
Extracellular Matrix Oxidised by the Granulocyte Oxidants Hypochlorous and Hypobromous Acid Reduces Lung Fibroblast Adhesion and Proliferation In Vitro

 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Primary Cell Isolation
2.2. Cell-Derived ECM
2.3. Preparation of HOCl Stocks
2.4. Preparation of HOBr Stocks
2.5. HOCl and HOBr Treatment of Cell-Derived ECM
2.6. Cell Attachment Assay
2.7. Proliferation Assay
2.8. ELISA
2.9. Data Analysis and Statistics
3. Results
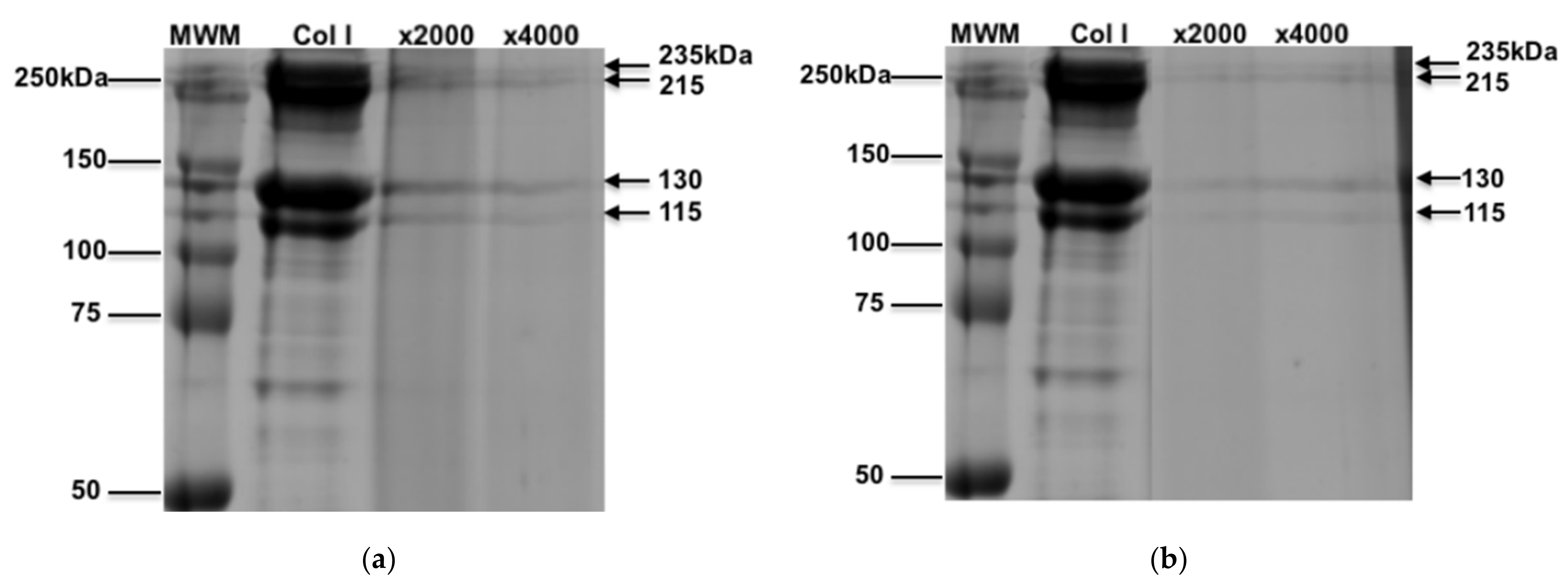
3.1. Characterization of Oxidized Collagen
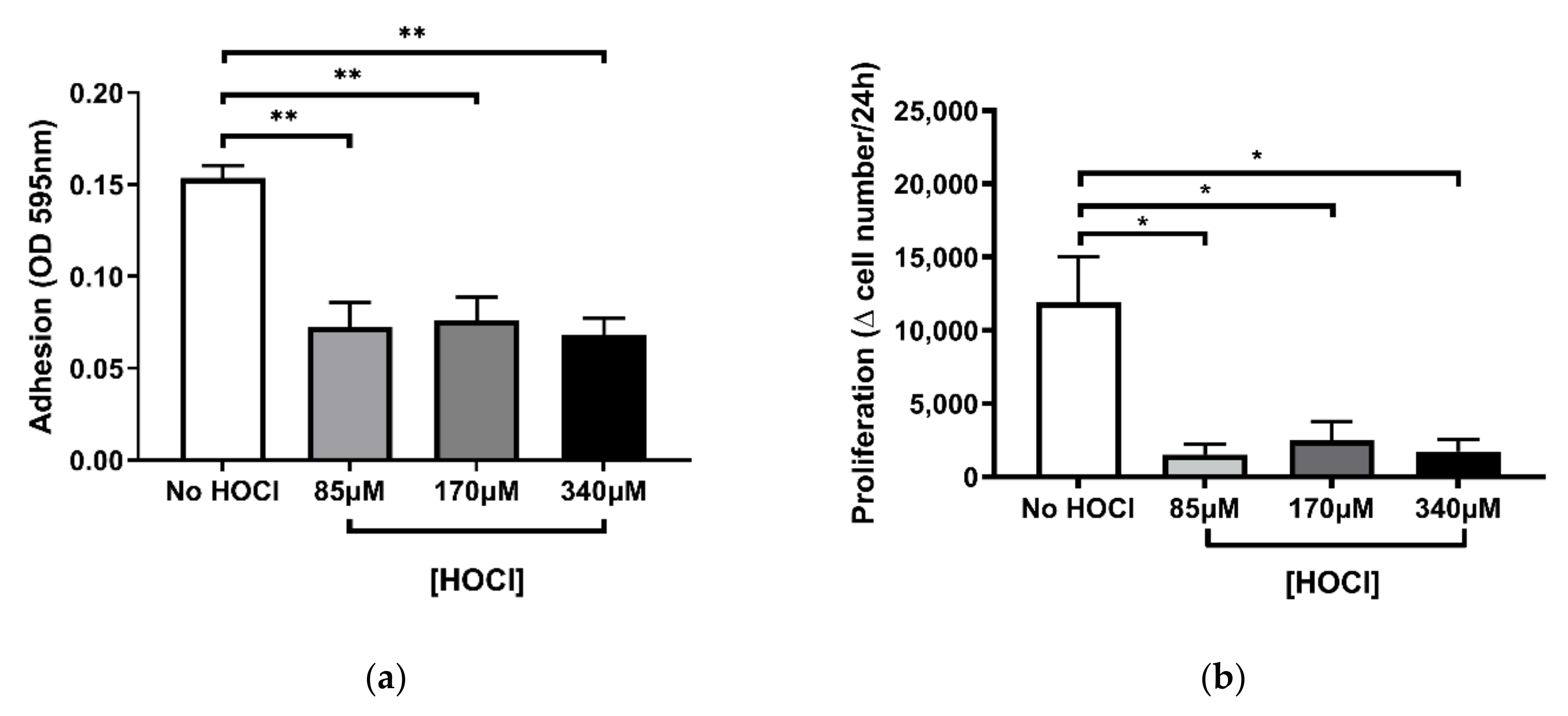
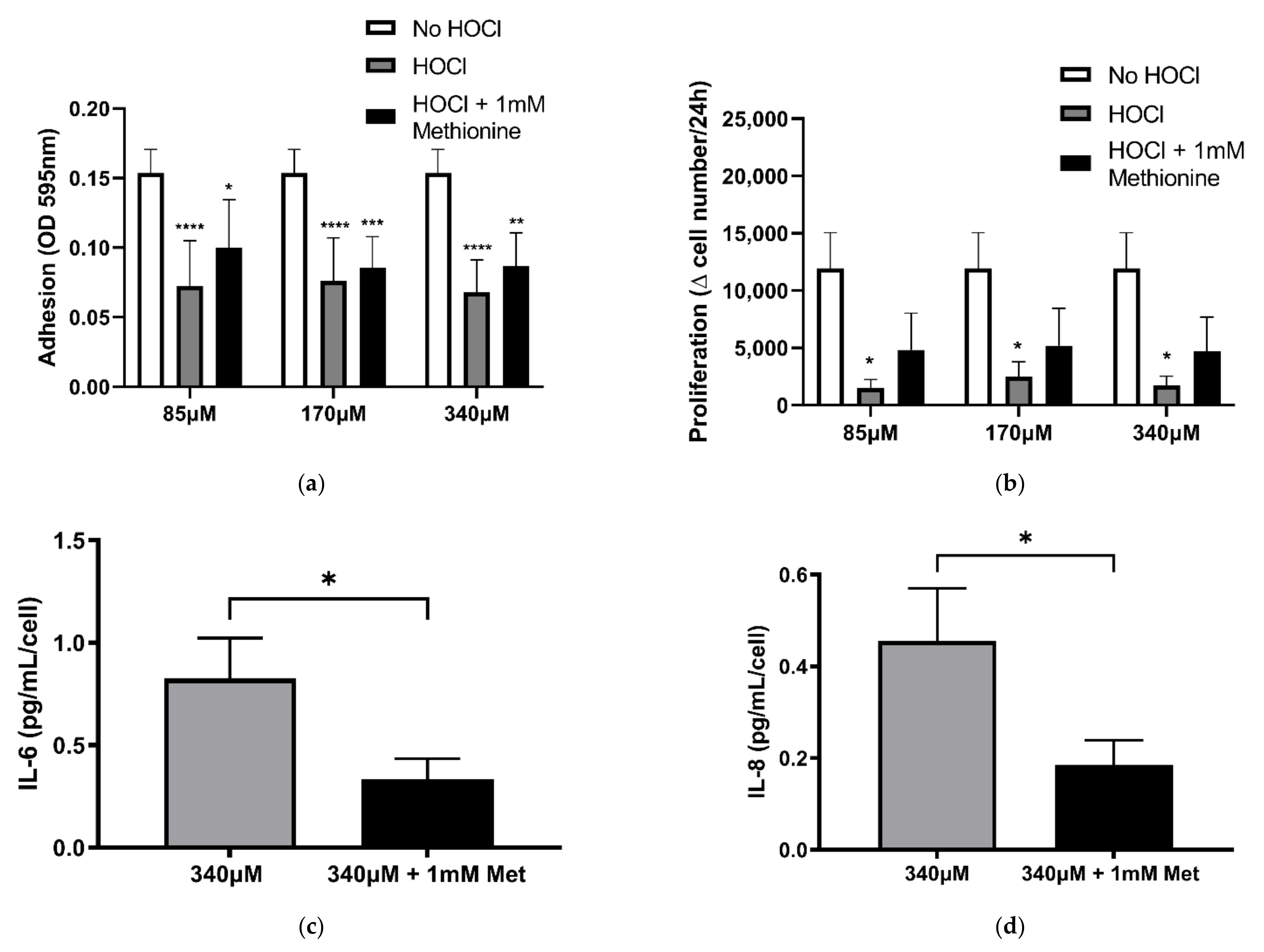
3.2. Altered Fibroblast Function Following HOCl-Induced Oxidation of the Cell-Derived ECM
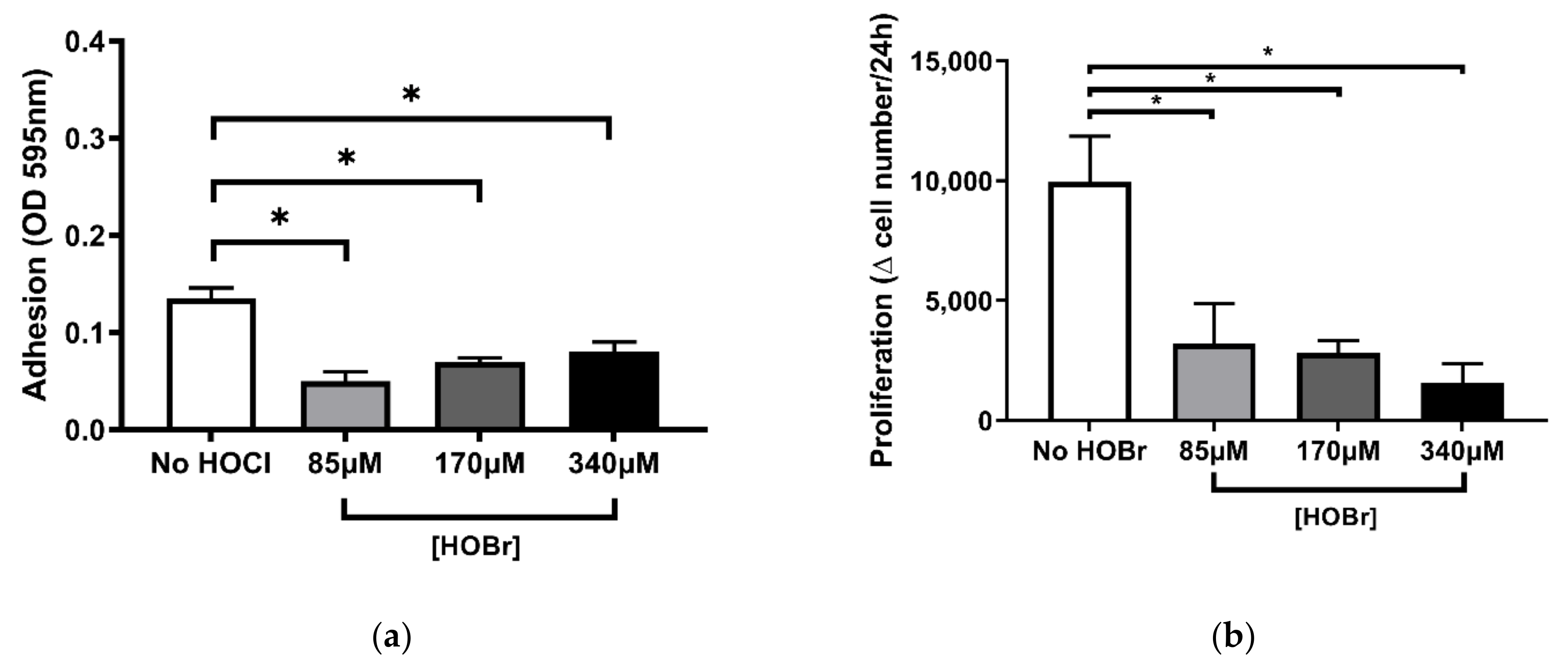
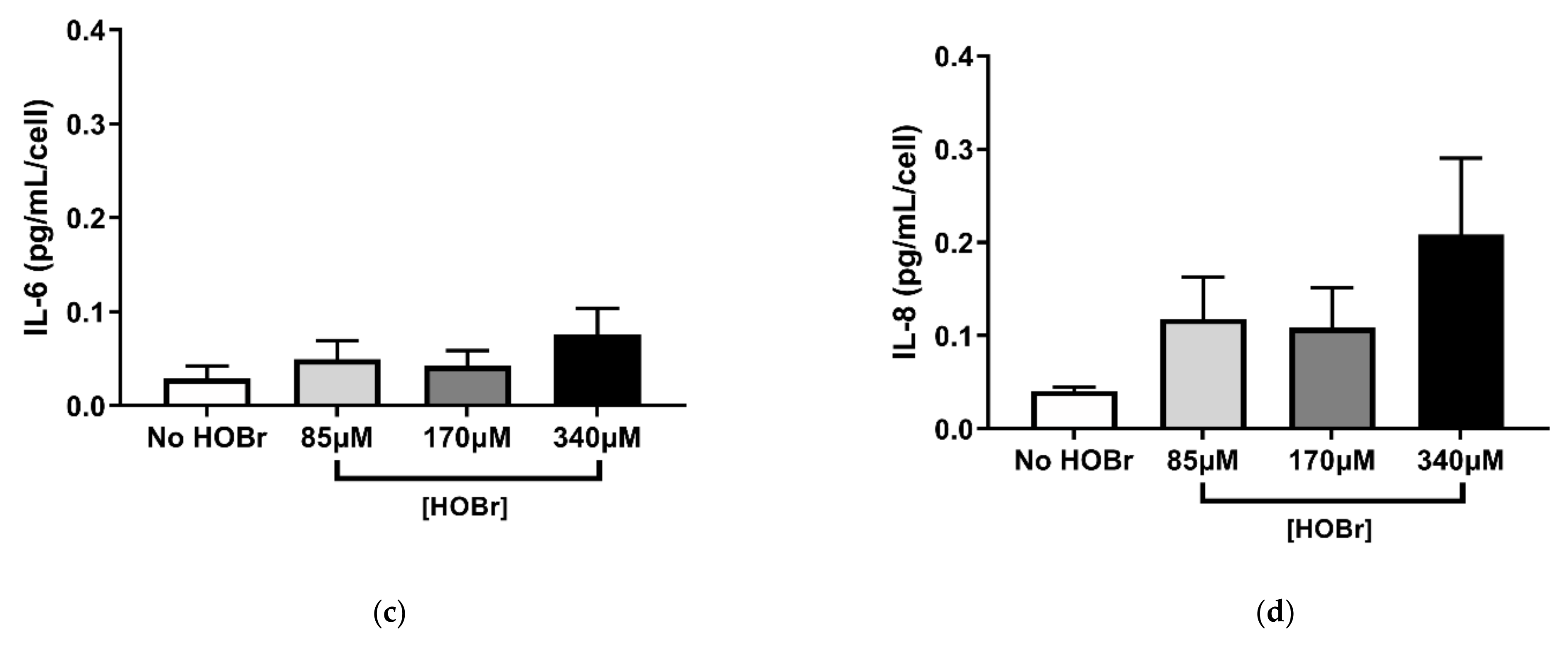
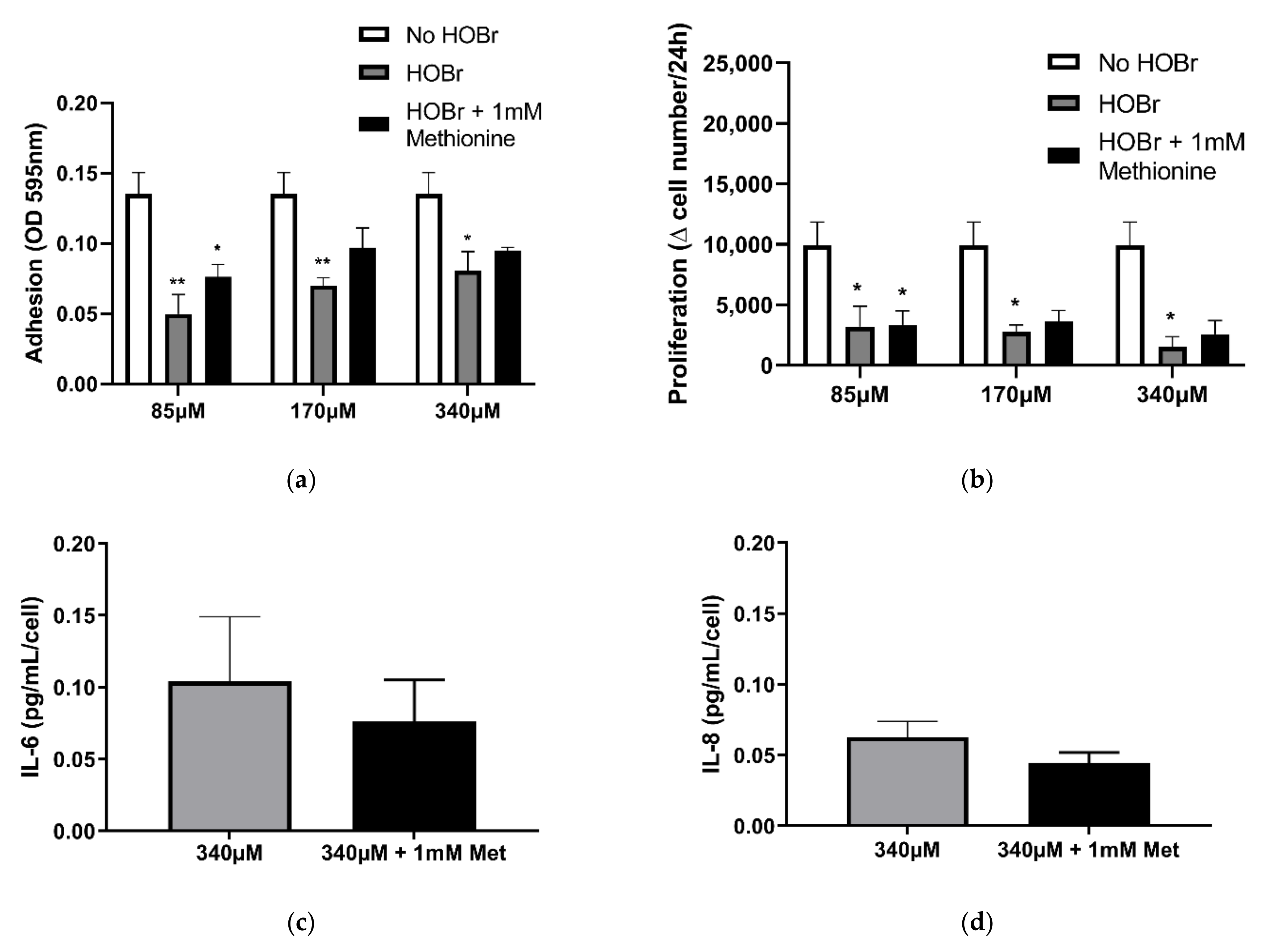
3.3. HOBr-Induced Oxidation of the Cell-Derived ECM on Fibroblast Function Compared to HOCl Treatment
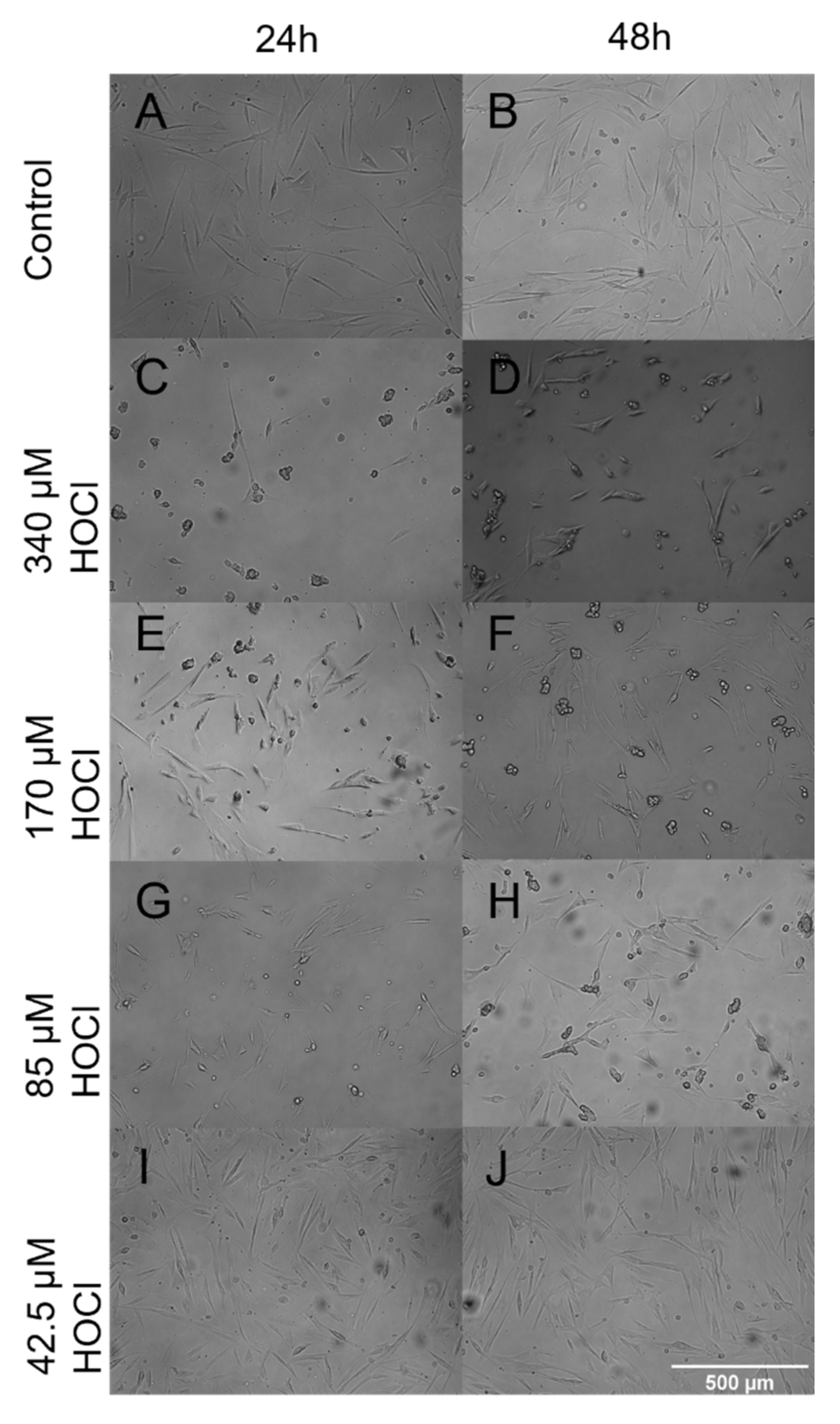
3.4. Fibroblast Morphology When Grown on Oxidized ECM
3.5. Involvement of Haloamines in the Modification of ECM on Fibroblast Function
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Shishido, N.; Nakamura, S.; Nakamura, M. Dissociation of DNA double strand by hypohalous acids. Redox Rep. 2000, 5, 243–247. [Google Scholar] [CrossRef] [PubMed]
