
Quiescence, Stemness and Adipogenic Differentiation Capacity in Human DLK1−/CD34+/CD24+ Adipose Stem/Progenitor Cells

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Donors
2.2. Isolation of the Stromal Vascular Fraction (SVF) from Subcutaneous White Adipose Tissue
2.3. Flow Cytometric Sorting and Analyses of Freshly Isolated Cells
2.4. Flow Cytometric Analysis of Cultivated ASCs
2.5. Cell Culture
2.6. Adipogenic Differentiation
2.7. Real-Time Quantitative PCR
2.8. Western Blot Analysis
2.9. Proliferation Assay
2.10. BrdU Assay
2.11. Cell Cycle Analysis
2.12. Colony Formation Assay
2.13. shRNA Mediated CD24 Knock Down
2.14. Statistics
3. Results
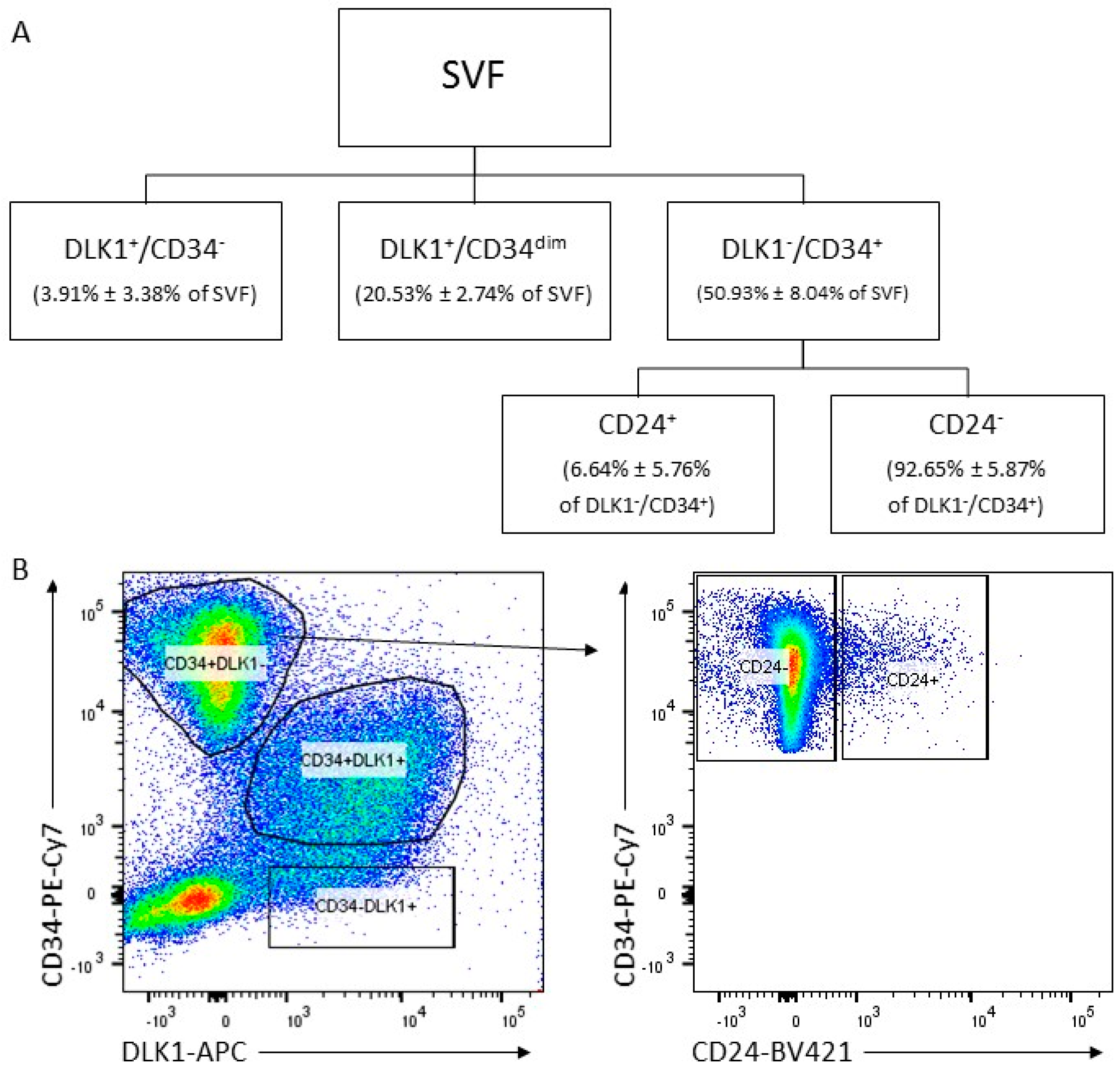
3.1. Isolation of ASC Populations Defined by DLK1, CD34 and CD24 from Human sWAT
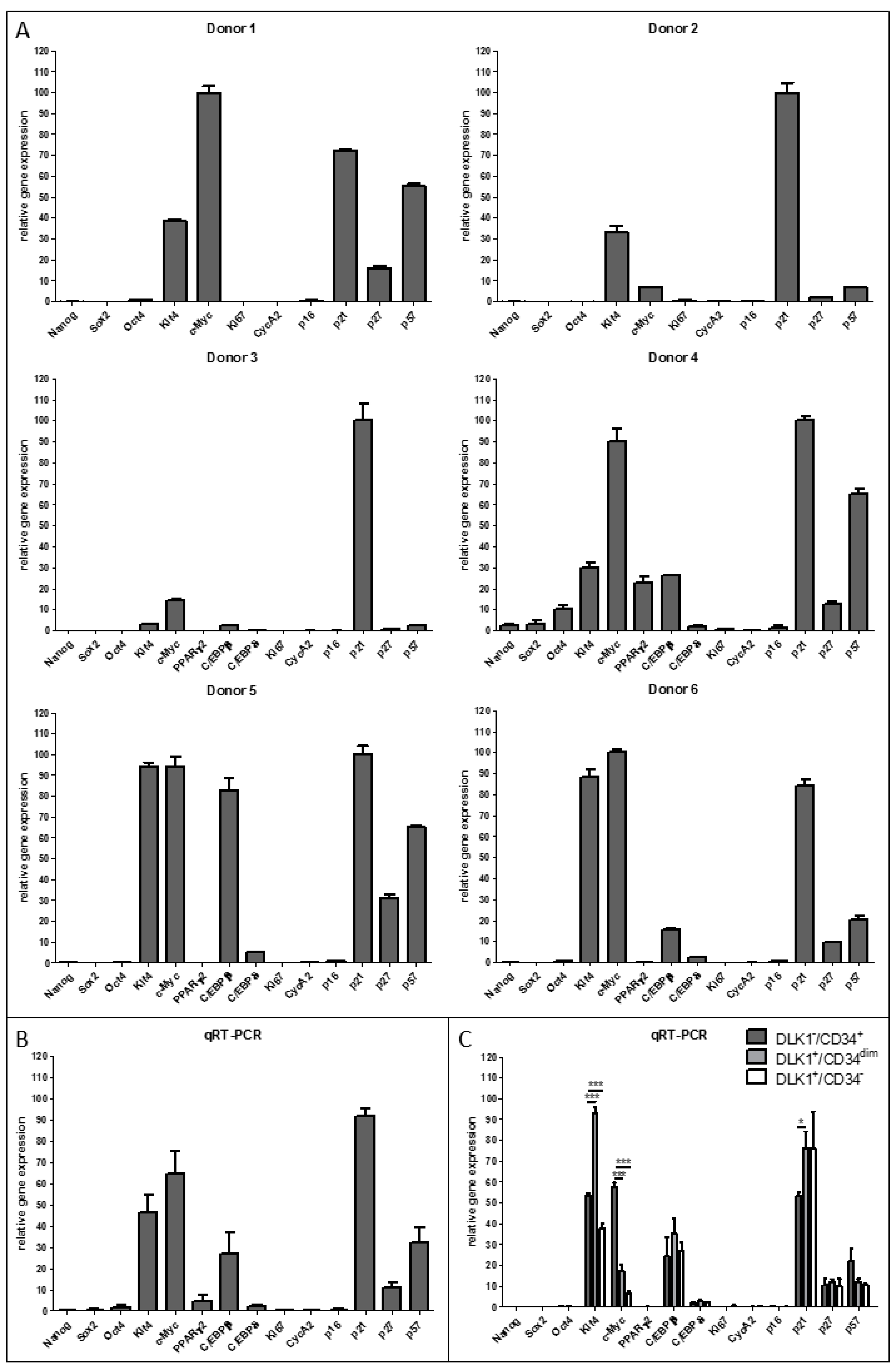
3.2. DLK−/CD34+ ASCs in Human sWAT Are in a Quiescent State and Express High Levels of the Stemness Factors KLF4 and c-MYC and of the Early Pro-Adipogenic Factor C/EBPβ
3.3. Expression of Quiescence, Proliferation, Senescence, Stemness and Adipogenesis Markers in DLK1−/CD34+ Relative to DLK1+/CD34dim and DLK1+/CD34− SVF Cell Populations
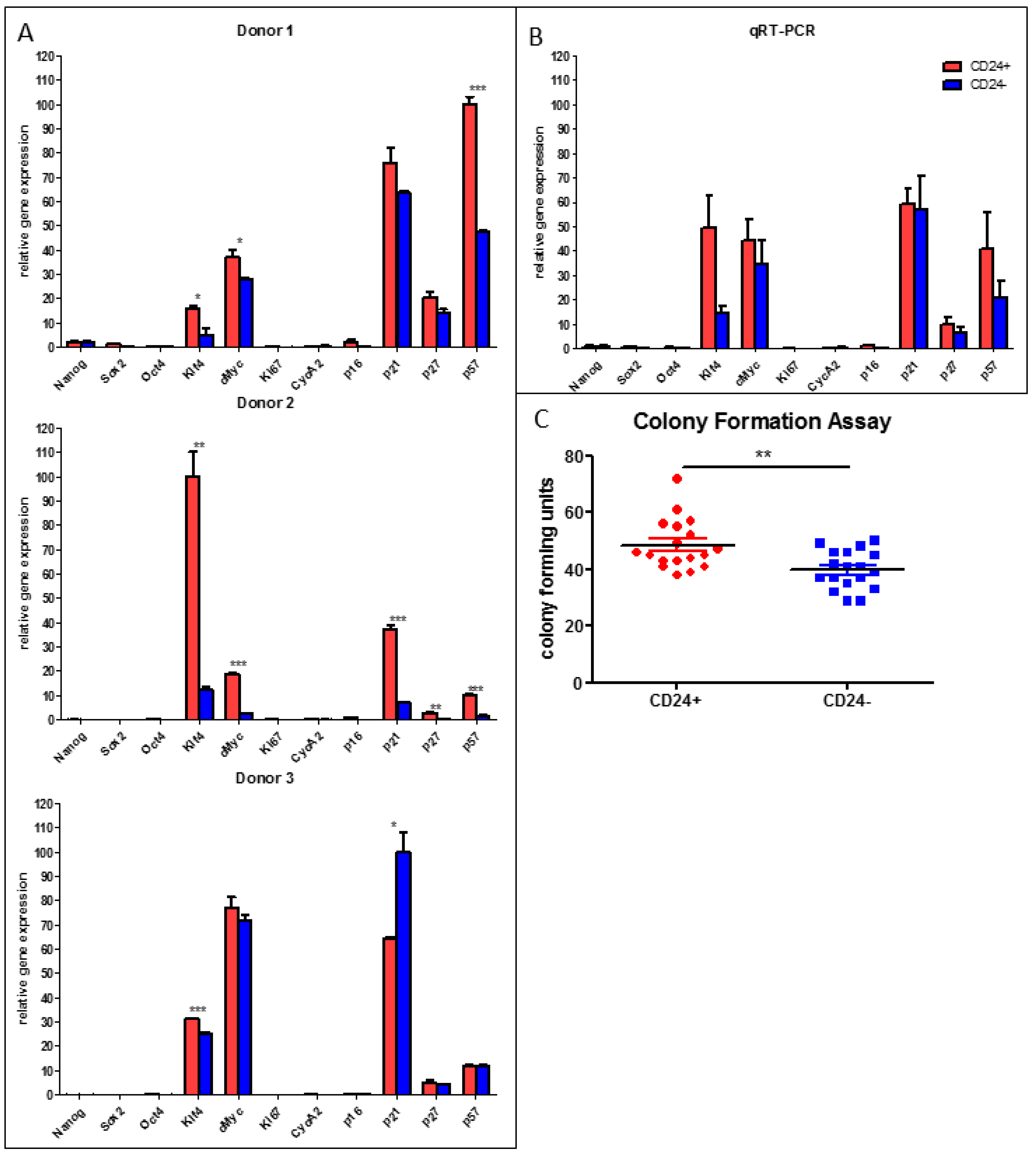
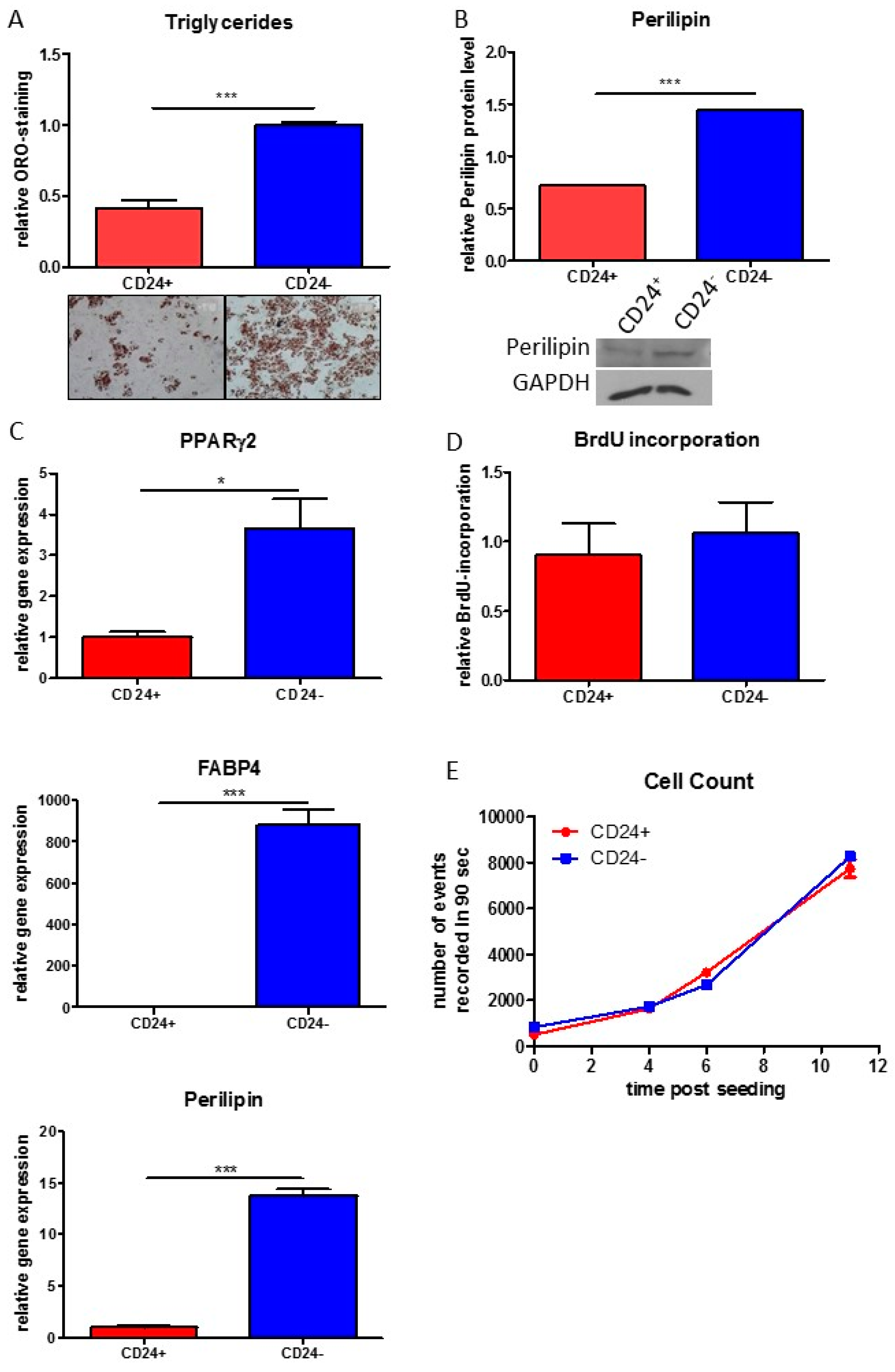
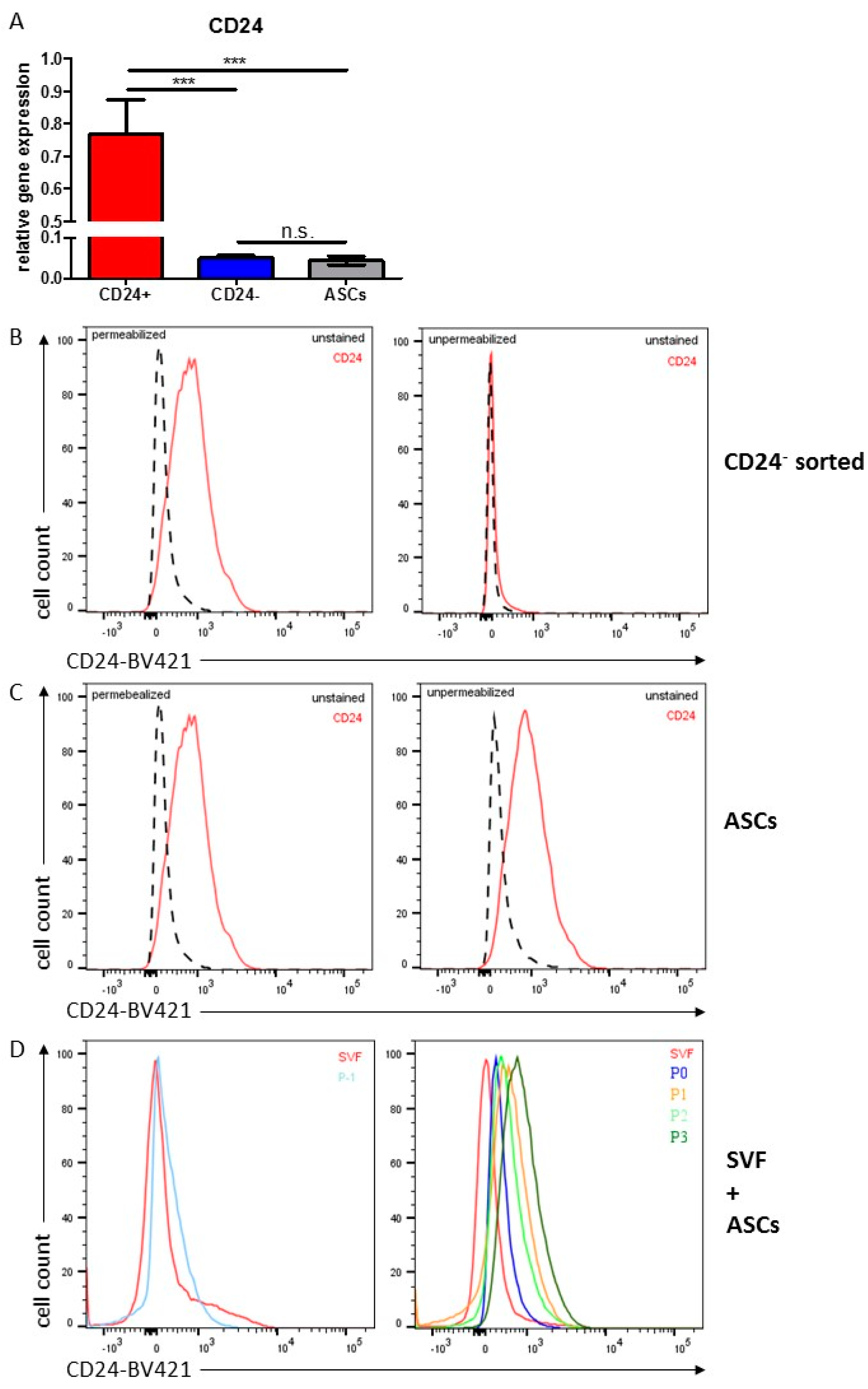
3.4. Stemness Is Higher and Adipogenic Differentiation Potential Lower in Human DLK−/CD34+/CD24+ than in DLK1−/CD34+/CD24− ASC Populations
3.5. Expression of Quiescence, Stemness, Proliferation, Senescence and Adipogenesis Markers in Routinely Isolated and In Vitro Cultivated Human DLK1−/CD34+ ASCs
3.6. Characterization of CD24 in Human ASCs
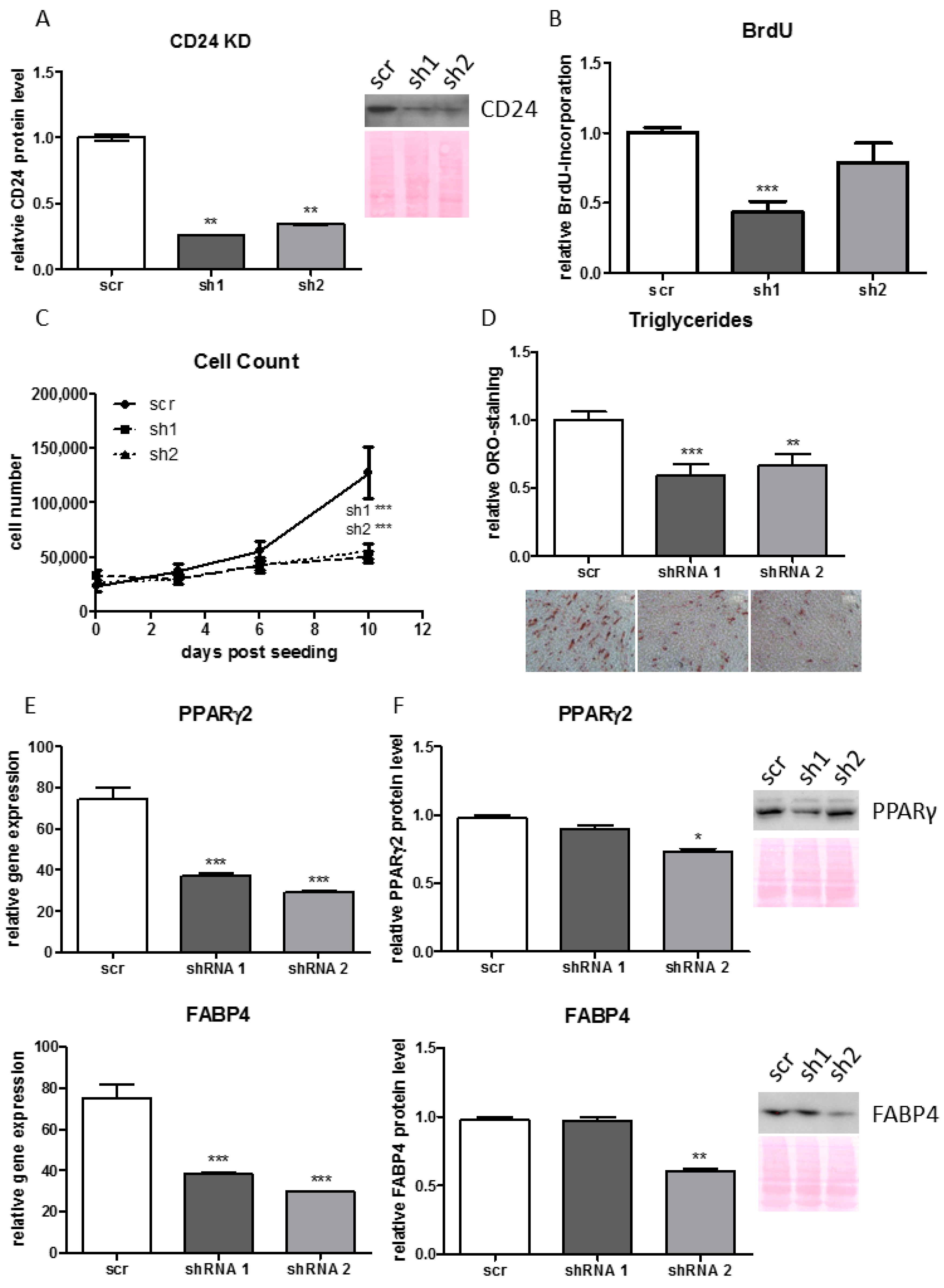
3.7. Knock-Down of CD24 Reduces Proliferative and Adipogenic Capacity of Human ASCs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Spalding, K.L.; Arner, E.; Westermark, P.O.; Bernard, S.; Buchholz, B.A.; Bergmann, O.; Blomqvist, L.; Hoffstedt, J.; Näslund, E.; Hassan, M.; et al. Dynamics of fat cell turnover in humans. Nature 2008, 453, 783–787. (In English) [Google Scholar] [CrossRef] [PubMed]
- Rodeheffer, M.S.; Birsoy, K.; Friedman, J.M. Identification of white adipocyte progenitor cells in vivo. Cell 2008, 135, 240–249. (In English) [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, N.; Akamatsu, H.; Hasegawa, S.; Yamada, T.; Nakata, S.; Ohkuma, M.; Miyachi, E.; Marunouchi, T.; Matsunaga, K. Isolation of multipotent stem cells from mouse adipose tissue. J. Derm. Sci. 2007, 48, 43–52. (In English) [Google Scholar] [CrossRef] [PubMed]
