Effect of Supplementary Light Intensity on Quality of Grafted Tomato Seedlings and Expression of Two Photosynthetic Genes and Proteins




Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Light Treatments
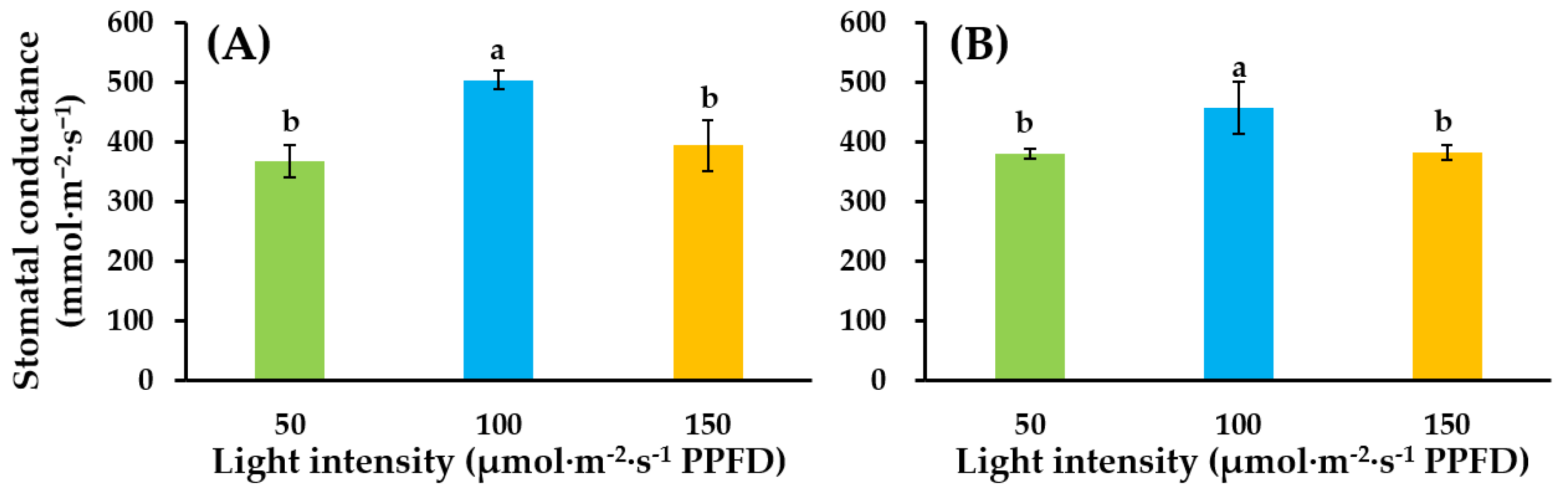
2.3. Stomatal Conductance
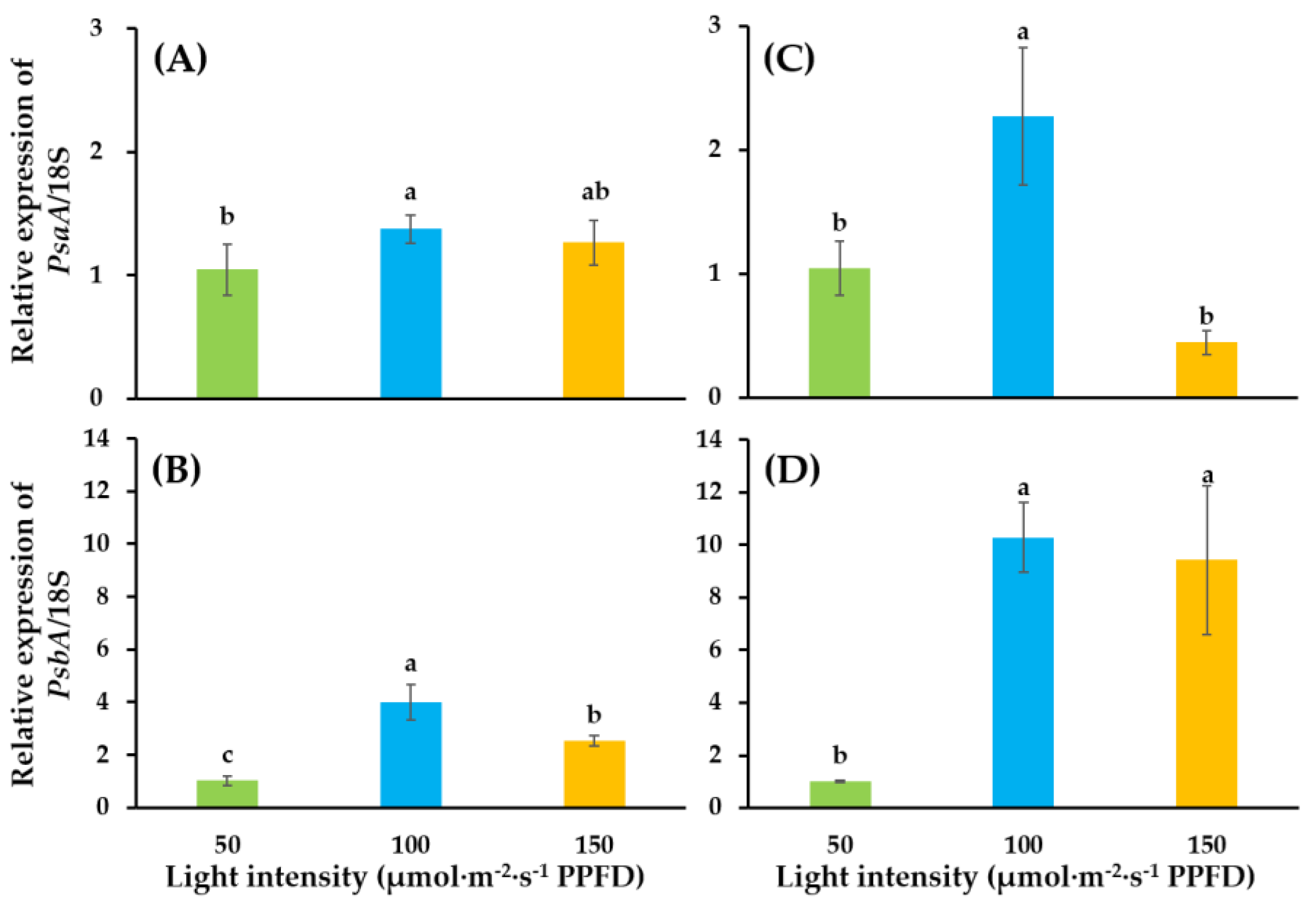
2.4. Quantitative Real-Time PCR Analysis
2.5. Protein Extraction and Western Blotting
2.6. Data Collection and Analysis
3. Results
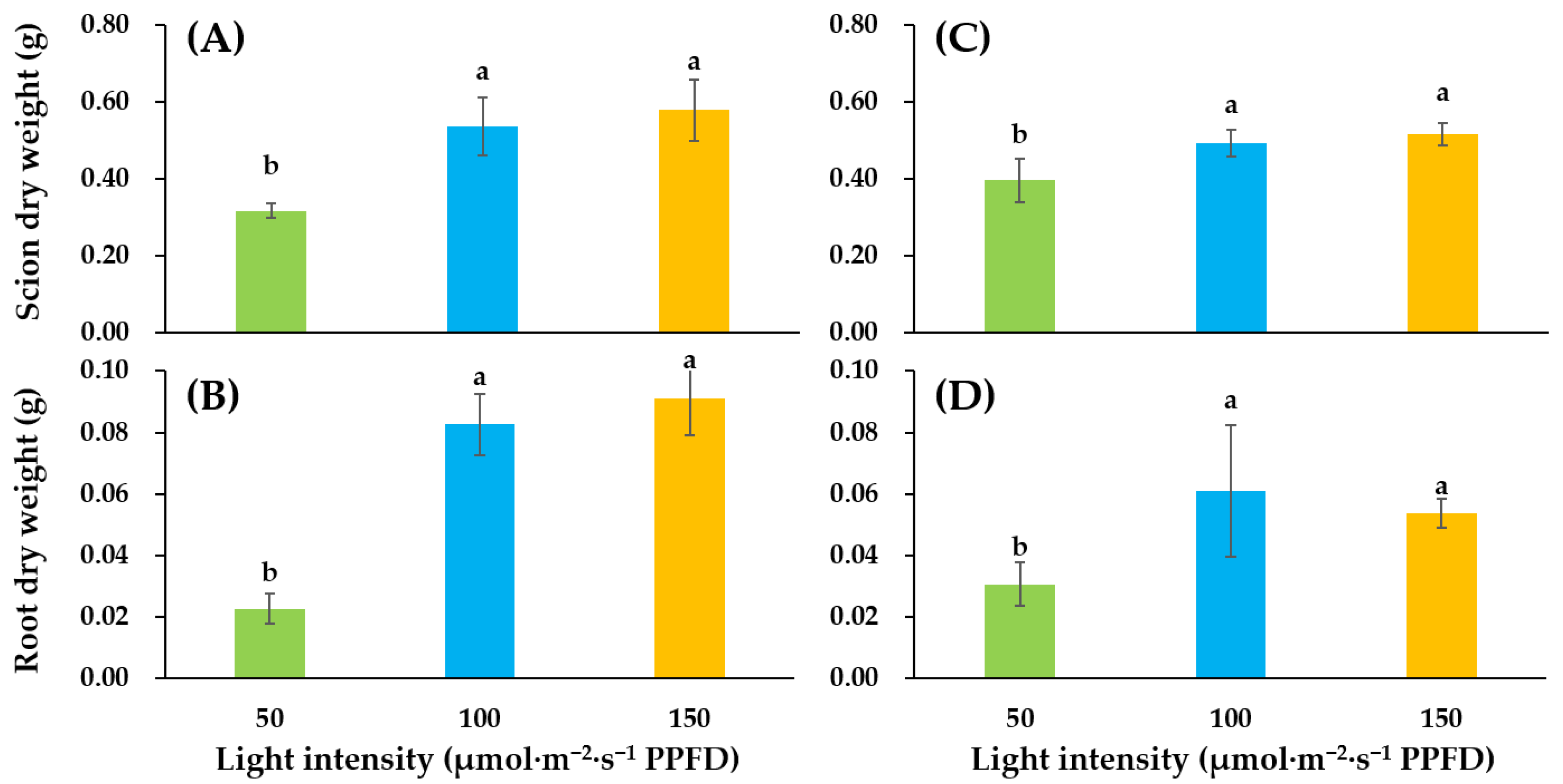
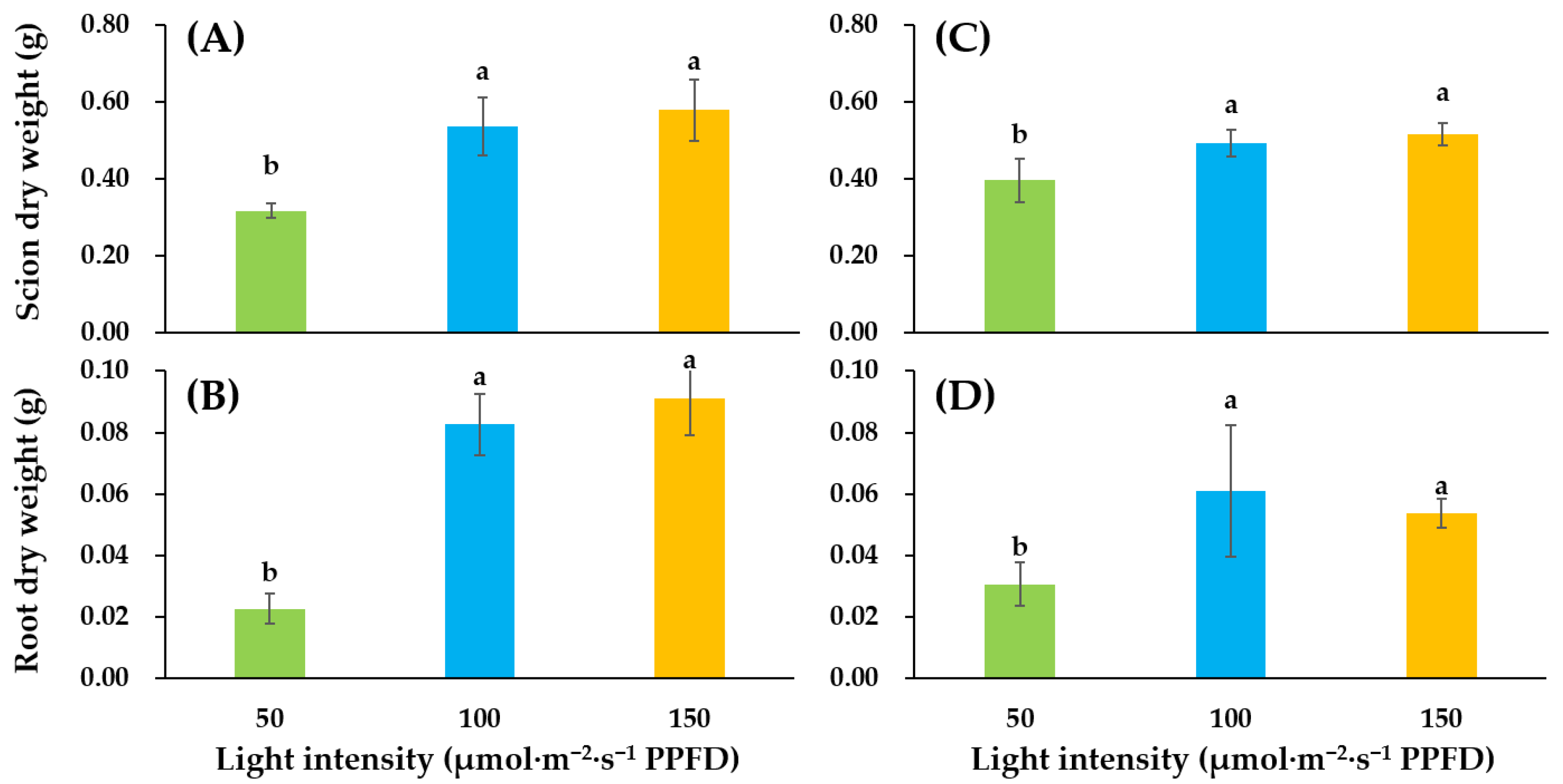
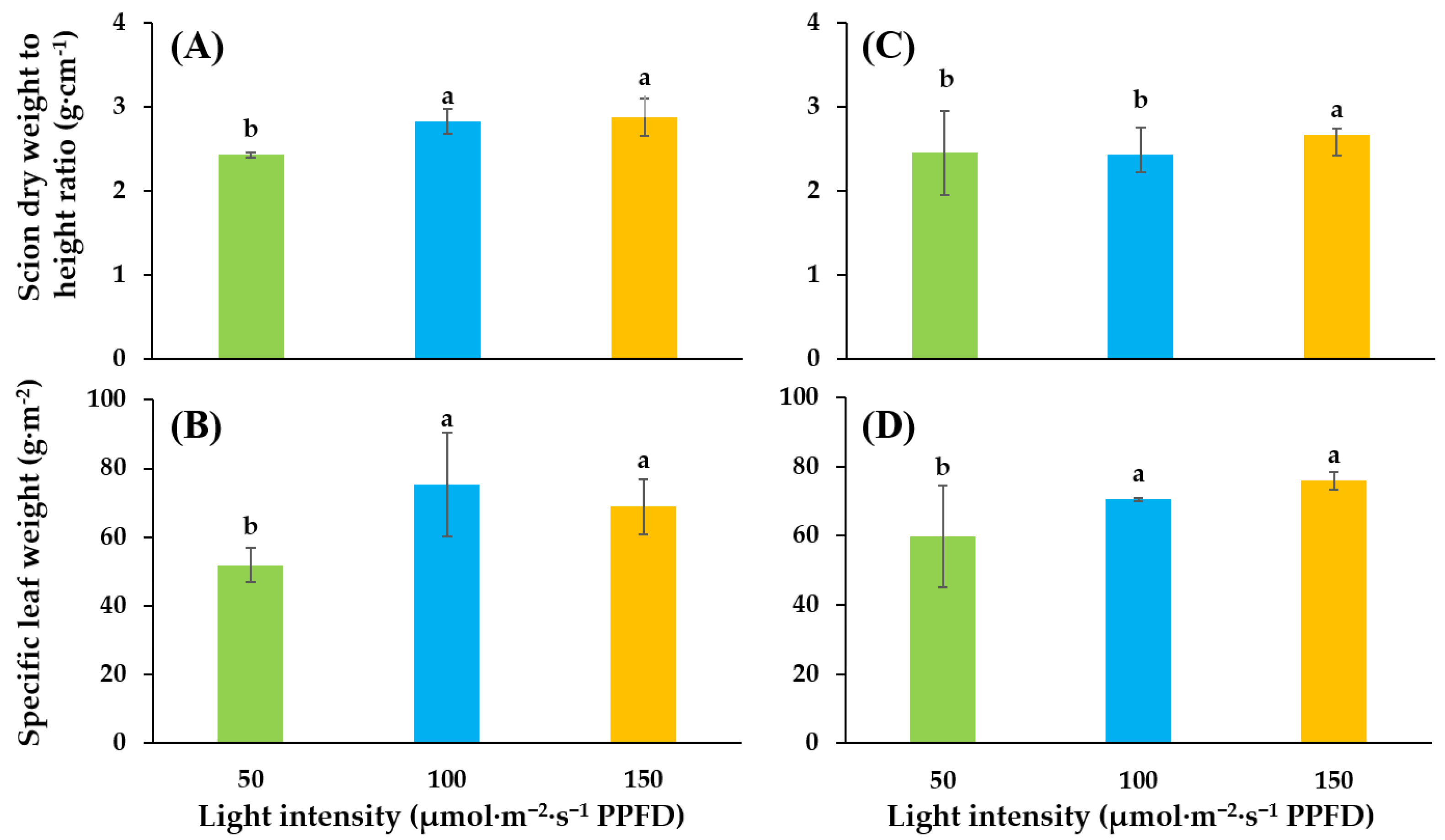
3.1. Growth and Development of Tomato Seedlings as Affected by Supplemental Lighting Treatment in the Glasshouse
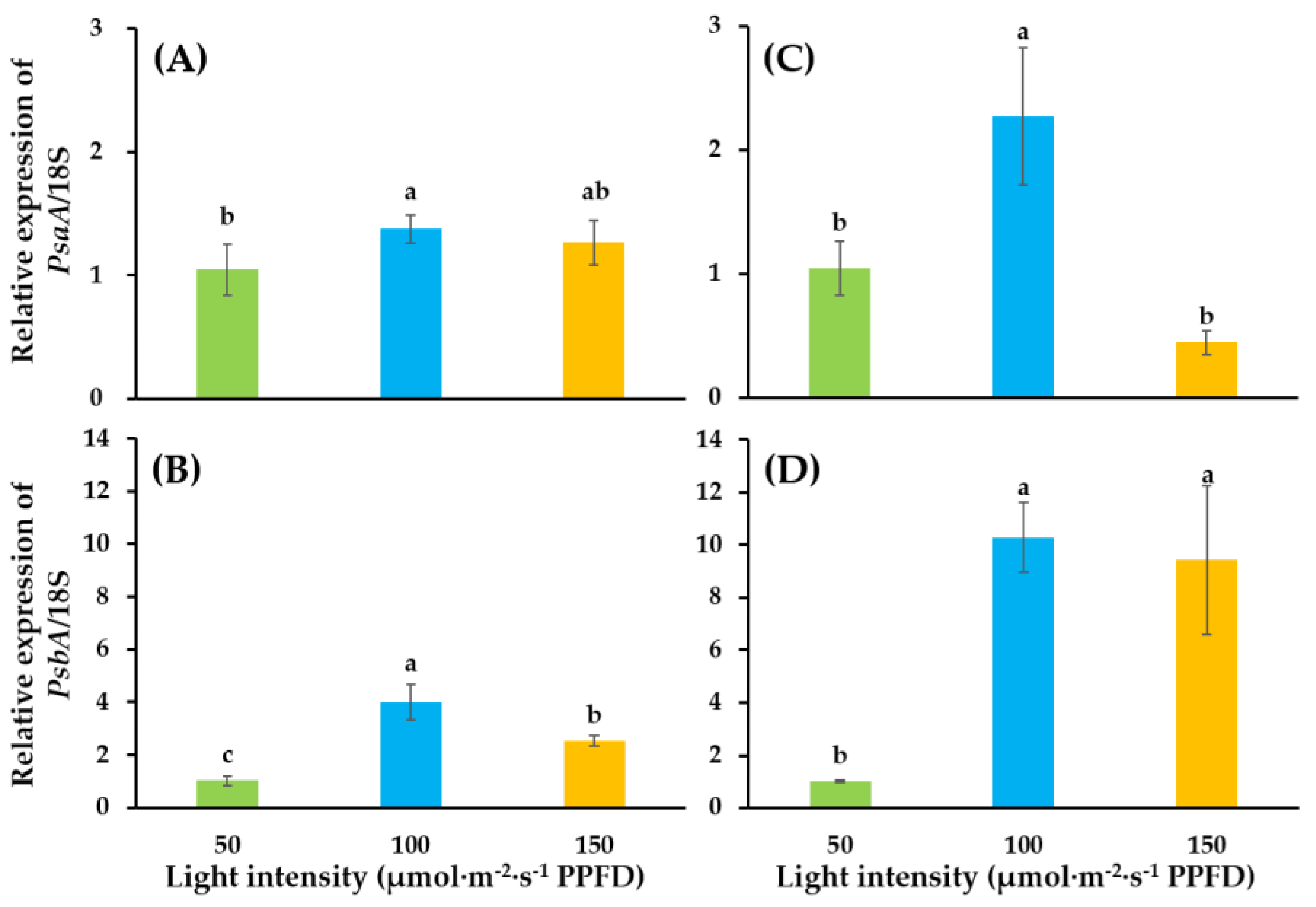
3.2. Expressions of Photosynthesis-Related Genes
3.3. PsaA and PsbA Immunoblots
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Muneer, S.; Kim, E.J.; Park, J.S.; Lee, J.H. Influence of green, red and blue light emitting diodes on multiprotein complex proteins and photosynthetic activity under different light intensities in lettuce leaves (Lactuca sativa L.). Int. J. Mol. Sci. 2014, 15, 4657–4670. [Google Scholar] [CrossRef] [PubMed]
- Berta, M.; Giovannelli, A.; Sebastiani, F.; Camussi, A.; Racchi, M.L. Transcriptome changes in the cambial region of poplar (Populus alba L.) in response to water deficit. Plant Biol. 2010, 12, 341–354. [Google Scholar] [CrossRef] [PubMed]
- Baba, K.; Karlberg, A.; Schmidt, J.; Schrader, J.; Hvidsten, T.R.; Bako, L.; Bhalerao, R.P. Activity-dormancy transition in the cambial meristem involves stage-specific modulation of auxin response in hybrid aspen. Proc. Natl. Acad. Sci. USA 2011, 108, 3418–3423. [Google Scholar] [CrossRef] [PubMed]
- Sorce, C.; Giovannelli, A.; Sebastiani, L.; Anfodillo, T. Hormonal signals involved in the regulation of cambial activity, xylogenesis and vessel patterning in trees. Plant Cell Rep. 2013, 32, 885–898. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.M.; Kubota, C.; Tsao, S.; Bie, Z.; Echevarria, P.H.; Morra, L.; Oda, M. Current status of vegetable grafting: Diffusion, grafting techniques, automation. Sci. Hortic. 2010, 127, 93–105. [Google Scholar] [CrossRef]
- Vu, N.T.; Kim, Y.S.; Kang, H.M.; Kim, I.S. Effect of red LEDs during healing and acclimatization process on the survival rate and quality of grafted tomato seedlings. Prot. Hortic. Plant Fact. 2014, 23, 43–49. [Google Scholar] [CrossRef]
- Yorio, N.C.; Goins, G.D.; Kagie, H.R.; Wheeler, R.M.; Sager, J.C. Improving spinach, radish, and lettuce growth under red light-emitting diodes (LEDs) with blue light supplementation. HortScience 2001, 36, 380–383. [Google Scholar] [CrossRef]
- Yano, A.; Fujiwara, K. Plant lighting system with five wavelength-band light-emitting diodes providing photon flux density and mixing ratio control. Plant Methods 2012, 8, 46. [Google Scholar] [CrossRef]
