Potential Use of Sweet Potato (Ipomoea batatas (L.) Lam.) to Suppress Three Invasive Plant Species in Agroecosystems (Ageratum conyzoides L., Bidens pilosa L., and Galinsoga parviflora Cav.)

 ,
,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Study Species
2.3. Experiment Design and Data Collection
2.4. Data Analyses
3. Results
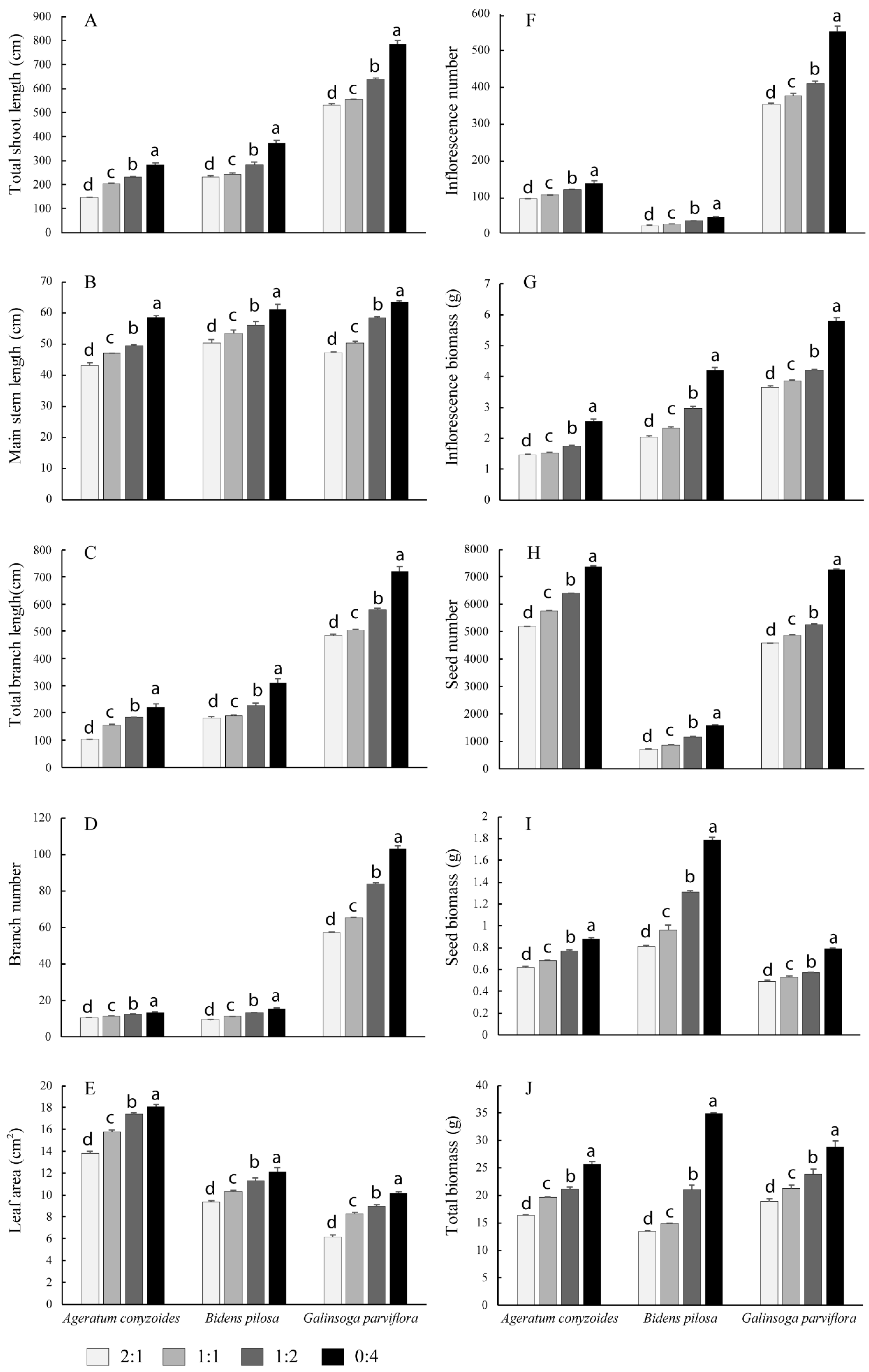
3.1. Plant Growth
3.2. Competitive Interactions
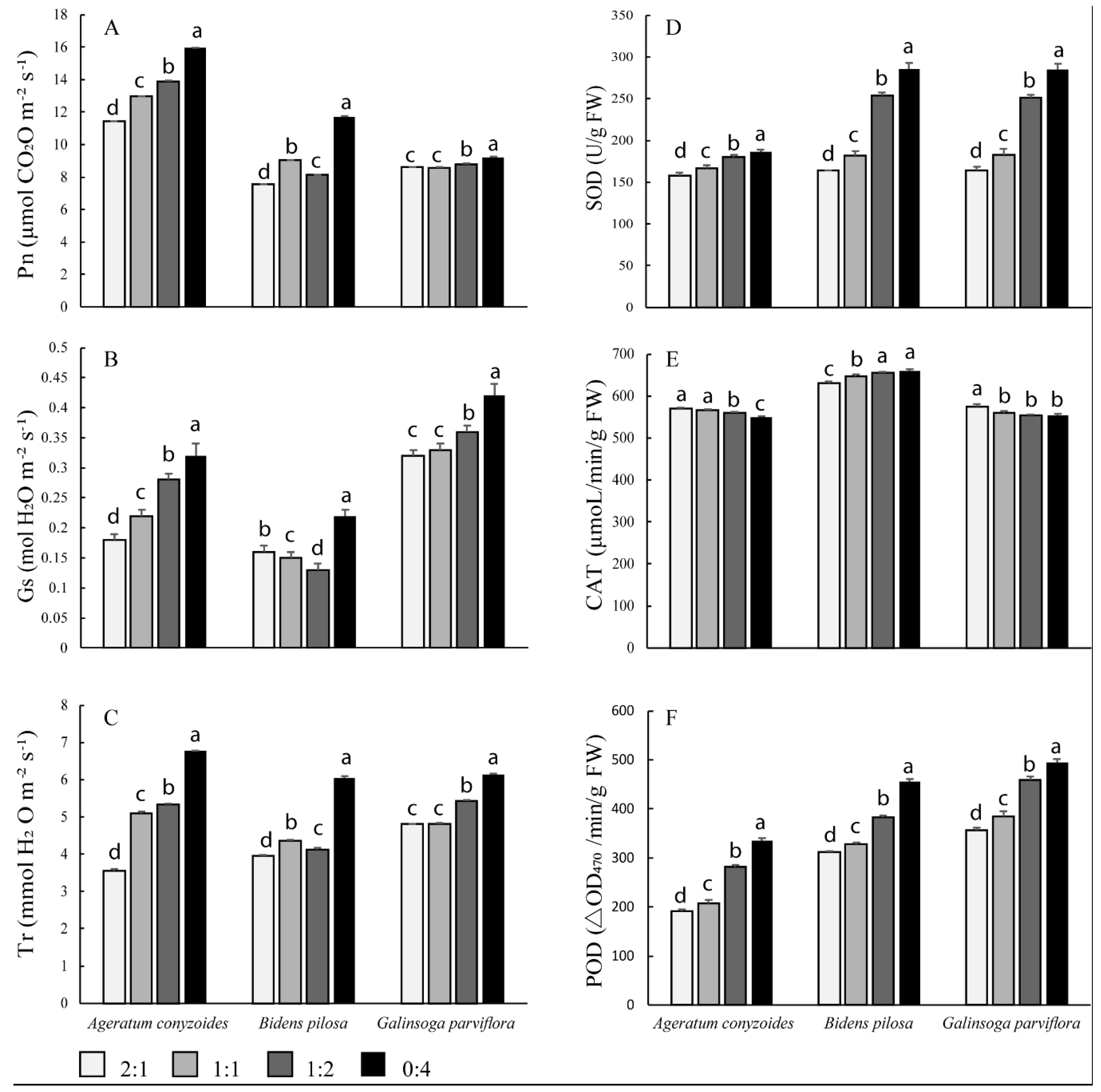
3.3. Photosynthesis and Enzyme Activities
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Simberloff, D.; Martin, J.L.; Genovesi, P.; Maris, V.; Wardle, D.A.; Aronson, J.; Courchamp, F.; Galil, B.; García-Berthou, E.; Pascal, M.; et al. Impacts of biological invasions: What’S what and the way forward. Trends Ecol. Evol. 2013, 28, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Blackburn, T.M.; Essl, F.; Evans, T.; Hulme, P.E.; Jeschke, J.M.; Kühn, I.; Kumschick, S.; Marková, Z.; Mrugała, A.; Nentwig, W.; et al. A unified classification of alien species based on the magnitude of their environmental impacts. PLoS Biol. 2014, 12, e1001850. [Google Scholar] [CrossRef] [PubMed]
- Day, M.D.; Clements, D.R.; Gile, C.; Senaratne, W.K.A.D.; Shen, S.C.; Weston, L.A.; Zhang, F.D. Biology and impacts of Pacific Islands invasive species. 13. Mikania micrantha Kunth (Asteraceae). Pac. Sci. 2016, 70, 257–285. [Google Scholar] [CrossRef]
- Van Bruggen, A.H.C.; He, M.M.; Shin, K.; Mai, V.; Jeong, K.C.; Finckh, M.R.; Morris, J.G., Jr. Environmental and health effects of the herbicide glyphosate. Sci. Total Environ. 2018, 616–617, 255–268. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.L.; Liu, W.X.; Wan, F.H.; Li, Z.Y. Measurements of plant competition ability and their applications: A review. Chin. J. Ecol. 2008, 27, 985–992. [Google Scholar]
- Shen, S.C.; Xu, G.F.; Clements, D.R.; Jin, G.M.; Chen, A.D.; Zhang, F.D.; Hisashi, K.N. Suppression of the invasive plant mile-a-minute (Mikania micrantha) by local crop sweet potato (Ipomoea batatas) by means of higher growth rate and competition for soil nutrients. BMC Ecol. 2015, 15, 1. [Google Scholar] [CrossRef] [PubMed]
- Lugo, A.E. The apparent paradox of reestablishing species richness on degraded lands with tree monocultures. Forest Ecol. Manag. 1997, 99, 9–19. [Google Scholar] [CrossRef]
