
The Cellular Innate Immune Response of the Invasive Pest Insect Drosophila suzukii against Pseudomonas entomophila Involves the Release of Extracellular Traps

 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Drosophila suzukii Fly Stocks
2.2. Hemocyte Collection and Identification
2.3. Cell Viability Assay
2.4. Immunofluorescence Staining
2.5. Detection of Plasmatocyte Phagocytosis and Extracellular Traps
2.6. Scanning Electron Microscopy (SEM)
3. Results
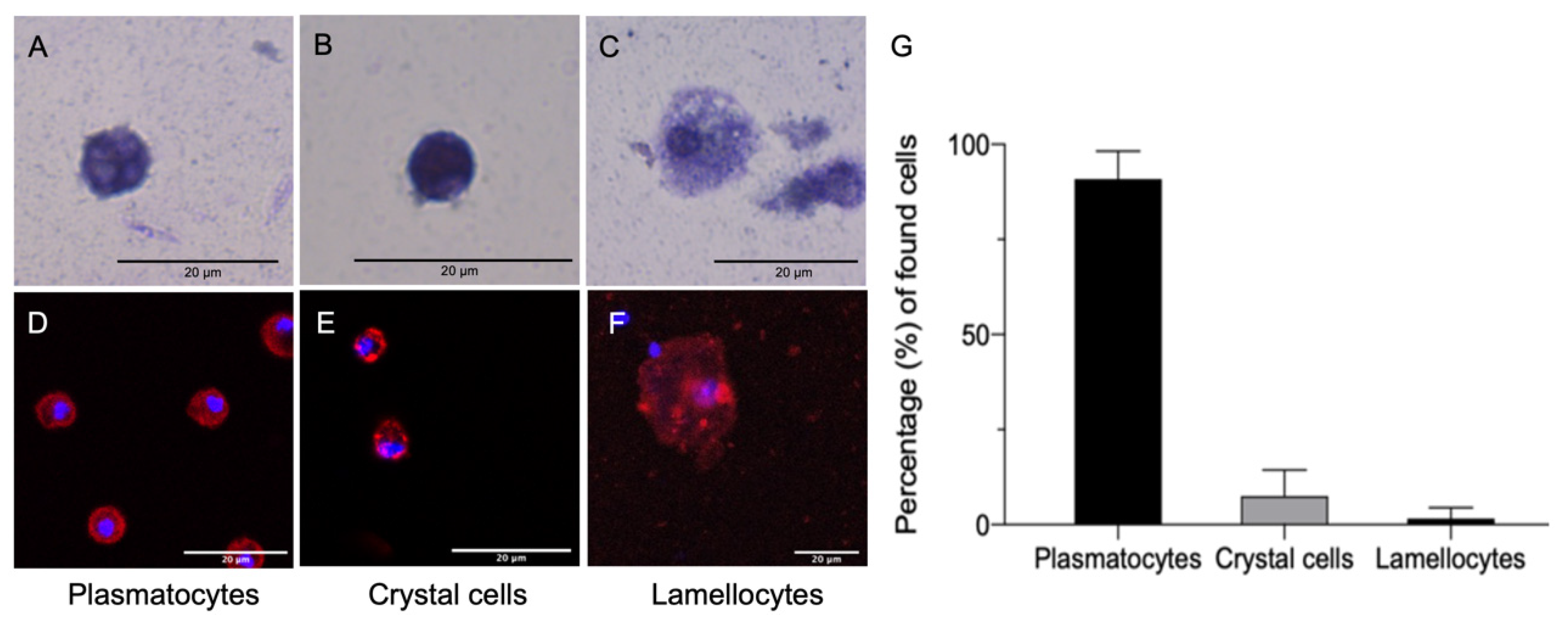
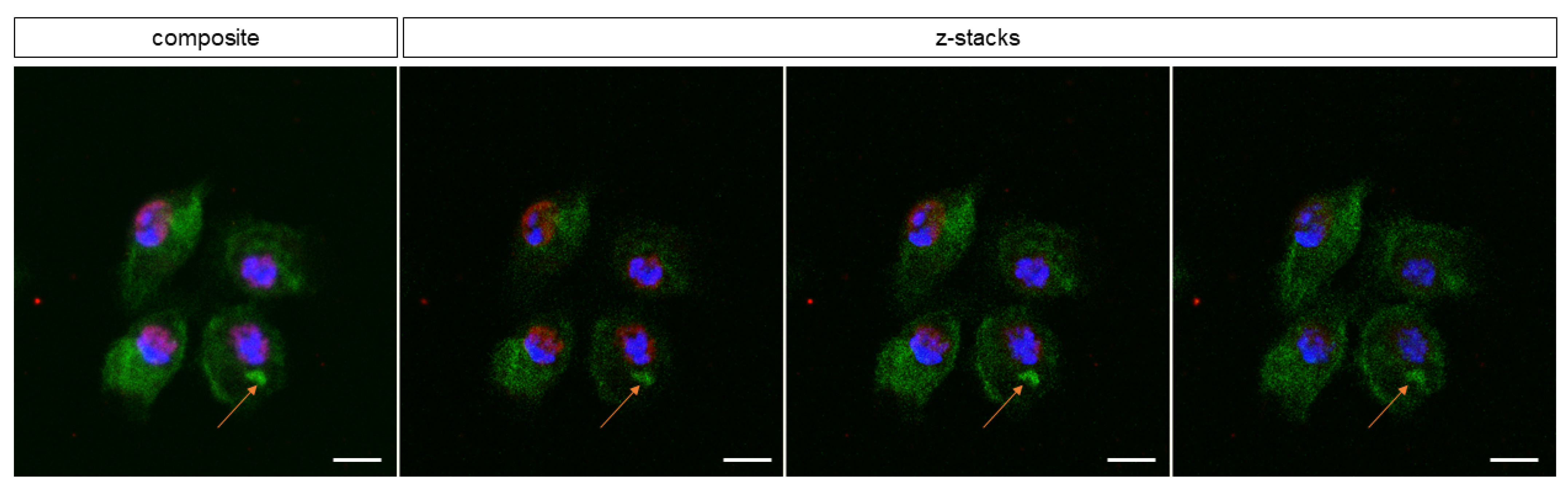
3.1. Characterization of D. suzukii Larval Hemocytes
3.2. Viability of Freshly Isolated D. suzukii Larval Plasmatocytes
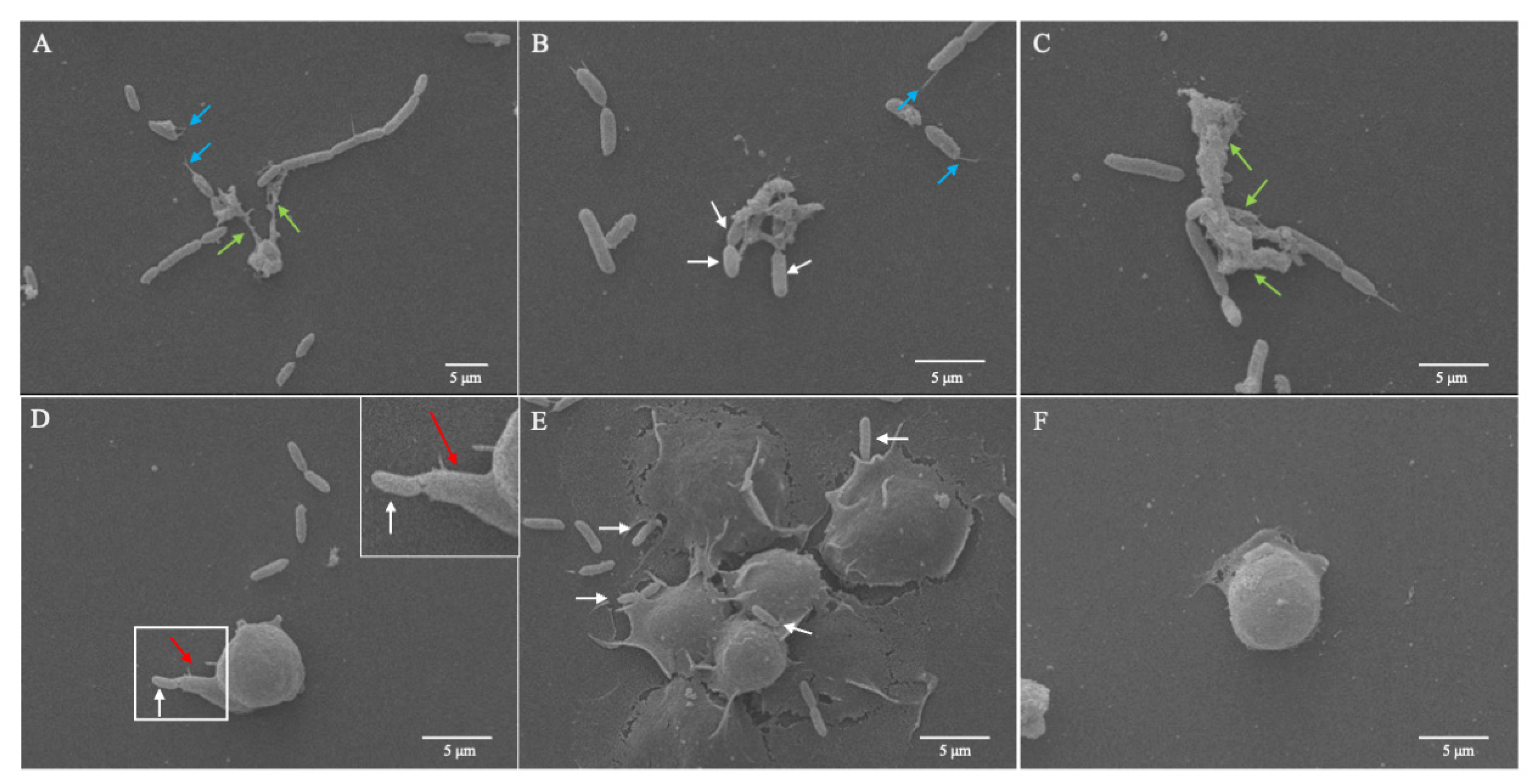
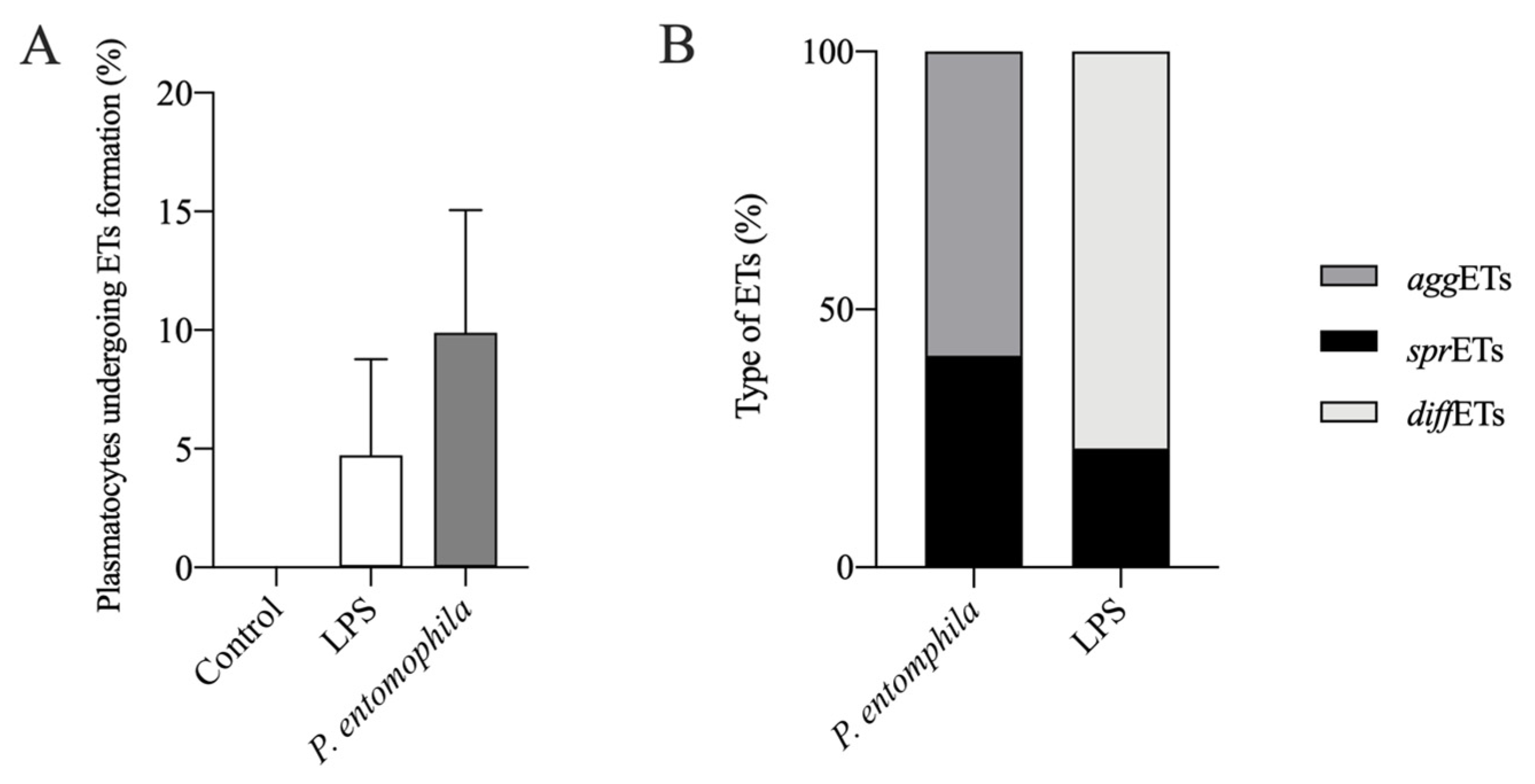
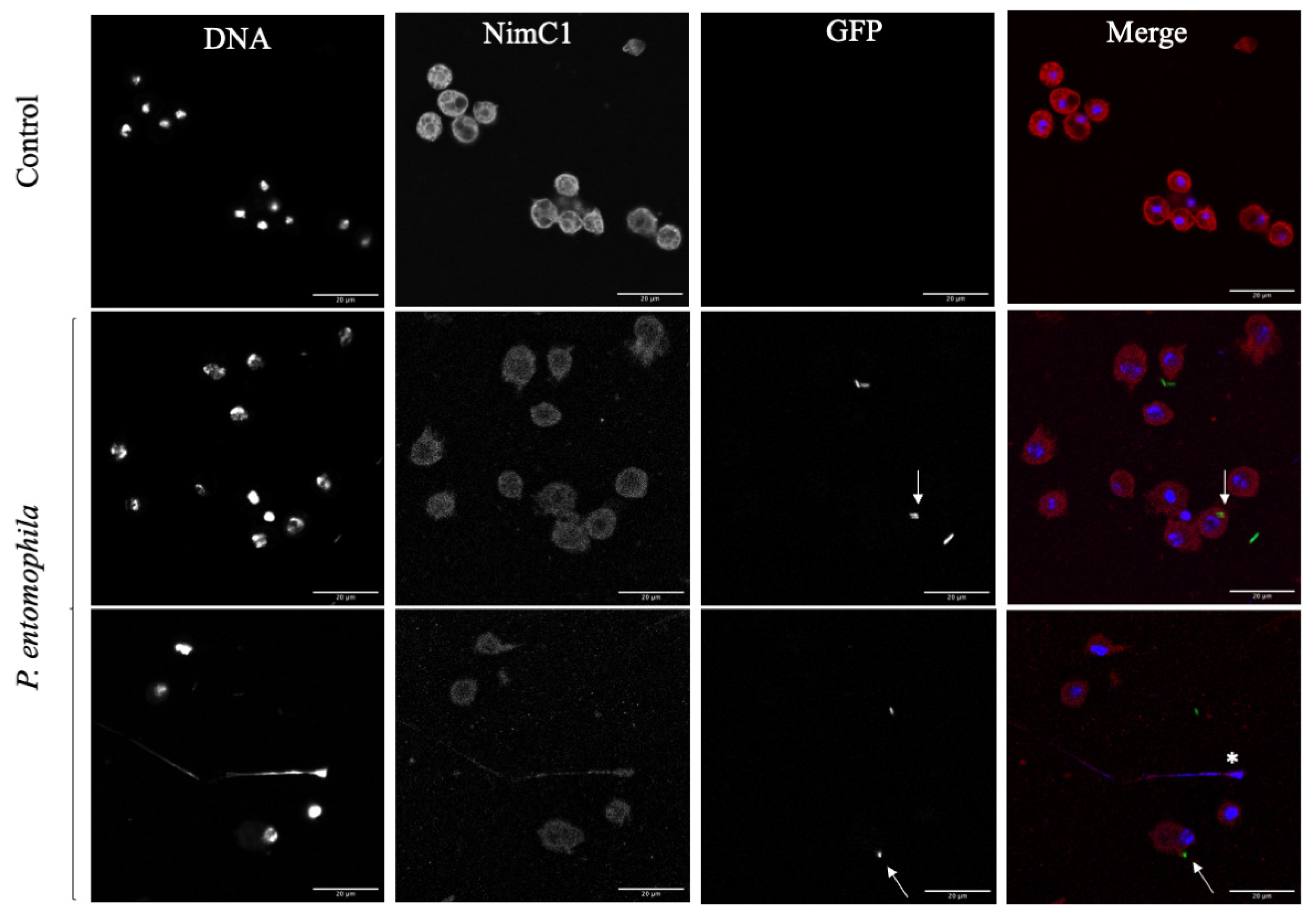
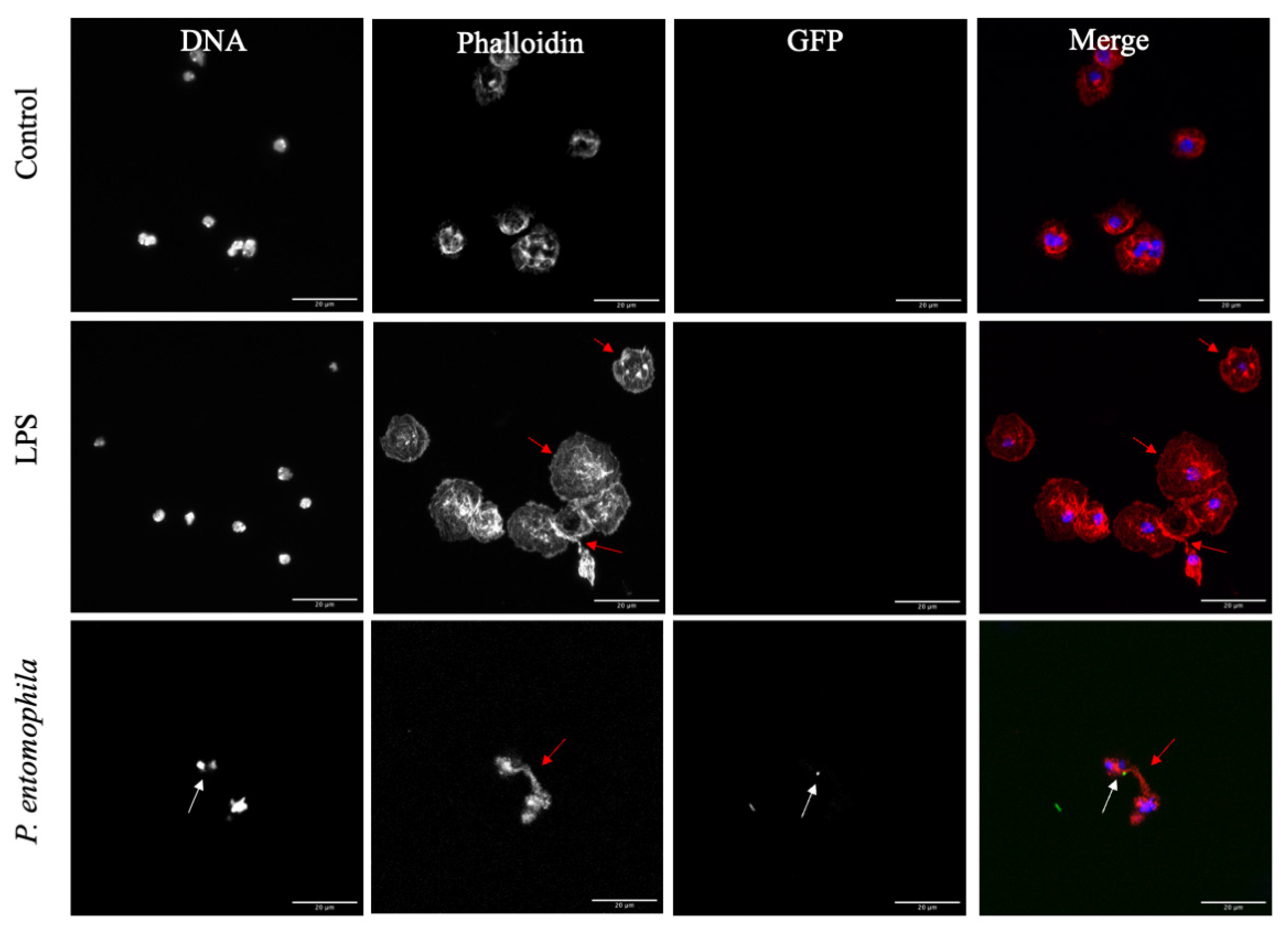
3.3. Response of D. suzukii L3 Plasmatocytes to P. entomophila
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Calabria, G.; Máca, J.; Serra, L.; Pascual, M. First Records of the Potential Pest Species Drosophila suzukii (Diptera: Drosophilidae) in Europe. J. Appl. Entomol. 2012, 136, 139–147. [Google Scholar] [CrossRef]
- Walsh, D.B.; Bolda, M.P.; Goodhue, R.E.; Dreves, A.J.; Lee, J.; Bruck, D.J.; Walton, V.M.; O’Neal, S.D.; Zalom, F.G. Drosophila suzukii (Diptera: Drosophilidae): Invasive Pest of Ripening Soft Fruit Expanding Its Geographic Range and Damage Potential. J. Integr. Pest Manag. 2011, 2, G1–G7. [Google Scholar] [CrossRef]
- Andreazza, F.; Bernardi, D.; Dos Santos, R.S.S.; Garcia, F.R.M.; Oliveira, E.E.; Botton, M.; Nava, D.E. Drosophila suzukii in Southern Neotropical Region: Current Status and Future Perspectives. Neotrop. Entomol. 2017, 46, 591–605. [Google Scholar] [CrossRef] [PubMed]
- Ørsted, I.V.; Ørsted, M. Species Distribution Models of the Spotted Wing Drosophila (Drosophila suzukii, Diptera: Drosophilidae) in Its Native and Invasive Range Reveal an Ecological Niche Shift. J. Appl. Ecol. 2019, 56, 423–435. [Google Scholar] [CrossRef]