- Tang, W.; Zeve, D.; Suh, J.M.; Bosnakovski, D.; Kyba, M.; Hammer, R.E.; Tallquist, M.D.; Graff, J.M. White fat progenitor cells reside in the adipose vasculature. Science 2008, 322, 583–586. (In English) [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vishvanath, L.; Gupta, R.K. Contribution of adipogenesis to healthy adipose tissue expansion in obesity. J. Clin. Investig. 2019, 129, 4022–4031. (In English) [Google Scholar] [CrossRef]
- Minamino, T.; Dai, Z.; Jabbari, A.; Cerise, J.E.; Higgins, C.A.; Gong, W.; de Jong, A.; Harel, S.; DeStefano, G.M.; Rothman, L.; et al. A crucial role for adipose tissue p53 in the regulation of insulin resistance. Nat. Med. 2009, 15, 1082–1087. (In English) [Google Scholar] [CrossRef]
- Mitterberger, M.C.; Lechner, S.; Mattesich, M.; Zwerschke, W. Adipogenic differentiation is impaired in replicative senescent human subcutaneous adipose-derived stromal/progenitor cells. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2014, 69, 13–24. [Google Scholar] [CrossRef] [Green Version]
- Berry, D.C.; Jiang, Y.; Arpke, R.W.; Berglund, E.D.; Kyba, M.; Graff, J.M. Cellular aging contributes to failure of cold-induced beige adipocyte formation in old mice and humans. Cell Metab. 2017, 25, 481. (In English) [Google Scholar] [CrossRef] [Green Version]
- Guillermier, C.; Fazeli, P.K.; Kim, S.; Lun, M.; Zuflacht, J.P.; Milian, J.; Lee, H.; Francois-Saint-Cyr, H.; Horreard, F.; Larson, D.; et al. Imaging mass spectrometry demonstrates age-related decline in human adipose plasticity. JCI Insight 2017, 2, e90349. (In English) [Google Scholar] [CrossRef]
- Ghaben, L.; Scherer, P.E. Adipogenesis and metabolic health. Nat. Rev. Mol. Cell Biol. 2019, 20, 242–258. (In English) [Google Scholar] [CrossRef]
- Gimble, J.M.; Bunnell, B.A.; Chiu, E.S.; Guilak, F. Concise review: Adipose-derived stromal vascular fraction cells and stem cells: Let’s not get lost in translation. Stem. Cells 2011, 29, 749–754. (In English) [Google Scholar] [CrossRef] [PubMed]
- Ballini, S.; Scacco, S.; Coletti, D.; Pluchino, S.; Tatullo, M. Mesenchymal stem cells as promoters, enhancers, and playmakers of the translational regenerative medicine. Stem. Cells Int. 2017, 2017, 3292810. (In English) [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballini, S.; Cantore, S.; Scacco, S.; Coletti, D.; Tatullo, M. Mesenchymal stem cells as promoters, enhancers, and playmakers of the translational regenerative medicine 2018. Stem. Cells Int. 2018, 2018, 6927401. (In English) [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spagnuolo, G.; Codispoti, B.; Marrelli, M.; Rengo, C.; Rengo, S.; Tatullo, M. Commitment of oral-derived stem cells in dental and maxillofacial applications. Dent. J. 2018, 6, 72. (In English) [Google Scholar] [CrossRef] [Green Version]