- Kim, K.; Kook, H.S.; Jang, Y.J.; Lee, W.H.; Kamala-Kannan, S.; Chae, J.C.; Lee, K.J. The effect of blue-light-emitting diodes on antioxidant properties and resistance to Botrytis cinerea in tomato. J. Plant Pathol. Microbiol. 2013, 4, 203. [Google Scholar] [CrossRef]
- Beon, M.S.; Bartsch, N. Early seedling growth of pine (Pinus densiflora) and oaks (Quercus serrata, Q. mongolica, Q. variabilis) in response to light intensity and soil moisture. Plant Ecol. 2003, 167, 97–105. [Google Scholar] [CrossRef]
- Du, Y.X.; Xin, J.; Zhang, J.; Li, J.Z.; Sun, H.Z.; Zhao, Q.Z. Research progress on the impacts of low light intensity on rice growth and development. Chin. J. Eco-Agric. 2013, 11, 1307–1317. (In Chinese) [Google Scholar] [CrossRef]
- Perrin, P.M.; Mitchell, F.J. Effects of shade on growth, biomass allocation and leaf morphology in European yew (Taxus baccata L.). Eur. J. For. Res. 2013, 132, 211–218. [Google Scholar] [CrossRef]
- Shao, Q.; Wang, H.; Guo, H.; Zhou, A.; Huang, Y.; Sun, Y.; Li, M. Effects of shade treatments on photosynthetic characteristics, chloroplast ultrastructure, and physiology of Anoectochilus roxburghii. PLoS ONE 2014, 9, e85996. [Google Scholar] [CrossRef] [PubMed]
- Nagel, K.A.; Schurr, U.; Walter, A. Dynamics of root growth stimulation in Nicotiana tabacum in increasing light intensity. Plant Cell Environ. 2006, 29, 1936–1945. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Hu, J.T.; Liu, C.; Wang, M.Z.; Zhao, J.; Kang, D.I.; Jeong, B.R. Effect of supplementary light source on quality of grafted tomato seedlings and expression of two photosynthetic genes. Agronomy 2018, 8, 207. [Google Scholar] [CrossRef]
- Li, Y.Y.; Mao, K.; Zhao, C.; Zhao, X.Y.; Zhang, H.L.; Shu, H.R.; Hao, Y.J. MdCOP1 ubiquitin E3 ligases interact with MdMYB1 to regulate light-induced anthocyanin biosynthesis and red fruit coloration in apple. Plant Physiol. 2012, 160, 1011–1022. [Google Scholar] [CrossRef]
- Choquet, Y.; Goldschmidt-Clermont, M.; Girard-Bascou, J.; Kück, U.; Bennoun, P.; Rochaix, J.D. Mutant phenotypes support a trans-splicing mechanism for the expression of the tripartite psaA gene in the C. reinhardtii chloroplast. Cell 1988, 52, 903–913. [Google Scholar] [CrossRef]
- Singh, M. Turnover of D1 protein encoded by PsbA gene in higher plants and cyanobacteria sustains photosynthetic efficiency to maintain plant productivity under photoinhibitory irradiance. Photosynthetica 2000, 38, 161–169. [Google Scholar] [CrossRef]
- Kang, J.H.; Krishna, K.S.; Atulba, S.L.S.; Jeong, B.R.; Hwang, S.J. Light intensity and photoperiod influence the growth and development of hydroponically grown leaf lettuce in a closed-type plant factory system. Hortic. Environ. Biotechnol. 2013, 54, 501–509. [Google Scholar] [CrossRef]
- Hunter, D.C.; Burritt, D.J. Light quality influences adventitious shoot production from cotyledon explants of lettuce (Lactuca sativa L). In Vitro Cell Dev. Biol. 2004, 40, 215–220. [Google Scholar] [CrossRef]
- Li, Q.; Kubota, C. Effects of supplemental light quality on growth and phytochemicals of baby leaf lettuce. Environ. Exp. Bot. 2009, 67, 59–64. [Google Scholar] [CrossRef]
