Chromosomal Analysis in Crotophaga ani (Aves, Cuculiformes) Reveals Extensive Genomic Reorganization and an Unusual Z-Autosome Robertsonian Translocation

 ,
, 

 , ,
, ,  , and
, and 
Abstract
:1. Introduction
2. Material and Methods
2.1. Specimens and Chromosome Preparation
2.2. Diploid Number, Karyotype Description, C-Banding, and G-Banding
2.3. Fluorescent in Situ Hybridization (FISH)
2.4. Microscope Analysis and Image Capturing
3. Results
3.1. Diploid Number, Karyotype Description, C-Banding, and G-Banding
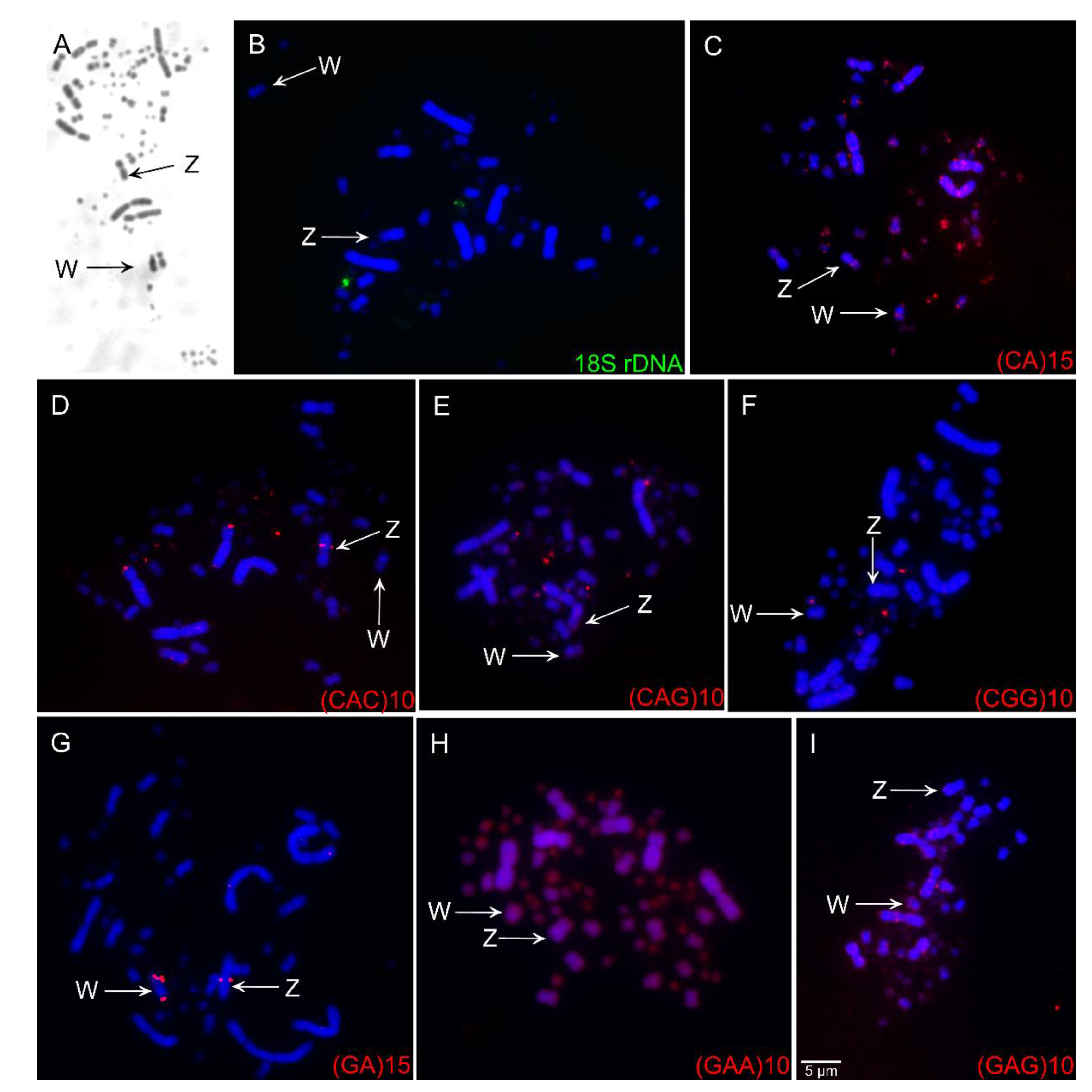
3.2. Fluorescence in Situ Hybridization (FISH) of Repetitive Sequences
3.3. Fluorescence in Situ Hybridization (FISH) of Chicken and Zebra Finch BAC Clones
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ellegren, H. Evolution of the avian sex chromosomes and their role in sex determination. Trends Ecol Evol. 2000, 15, 188–192. [Google Scholar] [CrossRef]
- Schartl, M.; Schmid, M.; Nanda, I. Dynamics of vertebrate sex chromosome evolution: From equal size to giants and dwarfs. Chromosoma 2016, 125, 553–571. [Google Scholar] [CrossRef] [PubMed]
- Kretschmer, R.; Ferguson-Smith, M.A.; de Oliveira, E.H.C. Karyotype Evolution in Birds: From Conventional Staining to Chromosome Painting. Genes 2018, 9, 181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nanda, I.; Schmid, M. Conservation of avian Z chromosomes as revealed by comparative mapping of the Z-linked aldolase B gene. Cytogenet. Genome Res. 2002, 96, 176–178. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, T.D.; Kretschmer, R.; Bertocchi, N.A.; Degrandi, T.M.; de Oliveira, E.H.C.; Cioffi, M.B.; Garnero, A.D.V.; Gunski, R.J. Genomic organization of repetitive DNA in woodpeckers (Aves, Piciformes): Implications for karyotype and ZW sex chromosome differentiation. PLoS ONE 2017, 12, e0169987. [Google Scholar] [CrossRef] [PubMed]
- Kretschmer, R.; Furo, I.O.; Cioffi, M.B.; Gunski, R.J.; Garnero, A.D.V.; O’Brien, P.C.M.; Ferguson-Smith, M.A.; de Freitas, T.R.O.; de Oliveira, E.H.C. Extensive chromosomal fissions and repetitive DNA accumulation shaped the atypical karyotypes of two Ramphastidae (Aves: Piciformes) species. Biol. J. Linn. Soc. Lond. 2020, 130, 839–849. [Google Scholar] [CrossRef]
- Nieto, L.M.; Kretschmer, R.; Ledesma, M.A.; Garnero, A.D.V.; Gunski, R.J. Karyotype morphology suggests that the Nyctibius griseus (Gmelin, 1789) carries an ancestral ZW-chromosome pair to the order Caprimulgiformes (Aves). Comp. Cytogenet. 2012, 6, 379–387. [Google Scholar] [CrossRef]
