Influence of Hydroxyapatite Nanoparticles on Germination and Plant Metabolism of Tomato (Solanum lycopersicum L.): Preliminary Evidence

 ,
,  ,
,  ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Hydroxyapatite Nanoparticles Preparation
2.2. Hydroxyapatite Chemical, Morphological, and Structural Characterization
2.3. Plant Experiment
2.3.1. Treatments
2.3.2. Seed Germination and Root Elongation
2.3.3. Seedling Hydroponic Culture
2.3.4. ICP-MS Analysis
2.3.5. Reactive Oxygen Species (ROS) Determination
2.3.6. Photosynthetic Pigments Quantification
2.3.7. Determination of Cellular ATP
2.3.8. Data Analysis
3. Results
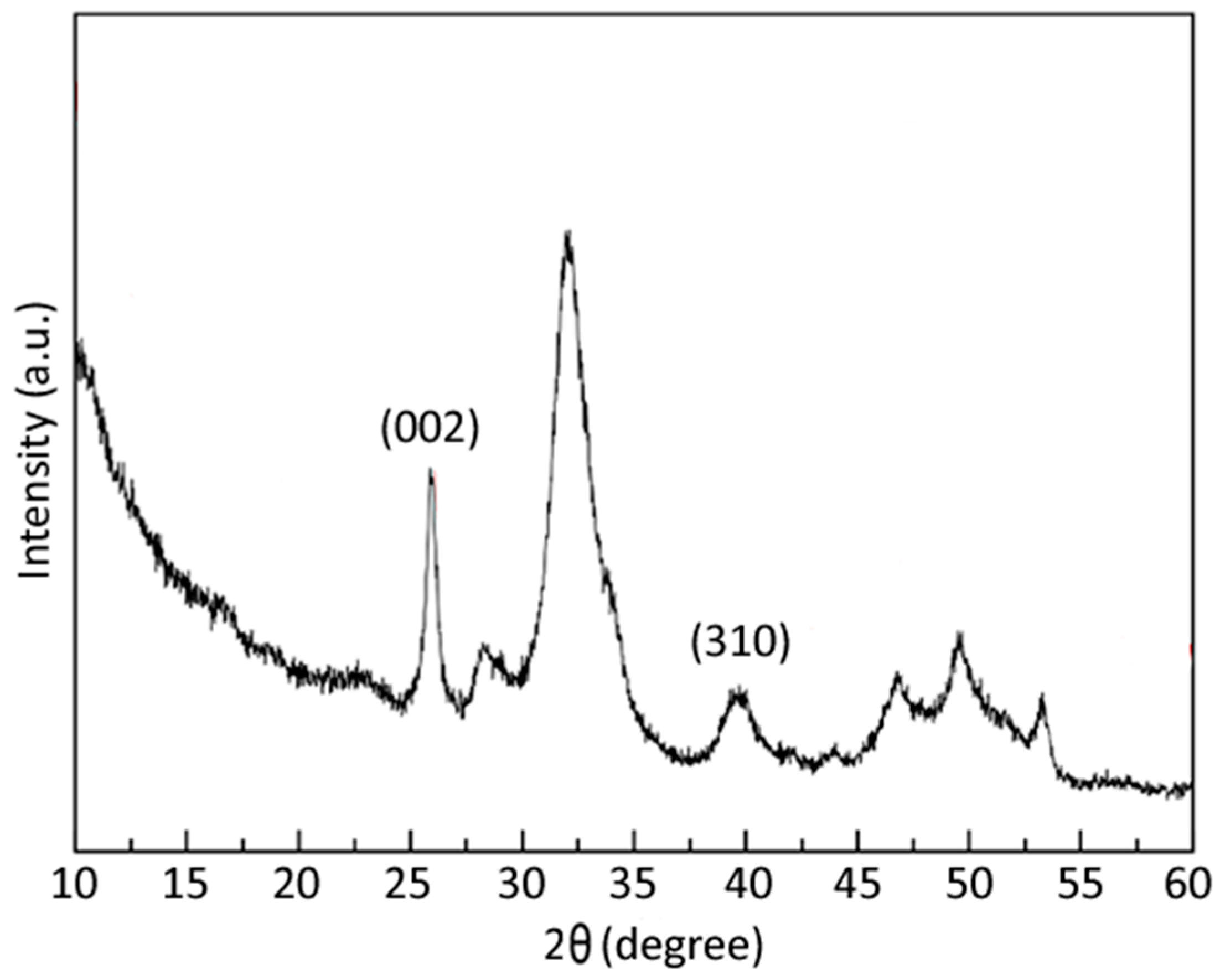
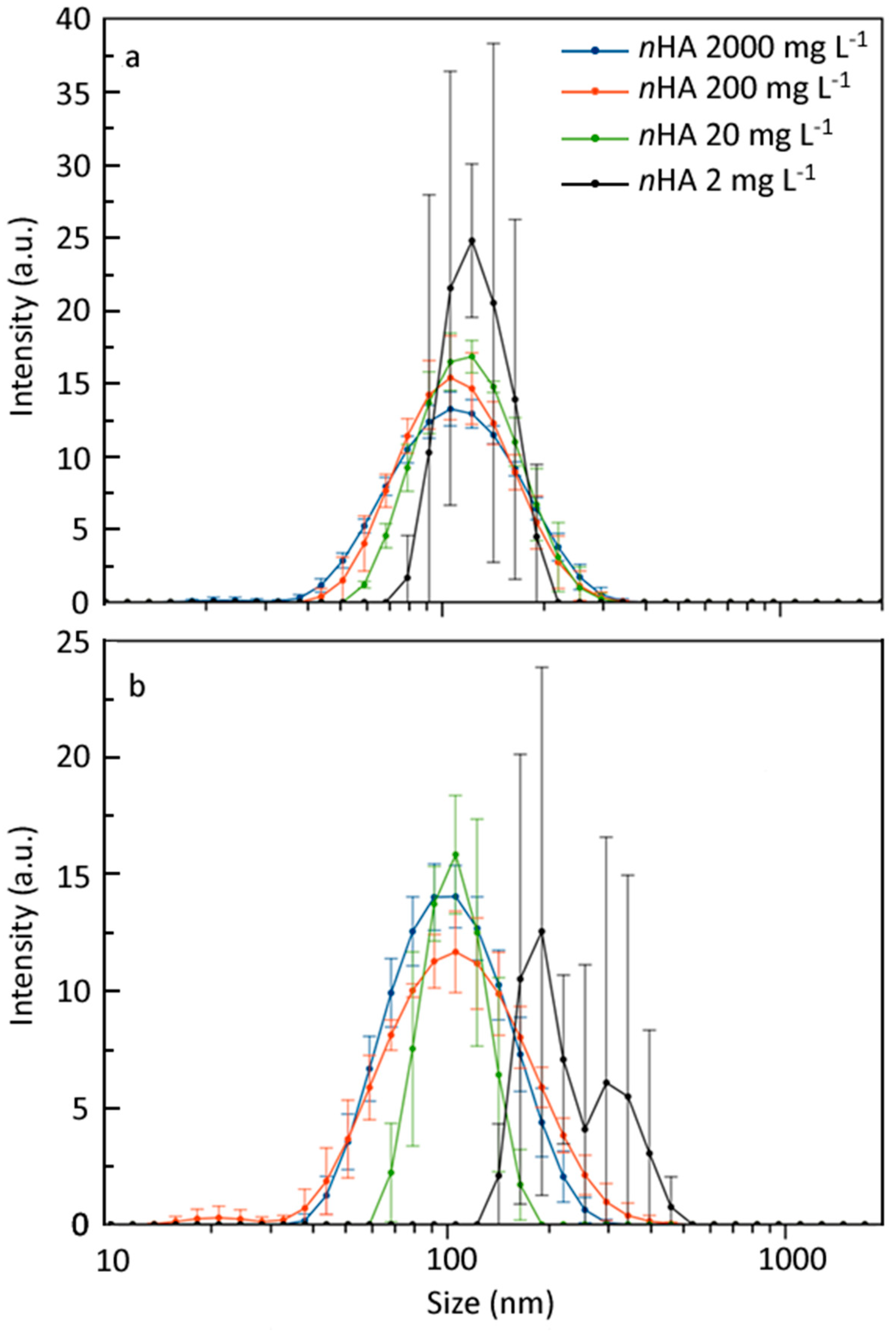
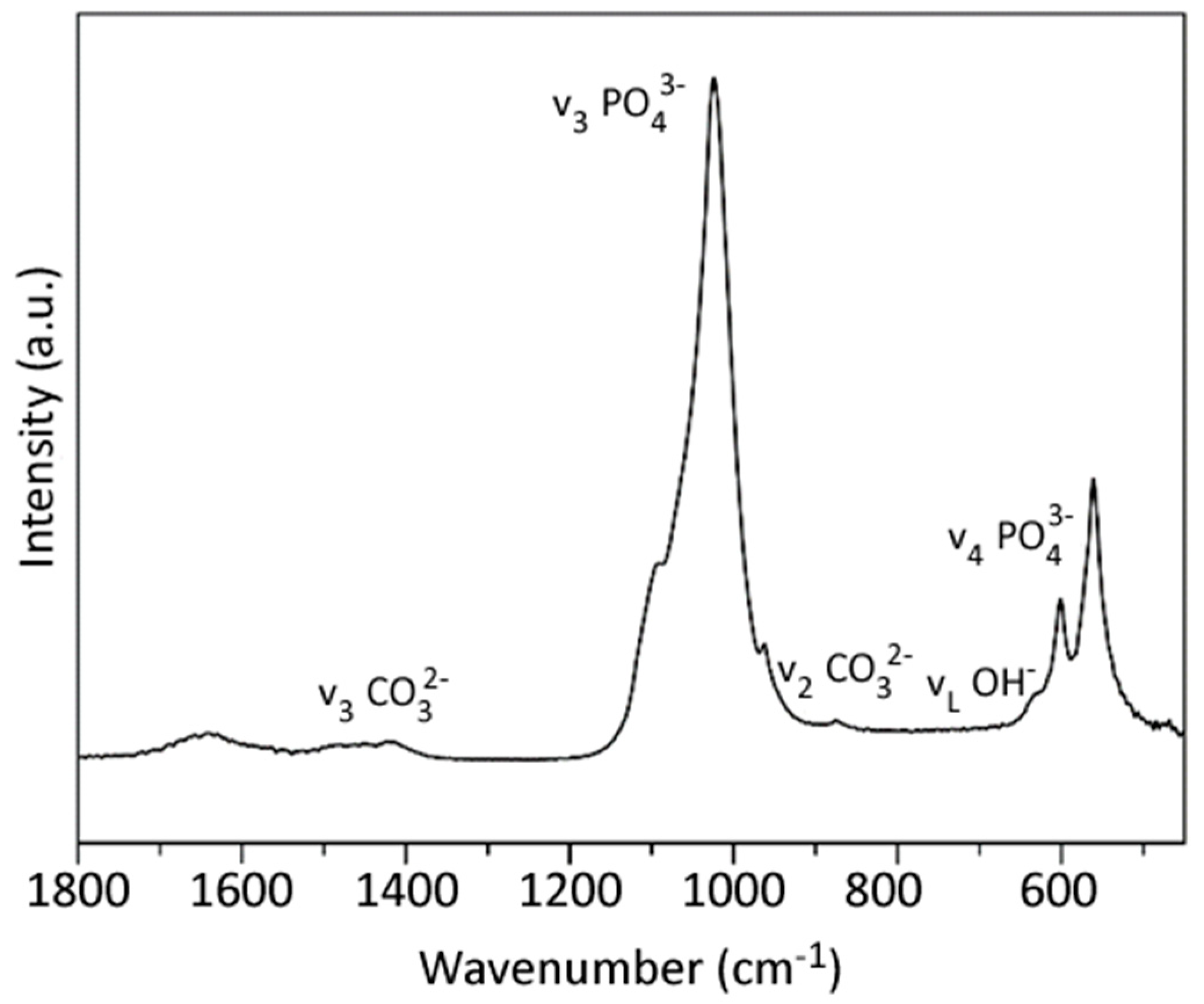
3.1. nHA and CMC-nHA Characterization
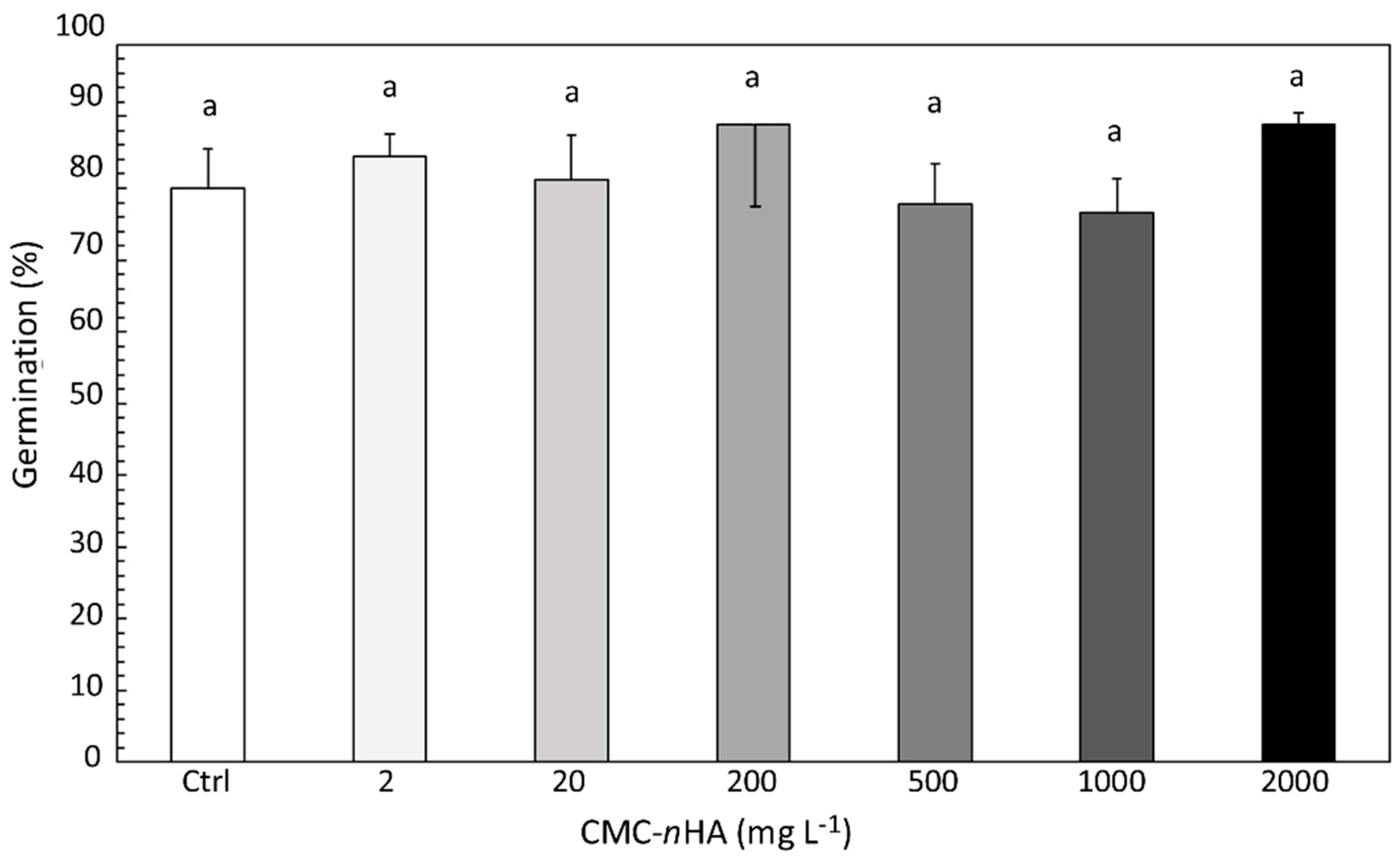
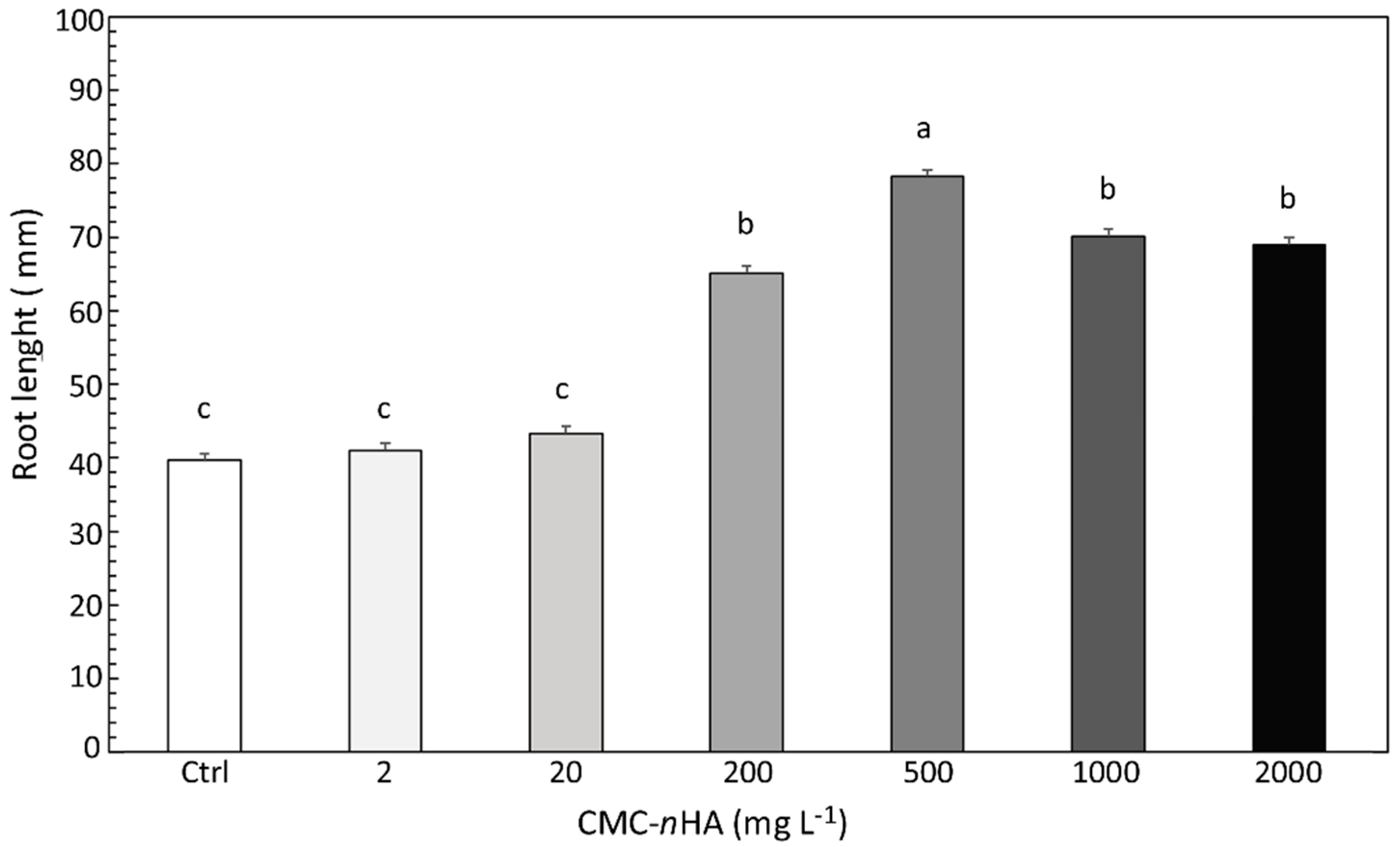
3.2. Seed Germination and Root Elongation
3.3. Plant Metabolism
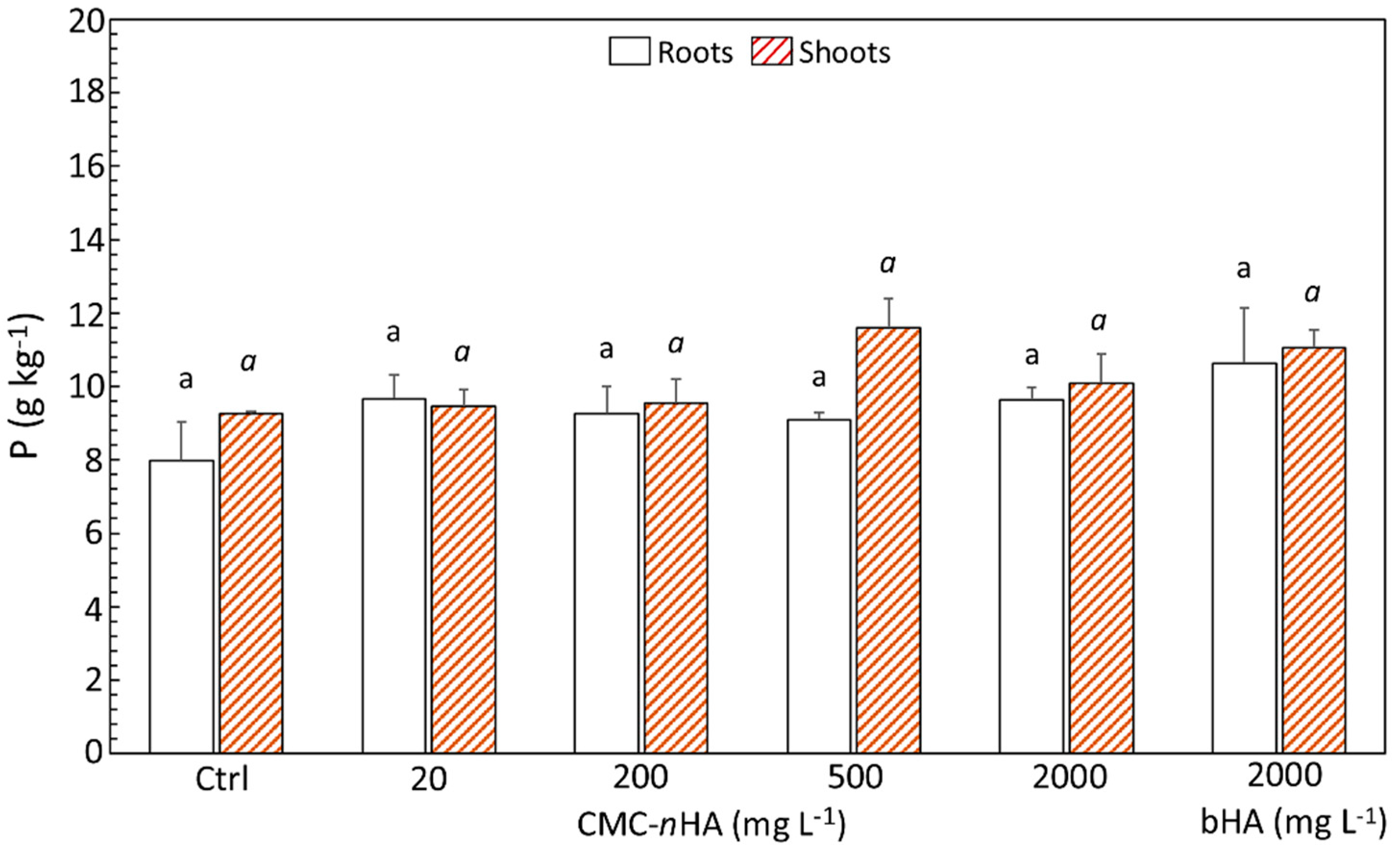
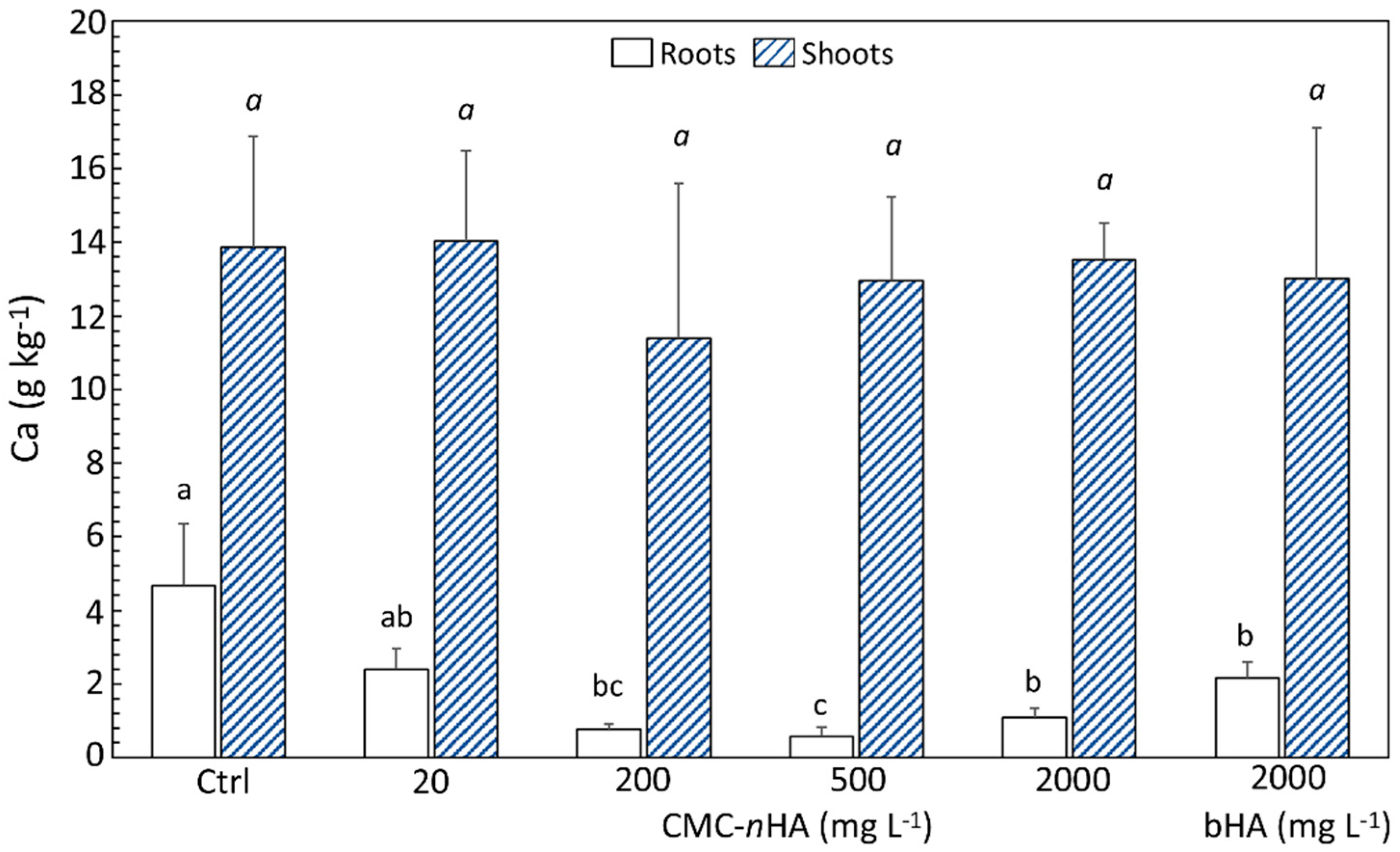
3.4. P and Ca Content
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pisante, M.; Stagnari, F.; Grant, C.A. Agricultural innovations for sustainable crop production intensification. Ital. J. Agron. 2012, 7, 40. [Google Scholar] [CrossRef]
- López-Arredondo, D.L.; Sánchez-Calderón, L.; Yong-Villalobos, L. Molecular and genetic basis of plant macronutrient use efficiency: Concepts, opportunities and challenges. In Plant Macronutrient Use Efficiency Molecular and Genomic Perspectives in Crop Plants; Hossain, M.A., Kamiya, T., Burritt, D.Y., Phan Tran, L.-S., Fujiwara, T., Eds.; Academic Press: Cambridge, MA, USA, 2017; pp. 1–29. ISBN 978-0-12-811308-0. [Google Scholar]
- Baligar, V.C.; Fageria, N.K. Nutrient use efficiency in plants: An overview. In Nutrient Use Efficiency: From Basics to Advances; Rakshit, A., Singh, H.B., Sen, A., Eds.; Springer: New Delhi, India, 2015; pp. 1–14. ISBN 978-81-322-2168-5. [Google Scholar]
- Liu, R.; Lal, R. Potentials of engineered nanoparticles as fertilizers for increasing agronomic productions. Sci. Total Environ. 2015, 514, 131–139. [Google Scholar] [CrossRef]
- Wang, P.; Lombi, E.; Zhao, F.-J.; Kopittke, P.M. Nanotechnology: A new opportunity in plant sciences. Trends Plant Sci. 2016, 21, 699–712. [Google Scholar] [CrossRef]
