
In Silico Model Estimates the Clinical Trial Outcome of Cancer Vaccines

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Studies Included in the Meta-Analysis
2.2. In Silico Trial
2.3. Statistical Calculations
3. Results
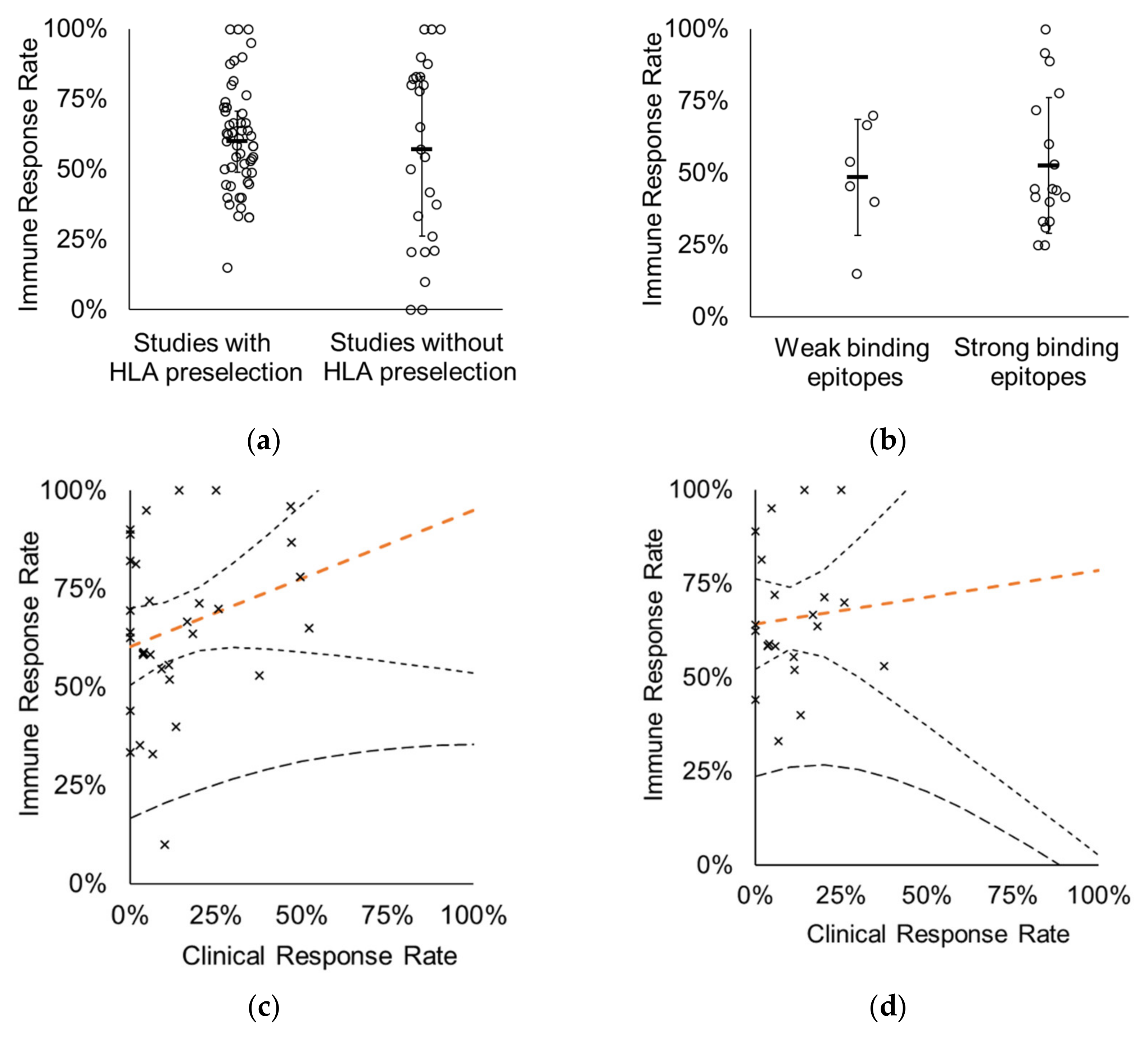
3.1. Preselection of HLA-Matched Subjects Does Not Improve Response Rate Obtained in Clinical Trials
3.2. Characterization of the In Silico Model
3.2.1. HLA Allele Frequency Analysis
3.2.2. Epitope-Binding Capabilities
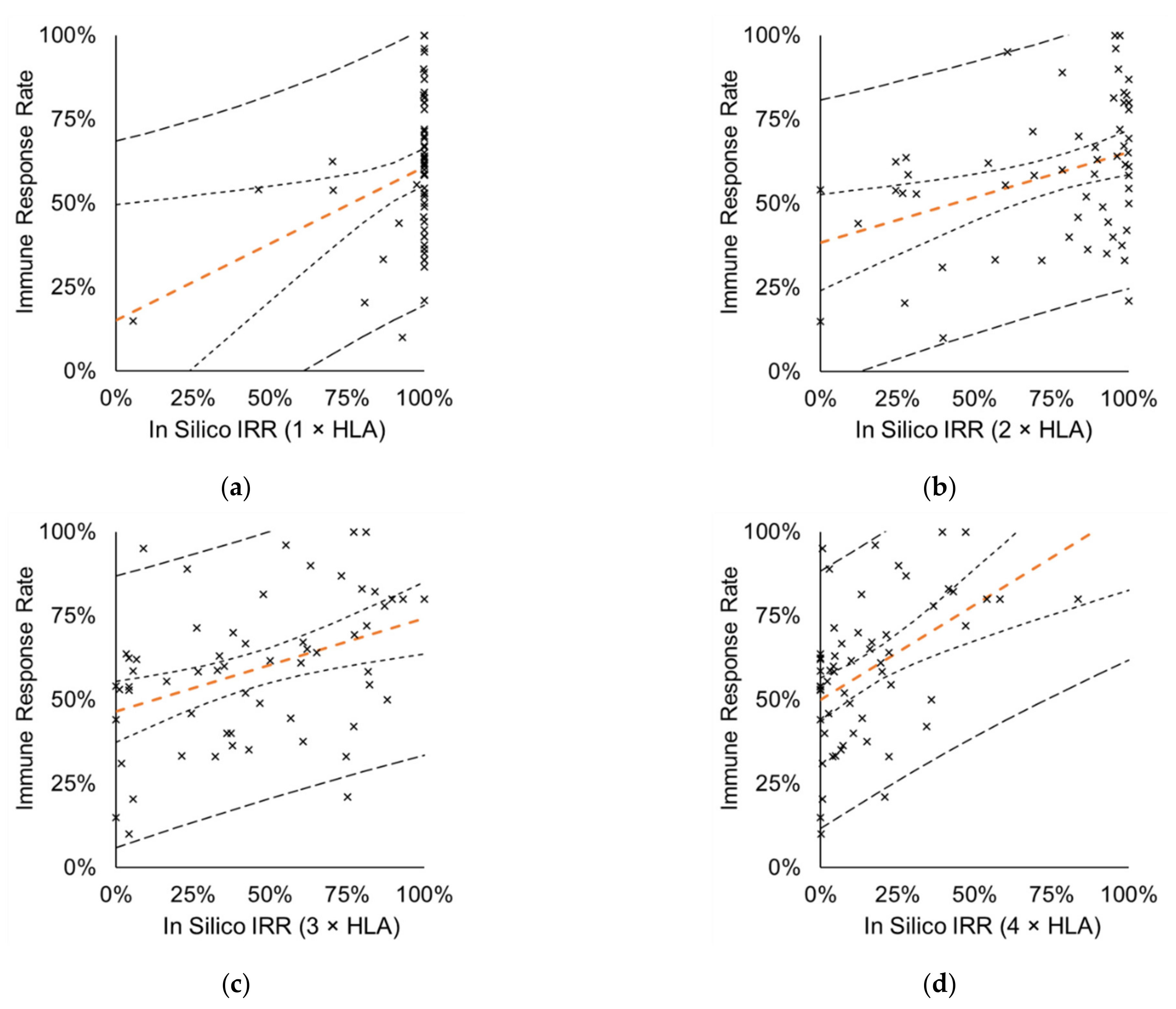
3.3. In Silico IRRs in the MP Correlate with IRRs Measured in CTs
3.3.1. Correlation Analysis between In Silico and Measured IRRs
3.3.2. Pairwise Comparison of In Silico and Measured IRRs
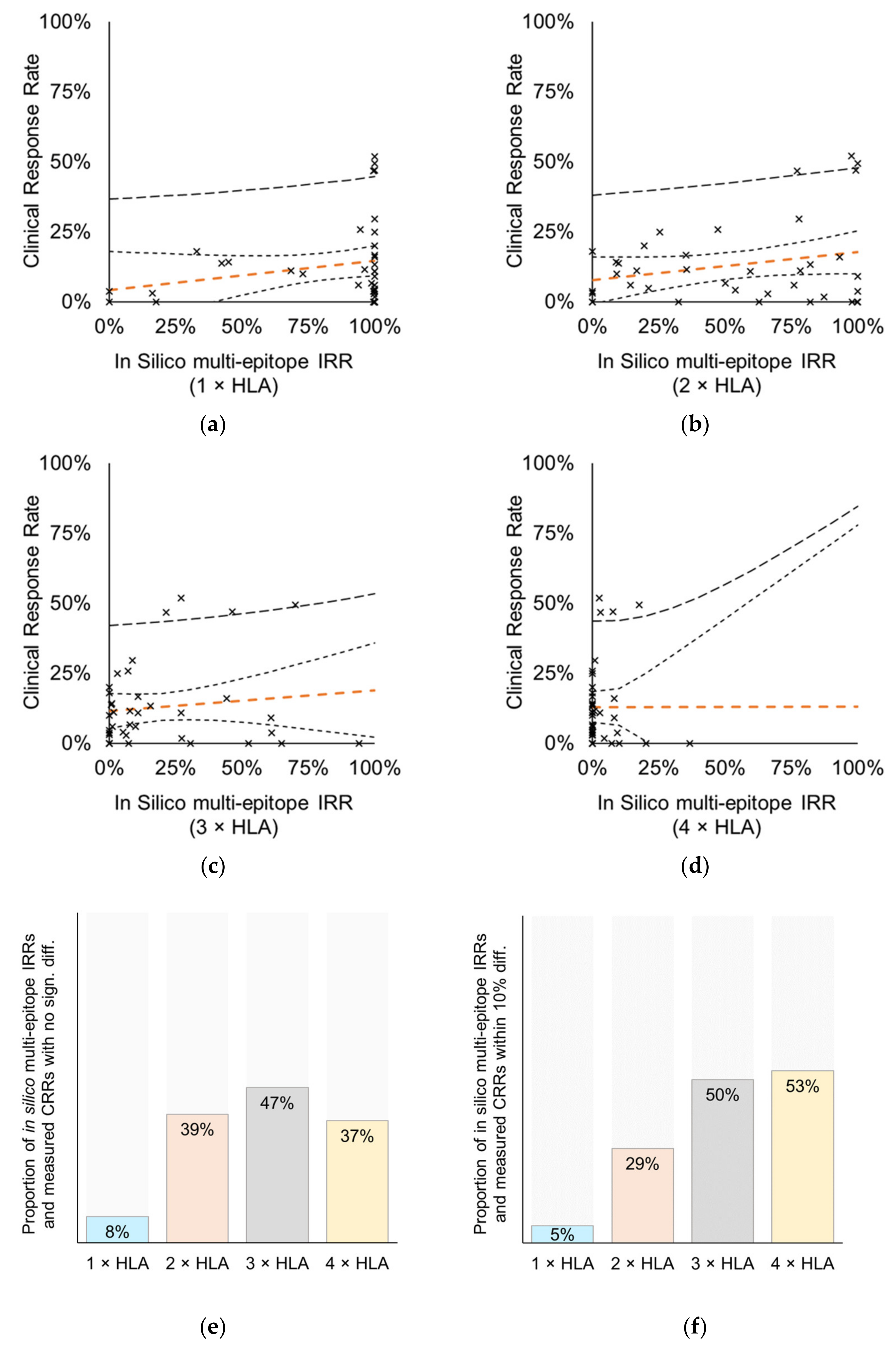
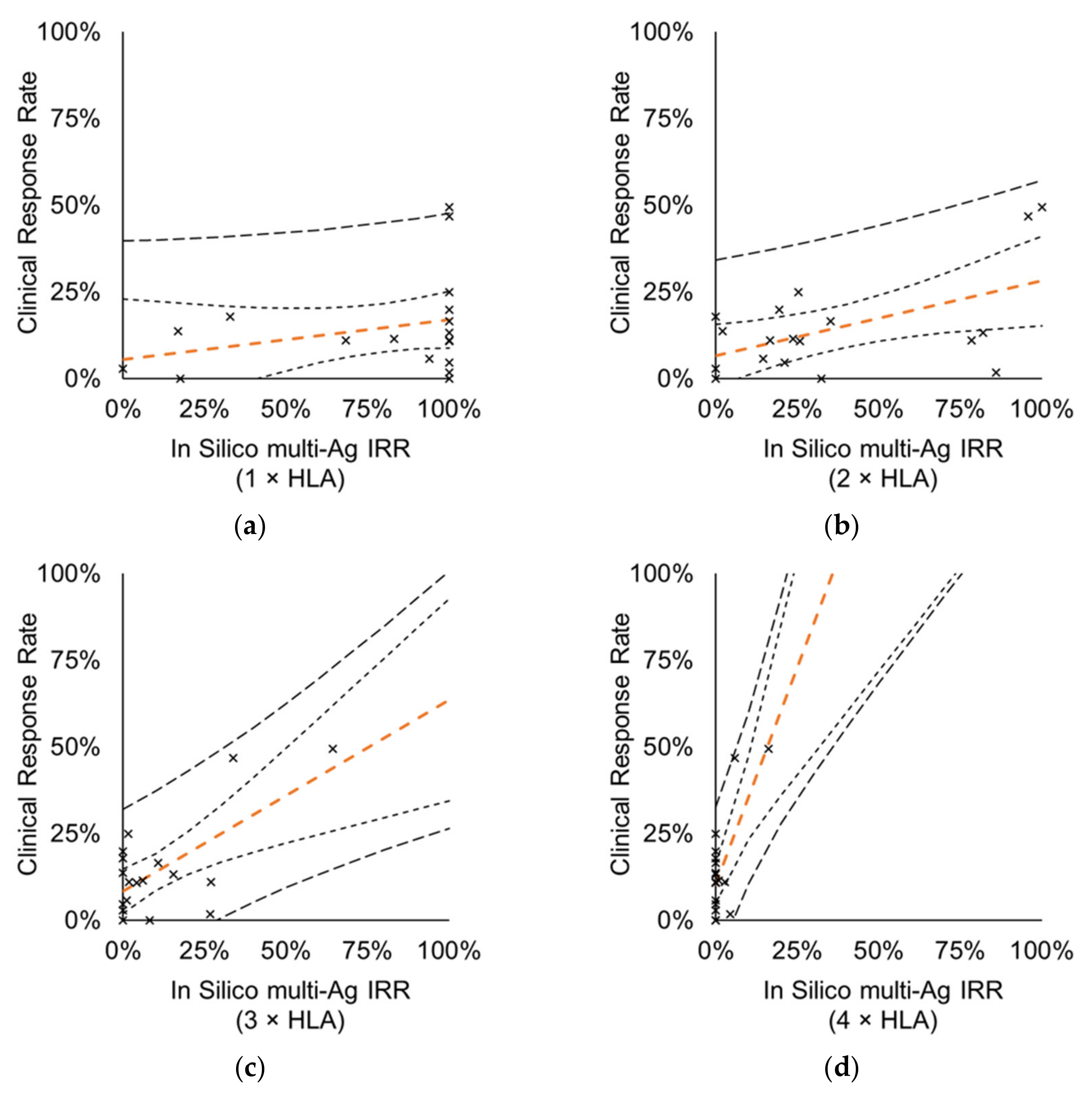
3.4. Relationship between Immune- and Clinical Response
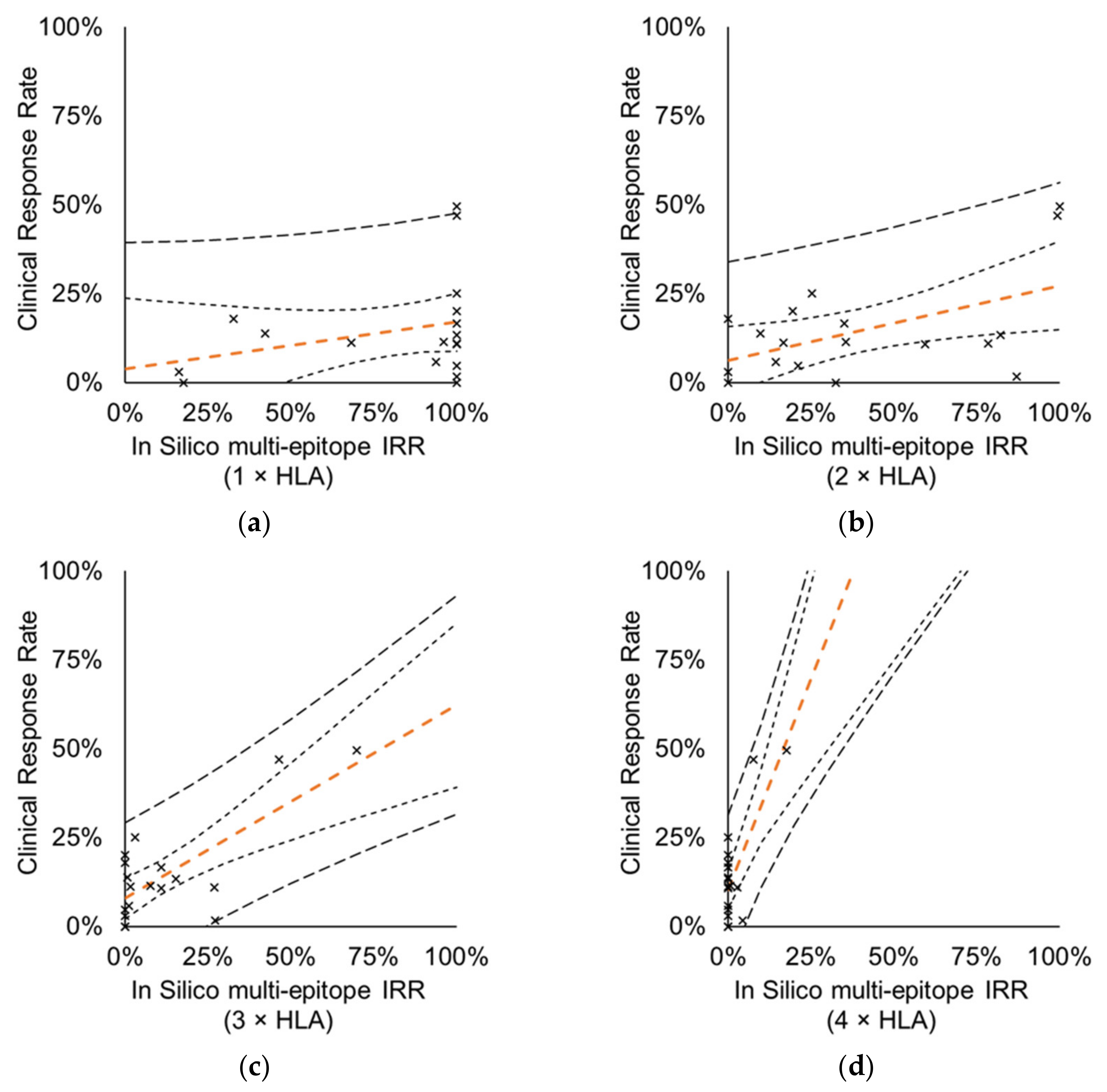
3.4.1. Vaccines Targeting Multiple Epitopes
3.4.2. Vaccines Targeting Multiple Protein Antigens
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vaccine | IRR | CRR | Combined IRR | Combined CRR | Ref. | Included in Analysis (X) | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| IRR-CRR Correlations | IRR in CTs with/without HLA Preselection | High Binding, Low Binding Epitope Vaccines | Measured and In Silico IRR Correlations | Measured CRR-In Silico IRR Correlations | MultiAg Correlations | ||||||
| 9-peptide breast cancer vaccine | 36% | --- | 36% | --- | [37] | X | X | ||||
| AFP-derived peptides | 33% | 7% | 33% | 7% | [38] | X | X | X | X | ||
| Antigen-pulsed DC vaccine | 38% | --- | 38% | --- | [39] | X | X | ||||
| CD34 DC vaccine | 44% | --- | 44% | --- | [40] | X | X | ||||
| CV9103 mRNA vaccine | 80% | --- | 80% | --- | [41,42] | X | X | ||||
| DCCP peptide vaccine | 56% | 11% | 56% | 11% | [23] | X | X | X | X | X | |
| DPX0907 peptide vaccine | 61% | --- | 61% | --- | [43,44] | X | X | ||||
| Elenagen pDNA vaccine | --- | 0% | --- | 0% | [45] | X | |||||
| EMD640744 peptide vaccine | 63% | --- | 63% | --- | [22] | X | X | ||||
| Five-peptide cancer vaccine | --- | 11% | --- | 11% | [46] | X | X | ||||
| GAA peptides vaccine | 100% | 29% | 71% | 20% | [47] | X | X | X | X | X | X |
| 55% | 15% | [48] | |||||||||
| GL-0817 (MAGE-A3 Trojan) | 0% | 0% | 33% | 0% | [49] | X | X | X | X | ||
| 33% | 0% | [50] | X | ||||||||
| 57% | --- | [51] | X | ||||||||
| Glypican-3 peptide vaccine (A24) | 33% | 33% | 64% | 18% | [52,53] | X | X | X | X | ||
| 100% | 0% | X | |||||||||
| Glypican-3 peptide vaccine (A02) | 67% | 0% | 53% | 38% | X | X | X | X | |||
| 45% | 60% | X | |||||||||
| GVX301 peptide vaccine | 64% | 0% | 64% | 0% | [54] | X | X | X | X | ||
| HER1 vaccine | 42% | 42% | --- | [55] | X | X | |||||
| HER2 B cell vaccine | --- | 6% | [56] | X | |||||||
| Her2/neu peptide vaccine (p369) | 62% | 62% | --- | [57] | X | X | |||||
| Her2/neu peptide vaccine (p688) | 31% | --- | X | X | |||||||
| Her2/neu peptide vaccine (p971) | 54% | --- | X | X | |||||||
| HIVIS (HIV gag) | 80% | 80% | --- | [58] | X | X | |||||
| HIVIS (HIV RT) | 50% | 50% | --- | [58] | X | X | |||||
| HPV-SLP | 83% | 60% | 87% | 47% | [59] | X | X | X | X | X | |
| 100% | 60% | [60] | X | ||||||||
| --- | 33% | [61] | |||||||||
| ICT107 | 33% | 33% | --- | [62] | X | X | |||||
| IDO and survivin peptide vaccine | 58% | 6% | 58% | 6% | [63] | X | X | X | X | X | X |
| IDOlong peptide vaccine | 10% | 10% | 10% | 10% | [64] | X | X | X | X | ||
| IMA901 | 64% | --- | 67% | --- | [5] | X | X | ||||
| 74% | --- | X | |||||||||
| IMA950 | 63% | 0% | 81% | 2% | [65] | X | X | X | X | X | |
| 81% | 3% | [66] | X | ||||||||
| 90% | --- | [67] | X | ||||||||
| ImMucin | 100% | 47% | 96% | 47% | [68,69] | X | X | X | X | ||
| IMP321/LAG-3Ig + peptides vaccine | 40% | --- | 40% | --- | [70] | X | X | X | |||
| INGN-225 p53 vaccine | 58% | 4% | 58% | 4% | [71] | X | X | X | X | ||
| KIF20A-66 peptide vaccine | 70% | 26% | 70% | 26% | [72] | X | X | X | X | X | |
| KRM-20 vaccine | 40% | 13% | 40% | 13% | [73] | X | X | X | X | X | |
| MART-1 Peptide Vaccine | 15% | --- | 15% | --- | [74] | X | X | X | |||
| Melanoma peptide vaccine | 52% | 12% | 52% | 12% | [75] | X | X | X | X | X | |
| MELITAC 12.1 | 49% | --- | 49% | --- | [24] | X | X | ||||
| Multiepitope peptide vaccine | 89% | 0% | 89% | 0% | [76] | X | X | X | X | X | X |
| NY-ESO-1 OLP | 82% | --- | 82% | 0% | [77] | X | X | X | |||
| [78] | |||||||||||
| NY-ESO-1f | 90% | 0% | 90% | 0% | [79] | X | X | X | X | ||
| OCV-C02 vaccine | 63% | 0% | 63% | 0% | [80] | X | X | X | X | X | |
| p53 SLP70-235 | 21% | --- | 21% | --- | [81] | X | X | ||||
| p53 SLP70-248 | 0% | --- | 69% | [82] | X | X | |||||
| 88% | 0% | 0% | [83] | X | X | X | |||||
| 100% | 0% | [84] | X | ||||||||
| p53MVA vaccine | 55% | 9% | 55% | 9% | [85] | X | X | X | X | ||
| PepCan | 65% | 52% | 65% | 52% | [86] | X | X | X | X | ||
| Peptide cocktail therapy | --- | 11% | --- | 11% | [87] | X | X | ||||
| --- | 14% | --- | 14% | X | X | ||||||
| Peptide vaccine 1 | 54% | --- | 54% | --- | [88] | X | X | ||||
| Peptide vaccine 2 | --- | 3% | --- | 3% | [89] | X | X | ||||
| Peptide vaccine 3 | 100% | 25% | 100% | 25% | [90] | X | X | X | X | X | |
| pNGVL4a-CRT/E7(detox) vaccine | --- | 30% | --- | 30% | [91] | X | |||||
| PR1 Peptide Vaccine | 53% | 53% | --- | [21] | X | X | X | ||||
| ProstVac | 45% | --- | 62% | --- | [97] | X | X | ||||
| 50% | --- | [93] | X | ||||||||
| 67% | --- | [92] | X | ||||||||
| 67% | --- | [94] | X | ||||||||
| 72% | --- | [96] | X | ||||||||
| 76% | --- | [95] | X | ||||||||
| PSMA-Survivin pulsed autologous DC vaccine | --- | 18% | --- | 18% | [98] | X | X | ||||
| PVX-410 peptide vaccine | 95% | 5% | 95% | 5% | [99] | X | X | X | X | X | |
| RHAMM-R3 | 44% | 0% | 44% | 0% | [100] | X | X | X | X | ||
| S-288310 peptide vaccine | 67% | 17% | 67% | 17% | [101] | X | X | X | X | X | |
| StimuVax | 21% | 21% | --- | [102] | X | X | |||||
| SVN-2B peptide vaccine | 60% | 60% | --- | [103] | X | X | X | ||||
| Synchrotope TA2M | 46% | --- | 46% | --- | [104] | X | |||||
| TARP vaccine | 80% | 80% | --- | [105] | X | X | |||||
| TG4010 | --- | 0% | 35% | 3% | [106] | X | X | X | X | ||
| 21% | --- | [107] | X | ||||||||
| 26% | 0% | [108] | X | ||||||||
| 38% | 13% | [109] | X | ||||||||
| TSPP peptide vaccine | --- | 5% | --- | 16% | [110] | X | |||||
| --- | 24% | [111] | |||||||||
| VGX-3100 | 78% | --- | 78% | 50% | [112] | X | X | X | X | X | |
| --- | 50% | [113] | |||||||||
| Vx-001 | 51% | --- | 59% | [114] | X | X | |||||
| 58% | 4% | 4% | [115] | X | X | X | |||||
| 66% | 7% | [117] | X | ||||||||
| 71% | 0% | [116] | X | ||||||||
| WT1 vaccine (1) | 64% | 0% | 59% | 4% | [118] | X | X | X | X | ||
| 40% | 10% | [119] | X | X | |||||||
| 86% | 0% | [120] | X | X | |||||||
| WT1 peptide vaccine | 72% | 6% | 72% | 6% | [121] | X | X | X | X | ||
| WT1 peptides | 100% | 14% | 100% | 14% | [122] | X | X | X | X | ||
| WT1 vaccine (2) | 83% | --- | 83% | --- | [123] | X | X | ||||
| HLA Allele | Frequency in CIWD |
|---|---|
| HLA-A*02:01 | 0.24065 |
| HLA-A*01:01 | 0.13343 |
| HLA-C*07:01 | 0.11948 |
| HLA-A*03:01 | 0.11852 |
| HLA-C*07:02 | 0.11093 |
| HLA-C*04:01 | 0.11062 |
| HLA-B*07:02 | 0.10205 |
| HLA-A*24:02 | 0.08948 |
| HLA-C*06:02 | 0.08771 |
| HLA-B*08:01 | 0.08462 |
| HLA-B*44:02 | 0.06295 |
| HLA-A*11:01 | 0.06098 |
| HLA-C*03:04 | 0.06076 |
| HLA-C*05:01 | 0.05597 |
| HLA-C*12:03 | 0.05491 |
| HLA-B*51:01 | 0.05432 |
| HLA-B*35:01 | 0.05270 |
| HLA-B*15:01 | 0.05000 |
| HLA-B*18:01 | 0.04534 |
| HLA-C*02:02 | 0.04281 |
| HLA-B*44:03 | 0.04280 |
| HLA-C*03:03 | 0.04237 |
| HLA-B*40:01 | 0.03957 |
| HLA-C*01:02 | 0.03512 |
| HLA-A*26:01 | 0.03352 |
| HLA-A*68:01 | 0.03338 |
| HLA-A*32:01 | 0.03200 |
| HLA-B*57:01 | 0.03134 |
| HLA-B*27:05 | 0.02952 |
| HLA-B*13:02 | 0.02899 |
| HLA-C*15:02 | 0.02573 |
| HLA-C*08:02 | 0.02483 |
| HLA-A*31:01 | 0.02462 |
| HLA-B*35:03 | 0.02344 |
| HLA-C*16:01 | 0.02241 |
| HLA-B*38:01 | 0.02218 |
| HLA-A*29:02 | 0.02166 |
| HLA-A*25:01 | 0.02124 |
| HLA-A*23:01 | 0.02096 |
| HLA-B*14:02 | 0.02042 |
| HLA-C*07:04 | 0.01709 |
| HLA-B*52:01 | 0.01659 |
| HLA-A*30:01 | 0.01589 |
| HLA-B*40:02 | 0.01534 |
| HLA-C*12:02 | 0.01526 |
| HLA-B*49:01 | 0.01498 |
| HLA-B*55:01 | 0.01438 |
| HLA-C*14:02 | 0.01383 |
| HLA-A*33:03 | 0.01360 |
| HLA-B*37:01 | 0.01334 |
| HLA-C*17:01 | 0.01273 |
| HLA-B*58:01 | 0.01224 |
| HLA-B*50:01 | 0.01189 |