- Gunski, R.J.; Kretschmer, R.; de Souza, M.S.; Furo, I.O.; Barcellos, S.; Costa, A.L.; Cioffi, M.B.; de Oliveira, E.H.C.; Garnero, A.V. Evolution of bird sex chromosomes narrated by repetitive sequences: Unusual W chromosome enlargement in Gallinula melanops (Aves: Gruiformes: Rallidae). Cytogenet. Genome Res. 2019, 158, 152–159. [Google Scholar] [CrossRef]
- Furo, I.O.; Kretschmer, R.; dos Santos, M.S.; Carvalho, C.A.; Gunski, R.J.; O’Brien, P.C.M.; Ferguson-Smith, M.A.; Cioffi, M.B.; de Oliveira, E.H.C. Chromosomal Mapping of Repetitive DNAs in Myiopsitta monachus and Amazona aestiva (Psittaciformes, Psittacidae), with emphasis on the sex chromosomes. Cytogenet. Genome Res. 2017, 151, 151–160. [Google Scholar] [CrossRef]
- Vozdova, M.; Ruiz-Herrera, A.; Fernandez, J.; Cernohorska, H.; Frohlich, J.; Sebestova, H.; Kubickova, S.; Rubes, J. Meiotic behaviour of evolutionary sex-autosome translocations in Bovidae. Chromosome Res. 2016, 24, 325–338. [Google Scholar] [CrossRef]
- Araújo, N.P.; Stanyon, R.; Pereira, V.S.; Svartman, M. Interspecific Chromosome Painting Provides Clues to the Ancestral Karyotype of the New World Monkey Genus. Aotus. J. Mammal. Evol. 2019, 26, 283–290. [Google Scholar] [CrossRef]
- Veyrunes, F.; Catalan, J.; Sicard, B.; Robinson, T.J.; Duplantier, J.M.; Granjon, L.; Dobigny, G.; Britton-Davidian, J. Autosome and sex chromosome diversity among the African pygmy mice, subgenus Nannomys (Murinae; Mus). Chromosome Res. 2004, 12, 369–382. [Google Scholar] [CrossRef] [PubMed]
- Zartman, D.L. Location of the pea comb gene. Poultry Sci. 1973, 52, 1455–1462. [Google Scholar] [CrossRef] [PubMed]
- Telloni, R.V.; Jaap, R.G.; Fechheimer, N.S. Cytogenetic and phenotypic effects of a part of the Z-chromosome attached to a microchromosome in the chicken. Poultry Sci. 1976, 55, 1886–1896. [Google Scholar] [CrossRef] [PubMed]
- Pala, I.; Hasselquist, D.; Bensch, S.; Hansson, B. Patterns of molecular evolution of an avian neo-sex chromosome. Mol. Biol. Evol. 2012, 29, 3741–3754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pala, I.; Naurin, S.; Stervander, M.; Hasselquist, D.; Bensch, S.; Hansson, B. Evidence of a neo-sex chromosome in birds. Heredity 2012, 108, 264–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sigeman, H.; Ponnikas, S.; Chauhan, P.; Dierickx, E.; Brooke, M.d.L.; Hansson, B. Repeated sex chromosome evolution in vertebrates supported by expanded avian sex chromosomes. Proc. R. Soc. B 2019, 286, 20192051. [Google Scholar] [CrossRef]