- Duhan, J.S.; Kumar, R.; Kumar, N.; Kaur, P.; Nehra, K.; Duhan, S. Nanotechnology: The new perspective in precision agriculture. Biotechnol. Rep. 2017, 125, 11–23. [Google Scholar] [CrossRef]
- Raliya, R.; Saharan, V.; Dimkpa, C.; Biswas, P. Nanofertilizer for precision and sustainable agriculture: Current state and future perspectives. J. Agric. Food Chem. 2018, 66, 6487–6503. [Google Scholar] [CrossRef]
- Gogos, A.; Knauer, K.; Bucheli, T.D. Nanomaterials in plant protection and fertilization: Current state, foreseen applications, and research priorities. J. Agric. Food Chem. 2012, 60, 9781–9792. [Google Scholar] [CrossRef] [PubMed]
- Kah, M.; Kookana, R.S.; Gogos, A.; Bucheli, T.D. A critical evaluation of nanopesticides and nanofertilizers against their conventional analogues. Nat. Nanotechnol. 2018, 13, 677–684. [Google Scholar] [CrossRef]
- Dimkpa, C.O.; Bindraban, P.S. Nanofertilizers: New products for the industry? J. Agric. Food Chem. 2018, 66, 6462–6473. [Google Scholar] [CrossRef] [PubMed]
- Calabi-Floody, M.; Medina, J.; Rumpel, C.; Condron, L.M.; Hernandez, M.; Dumont, M.; de la Luz Mora, M. Smart fertilizers as a strategy for sustainable agriculture. Adv. Agron. 2018, 147, 119–157. [Google Scholar] [CrossRef]
- Gómez-Morales, J.; Iafisco, M.; Delgado López, J.M.; Stéphanie, S.; Drouet, C. Progress on the preparation of nanocrystalline apatites and surface characterization: Overview of fundamental and applied aspects. Prog. Cryst. Growth Charact. Mater. 2013, 59, 1–46. [Google Scholar] [CrossRef] [Green Version]
- Tampieri, A.; Iafisco, M.; Sprio, S.; Ruffini, A.; Panseri, S.; Montesi, M.; Adamiano, A.; Sandri, M. Hydroxyapatite: From Nanocrystals to Hybrid Nanocomposites for Regenerative Medicine. In Handbook of Bioceramics and Biocomposites; Antoniac, I., Ed.; Springer: Cham, Switzerland, 2016; pp. 119–144. [Google Scholar] [CrossRef]
- Sprio, S.; Sandri, M.; Ruffini, A.; Adamiano, A.; Iafisco, M.; Dapporto, M.; Panseri, S.; Montesi, M.; Tampieri, A. Tissue Engineering and Biomimetics with Bioceramics. In Advances in Ceramic Biomaterials; Palmero, P., Cambier, F., De Barra, E., Eds.; Woodhead Publishing: Cambridge, UK, 2017; pp. 407–432. [Google Scholar] [CrossRef]
- Kottegoda, N.; Munaweera, I.; Madusanka, N.; Karunaratne, V. A green slow-release fertilizer composition based on urea-modified hydroxyapatite nanoparticles encapsulated wood. Curr. Sci. 2011, 101, 73–78. [Google Scholar]