- Atallah, J.; Teixeira, L.; Salazar, R.; Zaragoza, G.; Kopp, A. The Making of a Pest: The Evolution of a Fruit-Penetrating Ovipositor in Drosophila suzukii and Related Species. Proc. R. Soc. B Biol. Sci. 2014, 281, 20132840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tochen, S.; Dalton, D.T.; Wiman, N.; Hamm, C.; Shearer, P.W.; Walton, V.M. Temperature-Related Development and Population Parameters for Drosophila suzukii (Diptera: Drosophilidae) on Cherry and Blueberry. Environ. Entomol. 2014, 43, 501–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiman, N.G.; Walton, V.M.; Dalton, D.T.; Anfora, G.; Burrack, H.J.; Chiu, J.C.; Daane, K.M.; Grassi, A.; Miller, B.; Tochen, S.; et al. Integrating Temperature-Dependent Life Table Data into a Matrix Projection Model for Drosophila suzukii Population Estimation. PLoS ONE 2014, 9, e106909. [Google Scholar] [CrossRef]
- Kacsoh, B.Z.; Schlenke, T.A. High Hemocyte Load Is Associated with Increased Resistance against Parasitoids in Drosophila suzukii, a Relative of D. melanogaster. PLoS ONE 2012, 7, e34721. [Google Scholar] [CrossRef]
- Poyet, M.; Havard, S.; Prevost, G.; Chabrerie, O.; Doury, G.; Gibert, P.; Eslin, P. Resistance of Drosophila suzukii to the Larval Parasitoids Leptopilina heterotoma and Asobara japonica Is Related to Haemocyte Load. Physiol. Entomol. 2013, 38, 45–53. [Google Scholar] [CrossRef]
- Pech, L.L.; Strand, M.R. Granular Cells Are Required for Encapsulation of Foreign Targets by Insect Haemocytes. J. Cell Sci. 1996, 109, 2053–2060. [Google Scholar] [CrossRef]
- Schmidt, O.; Theopold, U.; Strand, M. Innate Immunity and Its Evasion and Suppression by Hymenopteran Endoparasitoids. BioEssays 2001, 23, 344–351. [Google Scholar] [CrossRef]
- Traver, D.; Zon, L.I. Walking the Walk: Migration and Other Common Themes in Blood and Vascular Development. Cell 2002, 108, 731–734. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, U.; Girard, J.R.; Goins, L.M.; Spratford, C.M. Drosophila as a Genetic Model for Hematopoiesis. Genetics 2019, 211, 367–417. [Google Scholar] [CrossRef] [Green Version]
- Meister, M.; Lagueux, M. Drosophila Blood Cells. Cell. Microbiol. 2003, 5, 573–580. [Google Scholar] [CrossRef]