- Sigeman, H.; Ponnikas, S.; Hansson, B. Whole-genome analysis across 10 songbird families within Sylvioidea reveals a novel autosome–sex chromosome fusion. Biol. Lett. 2020, 16, 20200082. [Google Scholar] [CrossRef] [Green Version]
- Gunski, R.J.; Cañedo, A.D.; Garnero, A.D.V.; Ledesma, M.A.; Coria, N.; Montalti, D.; Degrandi, T.M. Multiple sex chromosome system in penguins (Pygoscelis, Spheniscidae). Comp. Cytogenet. 2017, 11, 541–552. [Google Scholar] [CrossRef] [Green Version]
- Griffin, D.K.; Robertson, L.B.W.; Tempest, H.G.; Skinner, B.M. The evolution of the avian genome as revealed by comparative molecular cytogenetic. Cytogenet. Genome Res. 2007, 117, 64–77. [Google Scholar] [CrossRef]
- Kretschmer, R.; Furo, I.O.; Gomes, A.J.B.; Kiazim, L.G.; Gunski, R.J.; Garnero, A.D.V.; Pereira, J.C.; Ferguson-Smith, M.A.; de Oliveira, E.H.C.; Griffin, D.K.; et al. A Comprehensive Cytogenetic Analysis of Several Members of the Family Columbidae (Aves, Columbiformes). Genes 2020, 11, 632. [Google Scholar] [CrossRef] [PubMed]
- Furo, I.O.; Kretschmer, R.; O’Brien, P.C.M.; Pereira, J.; Garnero, A.D.V.; Gunski, R.J.; O’Connor, R.E.; Griffin, D.K.; Gomes, A.J.B.; Ferguson-Smith, M.A.; et al. Chromosomal evolution in the phylogenetic context in Neotropical Psittacidae with emphasis on a species with high karyotypic reorganization (Myiopsitta monachus). Front. Genet. 2020, 11, 721. [Google Scholar] [CrossRef]
- O’Connor, R.E.; Farré, M.; Joseph, S.; Damas, J.; Kiazim, L.; Jennings, R.; Bennett, S.; Slack, E.A.; Allanson, E.; Larkin, D.M.; et al. Chromosome-level assembly reveals extensive rearrangement in saker falcon and budgerigar, but not ostrich, genomes. Genome Biol. 2018, 19, 171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joseph, S.; O’Connor, R.E.; Al Mutery, A.F.; Watson, M.; Larkin, D.M.; Griffin, D.K. Chromosome Level Genome Assembly and Comparative Genomics between Three Falcon Species Reveals an Unusual Pattern of Genome Organisation. Diversity 2018, 10, 113. [Google Scholar] [CrossRef] [Green Version]
- O’Connor, R.E.; Kiazim, L.; Skinner, B.; Fonseka, G.; Joseph, S.; Jennings, R.; Larkin, D.M.; Griffin, D.K. Patterns of microchromosome organization remain highly conserved throughout avian evolution. Chromosoma 2019, 128, 21–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gill, F.; Donsker, D.; Rasmussen, P. IOC World Bird List (v10.2). 2020. Available online: https://www.worldbirdnames.org/new/ (accessed on 21 December 2020).