- Zimmerlin, L.; Donnenberg, V.S.; Pfeifer, M.E.; Meyer, E.M.; Péault, B.; Rubin, J.P.; Donnenberg, A.D. Stromal vascular progenitors in adult human adipose tissue. Cytometry. Part A J. Int. Soc. Analyt. Cytol. 2010, 77, 22–30. [Google Scholar] [CrossRef]
- Zwierzina, M.E.; Ejaz, A.; Bitsche, M.; Blumer, M.J.F.; Mitterberger, M.C.; Mattesich, M.; Amann, A.; Kaiser, A.; Pechriggl, E.J.; Hörl, S.; et al. Characterization of DLK1(PREF1) + /CD34+ cells in vascular stroma of human white adipose tissue. Stem. Cell Res. 2015, 15, 403–418. [Google Scholar] [CrossRef] [Green Version]
- Frazier, T.P.; Bowles, A.; Lee, S.; Abbott, R.; Tucker, H.A.; Kaplan, D.; Wang, M.; Strong, A.; Brown, Q.; He, J.; et al. Serially transplanted nonpericytic CD146(-) adipose stromal/stem cells in silk bioscaffolds regenerate adipose tissue in vivo. Stem. Cells 2016, 34, 1097–1111. (In English) [Google Scholar] [CrossRef] [Green Version]
- Hudak, S.; Sul, H.S. Pref-1, a gatekeeper of adipogenesis. Front. Endocrinol (Lausanne) 2013, 4, 79. (In English) [Google Scholar] [CrossRef] [Green Version]
- Traustadottir, G.A.; Kosmina, R.; Sheikh, S.P.; Jensen, C.H.; Andersen, D.C. Preadipocytes proliferate and differentiate under the guidance of Delta-like 1 homolog (DLK1). Adipocyte 2013, 2, 272–275. (In English) [Google Scholar] [CrossRef] [Green Version]
- Sengenès, C.; Lolmède, K.; Zakaroff-Girard, A.; Busse, R.; Bouloumié, A. Preadipocytes in the human subcutaneous adipose tissue display distinct features from the adult mesenchymal and hematopoietic stem cells. J. Cell Physiol. 2005, 205, 114–122. (In English) [Google Scholar] [CrossRef]
- Sidney, L.E.; Branch, M.J.; Dunphy, S.E.; Dua, H.S.; Hopkinson, A. Concise review: Evidence for CD34 as a common marker for diverse progenitors. Stem. Cells 2014, 32, 1380–1389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sammar, M.; Gulbins, E.; Hilbert, K.; Lang, F.; Altevogt, P. Mouse CD24 as a signaling molecule for integrin-mediated cell binding: Functional and physical association with src-kinases. Biochem. Biophys. Res. Commun. 1997, 234, 330–334. (In English) [Google Scholar] [CrossRef] [PubMed]