| HLA-B*39:01 | 0.01158 |
| HLA-B*35:02 | 0.01027 |
| HLA-A*02:05 | 0.00878 |
| HLA-A*30:02 | 0.00838 |
| HLA-A*68:02 | 0.00834 |
| HLA-B*56:01 | 0.00784 |
| HLA-C*03:02 | 0.00779 |
| HLA-A*33:01 | 0.00749 |
| HLA-B*40:06 | 0.00719 |
| HLA-B*53:01 | 0.00696 |
| HLA-B*45:01 | 0.00604 |
| HLA-B*07:05 | 0.00593 |
| HLA-B*41:01 | 0.00592 |
| HLA-B*41:02 | 0.00579 |
| HLA-B*14:01 | 0.00578 |
| HLA-A*02:06 | 0.00569 |
| HLA-C*15:05 | 0.00537 |
| HLA-C*08:01 | 0.00515 |
| HLA-B*39:06 | 0.00514 |
| HLA-A*02:11 | 0.00463 |
| HLA-A*66:01 | 0.00463 |
| HLA-A*29:01 | 0.00413 |
| HLA-A*03:02 | 0.00368 |
| HLA-B*15:03 | 0.00337 |
| HLA-B*15:18 | 0.00281 |
| HLA-C*02:10 | 0.00275 |
| HLA-B*15:02 | 0.00261 |
| HLA-A*02:02 | 0.00239 |
| HLA-A*24:03 | 0.00236 |
| HLA-B*48:01 | 0.00229 |
| HLA-B*13:01 | 0.00219 |
| HLA-B*46:01 | 0.00215 |
| HLA-C*16:04 | 0.00208 |
| HLA-A*74:01 | 0.00200 |
| HLA-B*57:03 | 0.00193 |
| HLA-B*42:01 | 0.00193 |
| HLA-A*02:07 | 0.00183 |
| HLA-A*30:04 | 0.00183 |
| HLA-A*02:03 | 0.00174 |
| HLA-B*38:02 | 0.00171 |
| HLA-A*34:02 | 0.00162 |
| HLA-B*39:05 | 0.00159 |
| HLA-B*15:10 | 0.00145 |
| HLA-C*18:01 | 0.00136 |
| HLA-B*58:02 | 0.00136 |
| HLA-A*36:01 | 0.00097 |
| HLA-B*15:16 | 0.00095 |
| HLA-A*68:03 | 0.00084 |
| HLA-B*54:01 | 0.00075 |
| HLA-C*08:03 | 0.00073 |
| HLA-B*35:17 | 0.00073 |
| HLA-B*81:01 | 0.00067 |
| HLA-C*14:03 | 0.00067 |
| HLA-A*11:02 | 0.00056 |
| HLA-B*39:24 | 0.00052 |
| HLA-B*15:07 | 0.00049 |
| HLA-A*80:01 | 0.00045 |
| HLA-B*57:02 | 0.00045 |
| HLA-B*35:43 | 0.00044 |
| HLA-B*39:10 | 0.00043 |
| HLA-B*42:02 | 0.00041 |
| HLA-B*27:04 | 0.00041 |
| HLA-C*08:04 | 0.00036 |
| HLA-B*78:01 | 0.00033 |
| HLA-C*12:04 | 0.00030 |
| HLA-B*39:09 | 0.00029 |
| HLA-A*66:02 | 0.00028 |
| HLA-A*02:22 | 0.00025 |
| HLA-B*15:04 | 0.00021 |
| HLA-B*40:04 | 0.00021 |
| HLA-B*15:11 | 0.00020 |
| HLA-B*67:01 | 0.00014 |
| HLA-B*35:20 | 0.00013 |
| HLA-A*11:03 | 0.00013 |
| HLA-C*04:04 | 0.00012 |
| HLA-A*29:10 | 0.00012 |
| HLA-B*14:03 | 0.00010 |
| HLA-A*26:02 | 0.00009 |
| HLA-B*59:01 | 0.00009 |
| HLA-C*01:03 | 0.00008 |
| HLA-B*40:11 | 0.00007 |
| HLA-A*66:03 | 0.00007 |
| HLA-A*26:03 | 0.00006 |
| HLA-A*02:24 | 0.00006 |
| HLA-B*39:13 | 0.00004 |
| HLA-C*06:06 | 0.00003 |
| HLA-A*02:14 | 0.00002 |
| HLA-A*43:01 | 0.00002 |
| HLA-A*36:03 | 0.00001 |
| HLA-B*40:64 | 0.00001 |
| HLA-A*02:87 | 0.00000 |
| HLA-C*05:11 | 0.00000 |
| HLA-A*03:17 | 0.00000 |
| HLA-A*24:04 | 0.00000 |
| HLA-B*13:03 | 0.00000 |
| HLA-C*06:11 | 0.00000 |
| HLA-A*24:24 | 0.00000 |
| HLA-B*41:04 | 0.00000 |
| HLA-B*49:02 | 0.00000 |
References
- Perez-Diez, A.; Spiess, P.J.; Restifo, N.P.; Matzinger, P.; Marincola, F.M. Intensity of the Vaccine-Elicited Immune Response Determines Tumor Clearance. J. Immunol. 2002, 168, 338–347. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.-H.; Wang, E.; Nielsen, M.-B.; Wunderlich, J.; Migueles, S.; Connors, M.; Steinberg, S.M.; Rosenberg, S.A.; Marincola, F.M. Increased Vaccine-Specific T Cell Frequency After Peptide-Based Vaccination Correlates with Increased Susceptibility to In Vitro Stimulation But Does Not Lead to Tumor Regression. J. Immunol. 1999, 163, 6292–6300. [Google Scholar]
- Disis, M.L. Immunologic biomarkers as correlates of clinical response to cancer immunotherapy. Cancer Immunol. Immunother. 2011, 60, 433–442. [Google Scholar] [CrossRef]
- Wheeler, C.J.; Black, K.L.; Liu, G.; Mazer, M.; Zhang, X.X.; Pepkowitz, S.; Goldfinger, D.; Ng, H.; Irvin, D.; Yu, J.S. Vaccination elicits correlated immune and clinical responses in glioblastoma multiforme patients. Cancer Res. 2008, 68, 5955–5964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walter, S.; Weinschenk, T.; Stenzl, A.; Zdrojowy, R.; Pluzanska, A.; Szczylik, C.; Staehler, M.; Brugger, W.; Dietrich, P.Y.; Mendrzyk, R.; et al. Multipeptide immune response to cancer vaccine IMA901 after single-dose cyclophosphamide associates with longer patient survival. Nat. Med. 2012, 18, 1254–1261. [Google Scholar] [CrossRef] [PubMed]
- Schneble, E.J.; Berry, J.S.; Trappey, F.A.; Clifton, G.T.; Ponniah, S.; Mittendorf, E.; Peoples, G.E. The HER2 peptide nelipepimut-S (E75) vaccine (NeuVax™) in breast cancer patients at risk for recurrence: Correlation of immunologic data with clinical response. Immunotherapy 2014, 6, 519–531. [Google Scholar] [CrossRef] [Green Version]
- Zinkernagel, R.M.; Doherty, P.C. Restriction of in vitro T cell-mediated cytotoxicity in lymphocytic choriomeningitis within a syngeneic or semiallogeneic system. Nature 1974, 248, 701–702. [Google Scholar] [CrossRef] [PubMed]
- Zinkernagel, R.M.; Doherty, P.C. Immunological surveillance against altered self components by sensitised T lymphocytes in lymphocytic choriomeningitis. Nature 1974, 251, 547–548. [Google Scholar] [CrossRef] [PubMed]
- Bjorkman, P.J.; Saper, M.A.; Samraoui, B.; Bennett, W.S.; Strominger, J.L.; Wiley, D.C. Structure of the human class I histocompatibility antigen, HLA-A2. Nature 1987, 329, 506–512. [Google Scholar] [CrossRef]
- Bjorkman, P.J.; Saper, M.A.; Samraoui, B.; Bennett, W.S.; Strominger, J.L.; Wiley, D.C. The foreign antigen binding site and T cell recognition regions of class I histocompatibility antigens. Nature 1987, 329, 512–518. [Google Scholar] [CrossRef]
- Fritsch, E.F.; Rajasagi, M.; Ott, P.A.; Brusic, V.; Hacohen, N.; Wu, C.J. HLA-binding properties of tumor neoepitopes in humans. Cancer Immunol. Res. 2014, 2, 522–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, I.; Gil, A.; Mishra, R.; Ghersi, D.; Selin, L.K.; Stern, L.J. Broad TCR repertoire and diverse structural solutions for recognition of an immunodominant CD8(+) T cell epitope. Nat. Struct. Mol. Biol. 2017, 24, 395–406. [Google Scholar] [CrossRef]
- Cachot, A.; Bilous, M.; Liu, Y.C.; Li, X.; Saillard, M.; Cenerenti, M.; Rockinger, G.A.; Wyss, T.; Guillaume, P.; Schmidt, J.; et al. Tumor-specific cytolytic CD4 T cells mediate immunity against human cancer. Sci. Adv. 2021, 7, eabe3348. [Google Scholar] [CrossRef]
- Dash, P.; Fiore-Gartland, A.J.; Hertz, T.; Wang, G.C.; Sharma, S.; Souquette, A.; Crawford, J.C.; Clemens, E.B.; Nguyen, T.H.O.; Kedzierska, K.; et al. Quantifiable predictive features define epitope-specific T cell receptor repertoires. Nature 2017, 547, 89–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jokinen, E.; Huuhtanen, J.; Mustjoki, S.; Heinonen, M.; Lahdesmaki, H. Predicting recognition between T cell receptors and epitopes with TCRGP. PLoS Comput. Biol. 2021, 17, e1008814. [Google Scholar] [CrossRef] [PubMed]
- Mei, S.; Li, F.; Leier, A.; Marquez-Lago, T.T.; Giam, K.; Croft, N.P.; Akutsu, T.; Smith, A.I.; Li, J.; Rossjohn, J.; et al. A comprehensive review and performance evaluation of bioinformatics tools for HLA class I peptide-binding prediction. Brief. Bioinform. 2020, 21, 1119–1135. [Google Scholar] [CrossRef]
- Van Rooij, N.; van Buuren, M.M.; Philips, D.; Velds, A.; Toebes, M.; Heemskerk, B.; van Dijk, L.J.; Behjati, S.; Hilkmann, H.; El Atmioui, D.; et al. Tumor exome analysis reveals neoantigen-specific T-cell reactivity in an ipilimumab-responsive melanoma. J. Clin. Oncol. 2013, 31, e439-442. [Google Scholar] [CrossRef]
- Ferretti, A.P.; Kula, T.; Wang, Y.; Nguyen, D.M.V.; Weinheimer, A.; Dunlap, G.S.; Xu, Q.; Nabilsi, N.; Perullo, C.R.; Cristofaro, A.W.; et al. Unbiased Screens Show CD8(+) T Cells of COVID-19 Patients Recognize Shared Epitopes in SARS-CoV-2 that Largely Reside outside the Spike Protein. Immunity 2020, 53, 1095–1107.e93. [Google Scholar] [CrossRef]
- Gonzalez-Galarza, F.F.; McCabe, A.; Santos, E.; Jones, J.; Takeshita, L.; Ortega-Rivera, N.D.; Cid-Pavon, G.M.D.; Ramsbottom, K.; Ghattaoraya, G.; Alfirevic, A.; et al. Allele frequency net database (AFND) 2020 update: Gold-standard data classification, open access genotype data and new query tools. Nucleic Acids Res. 2020, 48, D783–D788. [Google Scholar] [CrossRef] [PubMed]
- Maiers, M.; Gragert, L.; Klitz, W. High-resolution HLA alleles and haplotypes in the United States population. Hum. Immunol. 2007, 68, 779–788. [Google Scholar] [CrossRef]
- Qazilbash, M.H.; Wieder, E.; Thall, P.F.; Wang, X.; Rios, R.; Lu, S.; Kanodia, S.; Ruisaard, K.E.; Giralt, S.A.; Estey, E.H.; et al. PR1 peptide vaccine induces specific immunity with clinical responses in myeloid malignancies. Leukemia 2017, 31, 697–704. [Google Scholar] [CrossRef] [Green Version]