- Lebestky, T.; Chang, T.; Hartenstein, V.; Banerjee, U. Specification of Drosophila Hematopoietic Lineage by Conserved Transcription Factors. Science 2000, 288, 146–149. [Google Scholar] [CrossRef] [PubMed]
- Lavine, M.D.; Strand, M.R. Insect Hemocytes and Their Role in Immunity. Insect Biochem. Mol. Biol. 2002, 32, 1295–1309. [Google Scholar] [CrossRef]
- Tepass, U.; Fessler, L.I.; Aziz, A.; Hartenstein, V. Embryonic Origin of Hemocytes and Their Relationship to Cell Death in Drosophila. Development 1994, 120, 1829–1837. [Google Scholar] [CrossRef] [PubMed]
- Franc, N.C.; Heitzler, P.; Ezekowitz, R.A.B.; White, K. Requirement for Croquemort in Phagocytosis of Apoptotic Cells in Drosophila. Science 1999, 284, 1991–1994. [Google Scholar] [CrossRef] [PubMed]
- Müller, U.; Vogel, P.; Alber, G.; Schaub, G.A. The Innate Immune System of Mammals and Insects. Trends Innate Immun. 2008, 15, 21–44. [Google Scholar] [CrossRef]
- Franc, N.C.; Dimarcq, J.L.; Lagueux, M.; Hoffman, J.A.; Ezekowitz, R.A.B. Croquemort, A Novel Drosophila Hemocyte/Macrophage Receptor That Recognizes Apoptotic Cells. Immunity 1996, 4, 431–443. [Google Scholar] [CrossRef] [Green Version]
- Villagra-Blanco, R.; Silva, L.M.R.; Conejeros, I.; Taubert, A.; Hermosilla, C. Pinniped- and Cetacean-Derived ETosis Contributes to Combating Emerging Apicomplexan Parasites (Toxoplasma gondii, Neospora caninum) Circulating in Marine Environments. Biology 2019, 8, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neumann, A.; Brogden, G.; von Köckritz-Blickwede, M. Extracellular Traps: An Ancient Weapon of Multiple Kingdoms. Biology 2020, 9, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nathan, C. Neutrophils and Immunity: Challenges and Opportunities. Nat. Rev. Immunol. 2006, 6, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Urban, C.F.; Ermert, D.; Schmid, M.; Abu-Abed, U.; Goosmann, C.; Nacken, W.; Brinkmann, V.; Jungblut, P.R.; Zychlinsky, A. Neutrophil Extracellular Traps Contain Calprotectin, a Cytosolic Protein Complex Involved in Host Defense against Candida albicans. PLoS Pathog. 2009, 5, e1000639. [Google Scholar] [CrossRef] [Green Version]