- Franklin, K.A.; Larner, V.S.; Whitelam, G.C. The signal transducing photoreceptors of plants. ITLE Int. J. Dev. Biol. 2005, 49, 653–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, X.X.; Xu, Z.G.; Liu, X.Y.; Tang, C.M.; Wang, L.W.; Han, X.L. Effects of light intensity on the growth and leaf development of young tomato plants grown under a combination of red and blue light. Sci. Hortic. 2013, 153, 50–55. [Google Scholar] [CrossRef]
- Chitwood, D.H.; Kumar, R.; Ranjan, A.; Pelletier, J.M.; Townsley, B.T.; Ichihashi, Y.; Martinez, C.C.; Zumstein, K.; Harada, J.J.; Maloof, J.N.; et al. Light-Induced Indeterminacy Alters Shade-Avoiding Tomato Leaf Morphology. Plant Physiol. 2015, 169, 2030–2047. [Google Scholar] [CrossRef] [PubMed]
- Samach, A.; Lotan, H. The transition to flowering in tomato. Plant Biotechnol. 2007, 24, 71–82. [Google Scholar] [CrossRef] [Green Version]
- McCall, D. Effect of supplementary light on tomato transplant growth, and the after-effects on yield. Sci. Hortic. 1992, 51, 65–70. [Google Scholar] [CrossRef]
- Matsuda, R.; Yamano, T.; Murakami, K.; Fujiwara, K. Effects of spectral distribution and photosynthetic photon flux density for overnight LED light irradiation on tomato seedling growth and leaf injury. Sci. Hortic. 2016, 198, 363–369. [Google Scholar] [CrossRef]
- Fu, W.G.; Li, P.P.; Wu, Y.Y.; Tang, J. Effects of different light intensities on anti-oxidative enzyme activity, quality and biomass in lettuce. Sci. Hortic. 2012, 39, 129–134. [Google Scholar] [Green Version]
- Hole, C.C.; Dearman, J. The effect of photon flux density on distribution of assimilate between shoot and storage root of carrot, red beet and radish. Sci. Hortic. 1993, 55, 213–225. [Google Scholar] [CrossRef]
- Lee, S.H.; Tewari, R.K.; Hahn, E.J.; Paek, K.Y. Photon flux density and light quality induce changes in growth, stomatal development, photosynthesis and transpiration of Withania somnifera (L.) Dunal. plantlets. Plant Cell Tissue Organ Cult. 2007, 90, 141–151. [Google Scholar] [CrossRef]
- Seginer, I.; Albright, L.D.; Ioslovich, I. Improved strategy for a constant daily light integral in greenhouses. Biosyst. Eng. 2006, 93, 69–80. [Google Scholar] [CrossRef]
- Warner, R.M.; Erwin, J.E. Effect of photoperiod and daily light integral on flowering of five Hibiscus sp. Sci. Hortic. 2003, 97, 341–351. [Google Scholar] [CrossRef]
- Brown, C.S.; Schuerger, A.C.; Sager, J.C. Growth and photomorphogenesis of pepper plants under red light-emitting diodes with supplemental blue or far-red lighting. J. Am. Soc. Hortic. Sci. 1995, 120, 808–813. [Google Scholar] [CrossRef] [PubMed]
- Terashima, I.; Fujita, T.; Inoue, T.; Chow, W.S.; Oguchi, R. Green light drivesleaf photosynthesis more efficiently than red light in strong white light: Revisiting the enigmatic question of why leaves are green. Plant Cell Physiol. 2009, 50, 684–697. [Google Scholar] [CrossRef] [PubMed]