- Gross, S.; Lennerz, V.; Gallerani, E.; Mach, N.; Bohm, S.; Hess, D.; von Boehmer, L.; Knuth, A.; Ochsenbein, A.; Gnad-Vogt, U.; et al. Short Peptide Vaccine Induces CD4+ T Helper Cells in Patients with Different Solid Cancers. Cancer Immunol. Res. 2016, 4, 18–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukuda, K.; Funakoshi, T.; Sakurai, T.; Nakamura, Y.; Mori, M.; Tanese, K.; Tanikawa, A.; Taguchi, J.; Fujita, T.; Okamoto, M.; et al. Peptide-pulsed dendritic cell vaccine in combination with carboplatin and paclitaxel chemotherapy for stage IV melanoma. Melanoma Res. 2017, 27, 326–334. [Google Scholar] [CrossRef] [PubMed]
- Slingluff, C.L., Jr.; Petroni, G.R.; Chianese-Bullock, K.A.; Smolkin, M.E.; Ross, M.I.; Haas, N.B.; von Mehren, M.; Grosh, W.W. Randomized multicenter trial of the effects of melanoma-associated helper peptides and cyclophosphamide on the immunogenicity of a multipeptide melanoma vaccine. J. Clin. Oncol. 2011, 29, 2924–2932. [Google Scholar] [CrossRef] [Green Version]
- Sahin, U.; Derhovanessian, E.; Miller, M.; Kloke, B.P.; Simon, P.; Lower, M.; Bukur, V.; Tadmor, A.D.; Luxemburger, U.; Schrors, B.; et al. Personalized RNA mutanome vaccines mobilize poly-specific therapeutic immunity against cancer. Nature 2017, 547, 222–226. [Google Scholar] [CrossRef]
- Ott, P.A.; Hu, Z.; Keskin, D.B.; Shukla, S.A.; Sun, J.; Bozym, D.J.; Zhang, W.; Luoma, A.; Giobbie-Hurder, A.; Peter, L.; et al. An immunogenic personal neoantigen vaccine for patients with melanoma. Nature 2017, 547, 217–221. [Google Scholar] [CrossRef] [PubMed]
- Hilf, N.; Kuttruff-Coqui, S.; Frenzel, K.; Bukur, V.; Stevanovic, S.; Gouttefangeas, C.; Platten, M.; Tabatabai, G.; Dutoit, V.; van der Burg, S.H.; et al. Actively personalized vaccination trial for newly diagnosed glioblastoma. Nature 2019, 565, 240–245. [Google Scholar] [CrossRef] [PubMed]
- Want, M.Y.; Konstorum, A.; Huang, R.Y.; Jain, V.; Matsueda, S.; Tsuji, T.; Lugade, A.; Odunsi, K.; Koya, R.; Battaglia, S. Neoantigens retention in patient derived xenograft models mediates autologous T cells activation in ovarian cancer. Oncoimmunology 2019, 8, e1586042. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, M.; Lund, O.; Buus, S.; Lundegaard, C. MHC class II epitope predictive algorithms. Immunology 2010, 130, 319–328. [Google Scholar] [CrossRef]
- Bulik-Sullivan, B.; Busby, J.; Palmer, C.D.; Davis, M.J.; Murphy, T.; Clark, A.; Busby, M.; Duke, F.; Yang, A.; Young, L.; et al. Deep learning using tumor HLA peptide mass spectrometry datasets improves neoantigen identification. Nat. Biotechnol. 2018, 37, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Lam, H.; McNeil, L.K.; Starobinets, H.; DeVault, V.L.; Cohen, R.B.; Twardowski, P.; Johnson, M.L.; Gillison, M.L.; Stein, M.N.; Vaishampayan, U.N.; et al. An Empirical Antigen Selection Method Identifies Neoantigens That Either Elicit Broad Antitumor T-cell Responses or Drive Tumor Growth. Cancer Discov. 2021, 11, 696–713. [Google Scholar] [CrossRef]
- Finn, O.J.; Rammensee, H.G. Is It Possible to Develop Cancer Vaccines to Neoantigens, What Are the Major Challenges, and How Can These Be Overcome? Neoantigens: Nothing New in Spite of the Name. Cold Spring Harb. Perspect. Biol. 2018, 10, a028829. [Google Scholar] [CrossRef]
- Eisenhauer, E.A.; Therasse, P.; Bogaerts, J.; Schwartz, L.H.; Sargent, D.; Ford, R.; Dancey, J.; Arbuck, S.; Gwyther, S.; Mooney, M.; et al. New response evaluation criteria in solid tumours: Revised RECIST guideline (version 1.1). Eur. J. Cancer 2009, 45, 228–247. [Google Scholar] [CrossRef] [PubMed]
- Julka, P.K.; Doval, D.C.; Gupta, S.; Rath, G.K. Response assessment in solid tumours: A comparison of WHO, SWOG and RECIST guidelines. Br. J. Radiol. 2008, 81, 444–449. [Google Scholar] [CrossRef]
- Durie, B.G.; Harousseau, J.L.; Miguel, J.S.; Blade, J.; Barlogie, B.; Anderson, K.; Gertz, M.; Dimopoulos, M.; Westin, J.; Sonneveld, P.; et al. International uniform response criteria for multiple myeloma. Leukemia 2006, 20, 1467–1473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silverman, L.R.; Demakos, E.P.; Peterson, B.L.; Kornblith, A.B.; Holland, J.C.; Odchimar-Reissig, R.; Stone, R.M.; Nelson, D.; Powell, B.L.; DeCastro, C.M.; et al. Randomized controlled trial of azacitidine in patients with the myelodysplastic syndrome: A study of the cancer and leukemia group B. J. Clin. Oncol. 2002, 20, 2429–2440. [Google Scholar] [CrossRef] [PubMed]
- Dillon, P.M.; Petroni, G.R.; Smolkin, M.E.; Brenin, D.R.; Chianese-Bullock, K.A.; Smith, K.T.; Olson, W.C.; Fanous, I.S.; Nail, C.J.; Brenin, C.M.; et al. A pilot study of the immunogenicity of a 9-peptide breast cancer vaccine plus poly-ICLC in early stage breast cancer. J. Immunother. Cancer 2017, 5, 92–101. [Google Scholar] [CrossRef] [Green Version]
- Nakagawa, H.; Mizukoshi, E.; Kobayashi, E.; Tamai, T.; Hamana, H.; Ozawa, T.; Kishi, H.; Kitahara, M.; Yamashita, T.; Arai, K.; et al. Association Between High-Avidity T-Cell Receptors, Induced by alpha-Fetoprotein-Derived Peptides, and Anti-Tumor Effects in Patients With Hepatocellular Carcinoma. Gastroenterology 2017, 152, 1395–1406. [Google Scholar] [CrossRef]
- Mehrotra, S.; Britten, C.D.; Chin, S.; Garrett-Mayer, E.; Cloud, C.A.; Li, M.; Scurti, G.; Salem, M.L.; Nelson, M.H.; Thomas, M.B.; et al. Vaccination with poly(IC:LC) and peptide-pulsed autologous dendritic cells in patients with pancreatic cancer. J. Hematol. Oncol. 2017, 10, 82–94. [Google Scholar] [CrossRef] [Green Version]
- Banchereau, J.; Palucka, A.K.; Dhodapkar, M.; Burkeholder, S.; Taquet, N.; Rolland, A.; Taquet, S.; Coquery, S.; Wittkowski, K.M.; Bhardwaj, N.; et al. Immune and clinical responses in patients with metastatic melanoma to CD34(+) progenitor-derived dendritic cell vaccine. Cancer Res. 2001, 61, 6451–6458. [Google Scholar]
- Rausch, S.; Schwentner, C.; Stenzl, A.; Bedke, J. mRNA vaccine CV9103 and CV9104 for the treatment of prostate cancer. Hum. Vaccin. Immunother. 2014, 10, 3146–3152. [Google Scholar] [CrossRef] [Green Version]
- Kubler, H.; Scheel, B.; Gnad-Vogt, U.; Miller, K.; Schultze-Seemann, W.; Vom Dorp, F.; Parmiani, G.; Hampel, C.; Wedel, S.; Trojan, L.; et al. Self-adjuvanted mRNA vaccination in advanced prostate cancer patients: A first-in-man phase I/IIa study. J. Immunother. Cancer 2015, 3, 26–39. [Google Scholar] [CrossRef] [Green Version]
- Karkada, M.; Berinstein, N.L.; Mansour, M. Therapeutic vaccines and cancer: Focus on DPX-0907. Biologics 2014, 8, 27–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berinstein, N.L.; Karkada, M.; Morse, M.A.; Nemunaitis, J.J.; Chatta, G.; Kaufman, H.; Odunsi, K.; Nigam, R.; Sammatur, L.; MacDonald, L.D.; et al. First-in-man application of a novel therapeutic cancer vaccine formulation with the capacity to induce multi-functional T cell responses in ovarian, breast and prostate cancer patients. J. Transl. Med. 2012, 10, 156–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ponomarenko, D.M.; Klimova, I.D.; Chapygina, Y.A.; Dvornichenko, V.V.; Zhukova, N.V.; Orlova, R.V.; Manikhas, G.M.; Zyryanov, A.V.; Burkhanova, L.A.; Badrtdinova, I.I.; et al. Safety and efficacy of p62 DNA vaccine ELENAGEN in a first-in-human trial in patients with advanced solid tumors. Oncotarget 2017, 8, 53730–53739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murahashi, M.; Hijikata, Y.; Yamada, K.; Tanaka, Y.; Kishimoto, J.; Inoue, H.; Marumoto, T.; Takahashi, A.; Okazaki, T.; Takeda, K.; et al. Phase I clinical trial of a five-peptide cancer vaccine combined with cyclophosphamide in advanced solid tumors. Clin. Immunol. 2016, 166–167, 48–58. [Google Scholar] [CrossRef] [PubMed]
- Pollack, I.F.; Jakacki, R.I.; Butterfield, L.H.; Hamilton, R.L.; Panigrahy, A.; Normolle, D.P.; Connelly, A.K.; Dibridge, S.; Mason, G.; Whiteside, T.L.; et al. Immune responses and outcome after vaccination with glioma-associated antigen peptides and poly-ICLC in a pilot study for pediatric recurrent low-grade gliomas. Neuro-Oncol. 2016, 18, 1157–1168. [Google Scholar] [CrossRef] [PubMed]
- Pollack, I.F.; Jakacki, R.I.; Butterfield, L.H.; Hamilton, R.L.; Panigrahy, A.; Potter, D.M.; Connelly, A.K.; Dibridge, S.A.; Whiteside, T.L.; Okada, H. Antigen-specific immune responses and clinical outcome after vaccination with glioma-associated antigen peptides and polyinosinic-polycytidylic acid stabilized by lysine and carboxymethylcellulose in children with newly diagnosed malignant brainstem and nonbrainstem gliomas. J. Clin. Oncol. 2014, 32, 2050–2058. [Google Scholar] [CrossRef] [Green Version]