- Parker, H.; Albrett, A.M.; Kettle, A.J.; Winterbourn, C.C. Myeloperoxidase Associated with Neutrophil Extracellular Traps Is Active and Mediates Bacterial Killing in the Presence of Hydrogen Peroxide. J. Leukoc. Biol. 2012, 91, 369–376. [Google Scholar] [CrossRef]
- Webster, S.J.; Daigneault, M.; Bewley, M.A.; Preston, J.A.; Marriott, H.M.; Walmsley, S.R.; Read, R.C.; Whyte, M.K.B.; Dockrell, D.H. Distinct Cell Death Programs in Monocytes Regulate Innate Responses Following Challenge with Common Causes of Invasive Bacterial Disease. J. Immunol. 2010, 185, 2968–2979. [Google Scholar] [CrossRef] [Green Version]
- Bartneck, M.; Keul, H.A.; Zwadlo-Klarwasser, G.; Groll, J. Phagocytosis Independent Extracellular Nanoparticle Clearance by Human Immune Cells. Nano Lett. 2010, 10, 59–63. [Google Scholar] [CrossRef]
- Palić, D.; Ostojić, J.; Andreasen, C.B.; Roth, J.A. Fish Cast NETs: Neutrophil Extracellular Traps Are Released from Fish Neutrophils. Dev. Comp. Immunol. 2007, 31, 805–816. [Google Scholar] [CrossRef]
- Lange, M.K.; Penagos-Tabares, F.; Muñoz-Caro, T.; Gärtner, U.; Mejer, H.; Schaper, R.; Hermosilla, C.; Taubert, A. Gastropod-Derived Haemocyte Extracellular Traps Entrap Metastrongyloid Larval Stages of Angiostrongylus vasorum, Aelurostrongylus abstrusus and Troglostrongylus brevior. Parasites Vectors 2017, 10, 50. [Google Scholar] [CrossRef] [Green Version]
- Ng, T.H.; Chang, S.-H.; Wu, M.-H.; Wang, H.-C. Shrimp Hemocytes Release Extracellular Traps That Kill Bacteria. Dev. Comp. Immunol. 2013, 41, 644–651. [Google Scholar] [CrossRef]
- Silva, L.M.R.; Muñoz-Caro, T.; Burgos, R.A.; Hidalgo, M.A.; Taubert, A.; Hermosilla, C. Far beyond Phagocytosis: Phagocyte-Derived Extracellular Traps Act Efficiently against Protozoan Parasites In Vitro and In Vivo. Mediat. Inflamm. 2016, 2016, 5898074. [Google Scholar] [CrossRef] [Green Version]