- Li, W.H.; Luo, J.N.; Tian, X.S.; Chow, W.S.; Sun, Z.Y.; Zhang, T.J.; Peng, S.L.; Peng, C.L. A new strategy for controlling invasive weeds: selecting valuable native plants to defeat them. Sci. Rep. 2015, 5, 11004. [Google Scholar] [CrossRef]
- Sher, A.A.; Marshall, D.L.; Taylor, J.P. Establishment patterns of native Populus and Salix in the presence of invasive nonnative Tamarix. Ecol. Appl. 2002, 12, 760–772. [Google Scholar] [CrossRef]
- Shen, S.C.; Xu, G.F.; Clements, D.R.; Jin, G.M.; Liu, S.F.; Yang, Y.X.; Chen, A.D.; Zhang, F.D.; Hisashi, K.N. Suppression of reproductive characteristics of the invasive plant Mikania micrantha by sweet potato competition. BMC Ecol. 2016, 16, 30. [Google Scholar] [CrossRef]
- Sheley, R.L.; Mangold, J.M.; Anderson, J.L. Potential for successional theory to guide restoration of invasive-plant-dominated rangeland. Ecol. Monogr. 2006, 76, 365–379. [Google Scholar] [CrossRef]
- Keddy, P.; Nielsen, K.; Weiher, E.; Lawson, R. Relative competitive performance of 63 species of terrestrial herbaceous plants. J. Veg. Sci. 2002, 13, 5–16. [Google Scholar] [CrossRef]
- Williams, A.C.; McCarthy, B.C. A new index of interspecific competition for replacement and additive designs. Ecol. Res. 2001, 16, 29–40. [Google Scholar] [CrossRef]
- Xuan, T.D.; Shinkichi, T.; Hong, N.H.; Khanh, T.D.; Min, C. Assessment of phototoxic action of Ageratum conyzoides L. (billy goat weed) on weeds. Crop Prot. 2004, 915–922. [Google Scholar] [CrossRef]
- Arthur, G.D.; Naidoo, K.K.; Coopoosamy, R.M. Bidens pilosa L.: Agricultural and pharmaceutical importance. J. Med. Plants Res. 2012, 6, 3282–3287. [Google Scholar] [CrossRef]
- De Cauwer, B.; Devos, R.; Claerhout, S.; Bulcke, R.; Reheul, D. Seed dormancy, germination, emergence and seed longevity in Galinsoga parviflora and G. quadriradiata. Weed Res. 2014, 54, 38–47. [Google Scholar] [CrossRef]
- Holm, L.G.; Plucknett, D.L.; Pancho, J.V.; Herberger, J.P. The World’s Worst Weeds: Distribution and Biology; An East-West Centre Book; University Press of Hawaii: Honolulu, HI, USA, 1977; p. 609. [Google Scholar]
- Damalas, C.A. Distribution, biology, and agricultural importance of Galinsoga parviflora (Asteraceae). Weed Biol. Manag. 2008, 8, 147–153. [Google Scholar] [CrossRef]