- Voskens, C.J.; Sewell, D.; Hertzano, R.; DeSanto, J.; Rollins, S.; Lee, M.; Taylor, R.; Wolf, J.; Suntharalingam, M.; Gastman, B.; et al. Induction of MAGE-A3 and HPV-16 immunity by Trojan vaccines in patients with head and neck carcinoma. Head Neck 2012, 34, 1734–1746. [Google Scholar] [CrossRef]
- Rapoport, A.P.; Aqui, N.A.; Stadtmauer, E.A.; Vogl, D.T.; Xu, Y.Y.; Kalos, M.; Cai, L.; Fang, H.B.; Weiss, B.M.; Badros, A.; et al. Combination immunotherapy after ASCT for multiple myeloma using MAGE-A3/Poly-ICLC immunizations followed by adoptive transfer of vaccine-primed and costimulated autologous T cells. Clin. Cancer Res. 2014, 20, 1355–1365. [Google Scholar] [CrossRef] [Green Version]
- Zandberg, D.P.; Rollins, S.; Goloubeva, O.; Morales, R.E.; Tan, M.; Taylor, R.; Wolf, J.S.; Schumaker, L.M.; Cullen, K.J.; Zimrin, A.; et al. A phase I dose escalation trial of MAGE-A3- and HPV16-specific peptide immunomodulatory vaccines in patients with recurrent/metastatic (RM) squamous cell carcinoma of the head and neck (SCCHN). Cancer Immunol. Immunother. 2015, 64, 367–379. [Google Scholar] [CrossRef] [PubMed]
- Tsuchiya, N.; Hosono, A.; Yoshikawa, T.; Shoda, K.; Nosaka, K.; Shimomura, M.; Hara, J.; Nitani, C.; Manabe, A.; Yoshihara, H.; et al. Phase I study of glypican-3-derived peptide vaccine therapy for patients with refractory pediatric solid tumors. Oncoimmunology 2017, 7, e1377872. [Google Scholar] [CrossRef] [Green Version]
- Tsuchiya, N.; Yoshikawa, T.; Fujinami, N.; Saito, K.; Mizuno, S.; Sawada, Y.; Endo, I.; Nakatsura, T. Immunological efficacy of glypican-3 peptide vaccine in patients with advanced hepatocellular carcinoma. Oncoimmunology 2017, 6, e1346764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fenoglio, D.; Traverso, P.; Parodi, A.; Tomasello, L.; Negrini, S.; Kalli, F.; Battaglia, F.; Ferrera, F.; Sciallero, S.; Murdaca, G.; et al. A multi-peptide, dual-adjuvant telomerase vaccine (GX301) is highly immunogenic in patients with prostate and renal cancer. Cancer Immunol. Immunother. 2013, 62, 1041–1052. [Google Scholar] [CrossRef]
- Caballero, I.; Aira, L.E.; Lavastida, A.; Popa, X.; Rivero, J.; Gonzalez, J.; Mesa, M.; Gonzalez, N.; Coba, K.; Lorenzo-Luaces, P.; et al. Safety and Immunogenicity of a Human Epidermal Growth Factor Receptor 1 (HER1)-Based Vaccine in Prostate Castration-Resistant Carcinoma Patients: A Dose-Escalation Phase I Study Trial. Front. Pharm. 2017, 8, 263–268. [Google Scholar] [CrossRef] [Green Version]
- Bekaii-Saab, T.; Wesolowski, R.; Ahn, D.H.; Wu, C.; Mortazavi, A.; Lustberg, M.B.; Ramaswamy, B.; Fowler, J.; Wei, L.; Overholser, J.; et al. Phase 1 Immunotherapy Trial with Two Chimeric HER-2 B-Cell Peptide Vaccines emulsified in Montanide ISA 720VG and nor-MDP Adjuvant in Advanced Solid Tumors. Clin. Cancer Res. 2019, 25, 3495–3507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knutson, K.L.; Schiffman, K.; Disis, M.L. Immunization with a HER-2/neu helper peptide vaccine generates HER-2/neu CD8 T-cell immunity in cancer patients. J. Clin. Investig. 2001, 107, 477–484. [Google Scholar] [CrossRef] [Green Version]
- Gudmundsdotter, L.; Wahren, B.; Haller, B.K.; Boberg, A.; Edback, U.; Bernasconi, D.; Butto, S.; Gaines, H.; Imami, N.; Gotch, F.; et al. Amplified antigen-specific immune responses in HIV-1 infected individuals in a double blind DNA immunization and therapy interruption trial. Vaccine 2011, 29, 5558–5566. [Google Scholar] [CrossRef]
- Kenter, G.G.; Welters, M.J.; Valentijn, A.R.; Lowik, M.J.; Berends-van der Meer, D.M.; Vloon, A.P.; Essahsah, F.; Fathers, L.M.; Offringa, R.; Drijfhout, J.W.; et al. Vaccination against HPV-16 oncoproteins for vulvar intraepithelial neoplasia. New Engl. J. Med. 2009, 361, 1838–1847. [Google Scholar] [CrossRef] [Green Version]
- Welters, M.J.; Kenter, G.G.; Piersma, S.J.; Vloon, A.P.; Lowik, M.J.; Berends-van der Meer, D.M.; Drijfhout, J.W.; Valentijn, A.R.; Wafelman, A.R.; Oostendorp, J.; et al. Induction of tumor-specific CD4+ and CD8+ T-cell immunity in cervical cancer patients by a human papillomavirus type 16 E6 and E7 long peptides vaccine. Clin. Cancer Res. 2008, 14, 178–187. [Google Scholar] [CrossRef] [Green Version]
- Massarelli, E.; William, W.; Johnson, F.; Kies, M.; Ferrarotto, R.; Guo, M.; Feng, L.; Lee, J.J.; Tran, H.; Kim, Y.U.; et al. Combining Immune Checkpoint Blockade and Tumor-Specific Vaccine for Patients With Incurable Human Papillomavirus 16-Related Cancer: A Phase 2 Clinical Trial. JAMA Oncol. 2018, 5, 67–73. [Google Scholar] [CrossRef] [Green Version]
- Phuphanich, S.; Wheeler, C.J.; Rudnick, J.D.; Mazer, M.; Wang, H.; Nuno, M.A.; Richardson, J.E.; Fan, X.; Ji, J.; Chu, R.M.; et al. Phase I trial of a multi-epitope-pulsed dendritic cell vaccine for patients with newly diagnosed glioblastoma. Cancer Immunol. Immunother. 2013, 62, 125–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nitschke, N.J.; Bjoern, J.; Iversen, T.Z.; Andersen, M.H.; Svane, I.M. Indoleamine 2,3-dioxygenase and survivin peptide vaccine combined with temozolomide in metastatic melanoma. Stem Cell Investig. 2017, 4, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bjoern, J.; Iversen, T.Z.; Nitschke, N.J.; Andersen, M.H.; Svane, I.M. Safety, immune and clinical responses in metastatic melanoma patients vaccinated with a long peptide derived from indoleamine 2,3-dioxygenase in combination with ipilimumab. Cytotherapy 2016, 18, 1043–1055. [Google Scholar] [CrossRef] [PubMed]
- Migliorini, D.; Dutoit, V.; Allard, M.; Grandjean Hallez, N.; Marinari, E.; Widmer, V.; Philippin, G.; Corlazzoli, F.; Gustave, R.; Kreutzfeldt, M.; et al. Phase I/II trial testing safety and immunogenicity of the multipeptide IMA950/poly-ICLC vaccine in newly diagnosed adult malignant astrocytoma patients. Neuro-Oncol. 2019, 21, 923–933. [Google Scholar] [CrossRef] [Green Version]
- Dutoit, V.; Migliorini, D.; Ranzanici, G.; Marinari, E.; Widmer, V.; Lobrinus, J.A.; Momjian, S.; Costello, J.; Walker, P.R.; Okada, H.; et al. Antigenic expression and spontaneous immune responses support the use of a selected peptide set from the IMA950 glioblastoma vaccine for immunotherapy of grade II and III glioma. Oncoimmunology 2018, 7, e1391972. [Google Scholar] [CrossRef]
- Rampling, R.; Peoples, S.; Mulholland, P.J.; James, A.; Al-Salihi, O.; Twelves, C.J.; McBain, C.; Jefferies, S.; Jackson, A.; Stewart, W.; et al. A Cancer Research UK First Time in Human Phase I Trial of IMA950 (Novel Multipeptide Therapeutic Vaccine) in Patients with Newly Diagnosed Glioblastoma. Clin. Cancer Res. 2016, 22, 4776–4785. [Google Scholar] [CrossRef] [Green Version]
- Carmon, L.; Avivi, I.; Kovjazin, R.; Zuckerman, T.; Dray, L.; Gatt, M.E.; Or, R.; Shapira, M.Y. Phase I/II study exploring ImMucin, a pan-major histocompatibility complex, anti-MUC1 signal peptide vaccine, in multiple myeloma patients. Br. J. Haematol. 2015, 169, 44–56. [Google Scholar] [CrossRef] [Green Version]
- Kovjazin, R.; Volovitz, I.; Kundel, Y.; Rosenbaum, E.; Medalia, G.; Horn, G.; Smorodinsky, N.I.; Brenner, B.; Carmon, L. ImMucin: A novel therapeutic vaccine with promiscuous MHC binding for the treatment of MUC1-expressing tumors. Vaccine 2011, 29, 4676–4686. [Google Scholar] [CrossRef]
- Legat, A.; Maby-El Hajjami, H.; Baumgaertner, P.; Cagnon, L.; Abed Maillard, S.; Geldhof, C.; Iancu, E.M.; Lebon, L.; Guillaume, P.; Dojcinovic, D.; et al. Vaccination with LAG-3Ig (IMP321) and Peptides Induces Specific CD4 and CD8 T-Cell Responses in Metastatic Melanoma Patients--Report of a Phase I/IIa Clinical Trial. Clin. Cancer Res. 2016, 22, 1330–1340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiappori, A.A.; Soliman, H.; Janssen, W.E.; Antonia, S.J.; Gabrilovich, D.I. INGN-225: A dendritic cell-based p53 vaccine (Ad.p53-DC) in small cell lung cancer: Observed association between immune response and enhanced chemotherapy effect. Expert Opin. Biol. 2010, 10, 983–991. [Google Scholar] [CrossRef]
- Asahara, S.; Takeda, K.; Yamao, K.; Maguchi, H.; Yamaue, H. Phase I/II clinical trial using HLA-A24-restricted peptide vaccine derived from KIF20A for patients with advanced pancreatic cancer. J. Transl. Med. 2013, 11, 291–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noguchi, M.; Arai, G.; Matsumoto, K.; Naito, S.; Moriya, F.; Suekane, S.; Komatsu, N.; Matsueda, S.; Sasada, T.; Yamada, A.; et al. Phase I trial of a cancer vaccine consisting of 20 mixed peptides in patients with castration-resistant prostate cancer: Dose-related immune boosting and suppression. Cancer Immunol. Immunother. 2015, 64, 493–505. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Bade, E.; Kuniyoshi, C.; Spears, L.; Jeffery, G.; Marty, V.; Groshen, S.; Weber, J. Phase I trial of a MART-1 peptide vaccine with incomplete Freund’s adjuvant for resected high-risk melanoma. Clin. Cancer Res. 1999, 5, 2756–2765. [Google Scholar]