- Altincicek, B.; Stötzel, S.; Wygrecka, M.; Preissner, K.T.; Vilcinskas, A. Host-Derived Extracellular Nucleic Acids Enhance Innate Immune Responses, Induce Coagulation, and Prolong Survival upon Infection in Insects. J. Immunol. 2008, 181, 2705–2712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vodovar, N.; Vinals, M.; Liehl, P.; Basset, A.; Degrouard, J.; Spellman, P.; Boccard, F.; Lemaitre, B. Drosophila Host Defense after Oral Infection by an Entomopathogenic Pseudomonas Species. Proc. Natl. Acad. Sci. USA 2005, 102, 11414–11419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merkling, S.H.; van Rij, R.P. Analysis of Resistance and Tolerance to Virus Infection in Drosophila. Nat. Protoc. 2015, 10, 1084–1097. [Google Scholar] [CrossRef] [PubMed]
- Medd, N.C.; Fellous, S.; Waldron, F.M.; Xuéreb, A.; Nakai, M.; Cross, J.V.; Obbard, D.J. The Virome of Drosophila suzukii, an Invasive Pest of Soft Fruit. Virus Evol. 2018, 4, vey009. [Google Scholar] [CrossRef]
- Tracy, C.; Krämer, H. Isolation and Infection of Drosophila Primary Hemocytes. Bio-Protoc. 2017, 7, e2300. [Google Scholar] [CrossRef] [Green Version]
- Sandoz, P.A.; Tremblay, C.; Equis, S.; Pop, S.; Pollaro, L.; Cotte, Y.; van der Goot, F.G.; Frechin, M. Label Free 3D Analysis of Organelles in Living Cells by Refractive Index Shows Pre-Mitotic Organelle Spinning in Mammalian Stem Cells. bioRxiv 2018, 407239. [Google Scholar] [CrossRef] [Green Version]
- Velásquez, Z.D.; López-Osorio, S.; Mazurek, S.; Hermosilla, C.; Taubert, A. Eimeria Bovis Macromeront Formation Induces Glycolytic Responses and Mitochondrial Changes in Primary Host Endothelial Cells. Front. Cell. Infect. Microbiol. 2021, 11, 635. [Google Scholar] [CrossRef]
- Larrazabal, C.; Hermosilla, C.; Taubert, A.; Conejeros, I. 3D Holotomographic Monitoring of Ca++ Dynamics during Ionophore-Induced Neospora caninum Tachyzoite Egress from Primary Bovine Host Endothelial Cells. Parasitol. Res. 2021. [Google Scholar] [CrossRef]
- Kurucz, E.; Váczi, B.; Márkus, R.; Laurinyecz, B.; Vilmos, P.; Zsámboki, J.; Csorba, K.; Gateff, E.; Hultmark, D.; Andó, I. Definition of Drosophila Hemocyte Subsets by Cell-Type Specific Antigens. Acta Biol. Hung. 2007, 58, 95–111. [Google Scholar] [CrossRef] [Green Version]
