The Imaging of Guard Cells of thioglucosidase (tgg) Mutants of Arabidopsis Further Links Plant Chemical Defence Systems with Physical Defence Barriers

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Plant Material and Growth Conditions
2.2. Microscopy Analyses
2.3. ABA Application and Estimation of Stomatal Aperture
3. Results
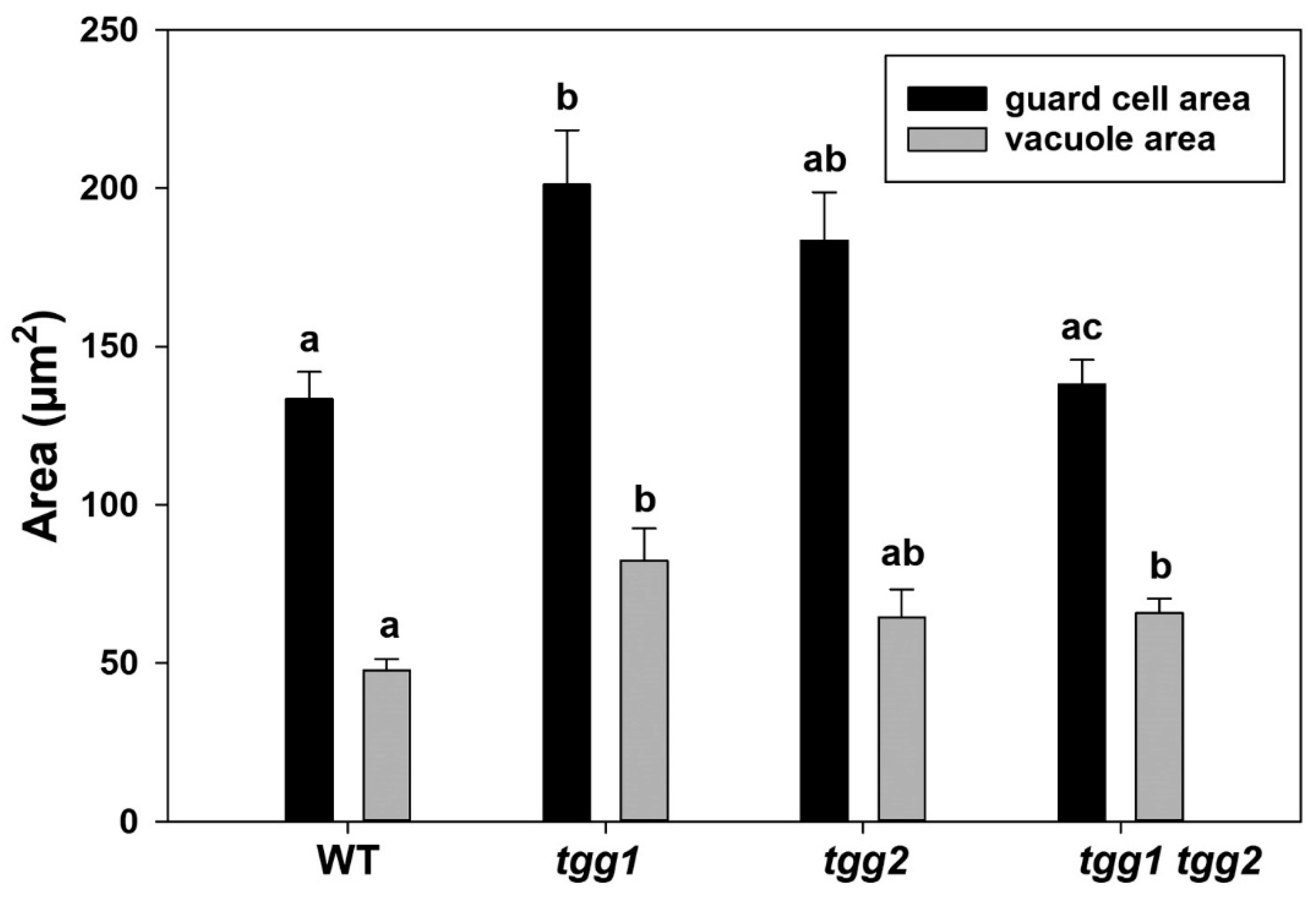
3.1. tgg Mutants Differ for Stomata and Area of Stomatal Complex
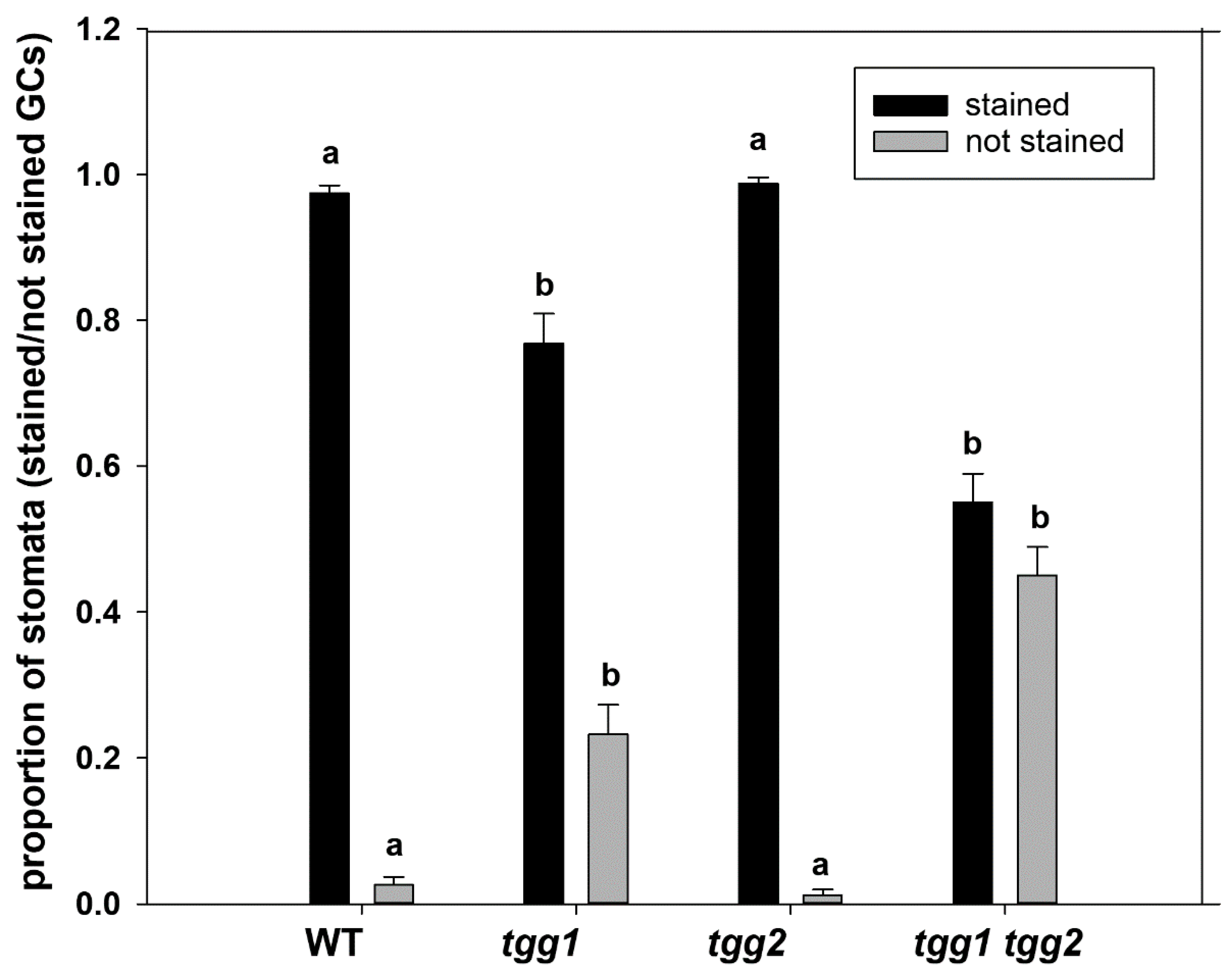
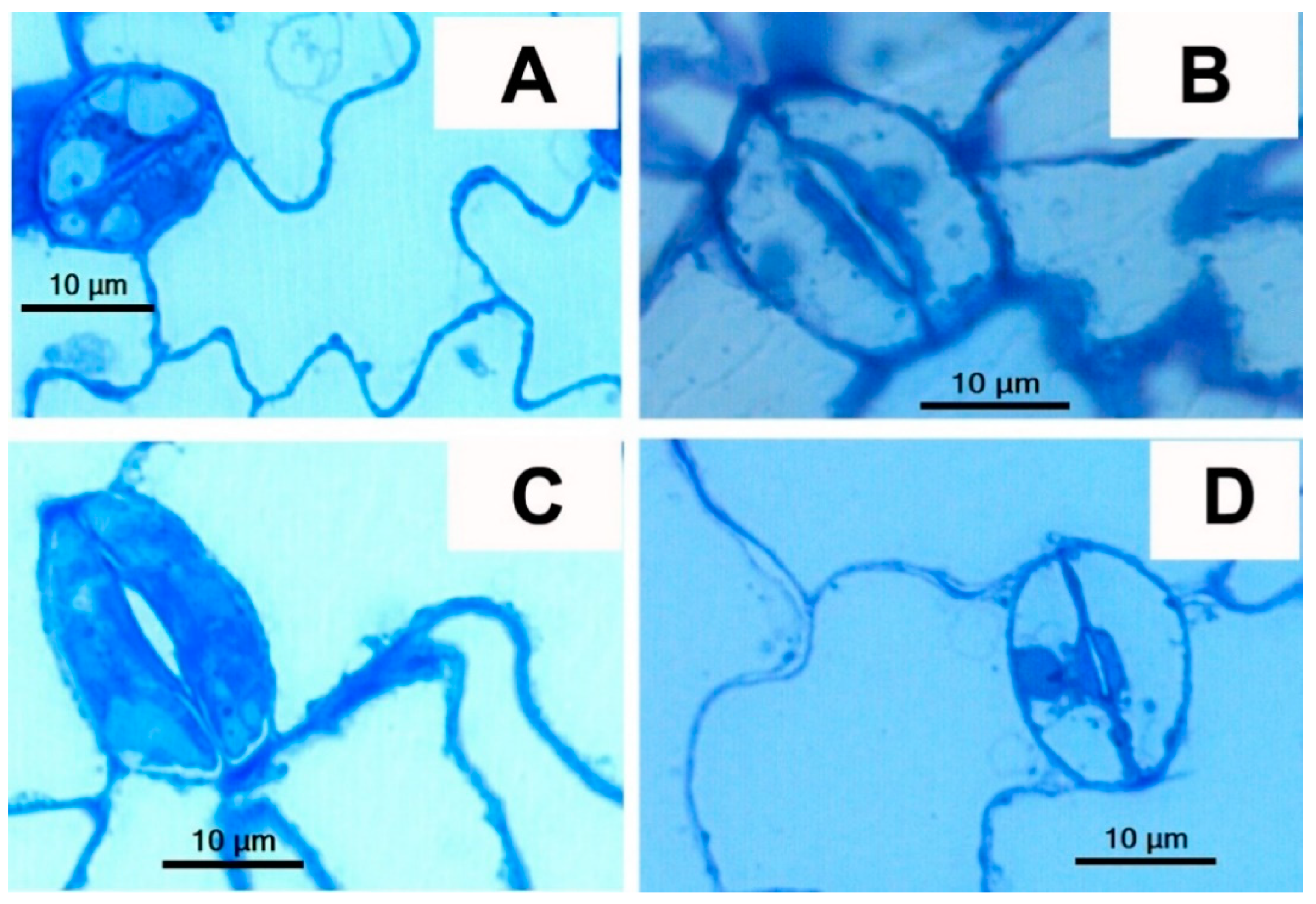
3.2. The GCs Vacuoles of tgg1 Single and tgg1 tgg2 Double Mutants Lack Staining with Toluidine Blue
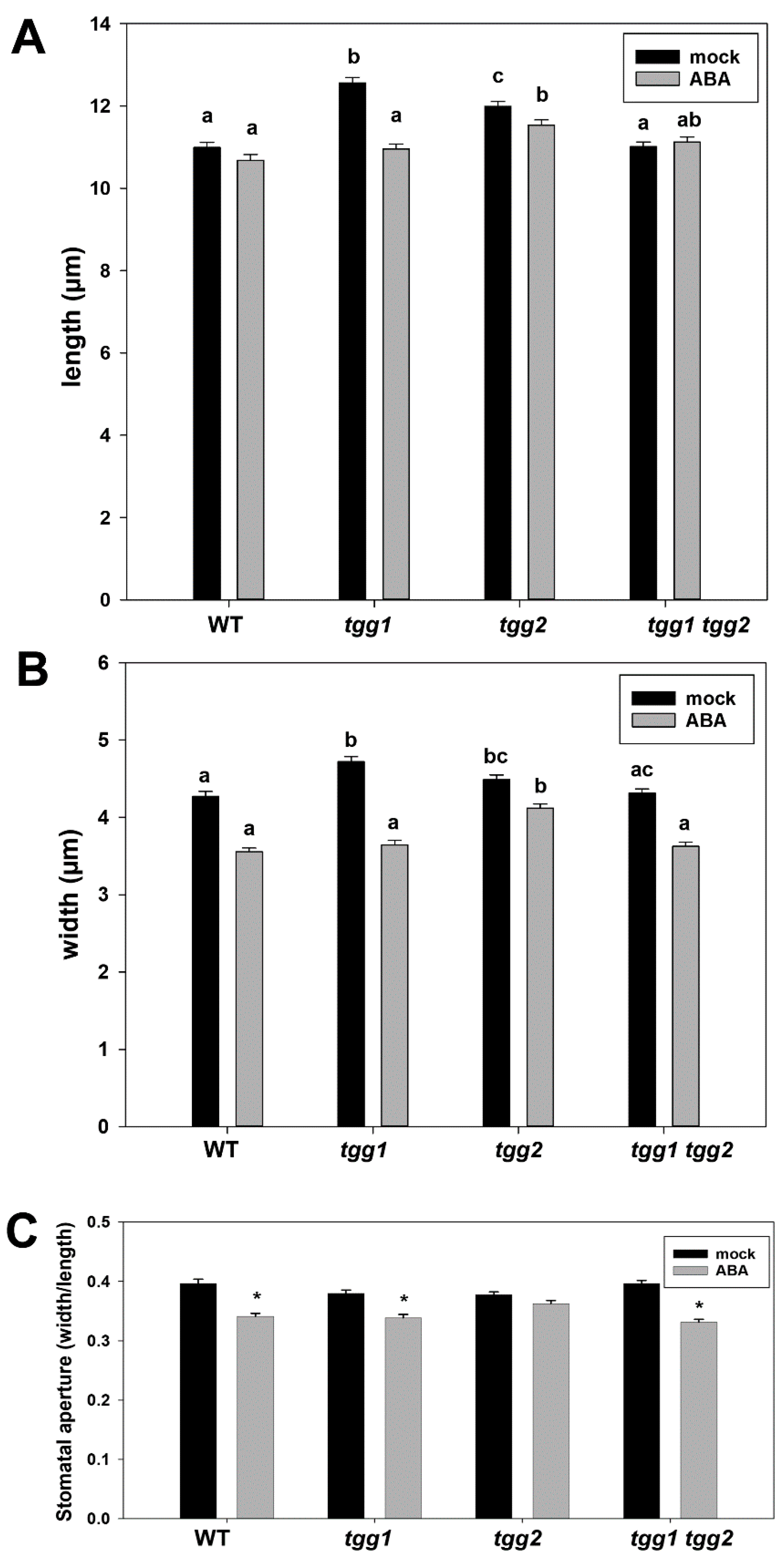