- Fang, X.; Zheng, P.; Tang, J.; Liu, Y. CD24: From A to Z. Cell Mol. Immunol. 2010, 7, 100–103. (In English) [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shackleton, M.; Vaillant, F.; Simpson, K.J.; Stingl, J.; Smyth, G.K.; Asselin-Labat, M.; Wu, L.; Lindeman, G.J.; Visvader, J.E. Generation of a functional mammary gland from a single stem cell. Nature 2006, 439, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Shakiba, N.; White, C.A.; Lipsitz, Y.Y.; Yachie-Kinoshita, A.; Tonge, P.D.; Hussein, S.M.I.; Puri, M.C.; Elbaz, J.; Morrissey-Scoot, J.; Li, M.; et al. CD24 tracks divergent pluripotent states in mouse and human cells. Nat. Commun. 2015, 6, 7329. (In English) [Google Scholar] [CrossRef] [Green Version]
- Tesio, M.; Trumpp, A. Breaking the cell cycle of HSCs by p57 and friends. Cell. Stem. Cell 2011, 9, 187–192. (In English) [Google Scholar] [CrossRef] [Green Version]
- van Velthoven, T.J.; Rando, T.A. Stem cell quiescence: Dynamism, restraint, and cellular idling. Cell. Stem. Cell 2019, 24, 213–225. (In English) [Google Scholar] [CrossRef] [Green Version]
- Boyer, L.A.; Lee, T.I.; Cole, M.F.; Johnstone, S.E.; Levine, S.S.; Zucker, J.P.; Guenther, M.G.; Kumar, R.M.; Murray, H.L.; Jenner, R.G.; et al. Core transcriptional regulatory circuitry in human embryonic stem cells. Cell 2005, 22, 947–956. (In English) [Google Scholar] [CrossRef] [Green Version]
- Izadpanah, R.; Trygg, C.; Patel, B.; Kriedt, C.; Dufour, J.; Gimble, J.M.; Bunnell, B.A. Biologic properties of mesenchymal stem cells derived from bone marrow and adipose tissue. J. Cell Biochem. 2006, 99, 1285–1297. (In English) [Google Scholar] [CrossRef] [Green Version]
- Berg, J.S.; Goodell, M.A. An argument against a role for Oct4 in somatic stem cells. Cell. Stem Cell 2007, 1, 359–360. (In English) [Google Scholar] [CrossRef] [Green Version]
- Greco, S.J.; Liu, K.; Rameshwar, P. Functional similarities among genes regulated by OCT4 in human mesenchymal and embryonic stem cells. Stem. Cells 2007, 25, 3143–3154. (In English) [Google Scholar] [CrossRef] [PubMed]
- Lengner, C.J.; Camargo, F.D.; Hochedlinger, K.; Scholer, H.R.; Tomilin, A.; Jaenisch, R. Oct4 expression is not required for mouse somatic stem cell self-renewal. Cell. Stem. Cell 2007, 1, 403–415. (In English) [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liedtke, S.; Enczmann, J.; Waclawczyk, S.; Wernet, P.; Kögler, G. Oct4 and its pseudogenes confuse stem cell research. Cell. Stem. Cell 2007, 1, 364–366. (In English) [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaltz, N.; Funari, A.; Hippauf, S.; Delorme, B.; Noël, D.; Riminucci, M.; Jacobs, V.R.; Häupl, T.; Jorgensen, C.; Charbord, P.; et al. In vivo osteoprogenitor potency of human stromal cells from different tissues does not correlate with expression of POU5F1 or its pseudogenes. Stem. Cells 2008, 26, 2419–2424. (In English) [Google Scholar] [CrossRef]