- Subbaiya, R.; Priyanka, M.; Masilamani Selvam, M. Formulation of green nano-fertilizer to enhance the plant growth through slow and sustained release of nitrogen. J. Pharm. Res. 2012, 5, 5178–5183. [Google Scholar]
- Kottegoda, N.; Sandaruwan, C.; Priyadarshana, G.; Siriwardhana, A.; Rathnayake, U.A.; Berugoda Arachchige, D.M.; Kumarasinghe, A.R.; Dahanayake, D.; Karunaratne, V.; Amaratunga, G.A.J. Urea-hydroxyapatite nanohybrids for slow release of nitrogen. ACS Nano 2017, 11, 1214–1221. [Google Scholar] [CrossRef]
- Gunaratne, G.P.; Kottegoda, N.; Madusanka, N.; Munaweera, I.; Sandaruwan, C.; Priyadarshana, W.M.G.I.; Siriwardhana, A.; Madhushanka, B.A.D.; Rathnayake, U.A.; Karunaratne, V. Two new plant nutrient nanocomposites based on urea coated hydroxyapatite: Efficacy and plant uptake. Indian J. Agric. Sci. 2016, 86, 494–499. [Google Scholar]
- Madusanka, N.; Sandaruwana, C.; Kottegoda, N.; Sirisena, D.; Munaweera, I.; De Alwis, A.; Karunaratnea, V.; Amaratunga, G.A.J. Urea–hydroxyapatite-montmorillonite nanohybrid composites as slow release nitrogen compositions. Appl. Clay Sci. 2017, 150, 303–308. [Google Scholar] [CrossRef]
- Liu, R.; Lal, R. Synthetic apatite nanoparticles as a phosphorus fertilizer for soybean (Glycine max). Sci. Rep. 2014, 4, 5686–5691. [Google Scholar] [CrossRef]
- Montalvo, D.; McLaughlin, M.J.; Degryse, F. Efficacy of hydroxyapatite nanoparticles as phosphorus fertilizer in andisols and oxisols. Soil Sci. Soc. Am. J. 2015, 79, 551–558. [Google Scholar] [CrossRef]
- Sandhöfer, B.; Meckel, M.; Delgado-López, J.M.; Patrício, T.; Tampieri, A.; Rösch, F.; Iafisco, M. Synthesis and preliminary in vivo evaluation of well-dispersed biomimetic nanocrystalline apatites labeled with positron emission tomographic imaging agents. ACS Appl. Mater. Interfaces 2015, 19, 10623–10633. [Google Scholar] [CrossRef]
- Filippi, A.; Zancani, M.; Petrussa, E.; Braidot, E. Caspase-3-like activity and proteasome degradation in grapevine suspension cell cultures undergoing silver-induced programmed cell death. J. Plant Physiol. 2018, 233, 42–51. [Google Scholar] [CrossRef]
- Mobin, M.; Khan, N.A. Photosynthetic activity, pigment composition and antioxidative response of two mustard (Brassica juncea) cultivars differing in photosynthetic capacity subjected to cadmium stress. J. Plant Physiol. 2007, 164, 601–610. [Google Scholar] [CrossRef]