- Colombo, G.; Clerici, M.; Altomare, A.; Rusconi, F.; Giustarini, D.; Portinaro, N.; Garavaglia, M.L.; Rossi, R.; Dalle-Donne, I.; Milzani, A. Thiol oxidation and di-tyrosine formation in human plasma proteins induced by inflammatory concentrations of hypochlorous acid. J. Proteom. 2017, 152, 22–32. [Google Scholar] [CrossRef] [PubMed]
- Tatsumi, T.; Fliss, H. Hypochlorous acid and chloramines increase endothelial permeability: Possible involvement of cellular zinc. Am. J. Physiol. Content 1994, 267, H1597–H1607. [Google Scholar] [CrossRef] [PubMed]
- Ge, Q.; Chen, L.; Jaffar, J.; Argraves, W.S.; Twal, W.O.; Hansbro, P.; Black, J.L.; Burgess, J.; Oliver, B. Fibulin1C peptide induces cell attachment and extracellular matrix deposition in lung fibroblasts. Sci. Rep. 2015, 5, 9496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kennett, E.C.; Chuang, C.Y.; Degendorfer, G.; Whitelock, J.M.; Davies, M.J. Mechanisms and consequences of oxidative damage to extracellular matrix. Biochem. Soc. Trans. 2011, 39, 1279–1287. [Google Scholar] [CrossRef] [PubMed]