- Sing, H.P.; Batish, D.R.; Kohli, R.K. Allelopathic effect of two volatile monoterpenes against billy goat weed (Ageratum conyzoides L.). Crop Prot. 2002, 21, 347–350. [Google Scholar] [CrossRef]
- Kong, C.H.; Hu, F.; Liang, W.J.; Wang, P.; Jiang, Y. Allelopathic potential of Ageratum conyzoides at various growth stages in different habitats. Allelopathy J. 2004, 13, 233–240. [Google Scholar]
- Shen, S.C.; Zhang, F.D.; Xu, G.F.; Li, T.L.; Wu, D.; Zhang, Y.H. Occurrence and infestation of invasive weed in crop field in Yunnan. Southwest China J. Agric. Sci. 2012, 25, 554–561. [Google Scholar]
- Xu, G.F.; Zhang, F.D.; Li, T.L.; Zhang, Y.; Zhang, Y.H. Allelopathy of 5 associated species on Mikania micrantha H.B.K. Southwest China J. Agric. Sci. 2009, 22, 1439–1443. [Google Scholar]
- Shen, S.C.; Xu, G.F.; Zhang, F.D.; Jin, G.M.; Zhang, Y.H. Effects of sweet potato (Ipomoea batas) cultivation on the community composition and diversity of weeds in arid fields. Biodivers Sci. 2014, 22, 485–491. [Google Scholar]
- Shen, S.C.; Xu, G.F.; Li, D.Y.; Clements, D.R.; Jin, G.M.; Liu, S.F.; Yang, Y.X.; Chen, A.D.; Zhang, F.D.; Hisashi, K.N. Allelopathic effects of three sweet potato cultivars (Ipomoea batatas) on the invasive plant Mikania micrantha. Pakistan J. Biol. Sci. 2018, 21, 8–15. [Google Scholar]
- Shen, S.C.; Xu, G.F.; Li, D.Y.; Clements, D.R.; Jin, G.M.; Liu, S.F.; Yang, Y.X.; Chen, A.D.; Zhang, F.D.; Hisashi, K.N. Allelopathic potential of sweet potato (Ipomoea batatas) germplasm resources of Yunnan Province in southwest China. Acta Ecol. Sin. 2018, 38, 444–449. [Google Scholar] [CrossRef]
- Shen, S.C.; Xu, G.F.; Zhang, F.D.; Jin, G.M.; Liu, S.F.; Yang, Y.X.; Zhang, Y.H. Allelopathic effects of water extracts from sweet potato (Ipomoea batatas) leaves on five major farming weeds. Acta Ecol. Sin. 2017, 37, 1931–1938. [Google Scholar]
- Jung, J.K.; Lee, S.U.; Kozukue, N.; Levin, C.E.; Friedman, M. Distribution of phenolic compounds and antioxidative activities in parts of sweet potato (Ipomoea batatas L.) plants and in home processed roots. J. Food Compos. Anal. 2011, 24, 29–37. [Google Scholar] [CrossRef]