- Mulet, M.; Gomila, M.; Lemaitre, B.; Lalucat, J.; García-Valdés, E. Taxonomic Characterisation of Pseudomonas Strain L48 and Formal Proposal of Pseudomonas entomophila Sp. Nov. Syst. Appl. Microbiol. 2012, 35, 145–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nascimento, M.T.C.; Silva, K.P.; Garcia, M.C.F.; Medeiros, M.N.; Machado, E.A.; Nascimento, S.B.; Saraiva, E.M. DNA Extracellular Traps Are Part of the Immune Repertoire of Periplaneta americana. Dev. Comp. Immunol. 2018, 84, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Caro, T.; Rubio, R.M.C.; Silva, L.M.R.; Magdowski, G.; Gärtner, U.; McNeilly, T.N.; Taubert, A.; Hermosilla, C. Leucocyte-Derived Extracellular Trap Formation Significantly Contributes to Haemonchus contortus Larval Entrapment. Parasites Vectors 2015, 8, 607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schauer, C.; Janko, C.; Munoz, L.E.; Zhao, Y.; Kienhöfer, D.; Frey, B.; Lell, M.; Manger, B.; Rech, J.; Naschberger, E.; et al. Aggregated Neutrophil Extracellular Traps Limit Inflammation by Degrading Cytokines and Chemokines. Nat. Med. 2014, 20, 511–517. [Google Scholar] [CrossRef]
- Hakkim, A.; Fuchs, T.A.; Martinez, N.E.; Hess, S.; Prinz, H.; Zychlinsky, A.; Waldmann, H. Activation of the Raf-MEK-ERK Pathway Is Required for Neutrophil Extracellular Trap Formation. Nat. Chem. Biol. 2011, 7, 75–77. [Google Scholar] [CrossRef]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil Extracellular Traps Kill Bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef]
- Zhou, E.; Silva, L.; Conejeros, I.; Velásquez, Z.; Hirz, M.; Gärtner, U.; Jacquiet, P.; Taubert, A.; Hermosilla, C. Besnoitia besnoiti bradyzoite stages induce suicidal- and rapid vital-NETosis. Parasitology 2020, 147, 401–409. [Google Scholar] [CrossRef]
- Beckert, A.; Wiesner, J.; Baumann, A.; Pöppel, A.-K.; Vogel, H.; Vilcinskas, A. Two C-Type Lysozymes Boost the Innate Immune System of the Invasive Ladybird Harmonia axyridis. Dev. Comp. Immunol. 2015, 49, 303–312. [Google Scholar] [CrossRef]
- Bidla, G.; Lindgren, M.; Theopold, U.; Dushay, M.S. Hemolymph Coagulation and Phenoloxidase in Drosophila Larvae. Dev. Comp. Immunol. 2005, 29, 669–679. [Google Scholar] [CrossRef]