- Midwinter, R.G.; Cheah, F.C.; Moskovitz, J.; Vissers, M.C.; Winterbourn, C.C. IκB is a sensitive target for oxidation by cell-permeable chloramines: Inhibition of NF-κB activity by glycine chloramine through methionine oxidation. Biochem. J. 2006, 396, 71–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bathish, B.; Paumann-Page, M.; Paton, L.N.; Kettle, A.J.; Winterbourn, C.C. Peroxidasin mediates bromination of tyrosine residues in the extracellular matrix. J. Biol. Chem. 2020, 295, 12697–12705. [Google Scholar] [CrossRef]
- Nybo, T.; Dieterich, S.; Gamon, L.; Chuang, C.Y.; Hammer, A.; Hoefler, G.; Malle, E.; Rogowska-Wrzesinska, A.; Davies, M.J. Chlorination and oxidation of the extracellular matrix protein laminin and basement membrane extracts by hypochlorous acid and myeloperoxidase. Redox Biol. 2019, 20, 496–513. [Google Scholar] [CrossRef]
- Badylak, S.F. The extracellular matrix as a scaffold for tissue reconstruction. Semin. Cell Dev. Biol. 2002, 13, 377–383. [Google Scholar] [CrossRef]
- Frantz, C.; Stewart, K.M.; Weaver, V.M. The extracellular matrix at a glance. J. Cell Sci. 2010, 123, 4195–4200. [Google Scholar] [CrossRef] [Green Version]
- Lu, P.; Takai, K.; Weaver, V.M.; Werb, Z. Extracellular Matrix Degradation and Remodeling in Development and Disease. Cold Spring Harb. Perspect. Biol. 2011, 3, a005058. [Google Scholar] [CrossRef]
- Hynes, R.O. The extracellular matrix: Not just pretty fibrils. Science 2009, 326, 1216–1219. [Google Scholar] [CrossRef] [Green Version]