- Mattiello, A.; Pošćić, F.; Musetti, R.; Giordano, C.; Vischi, M.; Filippi, A.; Bertolini, A.; Marchiol, L. Evidences of genotoxicity and phytotoxicity in Hordeum vulgare exposed to CeO2 and TiO2 nanoparticles. Front. Plant Sci. 2015, 6, 1043. [Google Scholar] [CrossRef]
- Willett, W.; Rockström, J.; Loken, B.; Springmann, M.; Lang, T.; Vermeulen, S.; Garnett, T.; Tilman, D.; DeClerck, F.; Wood, A.; et al. Food in the Anthropocene: The EAT–Lancet Commission on healthy diets from sustainable food systems. Lancet 2019. [Google Scholar] [CrossRef]
- Naderi, M.R.; Danesh-Shahraki, A. Nanofertilizers and their roles in sustainable agriculture. Int. J. Agric. Crop Sci. 2013, 5, 2229–2232. [Google Scholar]
- Sekhon, B.S. Nanotechnology in agri-food production: An overview. Nanotechnol. Sci. Appl. 2014, 7, 31–53. [Google Scholar] [CrossRef]
- Rajput, V.D.; Minkina, T.; Sushkova, S.; Tsitsuashvili, V.; Mandzhieva, S.; Gorovtsov, A.; Nevidomskyaya, D.; Gromakova, N. Effect of nanoparticles on crops and soil microbial communities. J. Soils Sedim. 2018, 18, 2179–2187. [Google Scholar] [CrossRef]
- Iavicoli, I.; Leso, V.; Beezhold, D.H.; Shvedova, A.A. Nanotechnology in agriculture: Opportunities, toxicological implications, and occupational risks. Toxicol. Appl. Pharm. 2017, 329, 96–111. [Google Scholar] [CrossRef]
- Lin, S.; Yu, T.; Yu, Z.; Hu, X.; Yin, D. Nanomaterials Safer-by-Design: An Environmental Safety Perspective. Adv. Mater. 2018, 1705691. [Google Scholar] [CrossRef]
- Iafisco, M.; Bosco, R.; Leeuwenburgh, S.C.; van den Beucken, J.J.J.P.; Jansen, J.A.; Prat, M.; Roveri, N. Electrostatic spray deposition of biomimetic nanocrystalline apatite coatings onto titanium. Adv. Eng. Mater. 2012, 14, B13–B20. [Google Scholar] [CrossRef]
- Bosco, R.; Iafisco, M.; Tampieri, A.; Jansen, J.A.; Leeuwenburgh, S.C.; Van Den Beucken, J.J. Hydroxyapatite nanocrystals functionalized with alendronate as bioactive components for bone implant coatings to decrease osteoclastic activity. Appl. Surf. Sci. 2015, 328, 516–524. [Google Scholar] [CrossRef]
- Husen, A.; Siddiqi, K.S. Carbon and fullerene nanomaterials in plant system. J. Nanobiotechnol. 2014, 12. [Google Scholar] [CrossRef]
- Vithanage, M.; Seneviratne, M.; Ahmad, M.; Sarkar, B.; Ok, Y.S. Contrasting effects of engineered carbon nanotubes on plants: A review. Environ. Geochem. Health 2017, 39, 1421–1439. [Google Scholar] [CrossRef]