- Hiebert, N.; Carrau, T.; Bartling, M.; Vilcinskas, A.; Lee, K.-Z. Identification of Entomopathogenic Bacteria Associated with the Invasive Pest Drosophila suzukii in Infested Areas of Germany. J. Invertebr. Pathol. 2020, 173, 107389. [Google Scholar] [CrossRef]
- Dieppois, G.; Opota, O.; Lalucat, J.; Lemaitre, B. Pseudomonas entomophila: A Versatile Bacterium with Entomopathogenic Properties. In Pseudomonas: New Aspects of Pseudomonas Biology; Ramos, J.-L., Goldberg, J.B., Filloux, A., Eds.; Springer: Dordrecht, The Netherlands, 2015; Volume 7, pp. 25–49. ISBN 978-94-017-9555-5. [Google Scholar]
- Johansson, K.C.; Metzendorf, C.; Söderhäll, K. Microarray Analysis of Immune Challenged Drosophila Hemocytes. Exp. Cell Res. 2005, 305, 145–155. [Google Scholar] [CrossRef]
- Frangou, E.; Vassilopoulos, D.; Boletis, J.; Boumpas, D.T. An Emerging Role of Neutrophils and NETosis in Chronic Inflammation and Fibrosis in Systemic Lupus Erythematosus (SLE) and ANCA-Associated Vasculitides (AAV): Implications for the Pathogenesis and Treatment. Autoimmun. Rev. 2019, 18, 751–760. [Google Scholar] [CrossRef] [PubMed]
- Mondotte, J.A.; Gausson, V.; Frangeul, L.; Blanc, H.; Lambrechts, L.; Saleh, M.-C. Immune Priming and Clearance of Orally Acquired RNA Viruses in Drosophila. Nat. Microbiol. 2018, 3, 1394–1403. [Google Scholar] [CrossRef] [PubMed]
- Mondotte, J.A.; Gausson, V.; Frangeul, L.; Suzuki, Y.; Vazeille, M.; Mongelli, V.; Blanc, H.; Failloux, A.-B.; Saleh, M.-C. Evidence For Long-Lasting Transgenerational Antiviral Immunity in Insects. Cell Rep. 2020, 33, 108506. [Google Scholar] [CrossRef] [PubMed]
- Cooper, D.; Eleftherianos, I. Memory and Specificity in the Insect Immune System: Current Perspectives and Future Challenges. Front. Immunol. 2017, 8, 539. [Google Scholar] [CrossRef]
- Pieterse, E.; Rother, N.; Yanginlar, C.; Hilbrands, L.B.; van der Vlag, J. Neutrophils Discriminate between Lipopolysaccharides of Different Bacterial Sources and Selectively Release Neutrophil Extracellular Traps. Front. Immunol. 2016, 7, 484. [Google Scholar] [CrossRef] [Green Version]