- Hoshiba, T.; Yamaoka, T. CHAPTER 1. Extracellular Matrix Scaffolds for Tissue Engineering and Biological Research. Biomater. Sci. Ser. 2019, 1–14. [Google Scholar] [CrossRef]
- Kusindarta, D.L.; Wihadmadyatami, H. The Role of Extracellular Matrix in Tissue Regeneration. In Tissue Regeneration; IntechOpen: London, UK, 2018; pp. 65–73. [Google Scholar]
- White, E. Lung Extracellular Matrix and Fibroblast Function. Ann. Am. Thorac. Soc. 2015, 12, S30–S33. [Google Scholar] [CrossRef] [Green Version]
- Bon, H.; Hales, P.; Lumb, S.; Holdsworth, G.; Johnson, T.; Qureshi, O.; Twomey, B.M. Spontaneous Extracellular Matrix Accumulation in a Human in vitro Model of Renal Fibrosis Is Mediated by αV Integrins. Nephron 2019, 142, 328–350. [Google Scholar] [CrossRef]
- Sakhneny, L.; Epshtein, A.; Landsman, L. Pericytes contribute to the islet basement membranes to promote beta-cell gene expression. Sci. Rep. 2021, 11, 2378. [Google Scholar] [CrossRef]
- Chuang, C.Y.-N.; Degendorfer, G.; Davies, M.J. Oxidation and modification of extracellular matrix and its role in disease. Free Radic. Res. 2014, 48, 970–989. [Google Scholar] [CrossRef]
- Watson, W.H.; Ritzenthaler, J.D.; Roman, J. Lung extracellular matrix and redox regulation. Redox Biol. 2016, 8, 305–315. [Google Scholar] [CrossRef] [Green Version]