- Dimkpa, C.O. Soil properties influence the response of terrestrial plants to metallic nanoparticles exposure. Curr. Opin. Environ. Sci. Health 2018, 6, 1–8. [Google Scholar] [CrossRef]
- Pandey, K.; Lahiani, M.H.; Hicks, V.K.; Hudson, M.K.; Green, M.J.; Khodakovskaya, M. Effects of carbon-based nanomaterials on seed germination, biomass accumulation and salt stress response of bioenergy crops. PLoS ONE 2018, 13, e0202274. [Google Scholar] [CrossRef]
- Rico, C.M.; Majumdar, S.; Duarte-Gardea, M.; Peralta-Videa, J.R.; Gardea-Torresdey, J.L. Interaction of nanoparticles with edible plants and their possible implications in the food chain. J. Agric. Food Chem. 2011, 59, 3485–3498. [Google Scholar] [CrossRef] [Green Version]
- Miralles, P.; Church, T.L.; Harris, A.T. Toxicity, uptake, and translocation of engineered nanomaterials in vascular plants. Environ. Sci. Technol. 2012, 46, 9224–9239. [Google Scholar] [CrossRef]
- Lahiani, M.H.; Dervishi, E.; Ivanov, I.; Chen, J.; Khodakovskaya, M. Comparative study of plant responses to carbon-based nanomaterials with different morphologies. Nanotechnology 2016, 27, 265102. [Google Scholar] [CrossRef]
- Siddiqui, M.H.; Al-Whaibi, M.H. Role of nano-SiO2 in germination of tomato (Lycopersicum esculentum) seeds mill. Saudi J. Biol. Sci. 2014, 21, 13–17. [Google Scholar] [CrossRef]
- Raliya, R.; Biswasa, P.; Tarafdarm, J.C. TiO2 nanoparticle biosynthesis and its physiological effect on mung bean (Vigna radiata L.). Biotechnol. Rep. 2015, 5, 22–26. [Google Scholar] [CrossRef]
- Andersen, C.P.; King, G.; Plocher, M.; Storm, M.; Pokhrel, L.R.; Johnson, M.G.; Rygiewicz, P.T. Germination and early plant development of ten plant species exposed to titanium dioxide and cerium oxide nanoparticles. Environ. Toxicol. Chem. 2016, 35, 2223–2229. [Google Scholar] [CrossRef]
- De la Rosa, G.; López-Moreno, M.L.; de Haro, D.; Botez, C.E.; Peralta-Videa, J.R.; Gardea-Torresdey, J.L. Effects of ZnO nanoparticles in alfalfa, tomato, and cucumber at the germination stage: Root development and X-ray absorption spectroscopy studies. Pure Appl. Chem. 2013, 85, 2161–2174. [Google Scholar] [CrossRef] [Green Version]