- Riekstina, U.; Cakstina, I.; Parfejevs, V.; Hoogduijn, M.; Jankovskis, G.; Muiznieks, I.; Muceniece, R.; Ancans, J. Embryonic stem cell marker expression pattern in human mesenchymal stem cells derived from bone marrow, adipose tissue, heart and dermis. Stem. Cell. Rev. Rep. 2009, 5, 378–386. (In English) [Google Scholar] [CrossRef]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. (In English) [Google Scholar] [CrossRef] [Green Version]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 2007, 131, 861–872. (In English) [Google Scholar] [CrossRef] [Green Version]
- Laurenti, E.; Wilson, A.; Trumpp, A. Myc’s other life: Stem cells and beyond. Curr. Opin. Cell. Biol. 2009, 21, 844–854. (In English) [Google Scholar] [CrossRef]
- Park, K.W.; Halperin, D.S.; Tontonoz, P. Before they were fat: Adipocyte progenitors. Cell Metab. 2008, 8, 454–457. (In English) [Google Scholar] [CrossRef] [Green Version]
- Seoane, J.; Le, H.V.; Massagué, J. Myc suppression of the p21(Cip1) Cdk inhibitor influences the outcome of the p53 response to DNA damage. Nature 2002, 419, 729–734. (In English) [Google Scholar] [CrossRef]
- Shields, J.M.; Christy, R.J.; Yang, V.W. Identification and characterization of a gene encoding a gut-enriched Krüppel-like factor expressed during growth arrest. J. Biol. Chem. 1996, 271, 20009–20017. (In English) [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Geiman, D.E.; Shields, J.M.; Dang, D.T.; Mahatan, C.S.; Kaestner, K.H.; Biggs, J.R.; Kraft, A.S.; Yang, V.W. The gut-enriched Kruppel-like factor (Kruppel-like factor 4) mediates the transactivating effect of p53 on the p21WAF1/Cip1 promoter. J. Biol. Chem. 2000, 275, 18391–18398. (In English) [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yusuf, I.; Kharas, M.G.; Chen, J.; Peralta, R.Q.; Maruniak, A.; Sareen, P.; Yang, V.W.; Kaestner, K.H.; Fruman, D.A. KLF4 is a FOXO target gene that suppresses B cell proliferation. Int. Immunol. 2008, 20, 671–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McConnell, B.B.; Yang, V.W. Mammalian Krüppel-like factors in health and diseases. Physiol. Rev. 2010, 90, 1337–1381. (In English) [Google Scholar] [CrossRef] [PubMed]
- Pulverer, B.; Sommer, A.; McArthur, G.A.; Eisenman, R.N.; Lüscher, B. Analysis of Myc/Max/Mad network members in adipogenesis: Inhibition of the proliferative burst and differentiation by ectopically expressed Mad1. J. Cell. Physiol. 2000, 183, 399–410. [Google Scholar] [CrossRef]