- Pace, R.D.; Sibiya, T.E.; Phills, B.R.; Dull, G.G. Ca, Fe, and Zn content of ‘Jewel’ sweet potato greens as affected by harvesting practices. J. Food Sci. 1985, 50, 940–941. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, Z.; Lu, X.T.; Zhang, C.L.; Zhang, T.T.; Ma, S.Z. Investigation of weeds in sweet potato field of Tai’an, Shandong Province. Weed Sci. 2012, 30, 43–45. [Google Scholar]
- Harrison, H.F.; Peterson, J.K. Allelopathic effects of sweet potatoes (Ipomoea batatas) on yellow nutsedge (Cyperus esculentus) and alfalfa (Medicago sativa). Weed Sci. 1986, 34, 623–627. [Google Scholar] [CrossRef]
- Walker, D.W.; Hubbell, T.J.; Sedberry, J.E. Influence of decaying sweet potato crop residues on nutrient uptake of sweet potato plants. Agr. Ecosyst. Environ. 1989, 26, 45–52. [Google Scholar] [CrossRef]
- Chon, S.U.; Boo, H.O. Difference in allelopathic potential as influenced by root periderm colour of sweet potato (Ipomoea batatas). J. Agron. Crop Sci. 2005, 191, 75–80. [Google Scholar] [CrossRef]
- Xuan, T.D.; Toyama, T.; Khanh, T.D.; Tawata, S.; Nakagoshi, N. Allelopathic interference of sweet potato with cogongrass and relevant species. Plant Ecol. 2012, 213, 1955–1961. [Google Scholar] [CrossRef]
- de Wit, C.T. On competition. Versl. Landbouwk. Onderzoek. 1960, 66, 1–82. [Google Scholar]
- Rodríguez, D.J. A method to study competition dynamics using de Wit replacement series experiments. Oikos 1997, 78, 411–415. [Google Scholar] [CrossRef]
- Roush, M.L.; Radosevich, S.R.; Wagner, R.G.; Maxwell, B.D.; Petersen, T.D. A comparison of methods for measuring effects of density and proportion in plant competition experiments. Weed Sci. 1989, 37, 268–275. [Google Scholar] [CrossRef]
- Watkinson, A.R. Density-dependence in single-species populations of plants. J. Theor. Biol. 1980, 83, 345–357. [Google Scholar] [CrossRef]
- Orcutt, D.M.; Nilsen, E.T. Physiology of Plants under Stress: Soil and Biotic Factors; John Wiley & Sons: New York, NY, USA, 2000; pp. 412–413. [Google Scholar]
- Ara, N.; Nakkanong, K.; Lv, W.H.; Yang, J.H.; Hu, Z.Y.; Zhang, M.F. Antioxidant enzymatic activities and gene expression associated with heat tolerance in the stems and roots of two Cucurbit species (“Cucurbita maxima” and “Cucurbita moschata”) and their interspecific inbred line “Maxchata”. Int. J. Mol. Sci. 2013, 14, 24008–24028. [Google Scholar] [CrossRef]