- Barnes, C.; Bugbee, B. Morphological responses of wheat to changes in phytochrome photoequilibrium. Plant Physiol. 1991, 97, 359–365. [Google Scholar] [CrossRef] [PubMed]
- Hogewoning, S.W.; Douwstra, P.; Trouwborst, G.; van Ieperen, W.; Harbinson, J. An artificial solar spectrum substantially alters plant development compared with usual climate room irradiance spectra. Exp. Bot. 2010, 61, 1267–1276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.Y.; Xu, X.M.; Cui, J. The importance of blue light for leaf area expansion, development of photosynthetic apparatus, and chloroplast ultrastructure of Cucumis sativus grown under weak light. Photosynthetica 2015, 53, 213–222. [Google Scholar] [CrossRef]
- Bach, A.; Krol, A. Effect of light quality on somatic embryogenesis in Hyacinthus orientalis L. ‘Delft’s Blue’. Biol. Bull. Pozn. 2001, 38, 103–107. [Google Scholar]
- Lander, G.C.; Estrin, E.; Matyskiela, M.E.; Bashore, C.; Nogales, E.; Martin, A. Complete subunit architecture of the proteasome regulatory particle. Nature 2012, 482, 186–191. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.; Zhang, S.B.; Liu, T. Moderate photoinhibition of photosystem II significantly affects linear electron flow in the shade-demanding plant Panax notoginseng. Front. Plant Sci. 2018, 9, 250–256. [Google Scholar] [CrossRef]
- Raymond, J.; Blankenship, R.E. The evolutionary development of the protein complement of photosystem II. Biochim. Biophys. Acta Bioenerg. 2004, 1655, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Grotjohann, I.; Jolley, C.; Fromme, P. Evolution of photosynthesis and oxygen evolution: Implications from the structural comparison of Photosystems I and II. Phys. Chem. Chem. Phys. 2004, 6, 4743–4753. [Google Scholar] [CrossRef]
- Trebitsh, T.; Danon, A. Translation of chloroplast psbA mRNA is regulated by signals initiated by both photosystems II and I. Proc. Natl. Acad. Sci. USA 2001, 98, 12289–12294. [Google Scholar] [CrossRef] [PubMed]
- Gruissem, W. Chloroplast gene expression: How plants turn their plastids on. Cell 1989, 56, 161–170. [Google Scholar] [CrossRef]
- Chitnis, P.R.; Xu, Q.; Chitnis, V.P.; Nechushtai, R. Function and organization of photosystem I polypeptides. Photosynth. Res. 1995, 44, 23–40. [Google Scholar] [CrossRef] [PubMed]
- Altman, A.; Cohen, B.N.; Weissbach, H.; Brot, N. Transcriptional activity of isolated maize chloroplasts. Arch. Biochem. Biophys. 1984, 235, 26–33. [Google Scholar] [CrossRef]
- Webber, A.N.; Malkin, R. Photosystem I reaction-centre proteins contain leucine zipper motifs: A proposed role in dimer formation. FEBS Lett. 1990, 264, 1–4. [Google Scholar] [CrossRef]
- Hiratsuka, J.; Shimada, H.; Whittier, R.; Ishibashi, T.; Sakamoto, M.; Mori, M.; Kondo, C.; Honji, Y.; Sun, C.R.; Meng, B.Y.; et al. The complete sequence of the rice (Oryza sativa) chloroplast genome: Intermolecular recombination between distinct tRNA genes accounts for a major plastid DNA inversion during the evolution of the cereals. Mol. Gen. Genet. 1989, 217, 185–194. [Google Scholar] [CrossRef]