- Slingluff, C.L., Jr.; Petroni, G.R.; Yamshchikov, G.V.; Barnd, D.L.; Eastham, S.; Galavotti, H.; Patterson, J.W.; Deacon, D.H.; Hibbitts, S.; Teates, D.; et al. Clinical and immunologic results of a randomized phase II trial of vaccination using four melanoma peptides either administered in granulocyte-macrophage colony-stimulating factor in adjuvant or pulsed on dendritic cells. J. Clin. Oncol. 2003, 21, 4016–4026. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, K.; Ikeda, Y.; Kunugi, Y.; Kurosaki, A.; Imai, Y.; Kohyama, S.; Nagao, S.; Kozawa, E.; Yoshida, K.; Tsunoda, T.; et al. Phase I Study of Multiple Epitope Peptide Vaccination in Patients With Recurrent or Persistent Cervical Cancer. J. Immunother. 2018, 41, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Wada, H.; Isobe, M.; Kakimi, K.; Mizote, Y.; Eikawa, S.; Sato, E.; Takigawa, N.; Kiura, K.; Tsuji, K.; Iwatsuki, K.; et al. Vaccination with NY-ESO-1 overlapping peptides mixed with Picibanil OK-432 and montanide ISA-51 in patients with cancers expressing the NY-ESO-1 antigen. J. Immunother. 2014, 37, 84–92. [Google Scholar] [CrossRef]
- Sabbatini, P.; Tsuji, T.; Ferran, L.; Ritter, E.; Sedrak, C.; Tuballes, K.; Jungbluth, A.A.; Ritter, G.; Aghajanian, C.; Bell-McGuinn, K.; et al. Phase I trial of overlapping long peptides from a tumor self-antigen and poly-ICLC shows rapid induction of integrated immune response in ovarian cancer patients. Clin. Cancer Res. 2012, 18, 6497–6508. [Google Scholar] [CrossRef] [Green Version]
- Kakimi, K.; Isobe, M.; Uenaka, A.; Wada, H.; Sato, E.; Doki, Y.; Nakajima, J.; Seto, Y.; Yamatsuji, T.; Naomoto, Y.; et al. A phase I study of vaccination with NY-ESO-1f peptide mixed with Picibanil OK-432 and Montanide ISA-51 in patients with cancers expressing the NY-ESO-1 antigen. Int. J. Cancer 2011, 129, 2836–2846. [Google Scholar] [CrossRef]
- Taniguchi, H.; Iwasa, S.; Yamazaki, K.; Yoshino, T.; Kiryu, C.; Naka, Y.; Liew, E.L.; Sakata, Y. Phase 1 study of OCV-C02, a peptide vaccine consisting of two peptide epitopes for refractory metastatic colorectal cancer. Cancer Sci. 2017, 108, 1013–1021. [Google Scholar] [CrossRef] [PubMed]
- Zeestraten, E.C.; Speetjens, F.M.; Welters, M.J.; Saadatmand, S.; Stynenbosch, L.F.; Jongen, R.; Kapiteijn, E.; Gelderblom, H.; Nijman, H.W.; Valentijn, A.R.; et al. Addition of interferon-alpha to the p53-SLP(R) vaccine results in increased production of interferon-gamma in vaccinated colorectal cancer patients: A phase I/II clinical trial. Int. J. Cancer 2013, 132, 1581–1591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Speetjens, F.M.; Kuppen, P.J.; Welters, M.J.; Essahsah, F.; Voet van den Brink, A.M.; Lantrua, M.G.; Valentijn, A.R.; Oostendorp, J.; Fathers, L.M.; Nijman, H.W.; et al. Induction of p53-specific immunity by a p53 synthetic long peptide vaccine in patients treated for metastatic colorectal cancer. Clin. Cancer Res. 2009, 15, 1086–1095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vermeij, R.; Leffers, N.; Hoogeboom, B.N.; Hamming, I.L.; Wolf, R.; Reyners, A.K.; Molmans, B.H.; Hollema, H.; Bart, J.; Drijfhout, J.W.; et al. Potentiation of a p53-SLP vaccine by cyclophosphamide in ovarian cancer: A single-arm phase II study. Int. J. Cancer 2012, 131, E670–E680. [Google Scholar] [CrossRef] [PubMed]
- Leffers, N.; Lambeck, A.J.; Gooden, M.J.; Hoogeboom, B.N.; Wolf, R.; Hamming, I.E.; Hepkema, B.G.; Willemse, P.H.; Molmans, B.H.; Hollema, H.; et al. Immunization with a P53 synthetic long peptide vaccine induces P53-specific immune responses in ovarian cancer patients, a phase II trial. Int. J. Cancer 2009, 125, 2104–2113. [Google Scholar] [CrossRef]
- Hardwick, N.R.; Frankel, P.; Ruel, C.; Kilpatrick, J.; Tsai, W.; Kos, F.; Kaltcheva, T.; Leong, L.; Morgan, R.; Chung, V.; et al. p53-Reactive T Cells Are Associated with Clinical Benefit in Patients with Platinum-Resistant Epithelial Ovarian Cancer After Treatment with a p53 Vaccine and Gemcitabine Chemotherapy. Clin. Cancer Res. 2018, 24, 1315–1325. [Google Scholar] [CrossRef] [Green Version]
- Greenfield, W.W.; Stratton, S.L.; Myrick, R.S.; Vaughn, R.; Donnalley, L.M.; Coleman, H.N.; Mercado, M.; Moerman-Herzog, A.M.; Spencer, H.J.; Andrews-Collins, N.R.; et al. A phase I dose-escalation clinical trial of a peptide-based human papillomavirus therapeutic vaccine with Candida skin test reagent as a novel vaccine adjuvant for treating women with biopsy-proven cervical intraepithelial neoplasia 2/3. Oncoimmunology 2015, 4, e1031439. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, N.; Hazama, S.; Iguchi, H.; Uesugi, K.; Tanaka, H.; Hirakawa, K.; Aruga, A.; Hatori, T.; Ishizaki, H.; Umeda, Y.; et al. Phase II clinical trial of peptide cocktail therapy for patients with advanced pancreatic cancer: VENUS-PC study. Cancer Sci. 2017, 108, 73–80. [Google Scholar] [CrossRef] [Green Version]
- Kawamura, J.; Sugiura, F.; Sukegawa, Y.; Yoshioka, Y.; Hida, J.I.; Hazama, S.; Okuno, K. Cytotoxic T lymphocyte response to peptide vaccination predicts survival in stage III colorectal cancer. Cancer Sci. 2018, 109, 1545–1551. [Google Scholar] [CrossRef] [Green Version]
- Yoshitake, Y.; Fukuma, D.; Yuno, A.; Hirayama, M.; Nakayama, H.; Tanaka, T.; Nagata, M.; Takamune, Y.; Kawahara, K.; Nakagawa, Y.; et al. Phase II clinical trial of multiple peptide vaccination for advanced head and neck cancer patients revealed induction of immune responses and improved OS. Clin. Cancer Res. 2015, 21, 312–321. [Google Scholar] [CrossRef] [Green Version]
- Rezvani, K.; Yong, A.S.; Mielke, S.; Jafarpour, B.; Savani, B.N.; Le, R.Q.; Eniafe, R.; Musse, L.; Boss, C.; Kurlander, R.; et al. Repeated PR1 and WT1 peptide vaccination in Montanide-adjuvant fails to induce sustained high-avidity, epitope-specific CD8+ T cells in myeloid malignancies. Haematologica 2011, 96, 432–440. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, R.D.; Huh, W.K.; Bae, S.; Lamb, L.S., Jr.; Conner, M.G.; Boyer, J.; Wang, C.; Hung, C.F.; Sauter, E.; Paradis, M.; et al. A pilot study of pNGVL4a-CRT/E7(detox) for the treatment of patients with HPV16+ cervical intraepithelial neoplasia 2/3 (CIN2/3). Gynecol. Oncol. 2016, 140, 245–252. [Google Scholar] [CrossRef] [Green Version]
- Arlen, P.M.; Skarupa, L.; Pazdur, M.; Seetharam, M.; Tsang, K.Y.; Grosenbach, D.W.; Feldman, J.; Poole, D.J.; Litzinger, M.; Steinberg, S.M.; et al. Clinical safety of a viral vector based prostate cancer vaccine strategy. J. Urol. 2007, 178, 1515–1520. [Google Scholar] [CrossRef] [PubMed]
- Arlen, P.M.; Gulley, J.L.; Todd, N.; Lieberman, R.; Steinberg, S.M.; Morin, S.; Bastian, A.; Marte, J.; Tsang, K.Y.; Beetham, P.; et al. Antiandrogen, vaccine and combination therapy in patients with nonmetastatic hormone refractory prostate cancer. J. Urol. 2005, 174, 539–546. [Google Scholar] [CrossRef] [PubMed]
- Madan, R.A.; Mohebtash, M.; Arlen, P.M.; Vergati, M.; Rauckhorst, M.; Steinberg, S.M.; Tsang, K.Y.; Poole, D.J.; Parnes, H.L.; Wright, J.J.; et al. Ipilimumab and a poxviral vaccine targeting prostate-specific antigen in metastatic castration-resistant prostate cancer: A phase 1 dose-escalation trial. Lancet Oncol. 2012, 13, 501–508. [Google Scholar] [CrossRef]
- Gulley, J.L.; Arlen, P.M.; Bastian, A.; Morin, S.; Marte, J.; Beetham, P.; Tsang, K.Y.; Yokokawa, J.; Hodge, J.W.; Menard, C.; et al. Combining a recombinant cancer vaccine with standard definitive radiotherapy in patients with localized prostate cancer. Clin. Cancer Res. 2005, 11, 3353–3362. [Google Scholar] [CrossRef] [Green Version]