- Khodakovskaya, M.V.; de Silva, K.; Nedosekin, D.A.; Dervishi, E.; Biris, A.S.; Shashkov, E.V.; Galanzha, E.I.; Zharov, V.P. Complex genetic, photothermal, and photoacoustic analysis of nanoparticle-plant interactions. Proc. Natl. Acad. Sci. USA 2011, 108, 1028–1033. [Google Scholar] [CrossRef]
- Haghighi, M.; Teixeira da Silva, J.A. The effect of carbon nanotubes on the seed germination and seedling growth of four vegetable species. J. Crop Sci. Biotechnol. 2014, 17, 201–208. [Google Scholar] [CrossRef]
- Morla, S.; Rao, C.R.; Chakrapani, R. Factors affecting seed germination and seedling growth of tomato plants cultured in vitro conditions. J. Chem. Biol. Phys. Sci. 2011, 1, 328–334. [Google Scholar]
- Tiwari, D.K.; Dasgupta-Schubert, N.; Villasenor, L.M.; Tripathi, D.; Villegas, J. Interaction of carbon nanotubes with mineral nutrients for the promotion of growth of tomato seedlings. Nano Stud. 2013, 7, 87–96. [Google Scholar]
- Zhang, M.; Gao, B.; Chen, J.; Li, Y.; Zhang, M.; Gao, B.; Chen, J.; Li, Y. Effects of graphene on seed germination and seedling growth. J. Nanoparticle Res. 2015, 17, 78. [Google Scholar] [CrossRef]
- Rouached, H.; Bulak Arpat, B.; Poirier, Y. Regulation of phosphate starvation responses in plants: Signaling players and cross-talks. Mol. Plant 2010, 3, 288–299. [Google Scholar] [CrossRef]
- Ogawa, S.; Gomez Selvaraj, M.; Fernando, A.J.; Lorieux, M.; Ishitani, M.; McCouch, S.; Arbelaez, J.D. N- and P-mediated seminal root elongation response in rice seedlings. Plant Soil 2014, 375, 303–315. [Google Scholar] [CrossRef]
- Bengough, A.G.; McKenzie, B.M.; Hallett, P.D.; Valentine, T.A. Root elongation, water stress, and mechanical impedance: A review of limiting stresses and beneficial root tip traits. J. Exp. Bot. 2011, 62, 59–68. [Google Scholar] [CrossRef]
- Williamson, L.C.; Ribrioux, S.P.C.P.; Fitter, A.H.; Ottoline Leyser, H.M. Phosphate availability regulates root system architecture in Arabidopsis. Plant Physiol. 2001, 126, 875–882. [Google Scholar] [CrossRef]
- De Silva, D.L.R.; Hetherington, A.M.; Mansfield, T.A. Where does all the calcium go? Evidence of an important regulatory role for trichomes in two calcicoles. Plant Cell Environ. 1996, 19, 880–886. [Google Scholar] [CrossRef]
- Tanner, W.; Beevers, H. Transpiration, a prerequisite for long distance transport of minerals in plants? Proc. Natl. Acad. Sci. USA 2001, 98, 9443–9447. [Google Scholar] [CrossRef]