- Chen, S.G.; Cheng, M.C.; Chung, K.R.; Yu, N.J.; Chen, M.C. Expression of the rice chloroplast psaA-psaB-rps14 gene cluster. Plant Sci. 1992, 81, 93–102. [Google Scholar] [CrossRef]
- Tsinoremas, N.F.; Schaefer, M.R.; Golden, S.S. Blue and red light reversibly control psbA expression in the cyanobacterium Synechococcus sp. strain PCC 7942. J. Biol. Chem. 1994, 269, 16143–16147. [Google Scholar]
- Albers, S.C.; Peebles, C.A. Evaluating light-induced promoters for the control of heterologous gene expression in Synechocystis sp. PCC 6803. Biotechnol. Prog. 2017, 33, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Jia, H.; Tian, Q.; Du, L.; Gao, Y.; Miao, X.; Liu, Y. Protecting effect of phosphorylation on oxidative damage of D1 protein by down-regulating the production of superoxide anion in photosystem II membranes under high light. Photosynth. Res. 2012, 112, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Antal, T.K.; Kovalenko, I.B.; Rubin, A.B.; Tyystjärvi, E. Photosynthesis-related quantities for education and modeling. Photosynth. Res. 2013, 117, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Kok, B. On the inhibition of photosynthesis by intense light. Biochim. Biophys. Acta. 1956, 21, 234–244. [Google Scholar] [CrossRef]
- Powles, S.B. Photoinhibition of photosynthesis induced by visible light. Annu. Rev. Plant Physiol. 1984, 35, 15–44. [Google Scholar] [CrossRef]
- Aro, E.M.; Virgin, I.; Andersson, B. Photoinhibition of photosystem II. Inactivation, protein damage and turnover. Biochim. Biophys. Acta 1993, 1143, 113–134. [Google Scholar] [CrossRef]
- Henmi, T.; Yamasaki, H.; Sakuma, S.; Tomokawa, Y.; Tamura, N.; Shen, J.R.; Yamamoto, Y. Dynamic Interaction between the D1 protein, CP43 and OEC33 at the lumenal side of photosystem II in spinach chloroplasts: Evidence from light-induced cross-Linking of the proteins in the donor-side photoinhibition. Plant Cell Physiol. 2003, 44, 451–456. [Google Scholar] [CrossRef]
- Terashima, I.; Funayama, S.; Sonoike, K. The site of photoinhibition in leaves of Cucumis sativus L. at low temperatures is photosystem I, not photosystem II. Planta 1994, 193, 300–306. [Google Scholar] [CrossRef]
- Huang, W.; Yang, Y.J.; Zhang, J.L.; Hu, H.; Zhang, S.B. PSI photoinhibition is more related to electron transfer from PSII to PSI rather than PSI redox state in Psychotria rubra. Photosynth. Res. 2016, 129, 85–92. [Google Scholar] [CrossRef]
- Gollan, P.J.; Lima-Melo, Y.; Tiwari, A.; Tikkanen, M.; Aro, E.M. Interaction between photosynthetic electron transport and chloroplast sinks triggers protection and signalling important for plant productivity. Philos. Trans. R. Soc. B 2017, 372, 20160390. [Google Scholar] [CrossRef] [Green Version]
- Tjus, S.E.; Moller, B.L.; Scheller, H.V. Photosystem I is an early target of photoinhibition in barley illuminated at chilling temperatures. Plant Physiol. 1998, 116, 755–764. [Google Scholar] [CrossRef] [PubMed]
