| Fusarium spp. | % | |
| Graphyum sp. | % | |
| Penicillium spp. | % | |
| Phytophthora sp. | % | |
| Rhizopus nigricans | % | |
| Staphylotrichum sp. | % | |
| Trichoderma spp. | % | |
| Verticillium spp. | % | |
| Others (20 genera) | % | |
| Colony Forming Unit (Ln-CFU) | Ln (n g−1) | |
| Fungi and oomycetes Diversity (H-fungi) | - | |
| Fungi and oomycetes Richness (S-fungi) | - |
| PC1 | PC2 | PC3 | PC4 | PC5 | PC6 | |
|---|---|---|---|---|---|---|
| TN | −0.33 | 0.42 | 0.35 | 0.13 | −0.43 | −0.14 |
| TOC | −0.42 | 0.38 | 0.34 | 0.15 | −0.53 | −0.01 |
| N-NO3− | 0.09 | −0.35 | 0.08 | −0.07 | 0.50 | 0.12 |
| AMARE | −0.37 | −0.36 | 0.38 | 0.34 | 0.40 | −0.17 |
| BRSNI | −0.29 | −0.18 | 0.02 | 0.02 | −0.47 | 0.07 |
| CAPBP | 0.57 | −0.18 | 0.04 | −0.04 | −0.10 | 0.04 |
| CYPRO | −0.51 | −0.31 | −0.14 | −0.11 | 0.05 | −0.09 |
| DIPER | 0.42 | 0.05 | −0.08 | 0.32 | −0.18 | −0.11 |
| FUMOF | 0.70 | −0.09 | 0.05 | −0.04 | −0.09 | 0.04 |
| LAMAM | 0.53 | −0.03 | −0.19 | 0.40 | −0.17 | −0.16 |
| MALSI | −0.23 | 0.01 | −0.06 | −0.09 | −0.42 | 0.11 |
| POROL | −0.28 | −0.38 | 0.43 | 0.37 | 0.31 | −0.14 |
| Sonchus spp. | 0.37 | −0.03 | 0.49 | −0.23 | −0.01 | 0.33 |
| STEME | 0.61 | 0.13 | 0.11 | −0.11 | −0.10 | 0.06 |
| URTUR | 0.42 | 0.19 | −0.10 | 0.04 | −0.14 | −0.18 |
| Weed density | −0.02 | −0.53 | −0.10 | 0.04 | −0.04 | −0.08 |
| Weed evenness | 0.37 | −0.40 | 0.30 | 0.07 | 0.03 | 0.03 |
| Weed R | 0.36 | 0.40 | 0.14 | 0.12 | 0.27 | 0.11 |
| Fusarium spp. | 0.39 | −0.10 | 0.19 | 0.17 | −0.03 | −0.55 |
| Cladosporium spp. | −0.07 | 0.07 | −0.20 | −0.02 | 0.07 | 0.51 |
| Rhizopus nigricans | 0.23 | 0.05 | 0.11 | 0.23 | −0.04 | 0.45 |
| Ln (CFU) | 0.07 | 0.60 | 0.18 | 0.23 | 0.35 | −0.34 |
| H-fungi | −0.11 | −0.01 | −0.48 | 0.61 | 0.04 | 0.23 |
| S-fungi | −0.05 | −0.07 | −0.43 | 0.62 | −0.01 | 0.14 |
| PC1 | PC2 | PC3 | PC4 | PC5 | PC6 | |
|---|---|---|---|---|---|---|
| Year (Y) | ||||||
| Y 1 | 0.90 a 2 | 0.32 b | 0.10 | 0.10 | 0.05 | −0.00 |
| Y 2 | −0.63 b | −0.22 a | −0.07 | −0.07 | −0.03 | 0.00 |
| sig. 1 | *** | *** | n.s. | n.s. | n.s. | n.s. |
| System (S) | ||||||
| Cont | 1.03 a | −1.11 c | 0.23 a | 0.24 | 0.43 a | 0.04 ab |
| Barley (B) | 0.07 b | 1.14 a | −0.57 b | −0.14 | 0.53 a | −0.32 b |
| Horse Bean (HB) | −1.09 c | −0.05 b | 0.36 a | −0.10 | −0.91 b | 0.28 a |
| sig. | *** | *** | *** | n.s. | *** | ** |
| Fertilizer (F) | ||||||
| Min | 11.3 | −0.14 b | 0.21 a | −0.06 | 0.05 | 0.14 |
| Comp | −11.3 | 0.14 a | −0.21 a | 0.06 | −0.05 | −0.14 |
| sig. | n.s. | *** | *** | n.s. | n.s. | n.s. |
| Y × S | *** | ** | *** | n.s. | *** | n.s. |
| Y × F | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. |
| S × F | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. |
| Y × S × F | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. |
| PC1 | PC2 | PC3 | PC5 | |
|---|---|---|---|---|
| Y 1 Cont | 3.22 a2 | −0.77 b | −0.38 c | −0.31 c |
| Y 2 Cont | −0.52 c | −1.36 c | 0.65 b | 1.17 a |
| Y 1 B | 0.54 b | 1.07 a | −0.61 c | 0.82 a |
| Y 2 B | −0.62 c | 1.19 a | −0.55 c | 0.24 b |
| Y 1 HB | 0.89 b | 0.65 a | 1.27 a | −0.37 c |
| Y 2 HB | −1.92 d | −0.53 b | −0.29 c | −1.45 d |
| Sig.1 | *** | ** | *** | *** |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ciaccia, C.; La Torre, A.; Ferlito, F.; Testani, E.; Battaglia, V.; Salvati, L.; Roccuzzo, G. Agroecological Practices and Agrobiodiversity: A Case Study on Organic Orange in Southern Italy. Agronomy 2019, 9, 85. https://doi.org/10.3390/agronomy9020085
Ciaccia C, La Torre A, Ferlito F, Testani E, Battaglia V, Salvati L, Roccuzzo G. Agroecological Practices and Agrobiodiversity: A Case Study on Organic Orange in Southern Italy. Agronomy. 2019; 9(2):85. https://doi.org/10.3390/agronomy9020085
Chicago/Turabian StyleCiaccia, Corrado, Anna La Torre, Filippo Ferlito, Elena Testani, Valerio Battaglia, Luca Salvati, and Giancarlo Roccuzzo. 2019. "Agroecological Practices and Agrobiodiversity: A Case Study on Organic Orange in Southern Italy" Agronomy 9, no. 2: 85. https://doi.org/10.3390/agronomy9020085
APA StyleCiaccia, C., La Torre, A., Ferlito, F., Testani, E., Battaglia, V., Salvati, L., & Roccuzzo, G. (2019). Agroecological Practices and Agrobiodiversity: A Case Study on Organic Orange in Southern Italy. Agronomy, 9(2), 85. https://doi.org/10.3390/agronomy9020085