- Birsoy, Z.; Chen, E.C.; Friedman, J. Transcriptional regulation of adipogenesis by KLF4. Cell Metab. 2008, 7, 339–347. (In English) [Google Scholar] [CrossRef] [Green Version]
- Ntambi, M.; Young-Cheul, K. Adipocyte differentiation and gene expression. J. Nutr. 2000, 130, 3122S–3126S. (In English) [Google Scholar] [CrossRef]
- Lefterova, I.; Haakonsson, A.K.; Lazar, M.A.; Mandrup, S. PPARγ and the global map of adipogenesis and beyond. Trends Endocrinol. Metab. 2014, 25, 293–302. (In English) [Google Scholar] [CrossRef] [Green Version]
- Cristancho, G.; Lazar, M.A. Forming functional fat: A growing understanding of adipocyte differentiation. Nat. Rev. Mol. Cell Biol. 2011, 12, 722–734. (In English) [Google Scholar] [CrossRef]
- Mandl, M.; Wagner, S.A.; Hatzmann, F.M.; Mitterberger-Vogt, M.C.; Zwierzina, M.E.; Mattesich, M.; Zwerschke, W. Sprouty1 is a weight-loss target gene in human adipose stem/progenitor cells that is mandatory for the initiation of adipogenesis. Cell. Death Dis. 2019, 10, 411. (In English) [Google Scholar] [CrossRef] [Green Version]
- Mitterberger, C.; Lechner, S.; Mattesich, M.; Kaiser, A.; Probst, D.; Wenger, N.; Pierer, G.; Zwerschke, W. DLK1(PREF1) is a negative regulator of adipogenesis in CD105(+)/CD90(+)/CD34(+)/CD31(-)/FABP4(-) adipose-derived stromal cells from subcutaneous abdominal fat pats of adult women. Stem. Cell Res. 2012, 9, 35–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitchell, J.B.; McIntosh, K.; Zvonic, S.; Garrett, S.; Floyd, Z.E.; Kloster, A.; di Halvorsen, Y.; Storms, R.W.; Goh, B.; Kilroy, G.; et al. Immunophenotype of human adipose-derived cells: Temporal changes in stromal-associated and stem cell-associated markers. Stem. Cells 2006, 24, 376–385. (In English) [Google Scholar] [CrossRef] [PubMed]
- Mitterberger, M.C.; Mattesich, M.; Zwerschke, W. Bariatric surgery and diet-induced long-term caloric restriction protect subcutaneous adipose-derived stromal/progenitor cells and prolong their life span in formerly obese humans. Exp. Gerontol. 2014, 56, 106–113. (In English) [Google Scholar] [CrossRef] [PubMed]
- Pack, L.R.; Daigh, L.H.; Meyer, T. Putting the brakes on the cell cycle: Mechanisms of cellular growth arrest. Curr. Opin. Cell. Biol. 2019, 60, 106–113. (In English) [Google Scholar] [CrossRef] [PubMed]