- Sorenson, M.D.; Payne, R.B. A molecular genetic analysis of cuckoo phylogeny. In The Cuckoos; Payne, R.B., Ed.; Oxford University Press: Oxford, UK, 2005; pp. 68–94. [Google Scholar]
- Waldrigues, A.; Ferrari, I. Karyotypic study of Cuculiform Birds. I. Karyotype of the Smooth-Billed Ani (Crotophaga ani). Rev. Brasil. Genet. 1982, 1, 121–129. [Google Scholar]
- Waldrigues, A.; Ferrari, I.; Neto, A.F. Estudo cariotípico em duas espécies de Cuculiformes Americanos (Aves). Acta Amaz. 1983, 13, 37–50. [Google Scholar] [CrossRef] [Green Version]
- dos Santos, M.S.; Kretschmer, R.; Furo, I.O.; Gunski, R.J.; Garnero, A.V.; Valeri, M.P.; O’Brien, P.C.M.; Ferguson-Smith, M.A.; de Oliveira, E.H.C. Chromosomal evolution and phylogenetic considerations in cuckoos (Aves, Cuculiformes, Cuculidae). PLoS ONE 2020, 15, e0232509. [Google Scholar] [CrossRef]
- Sasaki, M.; Ikeuchi, T.; Makino, S. A feather pulp culture technique for avian chromosomes, with notes on the chromosomes of the peafowl and the ostrich. Experientia 1968, 24, 1292–1293. [Google Scholar] [CrossRef]
- Guerra, M.S. Reviewing the chromosome nomenclature of Levan et al. Rev. Bras. Genet. 1986, 9, 741–743. [Google Scholar]
- Sumner, A.T. A simple technique for demonstrating centromeric heterochromatin. Exp. Cell Res. 1972, 75, 304–306. [Google Scholar] [CrossRef]
- Kubat, Z.; Hobza, R.; Vyskot, B.; Kejnovsky, E. Microsatellite accumulation on the Y chromosome of Silene latifolia. Genome 2008, 51, 350–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cioffi, M.B.; Martins, C.; Centofante, L.; Jacobina, U.; Bertollo, L.A.C. Chromosomal variability among allopatric populations of Erythrinidae fish Hoplias malabaricus: Mapping of three classes of repetitive DNAs. Cytogenet. Genome Res. 2009, 125, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Degrandi, T.M.; Gunski, R.J.; Garnero, A.D.V.; de Oliveira, E.H.C.; Kretschmer, R.; de Souza, M.S.; Barcellos, S.A.; Hass, I. The distribution of 45S rDNA sites in bird chromosomes suggests multiple evolutionary histories. Genet. Mol. Biol. 2020, 43, e20180331. [Google Scholar] [CrossRef] [PubMed]
- Nishida-Umehara, C.; Tsuda, Y.; Ishijima, J.; Ando, J.; Fujiwara, A.; Matsuda, Y.; Griffin, D.K. The molecular basis of chromosome orthologies and sex chromosomal differentiation in palaeognathous birds. Chromosome Res. 2007, 15, 721–734. [Google Scholar] [CrossRef] [PubMed]
- Kretschmer, R.; de Oliveira, T.D.; Furo, I.O.; Silva, F.A.O.; Gunski, R.J.; Garnero, A.V.; Cioffi, M.B.; de Oliveira, E.H.C.; de Freitas, T.R.O. Repetitive DNAs and shrink genomes: A chromosomal analysis in nine Columbidae species (Aves, Columbiformes). Genet. Mol. Biol. 2018, 41, 98–106. [Google Scholar] [CrossRef]
- Barcellos, S.; Kretschmer, R.; de Souza, M.S.; Costa, A.L.; Degrandi, T.M.; dos Santos, M.S.; de Oliveira, E.H.C.; Cioffi, M.B.; Gunski, R.J.; Garnero, A.V. Karyotype Evolution and Distinct Evolutionary History of the W Chromosomes in Swallows (Aves, Passeriformes). Cytogenet. Genome Res. 2019, 158, 98–105. [Google Scholar] [CrossRef]