- Lechleider, R.J.; Arlen, P.M.; Tsang, K.Y.; Steinberg, S.M.; Yokokawa, J.; Cereda, V.; Camphausen, K.; Schlom, J.; Dahut, W.L.; Gulley, J.L. Safety and immunologic response of a viral vaccine to prostate-specific antigen in combination with radiation therapy when metronomic-dose interleukin 2 is used as an adjuvant. Clin. Cancer Res. 2008, 14, 5284–5291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gulley, J.L.; Arlen, P.M.; Madan, R.A.; Tsang, K.Y.; Pazdur, M.P.; Skarupa, L.; Jones, J.L.; Poole, D.J.; Higgins, J.P.; Hodge, J.W.; et al. Immunologic and prognostic factors associated with overall survival employing a poxviral-based PSA vaccine in metastatic castrate-resistant prostate cancer. Cancer Immunol. Immunother. 2010, 59, 663–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xi, H.B.; Wang, G.X.; Fu, B.; Liu, W.P.; Li, Y. Survivin and PSMA Loaded Dendritic Cell Vaccine for the Treatment of Prostate Cancer. Biol. Pharm. Bull. 2015, 38, 827–835. [Google Scholar] [CrossRef] [Green Version]
- Nooka, A.K.; Wang, M.L.; Yee, A.J.; Kaufman, J.L.; Bae, J.; Peterkin, D.; Richardson, P.G.; Raje, N.S. Assessment of Safety and Immunogenicity of PVX-410 Vaccine With or Without Lenalidomide in Patients With Smoldering Multiple Myeloma: A Nonrandomized Clinical Trial. JAMA Oncol. 2018, 4, e183267. [Google Scholar] [CrossRef]
- Greiner, J.; Schmitt, A.; Giannopoulos, K.; Rojewski, M.T.; Gotz, M.; Funk, I.; Ringhoffer, M.; Bunjes, D.; Hofmann, S.; Ritter, G.; et al. High-dose RHAMM-R3 peptide vaccination for patients with acute myeloid leukemia, myelodysplastic syndrome and multiple myeloma. Haematologica 2010, 95, 1191–1197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Obara, W.; Ohsawa, R.; Kanehira, M.; Takata, R.; Tsunoda, T.; Yoshida, K.; Takeda, K.; Katagiri, T.; Nakamura, Y.; Fujioka, T. Cancer peptide vaccine therapy developed from oncoantigens identified through genome-wide expression profile analysis for bladder cancer. Jpn. J. Clin. Oncol. 2012, 42, 591–600. [Google Scholar] [CrossRef]
- Butts, C.; Murray, N.; Maksymiuk, A.; Goss, G.; Marshall, E.; Soulieres, D.; Cormier, Y.; Ellis, P.; Price, A.; Sawhney, R.; et al. Randomized phase IIB trial of BLP25 liposome vaccine in stage IIIB and IV non-small-cell lung cancer. J. Clin. Oncol. 2005, 23, 6674–6681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satomi, F. Phase I Clinical Study of Survivin-Derived Peptide Vaccine for Patients with Advanced Gastrointestinal Cancers. Int J. Cancer Clin. Res. 2015, 2, 12. [Google Scholar] [CrossRef]
- Tagawa, S.T.; Lee, P.; Snively, J.; Boswell, W.; Ounpraseuth, S.; Lee, S.; Hickingbottom, B.; Smith, J.; Johnson, D.; Weber, J.S. Phase I study of intranodal delivery of a plasmid DNA vaccine for patients with Stage IV melanoma. Cancer 2003, 98, 144–154. [Google Scholar] [CrossRef]
- Wood, L.V.; Fojo, A.; Roberson, B.D.; Hughes, M.S.; Dahut, W.; Gulley, J.L.; Madan, R.A.; Arlen, P.M.; Sabatino, M.; Stroncek, D.F.; et al. TARP vaccination is associated with slowing in PSA velocity and decreasing tumor growth rates in patients with Stage D0 prostate cancer. Oncoimmunology 2016, 5, e1197459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rochlitz, C.; Figlin, R.; Squiban, P.; Salzberg, M.; Pless, M.; Herrmann, R.; Tartour, E.; Zhao, Y.; Bizouarne, N.; Baudin, M.; et al. Phase I immunotherapy with a modified vaccinia virus (MVA) expressing human MUC1 as antigen-specific immunotherapy in patients with MUC1-positive advanced cancer. J. Gene Med. 2003, 5, 690–699. [Google Scholar] [CrossRef] [PubMed]
- Dreicer, R.; Stadler, W.M.; Ahmann, F.R.; Whiteside, T.; Bizouarne, N.; Acres, B.; Limacher, J.M.; Squiban, P.; Pantuck, A. MVA-MUC1-IL2 vaccine immunotherapy (TG4010) improves PSA doubling time in patients with prostate cancer with biochemical failure. Investig. New Drugs 2009, 27, 379–386. [Google Scholar] [CrossRef] [PubMed]
- Oudard, S.; Rixe, O.; Beuselinck, B.; Linassier, C.; Banu, E.; Machiels, J.P.; Baudard, M.; Ringeisen, F.; Velu, T.; Lefrere-Belda, M.A.; et al. A phase II study of the cancer vaccine TG4010 alone and in combination with cytokines in patients with metastatic renal clear-cell carcinoma: Clinical and immunological findings. Cancer Immunol. Immunother. 2011, 60, 261–271. [Google Scholar] [CrossRef]
- Ramlau, R.; Quoix, E.; Rolski, J.; Pless, M.; Lena, H.; Levy, E.; Krzakowski, M.; Hess, D.; Tartour, E.; Chenard, M.P.; et al. A phase II study of Tg4010 (Mva-Muc1-Il2) in association with chemotherapy in patients with stage III/IV Non-small cell lung cancer. J. Thorac. Oncol. 2008, 3, 735–744. [Google Scholar] [CrossRef]
- Cusi, M.G.; Botta, C.; Pastina, P.; Rossetti, M.G.; Dreassi, E.; Guidelli, G.M.; Fioravanti, A.; Martino, E.C.; Gandolfo, C.; Pagliuchi, M.; et al. Phase I trial of thymidylate synthase poly-epitope peptide (TSPP) vaccine in advanced cancer patients. Cancer Immunol. Immunother. 2015, 64, 1159–1173. [Google Scholar] [CrossRef] [PubMed]
- Correale, P.; Botta, C.; Martino, E.C.; Ulivieri, C.; Battaglia, G.; Carfagno, T.; Rossetti, M.G.; Fioravanti, A.; Guidelli, G.M.; Cheleschi, S.; et al. Phase Ib study of poly-epitope peptide vaccination to thymidylate synthase (TSPP) and GOLFIG chemo-immunotherapy for treatment of metastatic colorectal cancer patients. Oncoimmunology 2016, 5, e1101205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bagarazzi, M.L.; Yan, J.; Morrow, M.P.; Shen, X.; Parker, R.L.; Lee, J.C.; Giffear, M.; Pankhong, P.; Khan, A.S.; Broderick, K.E.; et al. Immunotherapy against HPV16/18 generates potent TH1 and cytotoxic cellular immune responses. Sci. Transl. Med. 2012, 4, 155ra138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trimble, C.L.; Morrow, M.P.; Kraynyak, K.A.; Shen, X.; Dallas, M.; Yan, J.; Edwards, L.; Parker, R.L.; Denny, L.; Giffear, M.; et al. Safety, efficacy, and immunogenicity of VGX-3100, a therapeutic synthetic DNA vaccine targeting human papillomavirus 16 and 18 E6 and E7 proteins for cervical intraepithelial neoplasia 2/3: A randomised, double-blind, placebo-controlled phase 2b trial. Lancet 2015, 386, 2078–2088. [Google Scholar] [CrossRef] [Green Version]
- Vetsika, E.K.; Konsolakis, G.; Aggouraki, D.; Kotsakis, A.; Papadimitraki, E.; Christou, S.; Menez-Jamet, J.; Kosmatopoulos, K.; Georgoulias, V.; Mavroudis, D. Immunological responses in cancer patients after vaccination with the therapeutic telomerase-specific vaccine Vx-001. Cancer Immunol. Immunother. 2012, 61, 157–168. [Google Scholar] [CrossRef] [PubMed]
- Kotsakis, A.; Vetsika, E.K.; Christou, S.; Hatzidaki, D.; Vardakis, N.; Aggouraki, D.; Konsolakis, G.; Georgoulias, V.; Christophyllakis, C.; Cordopatis, P.; et al. Clinical outcome of patients with various advanced cancer types vaccinated with an optimized cryptic human telomerase reverse transcriptase (TERT) peptide: Results of an expanded phase II study. Ann. Oncol. 2012, 23, 442–449. [Google Scholar] [CrossRef] [PubMed]
- Bolonaki, I.; Kotsakis, A.; Papadimitraki, E.; Aggouraki, D.; Konsolakis, G.; Vagia, A.; Christophylakis, C.; Nikoloudi, I.; Magganas, E.; Galanis, A.; et al. Vaccination of patients with advanced non-small-cell lung cancer with an optimized cryptic human telomerase reverse transcriptase peptide. J. Clin. Oncol. 2007, 25, 2727–2734. [Google Scholar] [CrossRef] [PubMed]
- Kotsakis, A.; Papadimitraki, E.; Vetsika, E.K.; Aggouraki, D.; Dermitzaki, E.K.; Hatzidaki, D.; Kentepozidis, N.; Mavroudis, D.; Georgoulias, V. A phase II trial evaluating the clinical and immunologic response of HLA-A2(+) non-small cell lung cancer patients vaccinated with an hTERT cryptic peptide. Lung Cancer 2014, 86, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Tsuboi, A.; Hashimoto, N.; Fujiki, F.; Morimoto, S.; Kagawa, N.; Nakajima, H.; Hosen, N.; Nishida, S.; Nakata, J.; Morita, S.; et al. A phase I clinical study of a cocktail vaccine of Wilms’ tumor 1 (WT1) HLA class I and II peptides for recurrent malignant glioma. Cancer Immunol. Immunother. 2019, 68, 331–340. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Lu, X.; Cui, P.; Piao, C.; Xiao, M.; Liu, X.; Wang, Y.; Wu, X.; Liu, J.; Yang, L. Phase I/II clinical trial of a Wilms’ tumor 1-targeted dendritic cell vaccination-based immunotherapy in patients with advanced cancer. Cancer Immunol. Immunother. 2019, 68, 121–130. [Google Scholar] [CrossRef]
- Yanagisawa, R.; Koizumi, T.; Koya, T.; Sano, K.; Koido, S.; Nagai, K.; Kobayashi, M.; Okamoto, M.; Sugiyama, H.; Shimodaira, S. WT1-pulsed Dendritic Cell Vaccine Combined with Chemotherapy for Resected Pancreatic Cancer in a Phase I Study. Anticancer Res. 2018, 38, 2217–2225. [Google Scholar] [CrossRef] [Green Version]
- Keilholz, U.; Letsch, A.; Busse, A.; Asemissen, A.M.; Bauer, S.; Blau, I.W.; Hofmann, W.K.; Uharek, L.; Thiel, E.; Scheibenbogen, C. A clinical and immunologic phase 2 trial of Wilms tumor gene product 1 (WT1) peptide vaccination in patients with AML and MDS. Blood 2009, 113, 6541–6548. [Google Scholar] [CrossRef] [Green Version]