- Søndergaard, R.H.; Follin, B.; Lund, L.D.; Juhl, M.; Ekblond, A.; Kastrup, J.; Haack-Sørensen, M. Senescence and quiescence in adipose-derived stromal cells: Effects of human platelet lysate, fetal bovine serum and hypoxia. Cytotherapy 2017, 19, 95–106. (In English) [Google Scholar] [CrossRef] [Green Version]
- van Deursen, J.M. The role of senescent cells in ageing. Nature 2014, 509, 439–446. (In English) [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salotti, J.; Johnson, P.F. Regulation of senescence and the SASP by the transcription factor C/EBPβ. Exp. Gerontol. 2019, 128, 110752. (In English) [Google Scholar] [CrossRef]
- Baker, D.J.; Childs, B.G.; Durik, M.; Wijers, M.E.; Sieben, C.J.; Zhong, J.; Saltness, R.A.; Jeganathan, K.B.; Verzosa, G.C.; Pezeshki, A.; et al. Naturally occurring p16(Ink4a)-positive cells shorten healthy lifespan. Nature 2016, 530, 184–189. (In English) [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; Palmer, A.K.; Ding, H.; Weivoda, M.M.; Pirtskhalava, T.; White, T.A.; Sepe, A.; Johnson, K.O.; Stout, M.B.; Giorgadze, N.; et al. Targeting senescent cells enhances adipogenesis and metabolic function in old age. Elife 2015, 4, e12997. (In English) [Google Scholar] [CrossRef]
- Pierantozzi, E.; Gava, B.; Manini, I.; Roviello, F.; Marotta, G.; Chiavarelli, M.; Sorrentino, V. Pluripotency regulators in human mesenchymal stem cells: Expression of NANOG but not of OCT-4 and SOX-2. Stem. Cells Dev. 2011, 20, 915–923. (In English) [Google Scholar] [CrossRef]
- Chiche, A.; Chen, C.; Li, H. The crosstalk between cellular reprogramming and senescence in aging and regeneration. Exp. Gerontol. 2020, 138, 111005. (In English) [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Michowski, W.; Kolodziejczyk, A.; Sicinski, P. The cell cycle in stem cell proliferation, pluripotency and differentiation. Nat. Cell Biol. 2019, 21, 1060–1067. (In English) [Google Scholar] [CrossRef] [PubMed]
- Coller, A.; Sang, L.; Roberts, J.M. A new description of cellular quiescence. PLoS Biol. 2006, 4, e83. (In English) [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lefterova, M.I.; Zhang, Y.; Steger, D.J.; Schupp, M.; Schug, J.; Cristancho, A.; Feng, D.; Zhuo, D.; Stoeckert, C.J., Jr.; Liu, X.S.; et al. PPARgamma and C/EBP factors orchestrate adipocyte biology via adjacent binding on a genome-wide scale. Genes Dev. 2008, 22, 2941–2952. (In English) [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siersbæk, R.; Nielsen, R.; John, S.; Sung, M.-H.; Baek, S.; Loft, A.; Hager, G.L.; Mandrup, S. Extensive chromatin remodelling and establishment of transcription factor ‘hotspots’ during early adipogenesis. EMBO J. 2011, 30, 1459–1472. (In English) [Google Scholar] [CrossRef] [Green Version]