- Kunkemoeller, B.; Kyriakides, T.R. Redox signaling in diabetic wound healing regulates extracellular matrix deposition. Antioxid. Redox Signal. 2017, 27, 823–838. [Google Scholar] [CrossRef]
- Cai, H.; Chuang, C.Y.; Vanichkitrungruang, S.; Hawkins, C.; Davies, M.J. Hypochlorous acid-modified extracellular matrix contributes to the behavioral switching of human coronary artery smooth muscle cells. Free Radic. Biol. Med. 2019, 134, 516–526. [Google Scholar] [CrossRef]
- Feng, X.X.; Liu, M.; Yan, W.; Zhou, Z.Z.; Xia, Y.J.; Tu, W.; Li, P.-A.; Tian, D.A. β3 Integrin Promotes TGF-β1/H2O2/HOCl-Mediated Induction of Metastatic Phenotype of Hepatocellular Carcinoma Cells by Enhancing TGF-β1 Signaling. PLoS ONE 2013, 8, e79857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rees, M.D.; Whitelock, J.; Malle, E.; Chuang, C.Y.-N.; Iozzo, R.; Nilasaroya, A.; Davies, M. Myeloperoxidase-derived oxidants selectively disrupt the protein core of the heparan sulfate proteoglycan perlecan. Matrix Biol. 2010, 29, 63–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, G.; Khalil, N. TGF-β1 increases proliferation of airway smooth muscle cells by phosphorylation of map kinases. Respir. Res. 2006, 7, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zandvoort, A.; Postma, D.S.; Jonker, M.R.; Noordhoek, J.A.; Vos, J.T.W.M.; van der Geld, Y.M.; Timens, W. Altered expression of the Smad signalling pathway: Implications for COPD pathogenesis. Eur. Respir. J. 2006, 28, 533–541. [Google Scholar] [CrossRef] [Green Version]
- Soltani, A.; Sohal, S.S.; Reid, D.; Weston, S.; Wood-Baker, R.; Walters, E.H. Vessel-Associated Transforming Growth Factor-Beta1 (TGF-β1) Is Increased in the Bronchial Reticular Basement Membrane in COPD and Normal Smokers. PLoS ONE 2012, 7, e39736. [Google Scholar] [CrossRef]
- Hussell, T.; Lui, S.; Jagger, C.; Morgan, D.; Brand, O. The consequence of matrix dysfunction on lung immunity and the microbiome in COPD. Eur. Respir. Rev. 2018, 27, 180032. [Google Scholar] [CrossRef] [Green Version]
- Davoine, F.; Lacy, P. Eosinophil Cytokines, Chemokines, and Growth Factors: Emerging Roles in Immunity. Front. Immunol. 2014, 5, 570. [Google Scholar] [CrossRef] [Green Version]
- Moldoveanu, B.; Otmishi, P.; Jani, P.; Walker, J.; Sarmiento, X.; Guardiola, J.; Saad, M.; Yu, J. Inflammatory mechanisms in the lung. J. Inflamm. Res. 2009, 2, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Hua, G.; Reckhow, D.A. Characterization of Disinfection Byproduct Precursors Based on Hydrophobicity and Molecular Size. Environ. Sci. Technol. 2007, 41, 3309–3315. [Google Scholar] [CrossRef]