- Jiang, Y.W.; Huang, B.R.; Jiang, Y. Effects of calcium on antioxidant activities and water relations associated with heat tolerance in two cool-season grasses. J. Exp. Bot. 2001, 52, 341–349. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.I. The Water-culture Method for Growing Plants without Soil; Agricultural Experiment Station Circular, College of Agriculture University of California: Berkley, CA, USA, 1950. [Google Scholar]
- Fowler, N. Competition and coexistence in a North Carolina grassland: III. mixtures of component species. J. Ecol. 1982, 70, 77–92. [Google Scholar] [CrossRef]
- Wilson, J.B. Shoot competition and root competition. J. Appl. Ecol. 1988, 25, 279–296. [Google Scholar] [CrossRef]
- Ekeleme, F.; Forcella, F.; Archer, D.W.; Akobundu, I.O.; Chikoye, D. Seedling emergence model for tropic ageratum (Ageratum conyzoides). Weed Sci. 2005, 53, 55–61. [Google Scholar] [CrossRef]
- Baldwin, I.T.; Schmelz, E.A. Constraints on an induce defense: the role of leaf area. Oecologia 1994, 97, 424–430. [Google Scholar] [CrossRef] [PubMed]
- Lambers, H.; Poorter, H. Inherent variation in growth rate between higher plants: a search for physiological causes and ecological consequences. Adv. Ecol. Res. 1992, 23, 187–261. [Google Scholar]
- Shi, Q.H.; Bao, Z.Y.; Zhu, Z.J.; Ying, Q.S.; Qian, Q.Q. Effects of different treatments of salicylic acid on heat tolerance, chlorophyll fluorescence, and antioxidant enzyme activity in seedlings of Cucumis sativa L. Plant Growth Regul. 2006, 48, 127–135. [Google Scholar] [CrossRef]
- Zhang, L.L.; Wen, D.Z. Photosynthesis, chlorophyll fluorescence, and antioxidant enzyme responses of invasive weed Mikania micrantha to Bemisia tabaci infestation. Photosynthetica 2008, 46, 457–462. [Google Scholar] [CrossRef]
- Lei, G.S.; Wang, W.Y.; Jiang, Z.L.; Deng, D.D.; Liu, W.X.; Gui, F.R.; Li, Z.Y. Competitive effects and physiological and biochemical properties of Ageratina adenophora and its associated species Chenopodium serotinum. Ecol. Environ. Sci. 2014, 23, 16–21. [Google Scholar]

{kind=link}
{kind=link}
| Variables | Ratios (Sweet Potato:Invasive Plant) | |||
|---|---|---|---|---|
| 2:1 | 1:1 | 1:2 | ||