- Suorsa, M.; Järvi, S.; Grieco, M.; Nurmi, M.; Pietrzykowska, M.; Rantala, M.; Kangasjärvi, S.; Paakkarinen, V.; Tikkanen, M.; Jansson, S.; et al. PROTON GRADIENT REGULATION5 is essential for proper acclimation of Arabidopsis photosystem I to naturally and artificially fluctuating light conditions. Plant Cell 2012, 24, 2934–2948. [Google Scholar] [CrossRef]
- Tikkanen, M.; Grebe, S. Switching off photoprotection of photosystem I—A novel tool for gradual PSI photoinhibition. Physiol. Plant. 2018, 162, 156–161. [Google Scholar] [CrossRef] [PubMed]
- Munekage, Y.; Hojo, M.; Meurer, J.; Endo, T.; Tasaka, M.; Shikanai, T. PGR5 is involved in cyclic electron flow around photosystem I and is essential for photoprotection in Arabidopsis. Cell 2002, 110, 361–371. [Google Scholar] [CrossRef]
- Tikkanen, M.; Mekala, N.R.; Aro, E.M. Photosystem II photoinhibition-repair cycle protects Photosystem I from irreversible damage. Biochim. Biophys. Acta 2014, 1837, 210–215. [Google Scholar] [CrossRef] [Green Version]
- Kagawa, T.; Sakai, T.; Suetsugu, N.; Oikawa, K.; Ishiguro, S.; Kato, T.; Tabata, S.; Okada, K.; Wada, M. Arabidopsis NPL1: A phototropin homolog controlling the chloroplast high-light avoidance response. Science 2001, 291, 2138–2141. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.M.; Osmond, C.B. Shade-sun responses: Compromises between acclimation and photoinhibition. In Photoinhibition; Kyle, D.J., Osmond, C.B., Arntzen, C.J., Eds.; Elsevier Science BV: Amsterdam, The Netherlands, 1987; Volume 96, pp. 2–38. [Google Scholar]
- Demmig-Adams, B.; Adams Iii, W.W. Photoprotection and other responses of plants to high light stress. Annu. Rev. Plant Biol. 1992, 43, 599–626. [Google Scholar] [CrossRef]
- Sacharz, J.; Giovagnetti, V.; Ungerer, P.; Mastroianni, G.; Ruban, A.V. The xanthophyll cycle affects reversible interactions between PsbS and light-harvesting complex II to control non-photochemical quenching. Nat. Plants 2017, 3, 16225. [Google Scholar] [CrossRef]
- Bräutigam, A.; Gowik, U. Photorespiration connects C3 and C4 photosynthesis. J. Exp. Bot. 2016, 67, 2953–2962. [Google Scholar] [CrossRef] [PubMed]
- Badger, M.R.; Caemmerer, S.; Ruuska, S.; Nakano, H. Electron flow to oxygen in higher plants and algae: Rates and control of direct photoreduction (Mehler reaction) and rubisco oxygenase. Philos. Trans. R. Soc. B 2000, 355, 1433–1446. [Google Scholar] [CrossRef]
- Guo, Y.Y.; Tian, S.S.; Liu, S.S.; Wang, W.Q.; Sui, N. Energy dissipation and antioxidant enzyme system protect photosystem II of sweet sorghum under drought stress. Photosynthetica 2018, 56, 861–872. [Google Scholar] [CrossRef]
- Li, J.; Cang, Z.; Jiao, F.; Bai, X.; Zhang, D.; Zhai, R. Influence of drought stress on photosynthetic characteristics and protective enzymes of potato at seedling stage. J. Saud. Soc. Agric. Sci. 2017, 16, 82–88. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, H.; Iwai, M.; Takahashi, Y.; Minagawa, J. Identification of the mobile light-harvesting complex II polypeptides for state transitions in Chlamydomonas reinhardtii. Proc. Natl. Acad. Sci. USA 2006, 103, 477–482. [Google Scholar] [CrossRef] [PubMed]