- Rees, M.; McNiven, T.N.; Davies, M.J. Degradation of extracellular matrix and its components by hypobromous acid. Biochem. J. 2006, 401, 587–596. [Google Scholar] [CrossRef]
- Peskin, A.V.; Winterbourn, C.C. Kinetics of the reactions of hypochlorous acid and amino acid chloramines with thiols, methionine, and ascorbate. Free Radic. Biol. Med. 2001, 30, 572–579. [Google Scholar] [CrossRef]
- Peskin, A.V.; Turner, R.; Maghzal, G.J.; Winterbourn, C.C.; Kettle, A.J. Oxidation of Methionine to Dehydromethionine by Reactive Halogen Species Generated by Neutrophils. Biochemistry 2009, 48, 10175–10182. [Google Scholar] [CrossRef]
- Rosińczuk, J.; Taradaj, J.; Dymarek, R.; Sopel, M. Mechanoregulation of wound healing and skin homeostasis. Chronic Wounds Wound Dress. Wound Heal 2018, 6, 461–477. [Google Scholar]
- Hough, K.P.; Curtiss, M.L.; Blain, T.J.; Liu, R.M.; Trevor, J.; Deshane, J.S.; Thannickal, V.J. Airway Remodeling in Asthma. Front. Med. 2020, 7, 191. [Google Scholar] [CrossRef]
- Burgstaller, G.; Oehrle, B.; Gerckens, M.; White, E.S.; Schiller, H.B.; Eickelberg, O. The instructive extracellular matrix of the lung: Basic composition and alterations in chronic lung disease. Eur. Respir. J. 2017, 50, 1601805. [Google Scholar] [CrossRef] [Green Version]