- Heber, U.; Krause, G.H. What is the physiological role of photorespiration. Trends Biochem. Sci. 1980, 5, 32–34. [Google Scholar] [CrossRef]
- Kirkby, E.A.; Pilbeam, D.J. Calcium as a plant nutrient. Plant Cell Environ. 1984, 7, 397–405. [Google Scholar] [CrossRef]
- White, P.J.; Broadley, M.R. Calcium in plants. Ann. Bot. 2003, 92, 487–511. [Google Scholar] [CrossRef]
- Conn, S.; Gilliham, M. Comparative physiology of elemental distributions in plants. Ann Bot. 2010, 105, 1081–1102. [Google Scholar] [CrossRef] [Green Version]
- Dayod, M.; Tyerman, S.D.; Leigh, R.A.; Gilliham, M. Calcium storage in plants and the implications for calcium biofortification. Protoplasma 2010, 247, 215–231. [Google Scholar] [CrossRef]
- Hepler, P.H. Calcium: A Central Regulator of Plant Growth and Development. Plant Cell 2005, 17, 2142–2155. [Google Scholar] [CrossRef] [Green Version]
- Miedema, H.; Bothwell, J.H.; Brownlee, C.; Davies, J.M. Calcium uptake by plant cells -channels and pumps acting in concert. Trends Plant Sci. 2001, 6, 514–519. [Google Scholar] [CrossRef]
- Igamberdiev, A.U.; Hill, R.D. Elevation of cytosolic Ca2+ in response to energy deficiency in plants: The general mechanism of adaptation to low oxygen stress. Biochem. J. 2018, 475, 1411–1425. [Google Scholar] [CrossRef]
- Igamberdiev, A.U.; Kleczkowski, L.A. Magnesium and cell energetics in plants under anoxia. Biochem. J. 2011, 437, 373–379. [Google Scholar] [CrossRef] [Green Version]
- He, L.; Li, B.; Lu, X.; Yuan, L.; Yang, Y.; Yuan, Y.; Du, J.; Guo, S. The effect of exogenous calcium on mitochondria, respiratory metabolism enzymes and ion transport in cucumber roots under hypoxia. Sci. Rep. 2015, 5, 11391. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | D(002) (a) | D(310) (a) | Ca (b) | P (b) | Ca/P (b) | Carbonate (c) |
|---|---|---|---|---|---|---|
| Size (nm) | Size (nm) | w (%) | w (%) | (mol) | w (%) | |
| nHA | 16.5 | 3.6 | 31.6 ± 1.5 | 15.2 ± 0.3 | 1.60 ± 0.06 | 0.68 ± 0.07 |
| Treatment | Chl a | Chl b | Carotenoids | Chl a/Chl b | Pheophytin |
|---|---|---|---|---|---|
| (mg L−1) | (µg g−1 FW) | (µg g−1 FW) | (µg g−1 FW) | (%) | |
| 0 | 475 ± 100 a | 182 ± 47 ab | 131 ± 21 a | 3.18 ± 0.57 a | 8.75 ± 0.93 ab |
| 20 | 594 ± 174 a | 193 ± 30 a | 156 ± 47 a | 3.03 ± 0.44 a | 8.65 ± 0.34 b |
| 200 | 466 ± 148 a | 182 ± 18 ab | 115 ± 39 a | 2.66 ± 0.08 a | 10.01 ± 0.87 a |
| 500 | 430 ± 131 a | 154 ± 38 ab | 113 ± 34 a | 2.69 ± 0.43 a | 9.27 ± 1.22 ab |