- Falkowski, P.G.; Chen, Y.B. Photoacclimation of light harvesting systems in eukaryotic algae. In Light Harvesting Antennas in Photosynthesis; Green, B.R., Parson, W.W., Eds.; Springer: Dordrecht, The Netherlands, 2003; Volume 13, pp. 423–447. [Google Scholar]
- Lichtenthaler, H.K.; Meier, D.; Buschmann, C. Development of chloroplasts at high and low light quanta fluence rates. Isr. J. Bot. 1984, 33, 185–194. [Google Scholar]
- Pastenes, C.; Horton, P. Effect of high temperature on pho to synthesis in beans. Plant Physiol. 1996, 11, 1253–1260. [Google Scholar] [CrossRef] [PubMed]
- Mathur, S.; Jain, L.; Jajoo, A. Photosynthetic efficiency in sun and shade plants. Photosynthetica 2018, 56, 354–365. [Google Scholar] [CrossRef]
- Pearcy, R.W.; Osteryoung, K.; Calkin, H.W. Photosynthetic responses to dynamic light environments by Hawaiian trees: Time course of CO2 uptake and carbon gain during sunflecks. Plant Physiol. 1985, 79, 896–902. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.B.; Sun, G.Y.; Wang, X.C. Induction characteristics and response of photosynthetic quantum conversion to changes in irradiance in mulberry plants. J. Plant Physiol. 2007, 164, 959–968. [Google Scholar] [CrossRef] [PubMed]
- Hussain, S.; Iqbal, N.; Rahman, T.; Ting, L.; Brestic, M.; Safdar, M.E.; Asghar, M.A.; Farooq, M.U.; Shafiq, I.; Ali, A.; et al. Shade effect on carbohydrates dynamics and stem strength of soybean genotypes. Environ. Exp. Bot. 2019, 162, 374–382. [Google Scholar] [CrossRef]
- Hussain, S.; Iqbal, N.; Brestic, M.; Raza, M.A.; Pang, T.; Langham, D.R.; Safdar, M.E.; Ahmed, S.; Wen, B.; Gao, Y.; et al. Changes in morphology, chlorophyll fluorescence performance and Rubisco activity of soybean in response to foliar application of ionic titanium under normal light and shade environment. Sci. Total Environ. 2019, 658, 626–637. [Google Scholar] [CrossRef]
- Huang, D.; Wu, L.; Chen, J.R.; Dong, L. Morphological plasticity, photosynthesis and chlorophyll fluorescence of Athyrium pachyphlebium at different shade levels. Photosynthetica 2011, 49, 611–618. [Google Scholar] [CrossRef]
- Yao, X.; Li, C.; Li, S.; Zhu, Q.; Zhang, H.; Wang, H.; Yu, C.; Martin, S.K.S.; Xie, F. Effect of shade on leaf photosynthetic capacity, light-intercepting, electron transfer and energy distribution of soybeans. In Plant Growth Regulation; Springer: Berlin/Heidelberg, Germany, 2017; Volume 83, pp. 1–8. [Google Scholar]
- Kunderlikova, K.; Brestic, M.; Zivcak, M.; Kusniarova, P. Photosynthetic responses of sun-and shade-grown chlorophyll b deficient mutant of wheat. J. Cent. Eur. Agric. 2016, 17, 950–956. [Google Scholar] [CrossRef]
- Valladares, F.; Niinemets, Ü. Shade tolerance, a key plant feature of complex nature and consequences. Annu. Rev. Ecol. Syst. 2008, 39, 237–257. [Google Scholar] [CrossRef]