- Higham, A.; Quinn, A.M.; Cançado, J.E.D.; Singh, D. The pathology of small airways disease in COPD: Historical aspects and future directions. Respir. Res. 2019, 20, 49. [Google Scholar] [CrossRef] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Papanicolaou, M.; He, P.; Rutting, S.; Ammit, A.; Xenaki, D.; van Reyk, D.; Oliver, B.G. Extracellular Matrix Oxidised by the Granulocyte Oxidants Hypochlorous and Hypobromous Acid Reduces Lung Fibroblast Adhesion and Proliferation In Vitro. Cells 2021, 10, 3351. https://doi.org/10.3390/cells10123351
Papanicolaou M, He P, Rutting S, Ammit A, Xenaki D, van Reyk D, Oliver BG. Extracellular Matrix Oxidised by the Granulocyte Oxidants Hypochlorous and Hypobromous Acid Reduces Lung Fibroblast Adhesion and Proliferation In Vitro. Cells. 2021; 10(12):3351. https://doi.org/10.3390/cells10123351
Chicago/Turabian StylePapanicolaou, Michael, Patrick He, Sandra Rutting, Alaina Ammit, Dikaia Xenaki, David van Reyk, and Brian G. Oliver. 2021. "Extracellular Matrix Oxidised by the Granulocyte Oxidants Hypochlorous and Hypobromous Acid Reduces Lung Fibroblast Adhesion and Proliferation In Vitro" Cells 10, no. 12: 3351. https://doi.org/10.3390/cells10123351