3.3. ABA Treatment Affects Stomatal Aperture of tgg1 Single and tgg1 tgg2 Double Mutant
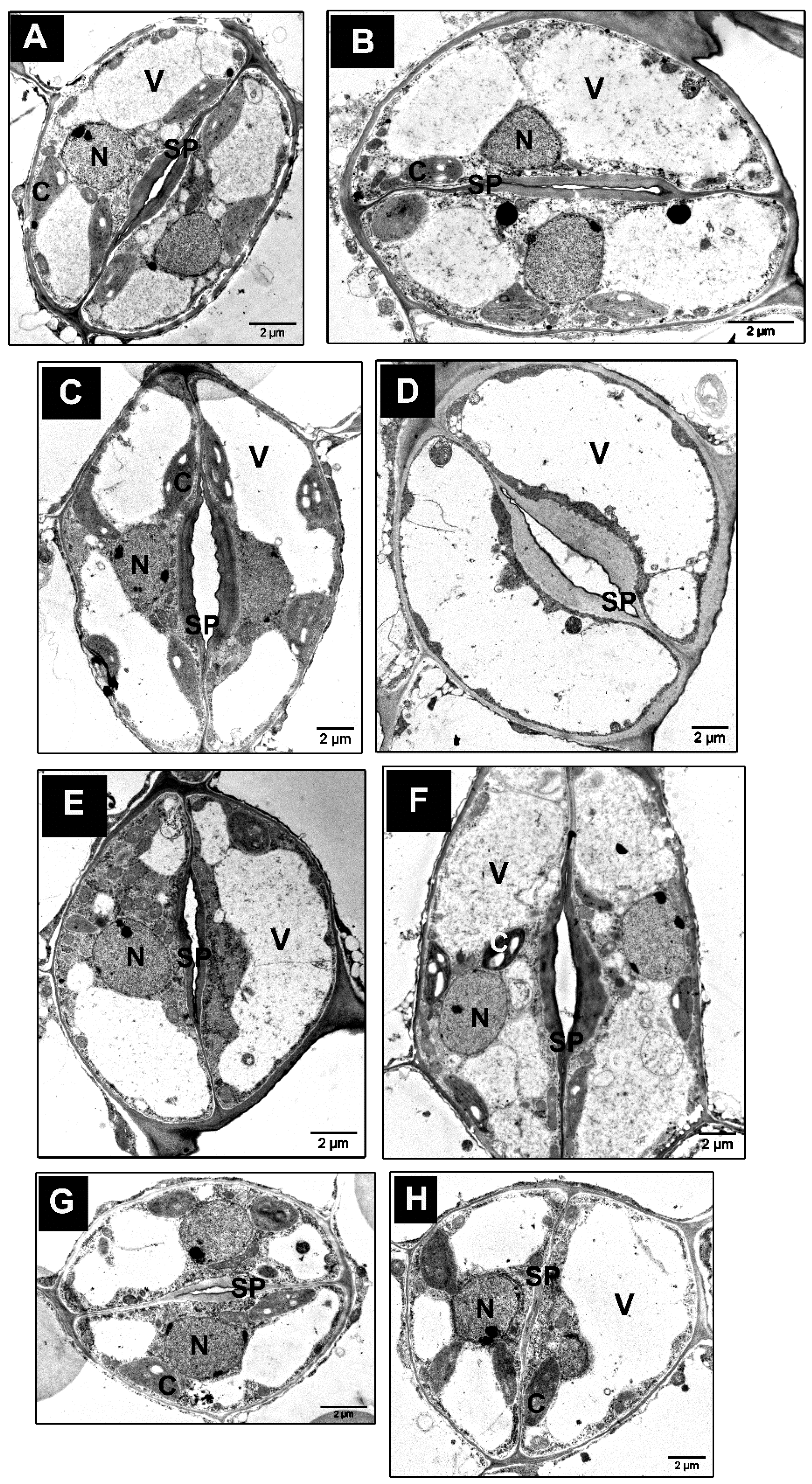
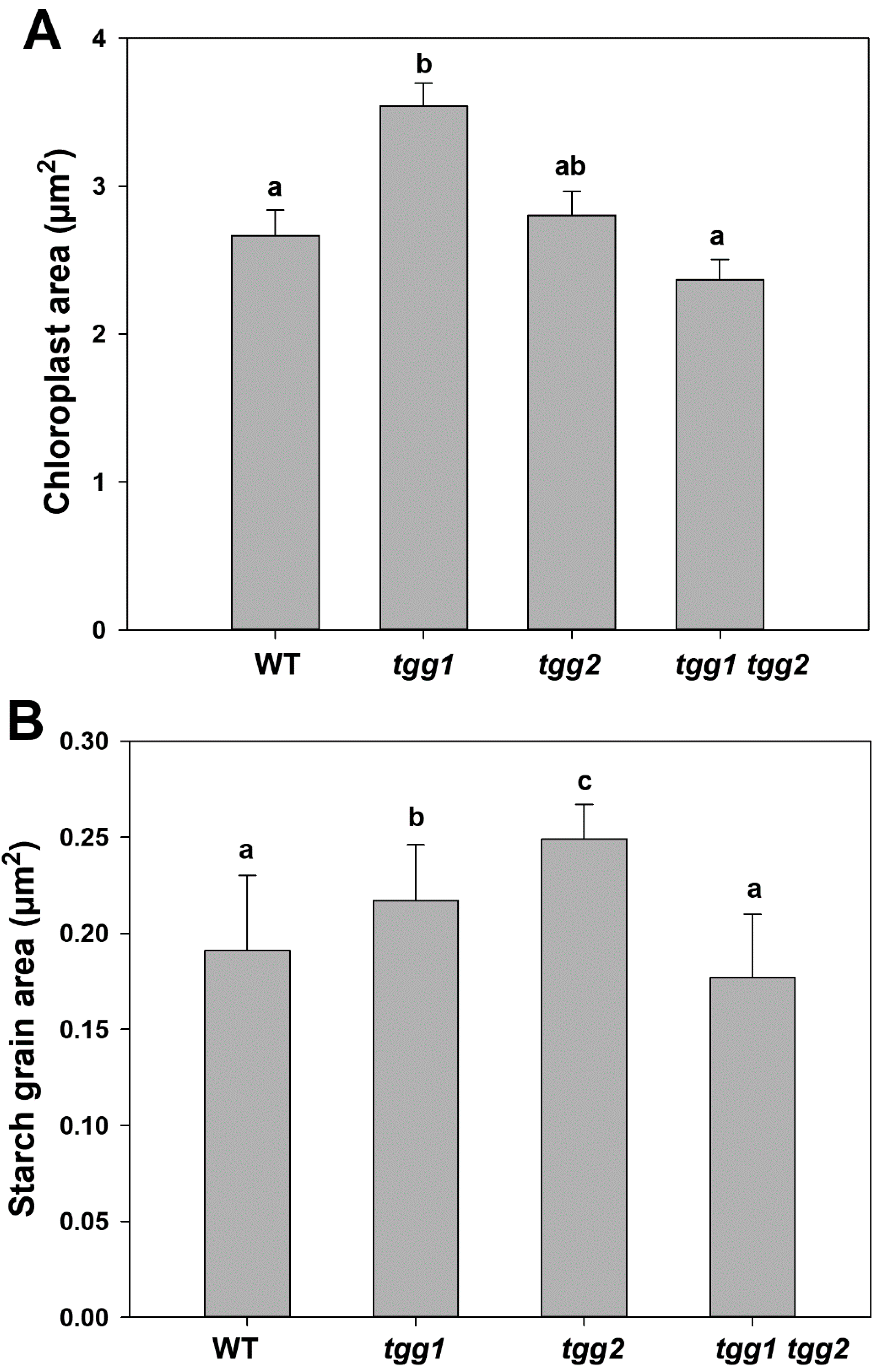
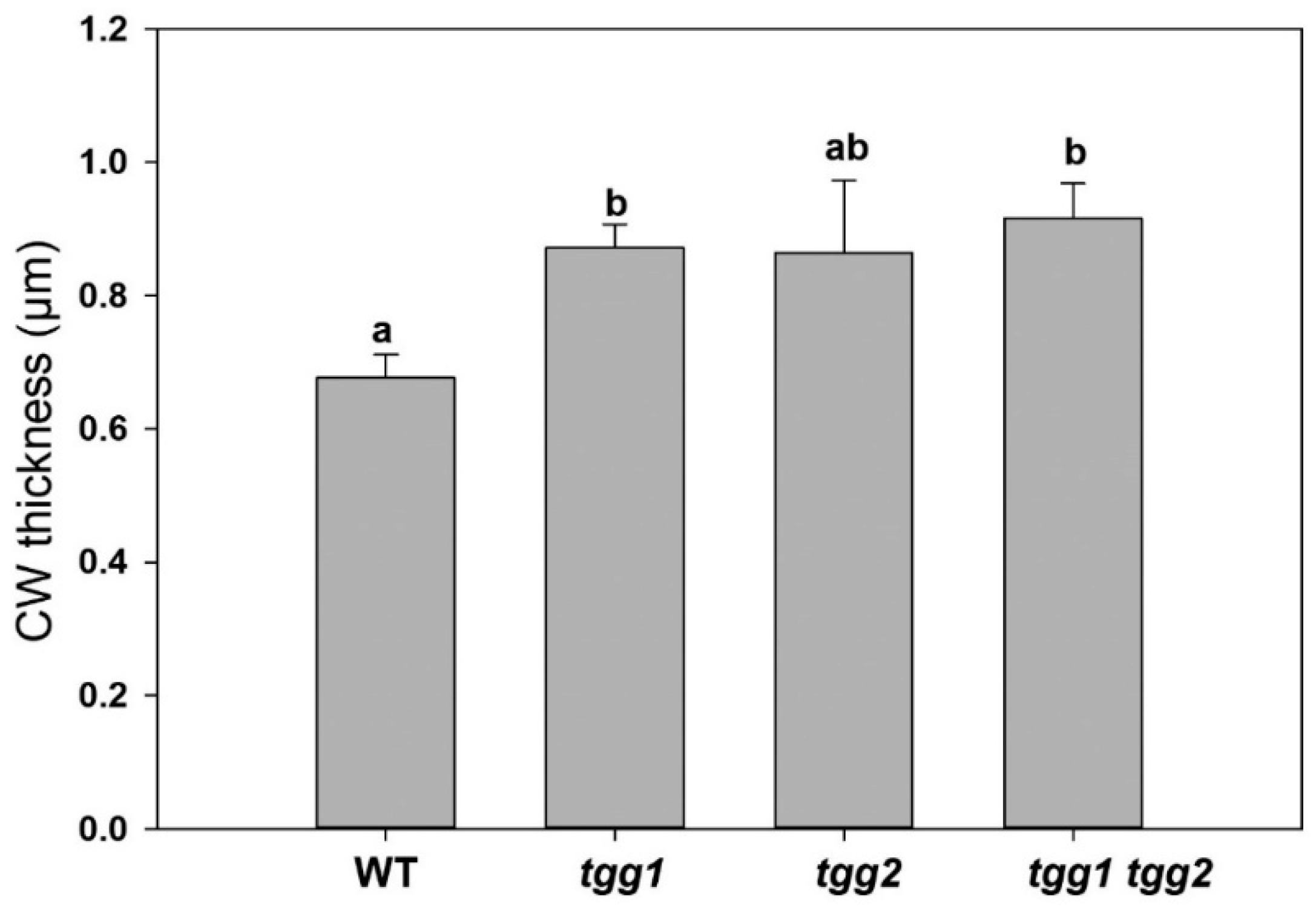
3.4. Ultrastructure Changes in Stomata and GCs of tgg Single and Double Mutants
3.5. Arabidopsis Plants Expressing TGG1-GFP Shows GC Expression Pattern and Localization of TGG1 Fusion Protein in GC Vacuoles