- Mikkelsen, T.S.; Xu, Z.; Zhang, X.; Gimble, J.M.; Lander, E.S.; Rosen, E.D. Comparative epigenomic analysis of murine and human adipogenesis. Cell 2010, 143, 156–169. (In English) [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berry, R.; Rodeheffer, M.S. Characterization of the adipocyte cellular lineage in vivo. Nat. Cell. Biol. 2013, 15, 302–308. [Google Scholar] [CrossRef]
- Wang, L.; Liu, R.; Ye, P.; Wong, C.; Chen, G.; Zhou, P.; Sakabe, K.; Zheng, X.; Wu, W.; Zhang, P.; et al. Intracellular CD24 disrupts the ARF-NPM interaction and enables mutational and viral oncogene-mediated p53 inactivation. Nat. Commun. 2015, 6, 5909. [Google Scholar] [CrossRef]
- Marrelli, M.; Pujia, A.; Palmieri, F.; Gatto, R.; Falisi, G.; Gargari, M.; Caruso, S.; Apicella, D.; Rastelli, C.; Nardi, G.M.; et al. Innovative approach for the in vitro research on biomedical scaffolds designed and customized with CAD-CAM technology. Int. J. Immunopathol. Pharmacol. 2016, 29, 778–783. (In English) [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hatzmann, F.M.; Ejaz, A.; Wiegers, G.J.; Mandl, M.; Brucker, C.; Lechner, S.; Rauchenwald, T.; Zwierzina, M.; Baumgarten, S.; Wagner, S.; et al. Quiescence, Stemness and Adipogenic Differentiation Capacity in Human DLK1−/CD34+/CD24+ Adipose Stem/Progenitor Cells. Cells 2021, 10, 214. https://doi.org/10.3390/cells10020214
Hatzmann FM, Ejaz A, Wiegers GJ, Mandl M, Brucker C, Lechner S, Rauchenwald T, Zwierzina M, Baumgarten S, Wagner S, et al. Quiescence, Stemness and Adipogenic Differentiation Capacity in Human DLK1−/CD34+/CD24+ Adipose Stem/Progenitor Cells. Cells. 2021; 10(2):214. https://doi.org/10.3390/cells10020214
Chicago/Turabian StyleHatzmann, Florian M., Asim Ejaz, G. Jan Wiegers, Markus Mandl, Camille Brucker, Stefan Lechner, Tina Rauchenwald, Marit Zwierzina, Saphira Baumgarten, Sonja Wagner, and et al. 2021. "Quiescence, Stemness and Adipogenic Differentiation Capacity in Human DLK1−/CD34+/CD24+ Adipose Stem/Progenitor Cells" Cells 10, no. 2: 214. https://doi.org/10.3390/cells10020214
APA StyleHatzmann, F. M., Ejaz, A., Wiegers, G. J., Mandl, M., Brucker, C., Lechner, S., Rauchenwald, T., Zwierzina, M., Baumgarten, S., Wagner, S., Mattesich, M., Waldegger, P., Pierer, G., & Zwerschke, W. (2021). Quiescence, Stemness and Adipogenic Differentiation Capacity in Human DLK1−/CD34+/CD24+ Adipose Stem/Progenitor Cells. Cells, 10(2), 214. https://doi.org/10.3390/cells10020214