- Deakin, J.E.; Edwards, M.J.; Patel, H.; O’Meally, D.; Lian, J.; Stenhouse, R.; Ryan, S.; Livernois, A.M.; Azad, B.; Holleley, C.E.; et al. Anchoring genome sequence to chromosomes of the central bearded dragon (Pogona vitticeps) enables reconstruction of ancestral squamate macrochromosomes and identifies sequence content of the Z chromosome. BMC Genom. 2016, 17, 447. [Google Scholar] [CrossRef] [Green Version]
- Searle, A.G.; Beechey, C.V.; Evans, E.P.; Kirk, M. Two new X autosome translocations in the mouse. Cytogenet Cell Genet. 1983, 35, 279–292. [Google Scholar] [CrossRef]
- Tease, C.; Fisher, G. Two new X-autosome Robertsonian translocations in the mouse. I. Meiotic chromosome segregation in male hemizygotes and female heterozygotes. Genet. Res. 1991, 58, 115–121. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.-W.; Park, S.-Y.; Ryu, H.-M.; Lee, D.-E.; Lee, B.-Y.; Kim, S.-Y.; Park, Y.-S.; Lee, H.-S.; Seo, J.-T. Molecular and clinical characteristics of 26 cases with structural Y chromosome aberrations. Cytogenet. Genome Res. 2012, 136, 270–277. [Google Scholar] [CrossRef] [PubMed]
- Ratomponirina, C.; Viegas-Péquignot, E.; Dutrillaux, B.; Petter, F.; Rumpler, Y. Synaptonemal complexes in Gerbillidae: Probable role of intercalated heterochromatin in gonosomeautosome translocations. Cytogenet. Cell Genet. 1986, 43, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Dobigny, G.; Ozouf-Costaz, C.; Bonillo, C.; Volobouev, V. Viability of X-autosome translocations in mammals: An epigenomic hypothesis from a rodent case-study. Chromosoma 2004, 113, 34–41. [Google Scholar] [CrossRef]
- Matveevsky, S.N.; Pavlova, S.V.; Atsaeva, M.M.; Searle, J.B.; Kolomiets, O.L. Dual mechanism of chromatin remodeling in the common shrew sex trivalent (XY1Y2). Comp. Cytogenet. 2017, 11, 727–745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellegren, H.; Hultin-Rosenberg, L.; Brunstrom, B.; Dencker, L.; Kultima, K.; Scholz, B. Faced with inequality: Chicken do not have a general dosage compensation of sex-linked genes. BMC Biol. 2007, 5, 40. [Google Scholar] [CrossRef] [Green Version]
- Ellegren, H. Characteristics, causes and evolutionary consequences of male-biased mutation. Proc. R Soc. B Biol Sci. 2007, 274, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Itoh, Y.; Replogle, K.; Kim, Y.H.; Wade, J.; Clayton, D.F.; Arnold, A.P. Sex bias and dosage compensation in the zebra finch versus chicken genomes: General and specialized patterns among birds. Genome Res. 2010, 20, 512–518. [Google Scholar] [CrossRef] [Green Version]
- Itoh, Y.; Melamed, E.; Yang, X.; Kampf, K.; Wang, S.; Yehya, N.; Nas, A.V.; Replogle, K.; Band, M.R.; Clayton, D.F.; et al. Dosage compensation is less effective in birds than in mammals. J. Biol. 2007, 6, 2. [Google Scholar] [CrossRef] [Green Version]