- Nishida, S.; Ishikawa, T.; Egawa, S.; Koido, S.; Yanagimoto, H.; Ishii, J.; Kanno, Y.; Kokura, S.; Yasuda, H.; Oba, M.S.; et al. Combination Gemcitabine and WT1 Peptide Vaccination Improves Progression-Free Survival in Advanced Pancreatic Ductal Adenocarcinoma: A Phase II Randomized Study. Cancer Immunol. Res. 2018, 6, 320–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krug, L.M.; Dao, T.; Brown, A.B.; Maslak, P.; Travis, W.; Bekele, S.; Korontsvit, T.; Zakhaleva, V.; Wolchok, J.; Yuan, J.; et al. WT1 peptide vaccinations induce CD4 and CD8 T cell immune responses in patients with mesothelioma and non-small cell lung cancer. Cancer Immunol. Immunother. 2010, 59, 1467–1479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erlich, R.L.; Jia, X.; Anderson, S.; Banks, E.; Gao, X.; Carrington, M.; Gupta, N.; DePristo, M.A.; Henn, M.R.; Lennon, N.J.; et al. Next-generation sequencing for HLA typing of class I loci. BMC Genom. 2011, 12, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, J.; Halliwell, J.A.; Marsh, S.G.E. IMGT/HLA and the Immuno Polymorphism Database. Immunoinformatics 2014, 1184, 109–121. [Google Scholar] [CrossRef]
- Yusim, K.; Korber, B.T.M.; Brander, C.; Barouch, D.; de Boer, R.; Haynes, B.F.; Koup, R.; Moore, J.P.; Walker, B.D.; Watkins, D.I. HIV molecular immunology database. Available online: https://www.hiv.lanl.gov/content/immunology/compendium.html (accessed on 21 May 2014).
- Vita, R.; Overton, J.A.; Greenbaum, J.A.; Ponomarenko, J.; Clark, J.D.; Cantrell, J.R.; Wheeler, D.K.; Gabbard, J.L.; Hix, D.; Sette, A.; et al. The immune epitope database (IEDB) 3.0. Nucleic Acids Res. 2015, 43, D405–D412. [Google Scholar] [CrossRef] [PubMed]
- Vita, R.; Mahajan, S.; Overton, J.A.; Dhanda, S.K.; Martini, S.; Cantrell, J.R.; Wheeler, D.K.; Sette, A.; Peters, B. The Immune Epitope Database (IEDB): 2018 update. Nucleic Acids Res. 2019, 47, D339–D343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hurley, C.K.; Kempenich, J.; Wadsworth, K.; Sauter, J.; Hofmann, J.A.; Schefzyk, D.; Schmidt, A.H.; Galarza, P.; Cardozo, M.B.R.; Dudkiewicz, M.; et al. Common, intermediate and well-documented HLA alleles in world populations: CIWD version 3.0.0. HLA 2020, 95, 516–531. [Google Scholar] [CrossRef] [Green Version]
- Gragert, L.; Madbouly, A.; Freeman, J.; Maiers, M. Six-locus high resolution HLA haplotype frequencies derived from mixed-resolution DNA typing for the entire US donor registry. Hum. Immunol. 2013, 74, 1313–1320. [Google Scholar] [CrossRef] [Green Version]
- MedCalc Software Ltd. Comparison of Proportions Calculator. (Version 20.010). Available online: https://www.medcalc.org/calc/comparison_of_proportions.php (accessed on 1 March 2021).
- Campbell, I. Chi-squared and Fisher-Irwin tests of two-by-two tables with small sample recommendations. Stat. Med. 2007, 26, 3661–3675. [Google Scholar] [CrossRef]
- Richardson, J.T. The analysis of 2 × 2 contingency tables--yet again. Stat. Med. 2011, 30, 890. [Google Scholar] [CrossRef] [PubMed]
- Newcombe, R.; Altman, D. Proportions and Their Differences. In Statistics with Confidence: Confidence Intervals and Statistical Guidelines, 2nd ed.; Altman, D., Machin, D., Bryant, T., Gardner, M., Eds.; BMJ Books: London, UK, 2000; pp. 45–57. [Google Scholar]
- Fawcett, T. An introduction to ROC analysis. Pattern Recognit. Lett. 2006, 27, 861–874. [Google Scholar] [CrossRef]
- Šimundić, A.-M. Measures of Diagnostic Accuracy: Basic Definitions. EJIFCC 2009, 19, 203–211. [Google Scholar] [PubMed]
- Somogyi, E.; Csiszovszki, Z.; Molnar, L.; Lorincz, O.; Toth, J.; Pattijn, S.; Schockaert, J.; Mazy, A.; Miklos, I.; Pantya, K.; et al. A Peptide Vaccine Candidate Tailored to Individuals’ Genetics Mimics the Multi-Targeted T Cell Immunity of COVID-19 Convalescent Subjects. Front. Genet. 2021, 12, 684152. [Google Scholar] [CrossRef]
- Toke, E.R.; Megyesi, M.; Molnar, L.; Tóth, J.; Lőrincz, O.; van der Burg, S.H.; Welters, M.; Melief, C.J.; Schönharting, W.; Urban, S.; et al. Prediction the clinical outcomes of cancer patients after peptide vaccination. J. Clin. Oncol. 2019, 37, e14295. [Google Scholar] [CrossRef]
- Yuan, J.; Ku, G.Y.; Gallardo, H.F.; Orlandi, F.; Manukian, G.; Rasalan, T.S.; Xu, Y.; Li, H.; Vyas, S.; Mu, Z.; et al. Safety and immunogenicity of a human and mouse gp100 DNA vaccine in a phase I trial of patients with melanoma. Cancer Immun. 2009, 9, 5. [Google Scholar]
- Bassani-Sternberg, M.; Bräunlein, E.; Klar, R.; Engleitner, T.; Sinitcyn, P.; Audehm, S.; Straub, M.; Weber, J.; Slotta-Huspenina, J.; Specht, K.; et al. Direct identification of clinically relevant neoepitopes presented on native human melanoma tissue by mass spectrometry. Nat. Commun. 2016, 7, 13404. [Google Scholar] [CrossRef] [Green Version]
- Wells, D.K.; van Buuren, M.M.; Dang, K.K.; Hubbard-Lucey, V.M.; Sheehan, K.C.F.; Campbell, K.M.; Lamb, A.; Ward, J.P.; Sidney, J.; Blazquez, A.B.; et al. Key Parameters of Tumor Epitope Immunogenicity Revealed Through a Consortium Approach Improve Neoantigen Prediction. Cell 2020, 183, 818–834.e13. [Google Scholar] [CrossRef]
- Kirner, A.; Mayer-Mokler, A.; Reinhardt, C. IMA901: A multi-peptide cancer vaccine for treatment of renal cell cancer. Hum. Vaccin. Immunother. 2014, 10, 3179–3189. [Google Scholar] [CrossRef] [Green Version]
- Boegel, S.; Löwer, M.; Bukur, T.; Sorn, P.; Castle, J.C.; Sahin, U. HLA and proteasome expression body map. BMC Med. Genom. 2018, 11, 36. [Google Scholar] [CrossRef]
- Zhang, Y.; Renkvist, N.; Sun, Z.; Schuler-Thurner, B.; Glaichenhaus, N.; Schuler, G.; Boon, T.; van der Bruggen, P.; Colau, D. A polyclonal anti-vaccine CD4 T cell response detected with HLA-DP4 multimers in a melanoma patient vaccinated with MAGE-3.DP4-peptide-pulsed dendritic cells. Eur. J. Immunol. 2005, 35, 1066–1075. [Google Scholar] [CrossRef] [PubMed]
- Hicklin, D.J.; Marincola, F.M.; Ferrone, S. HLA class I antigen downregulation in human cancers: T-cell immunotherapy revives an old story. Mol. Med. Today 1999, 5, 178–186. [Google Scholar] [CrossRef]
- Garrido, F.; Ruiz-Cabello, F.; Aptsiauri, N. Rejection versus escape: The tumor MHC dilemma. Cancer Immunol. Immunother. 2017, 66, 259–271. [Google Scholar] [CrossRef] [PubMed]
- Chowell, D.; Morris, L.G.T.; Grigg, C.M.; Weber, J.K.; Samstein, R.M.; Makarov, V.; Kuo, F.; Kendall, S.M.; Requena, D.; Riaz, N.; et al. Patient HLA class I genotype influences cancer response to checkpoint blockade immunotherapy. Science 2018, 359, 582–587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, J.I.; Decker, S.; Zaharevitz, D.; Rubinstein, L.V.; Venditti, J.M.; Schepartz, S.; Kalyandrug, S.; Christian, M.; Arbuck, S.; Hollingshead, M.; et al. Relationships between drug activity in NCI preclinical in vitro and in vivo models and early clinical trials. Br. J. Cancer 2001, 84, 1424–1431. [Google Scholar] [CrossRef]
- Hoos, A.; Parmiani, G.; Hege, K.; Sznol, M.; Loibner, H.; Eggermont, A.; Urba, W.; Blumenstein, B.; Sacks, N.; Keilholz, U.; et al. A clinical development paradigm for cancer vaccines and related biologics. J. Immunother. 2007, 30, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bateman, A.C.; Turner, S.J.; Theaker, J.M.; Howell, W.M. HLA-DQB1*0303 and *0301 alleles influence susceptibility to and prognosis in cutaneous malignant melanoma in the British Caucasian population. Tissue Antigens 1998, 52, 67–73. [Google Scholar] [CrossRef]
- Chaudhuri, S.; Cariappa, A.; Tang, M.; Bell, D.; Haber, D.A.; Isselbacher, K.J.; Finkelstein, D.; Forcione, D.; Pillai, S. Genetic susceptibility to breast cancer: HLA DQB*03032 and HLA DRB1*11 may represent protective alleles. Proc. Natl. Acad. Sci. USA 2000, 97, 11451–11454. [Google Scholar] [CrossRef] [Green Version]
- Aureli, A.; Canossi, A.; Del Beato, T.; Franceschilli, L.; Buonomo, O.; Papola, F.; De Sanctis, F.; Lanzilli, G.; Sileri, P.; Coppola, A.; et al. HLA-DRB1*13:01 allele in the genetic susceptibility to colorectal carcinoma. Int. J. Cancer 2015, 136, 2464–2468. [Google Scholar] [CrossRef]
- Lesseur, C.; Diergaarde, B.; Olshan, A.F.; Wünsch-Filho, V.; Ness, A.R.; Liu, G.; Lacko, M.; Eluf-Neto, J.; Franceschi, S.; Lagiou, P.; et al. Genome-wide association analyses identify new susceptibility loci for oral cavity and pharyngeal cancer. Nat. Genet. 2016, 48, 1544–1550. [Google Scholar] [CrossRef]