- Portsmuth, A.; Niinemets, U. Structural and physiological plasticity in response to light and nutrients in five temperate deciduous woody species of contrasting shade tolerance. Funct. Ecol. 2007, 21, 61–77. [Google Scholar] [CrossRef]
- Valladares, F.; Wright, S.J.; Lasso, E.; Kitajima, K.; Pearcy, R.W. Plastic phenotypic response to light of 16 congeneric shrubs from a Panamanian rainforest. Ecology 2000, 81, 1925–1936. [Google Scholar] [CrossRef]
- Aspinwall, M.J.; Loik, M.E.; Resco de Dios, V.; Tjoelker, M.G.; Payton, P.R.; Tissue, D.T. Utilizing intraspecific variation in phenotypic plasticity to bolster agricultural and forest productivity under climate change. Plant Cell Environ. 2015, 38, 1752–1764. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar (C) | Light Intensity (LI) (μmol·m−2·s−1 PPFD) | Scion | Leaf | Root | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Length (cm) | Stem Diameter(mm) | No. | Length(cm) | Width (cm) | Area (cm2) | Thickness (mm) | Chlorophyll (SPAD) | Length (cm) | Fresh Weight (g) | ||
| ‘SS’ | 50 | 13.2 b z | 5.13 a | 7.8 b | 14.3 | 8.7 b | 61.9 b | 0.35 b | 44.1 b | 8.6 b | 0.4 b |
| 100 | 19.1 a | 5.31 a | 8.8 a | 15.0 | 9.0 ab | 73.6 ab | 0.39 b | 47.6 ab | 17.2 a | 1.1 ab | |
| 150 | 20.2 a | 5.69 a | 9.3 a | 15.8 | 9.9 a | 84.5 a | 0.47 a | 50.3 a | 18.6 a | 1.9 a | |
| ‘SD’ | 50 | 16.3 b | 5.52 a | 8.7 a | 15.3 | 8.8 a | 70.0 a | 0.36 a | 39.5 c | 9.9 c | 0.4 b |
| 100 | 20.1 a | 5.63 a | 9.2 a | 15.7 | 9.3 a | 69.9 a | 0.39 a | 48.2 a | 14.6 b | 1.0 a | |
| 150 | 19.5 a | 5.96 a | 9.8 a | 16.0 | 8.7 a | 68.1 a | 0.39 a | 42.6 ab | 16.9 a | 1.0 a | |
| F-test | |||||||||||
| LI | *** y | * | *** | NS | * | * | ** | * | *** | *** | |
| C | NS | * | * | NS | NS | NS | NS | NS | NS | NS | |
| LI * C | NS | NS | NS | NS | NS | ** | NS | NS | NS | NS | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, H.; Zhao, J.; Hu, J.; Jeong, B.R. Effect of Supplementary Light Intensity on Quality of Grafted Tomato Seedlings and Expression of Two Photosynthetic Genes and Proteins. Agronomy 2019, 9, 339. https://doi.org/10.3390/agronomy9060339
Wei H, Zhao J, Hu J, Jeong BR. Effect of Supplementary Light Intensity on Quality of Grafted Tomato Seedlings and Expression of Two Photosynthetic Genes and Proteins. Agronomy. 2019; 9(6):339. https://doi.org/10.3390/agronomy9060339
Chicago/Turabian StyleWei, Hao, Jin Zhao, Jiangtao Hu, and Byoung Ryong Jeong. 2019. "Effect of Supplementary Light Intensity on Quality of Grafted Tomato Seedlings and Expression of Two Photosynthetic Genes and Proteins" Agronomy 9, no. 6: 339. https://doi.org/10.3390/agronomy9060339
APA StyleWei, H., Zhao, J., Hu, J., & Jeong, B. R. (2019). Effect of Supplementary Light Intensity on Quality of Grafted Tomato Seedlings and Expression of Two Photosynthetic Genes and Proteins. Agronomy, 9(6), 339. https://doi.org/10.3390/agronomy9060339