| 2000 | 451 ± 99 a | 142 ± 11 b | 121 ± 31 a | 2.97 ± 0.18 a | 9.11 ± 0.42 ab |
| 2000 bHA | 524 ± 127 a | 161 ± 40 ab | 140 ± 35 a | 3.17 ± 0.16 a | 9.04 ± 1.27 ab |
| F | 0.883 | 1.576 | 0.952 | 1.155 | 1.118 |
| p | 0.509 ns | 0.209 ns | 0.468 ns | 0.363 ns | 0.382 ns |
| Treatment | Root H2DCFDA Fluorescence | Shoot H2DCFDA Fluorescence | Root ATP | Shoot ATP |
|---|---|---|---|---|
| (mg L−1) | (A.U. mg−1 prot.) | (A.U. mg−1 prot.) | (nmol g−1 DW) | (nmol g−1 DW) |
| 0 | 93 ± 27 a | 57 ± 8 a | 0.90 ± 0.23 a | 28.87 ± 9.46 a |
| 20 | 75 ± 27 a | 50 ± 21 a | 0.91 ± 0.03 a | 28.37 ± 7.15 ab |
| 200 | 78 ± 20 a | 68 ± 15 a | 0.75 ± 0.23 ab | 25.66 ± 9.87 ab |
| 500 | 61 ± 9 a | 67 ± 14 a | 0.76 ± 0.19 ab | 18.18 ± 5.89 bc |
| 2000 | 74 ± 17 a | 68 ± 15 a | 0.50 ± 0.12 b | 14.27 ± 6.45 c |
| 2000 bHA | 83 ± 58 a | 66 ± 14 a | 0.68 ± 0.38 ab | 31.08 ± 2.07 a |
| F | 0.630 | 1.108 | 2.240 | 3.874 |
| p | 0.678 ns | 0.384 ns | 0.088 ns | 0.013 * |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marchiol, L.; Filippi, A.; Adamiano, A.; Degli Esposti, L.; Iafisco, M.; Mattiello, A.; Petrussa, E.; Braidot, E. Influence of Hydroxyapatite Nanoparticles on Germination and Plant Metabolism of Tomato (Solanum lycopersicum L.): Preliminary Evidence. Agronomy 2019, 9, 161. https://doi.org/10.3390/agronomy9040161
Marchiol L, Filippi A, Adamiano A, Degli Esposti L, Iafisco M, Mattiello A, Petrussa E, Braidot E. Influence of Hydroxyapatite Nanoparticles on Germination and Plant Metabolism of Tomato (Solanum lycopersicum L.): Preliminary Evidence. Agronomy. 2019; 9(4):161. https://doi.org/10.3390/agronomy9040161
Chicago/Turabian StyleMarchiol, Luca, Antonio Filippi, Alessio Adamiano, Lorenzo Degli Esposti, Michele Iafisco, Alessandro Mattiello, Elisa Petrussa, and Enrico Braidot. 2019. "Influence of Hydroxyapatite Nanoparticles on Germination and Plant Metabolism of Tomato (Solanum lycopersicum L.): Preliminary Evidence" Agronomy 9, no. 4: 161. https://doi.org/10.3390/agronomy9040161
APA StyleMarchiol, L., Filippi, A., Adamiano, A., Degli Esposti, L., Iafisco, M., Mattiello, A., Petrussa, E., & Braidot, E. (2019). Influence of Hydroxyapatite Nanoparticles on Germination and Plant Metabolism of Tomato (Solanum lycopersicum L.): Preliminary Evidence. Agronomy, 9(4), 161. https://doi.org/10.3390/agronomy9040161