- Krul, E.J.; Schipper, R.F.; Schreuder, G.M.; Fleuren, G.J.; Kenter, G.G.; Melief, C.J. HLA and susceptibility to cervical neoplasia. Hum. Immunol. 1999, 60, 337–342. [Google Scholar] [CrossRef]
- Kübler, K.; Arndt, P.F.; Wardelmann, E.; Krebs, D.; Kuhn, W.; van der Ven, K. HLA-class II haplotype associations with ovarian cancer. Int. J. Cancer 2006, 119, 2980–2985. [Google Scholar] [CrossRef] [PubMed]
- Marty, R.; Kaabinejadian, S.; Rossell, D.; Slifker, M.J.; van de Haar, J.; Engin, H.B.; de Prisco, N.; Ideker, T.; Hildebrand, W.H.; Font-Burgada, J.; et al. MHC-I Genotype Restricts the Oncogenic Mutational Landscape. Cell 2017, 171, 1272–1283.e15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, S.; Zhu, H.; Xia, X.; Liang, Z.; Ma, X.; Sun, B. Vaccine adjuvants: Understanding the structure and mechanism of adjuvanticity. Vaccine 2019, 37, 3167–3178. [Google Scholar] [CrossRef]
- Petrovsky, N.; Aguilar, J.C. Vaccine adjuvants: Current state and future trends. Immunol. Cell Biol. 2004, 82, 488–496. [Google Scholar] [CrossRef]
- Petrovsky, N. Comparative Safety of Vaccine Adjuvants: A Summary of Current Evidence and Future Needs. Drug Saf. 2015, 38, 1059–1074. [Google Scholar] [CrossRef] [PubMed]
- Aucouturier, J.; Dupuis, L.; Deville, S.; Ascarateil, S.; Ganne, V. Montanide ISA 720 and 51: A new generation of water in oil emulsions as adjuvants for human vaccines. Expert Rev. Vaccines 2002, 1, 111–118. [Google Scholar] [CrossRef]
- Janetzki, S.; Panageas, K.S.; Ben-Porat, L.; Boyer, J.; Britten, C.M.; Clay, T.M.; Kalos, M.; Maecker, H.T.; Romero, P.; Yuan, J.; et al. Results and harmonization guidelines from two large-scale international Elispot proficiency panels conducted by the Cancer Vaccine Consortium (CVC/SVI). Cancer Immunol. Immunother. 2008, 57, 303–315. [Google Scholar] [CrossRef] [Green Version]
- Janetzki, S.; Cox, J.H.; Oden, N.; Ferrari, G. Standardization and Validation Issues of the ELISPOT Assay. In Handbook of ELISPOT: Methods and Protocols; Kalyuzhny, A.E., Ed.; Humana Press: Totowa, NJ, USA, 2005; pp. 51–86. [Google Scholar]
- Lyerly, H.K. Quantitating cellular immune responses to cancer vaccines. Semin. Oncol. 2003, 30, 9–16. [Google Scholar] [CrossRef]
- Van der Burg, S.H.; Arens, R.; Ossendorp, F.; van Hall, T.; Melief, C.J. Vaccines for established cancer: Overcoming the challenges posed by immune evasion. Nat. Rev. Cancer 2016, 16, 219–233. [Google Scholar] [CrossRef]
- Galon, J.; Bruni, D. Approaches to treat immune hot, altered and cold tumours with combination immunotherapies. Nat. Rev. Drug Discov. 2019, 18, 197–218. [Google Scholar] [CrossRef] [PubMed]
- Saxena, M.; van der Burg, S.H.; Melief, C.J.M.; Bhardwaj, N. Therapeutic cancer vaccines. Nat. Rev. Cancer 2021, 21, 360–378. [Google Scholar] [CrossRef] [PubMed]
- Van der Burg, S.H. Correlates of immune and clinical activity of novel cancer vaccines. Semin. Immunol 2018, 39, 119–136. [Google Scholar] [CrossRef] [PubMed]



| Immunotherapy | Indication | Type | HLA Restriction | Ref. |
|---|---|---|---|---|
| 9-peptide breast cancer vaccine | BC | Peptide | HLA-A1, -A2, or -A3 | [37] |
| AFP-derived peptides | aHCC | Peptide | A24 | [38] |
| Antigen-pulsed DC vaccine | PAdC | pDC | A02 | [39] |
| CD34 DC vaccine | MEL | pDC | A02:01 | [40] |
| CV9103 mRNA vaccine | PC | mRNA | no | [41,42] |
| DCCP peptide vaccine | MEL | pDC | A24 or A02 | [23] |
| DPX0907 peptide vaccine | BC, OC, PC | Peptide | A02 | [43,44] |
| Elenagen pDNA vaccine | BC, CRC, KC, LC, OC, MEL | pDNA | no | [45] |
| EMD640744 peptide vaccine | S.tumors | Peptide | HLA-A1, -A2, -A3, -A24, -B7 | [22] |
| Five-peptide cancer vaccine | S.tumors | Peptide | A24:02 | [46] |
| GAA peptides vaccine | glioma | Peptide | A02 | [47,48] |
| GL-0817 (MAGE-A3 Trojan) | SCCHN | Peptide | no | [49] |
| MM | [50] | |||
| mSCCHN | [51] | |||
| Glypican-3 peptide vaccine | pediatric tumors | Peptide | A02 or A24:02 | [52] |
| HCC | Peptide | A02 or A24:02 | [53] | |
| GVX301 peptide vaccine | PC, KC | Peptide | A2 | [54] |
| HER1 vaccine | PC | Peptide | no | [55] |
| Her2 B-cell peptide vaccine | S.tumors | Peptide | no | [56] |
| Her2/neu peptide vaccine | BC, OC | Peptide | A02 | [57] |
| HIVIS | HIV positivity | pDNA | no | [58] |
| HPV-SLP | VIN3 | Peptide | no | [59] |
| CC | [60] | |||
| OrC, CC, AC | [61] | |||
| ICT107 | GB or BSG | Peptide | A1 or A2 | [62] |
| IDO and survivin peptide vaccine | mMEL | Peptide | A02 | [63] |
| IDO long peptide vaccine | adv. MEL | Peptide | no | [64] |
| IMA901 | RCC | Peptide | A02 | [5] |
| mRCC | ||||
| IMA950 | AC, glioma | Peptide | A02 | [65] |
| AC, ODG | [66] | |||
| GB | [67] | |||
| ImMucin | MM | Peptide | no | [68] |
| CRC, LC, PC, TT | [69] | |||
| IMP321/LAG-3Ig + peptides vaccine | mMEL | Peptide | A02 | [70] |
| INGN-225 p53 vaccine | SCLC | VV | A02 | [71] |
| KIF20A-66 peptide vaccine | met. PaC | Peptide | A24:02 | [72] |
| KRM-20 vaccine | PC | Peptide | A24 | [73] |
| MART-1 Peptide Vaccine | MEL | Peptide | A02 | [74] |
| Melanoma peptide vaccine | MEL | Peptide or pDC | A1, -A2 or -A3 | [75] |
| MELITAC 12.1 | MEL | Peptide | A1, -A2, or -A3; HLA-DR1, -DR4, -DR11, -DR13, or -DR15 | [24] |
| Multiepitope peptide vaccine | CC | Peptide | A24:02 | [76] |
| NY-ESO-1 OLP | MEL, EC, LC | Peptide | no | [77] |
| OC, FTC, PerC | [78] | |||
| NY-ESO-1f | EC, GC, NSCLC | Peptide | no | [79] |
| OCV-C02 vaccine | CRC | Peptide | A24:02 | [80] |
| p53 SLP70-235 | CRC | Peptide | no | [81] |
| p53 SLP70-248 | CRC | Peptide | no | [82] |
| EOC | [83,84] | |||
| p53MVA vaccine | EOC, FTC, PerC | VV | no | [85] |
| PepCan | CIN2/3 | Peptide | no | [86] |
| Peptide cocktail therapy | PaC | Peptide | A24:02 w/o | [87] |
| Peptide vaccine 1 | CRC | Peptide | A24:02 | [88] |
| Peptide vaccine 2 | HNSCC | Peptide | A24:02 | [89] |
| Peptide vaccine 3 | AML | Peptide | A02:01 | [90] |
| pNGVL4a-CRT/E7(detox) vaccine | CIN2/3 | pDNA | No | [91] |
| PR1 Peptide Vaccine | AML, MDS, CML | Peptide | A02 | [21] |
| ProstVac | PC | VV | A02 | [92,93,94,95,96,97] |
| PSMA-Survivin pulsed autologous DC vaccine | PC | pDC | A02:01 | [98] |
| PVX-410 peptide vaccine | MM | Peptide | A02 | [99] |
| RHAMM-R3 | AML | Peptide | A02 | [100] |
| S-288310 peptide vaccine | BLC | Peptide | A24:02 | [101] |
| StimuVax | NSCLC | Peptide | no | [102] |
| SVN-2B peptide vaccine | GIC, BDC, PaC | Peptide | A24:02 | [103] |
| Synchrotope TA2M | MEL | pDNA | A02, but pts were not typed | [104] |
| TARP vaccine | PC | Peptide or pDC | A02:01 | [105] |
| TG4010 | cancer | VV | no | [106] |
| PC | no | [107] | ||
| RCC | no | [108] | ||
| NSCLC | no | [109] | ||
| TSPP peptide vaccine | cancer | Peptide | no | [110] |
| mCRC | [111] | |||
| VGX-3100 | CIN2/3 | pDNA | no | [112,113] |
| Vx-001 | S.tumors | Peptide | A02:01 | [114,115] |
| NSCLC | [116,117] | |||
| WT1 vaccine (1) | glioma | Peptide | A24:02 | [118] |
| PaC | pDC | A24:02 | [119] | |
| BC, OC, GC | [120] | |||
| WT1 peptide vaccine | AML MDS | Peptide | A02 | [121] |
| WT1 peptides | PaC | Peptide | A02:01 or A24:02 | [122] |
| WT1 vaccine (2) | mPM | Peptide | no | [123] |
| Parameters | Predicted Percentage of Subjects in the MP: | Analysis Performed | ||
|---|---|---|---|---|
| Number of Vaccine-Specific Epitopes | Number of Vaccine-Specific Proteins | Threshold of HLA Alleles Binding the Epitope (n) | ||
| In Silico IRR (n × HLA) | ≥ 1 | ≥ 1 | ≥ 1, ≥ 2, ≥ 3 or ≥ 4 | Measured and predicted IRR correlations |
| In Silico multi-epitope IRR (n × HLA) | ≥ 2 | ≥ 1 | ≥ 1, ≥ 2, ≥ 3 or ≥ 4 | Measured and predicted CRR correlations |
| In Silico multi-Ag IRR (n × HLA) | ≥ 2 | ≥ 2 | ≥ 1, ≥ 2, ≥ 3 or ≥ 4 | Measured and predicted CRR correlations |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lőrincz, O.; Tóth, J.; Molnár, L.; Miklós, I.; Pántya, K.; Megyesi, M.; Somogyi, E.; Csiszovszki, Z.; Tőke, E.R. In Silico Model Estimates the Clinical Trial Outcome of Cancer Vaccines. Cells 2021, 10, 3048. https://doi.org/10.3390/cells10113048
Lőrincz O, Tóth J, Molnár L, Miklós I, Pántya K, Megyesi M, Somogyi E, Csiszovszki Z, Tőke ER. In Silico Model Estimates the Clinical Trial Outcome of Cancer Vaccines. Cells. 2021; 10(11):3048. https://doi.org/10.3390/cells10113048
Chicago/Turabian StyleLőrincz, Orsolya, József Tóth, Levente Molnár, István Miklós, Kata Pántya, Mónika Megyesi, Eszter Somogyi, Zsolt Csiszovszki, and Enikő R. Tőke. 2021. "In Silico Model Estimates the Clinical Trial Outcome of Cancer Vaccines" Cells 10, no. 11: 3048. https://doi.org/10.3390/cells10113048