| Sweet potato RY | A. conyzoides | 1.06 c ** | 1.10 b ** | 1.16 a ** |
| B. pilosa | 1.04 c ** | 1.10 b ** | 1.13 a ** | |
| G. parviflora | 1.08 c ** | 1.10 bc ** | 1.13 ab ** | |
| Competitive species RY | A. conyzoides | 0.63 c ** | 0.76 b ** | 0.82 a ** |
| B. pilosa | 0.38 c ** | 0.42 b ** | 0.60 a ** | |
| G. parviflora | 0.65 c ** | 0.74 b ** | 0.83 a ** | |
| RYT | A. conyzoides | 0.85 c ** | 0.93 b ** | 0.99 a * |
| B. pilosa | 0.71 c ** | 0.76 b ** | 0.87 a ** | |
| G. parviflora | 0.87 c ** | 0.92 b ** | 0.98 a ** | |
| CB index for sweet potato | A. conyzoides | 0.50 a ** | 0.37 bc ** | 0.34 c ** |
| B. pilosa | 0.99 a * | 0.95 b ** | 0.63 c ** | |
| G. parviflora | 0.50 a ** | 0.41 b ** | 0.31 c ** | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, S.; Xu, G.; Li, D.; Jin, G.; Liu, S.; Clements, D.R.; Yang, Y.; Rao, J.; Chen, A.; Zhang, F.; et al. Potential Use of Sweet Potato (Ipomoea batatas (L.) Lam.) to Suppress Three Invasive Plant Species in Agroecosystems (Ageratum conyzoides L., Bidens pilosa L., and Galinsoga parviflora Cav.). Agronomy 2019, 9, 318. https://doi.org/10.3390/agronomy9060318
Shen S, Xu G, Li D, Jin G, Liu S, Clements DR, Yang Y, Rao J, Chen A, Zhang F, et al. Potential Use of Sweet Potato (Ipomoea batatas (L.) Lam.) to Suppress Three Invasive Plant Species in Agroecosystems (Ageratum conyzoides L., Bidens pilosa L., and Galinsoga parviflora Cav.). Agronomy. 2019; 9(6):318. https://doi.org/10.3390/agronomy9060318
Chicago/Turabian StyleShen, Shicai, Gaofeng Xu, Diyu Li, Guimei Jin, Shufang Liu, David Roy Clements, Yanxian Yang, Jia Rao, Aidong Chen, Fudou Zhang, and et al. 2019. "Potential Use of Sweet Potato (Ipomoea batatas (L.) Lam.) to Suppress Three Invasive Plant Species in Agroecosystems (Ageratum conyzoides L., Bidens pilosa L., and Galinsoga parviflora Cav.)" Agronomy 9, no. 6: 318. https://doi.org/10.3390/agronomy9060318
APA StyleShen, S., Xu, G., Li, D., Jin, G., Liu, S., Clements, D. R., Yang, Y., Rao, J., Chen, A., Zhang, F., Zhu, X., & Weston, L. A. (2019). Potential Use of Sweet Potato (Ipomoea batatas (L.) Lam.) to Suppress Three Invasive Plant Species in Agroecosystems (Ageratum conyzoides L., Bidens pilosa L., and Galinsoga parviflora Cav.). Agronomy, 9(6), 318. https://doi.org/10.3390/agronomy9060318