- Gegner, T.; Schmidtberg, H.; Vogel, H.; Vilcinskas, A. Population-Specific Expression of Antimicrobial Peptides Conferring Pathogen Resistance in the Invasive Ladybird Harmonia axyridis. Sci. Rep. 2018, 8, 3600. [Google Scholar] [CrossRef]
- Vilcinskas, A.; Stoecker, K.; Schmidtberg, H.; Röhrich, C.R.; Vogel, H. Invasive Harlequin Ladybird Carries Biological Weapons against Native Competitors. Science 2013, 340, 862–863. [Google Scholar] [CrossRef]
- Röhrich, C.R.; Ngwa, C.J.; Wiesner, J.; Schmidtberg, H.; Degenkolb, T.; Kollewe, C.; Fischer, R.; Pradel, G.; Vilcinskas, A. Harmonine, a Defence Compound from the Harlequin Ladybird, Inhibits Mycobacterial Growth and Demonstrates Multi-Stage Antimalarial Activity. Biol. Lett. 2012, 8, 308–311. [Google Scholar] [CrossRef]
- Lee, K.A.; Klasing, K.C. A Role for Immunology in Invasion Biology. Trends Ecol. Evol. 2004, 19, 523–529. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carrau, T.; Thümecke, S.; Silva, L.M.R.; Perez-Bravo, D.; Gärtner, U.; Taubert, A.; Hermosilla, C.; Vilcinskas, A.; Lee, K.-Z. The Cellular Innate Immune Response of the Invasive Pest Insect Drosophila suzukii against Pseudomonas entomophila Involves the Release of Extracellular Traps. Cells 2021, 10, 3320. https://doi.org/10.3390/cells10123320
Carrau T, Thümecke S, Silva LMR, Perez-Bravo D, Gärtner U, Taubert A, Hermosilla C, Vilcinskas A, Lee K-Z. The Cellular Innate Immune Response of the Invasive Pest Insect Drosophila suzukii against Pseudomonas entomophila Involves the Release of Extracellular Traps. Cells. 2021; 10(12):3320. https://doi.org/10.3390/cells10123320
Chicago/Turabian StyleCarrau, Tessa, Susanne Thümecke, Liliana M. R. Silva, David Perez-Bravo, Ulrich Gärtner, Anja Taubert, Carlos Hermosilla, Andreas Vilcinskas, and Kwang-Zin Lee. 2021. "The Cellular Innate Immune Response of the Invasive Pest Insect Drosophila suzukii against Pseudomonas entomophila Involves the Release of Extracellular Traps" Cells 10, no. 12: 3320. https://doi.org/10.3390/cells10123320