- Wolf, J.B.; Bryk, J. General lack of global dosage compensation in ZZ/ZW systems? Broadening the perspective with RNA-seq. Bmc Genom. 2011, 12, 91. [Google Scholar] [CrossRef] [Green Version]
- Julien, P.; Brawand, D.; Soumillon, M.; Necsulea, A.; Liechti, A.; Schütz, F.; Daish, T.; Grützner, F.; Kaessmann, H. Mechanisms and evolutionary patterns of mammalian and avian dosage compensation. PLoS Biol. 2012, 10, e1001328. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chicken Chromosomes | Equivalent Guira Cuckoo, Guira Guira [30] | Equivalent Squirrel Cuckoo, Piaya Cayana [30] | Equivalent Smooth Billed Ani, Crotophaga Ani (Present Study) |
|---|---|---|---|
| 1 | 1 | 1 and 6 | 1 |
| 2 | 2 | 2, 13 and 15 | 2 |
| 3 | 3 | 5 and 10 | 3 |
| 4q | 4p and 6 | 3 | 6p, 7 and 12p |
| 4p | 9p | 14 | 9p |
| 5 | 5q, 7p and 8p | 4 | 4p, 5p and 14 |
| 6 | 8q and 12q | 8 | 4q and 11q |
| 7 | 4q and 10 | 7 | 6q |
| 8 | 9q | 9 | 8q |
| 9 | 11 | 11 | 13 |
| 10 | 5p | 12 | 10q |
| 11 | - | - | 8p |
| 12 | - | - | 9q |
| 13 | - | - | 12q |
| 14 | - | - | 11p |
| 15 | - | - | 5q |
| 16 | - | - | No data |
| 17 | - | - | Z |
| 18 | - | - | 17 |
| 19 | - | - | 18 |
| 20 | - | - | 19 |
| 21 | - | - | 20 |
| 22 | - | - | 21 |
| 23 | - | - | 22 |
| 24 | - | - | 23 |
| 25 | - | - | 10p |
| 26 | - | - | 24 |
| 27 | - | - | 25 |
| 28 | - | - | 26 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kretschmer, R.; Gunski, R.J.; Garnero, A.d.V.; de Freitas, T.R.O.; Toma, G.A.; Cioffi, M.d.B.; Oliveira, E.H.C.d.; O’Connor, R.E.; Griffin, D.K. Chromosomal Analysis in Crotophaga ani (Aves, Cuculiformes) Reveals Extensive Genomic Reorganization and an Unusual Z-Autosome Robertsonian Translocation. Cells 2021, 10, 4. https://doi.org/10.3390/cells10010004
Kretschmer R, Gunski RJ, Garnero AdV, de Freitas TRO, Toma GA, Cioffi MdB, Oliveira EHCd, O’Connor RE, Griffin DK. Chromosomal Analysis in Crotophaga ani (Aves, Cuculiformes) Reveals Extensive Genomic Reorganization and an Unusual Z-Autosome Robertsonian Translocation. Cells. 2021; 10(1):4. https://doi.org/10.3390/cells10010004
Chicago/Turabian StyleKretschmer, Rafael, Ricardo José Gunski, Analía del Valle Garnero, Thales Renato Ochotorena de Freitas, Gustavo Akira Toma, Marcelo de Bello Cioffi, Edivaldo Herculano Corrêa de Oliveira, Rebecca E. O’Connor, and Darren K. Griffin. 2021. "Chromosomal Analysis in Crotophaga ani (Aves, Cuculiformes) Reveals Extensive Genomic Reorganization and an Unusual Z-Autosome Robertsonian Translocation" Cells 10, no. 1: 4. https://doi.org/10.3390/cells10010004
APA StyleKretschmer, R., Gunski, R. J., Garnero, A. d. V., de Freitas, T. R. O., Toma, G. A., Cioffi, M. d. B., Oliveira, E. H. C. d., O’Connor, R. E., & Griffin, D. K. (2021). Chromosomal Analysis in Crotophaga ani (Aves, Cuculiformes) Reveals Extensive Genomic Reorganization and an Unusual Z-Autosome Robertsonian Translocation. Cells, 10(1), 4. https://doi.org/10.3390/cells10010004