4. Discussion
4.1. Scarcity of TEM Data on Stomata and GCs of Arabidopsis
4.2. Bigger and Less Electron-Dense Vacuoles in GCs of tgg1 Single and tgg1 tgg2 Double Mutants Most Likely due to Lack of Myrosinase/TGG1
4.3. Stomatal Opening/Closing Possibly Affects Vacuolation and Stomatal Complexes in tgg Mutants
4.4. Cuticle, Transpiration, Stomatal Ledges, and Closed Stomata in tgg1 tgg2 Double Mutant
4.5. Chloroplasts, Starch Degradation in GCs, Glucosinolates, Myrosinases, and Responses to Blue Light
4.6. Glucosinolates, Myrosinases, and Stomata/GC-ABA Responses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Thangstad, O.P.; Gilde, B.; Chadchawan, S.; Seem, M.; Husebye, H.; Bradley, D.; Bones, A.M. Cell specific, cross-species expression of myrosinases in Brassica napus, Arabidopsis thaliana and Nicotiana tabacum. Plant Mol. Biol. 2004, 54, 597–611. [Google Scholar] [CrossRef] [PubMed]
- Shirakawa, M.; Ueda, H.; Shimada, T.; Hara-Nishimura, I. FAMA: A molecular link between stomata and myrosin cells. Trends Plant Sci. 2016, 21, 861–871. [Google Scholar] [CrossRef] [PubMed]
- Ahuja, I.; Rohloff, J.; Bones, A.M. Defence mechanisms of Brassicaceae: Implications for plant-insect interactions and potential for integrated pest management. A Review. Agron. Sustain. Dev. 2010, 30, 311–348. [Google Scholar] [CrossRef] [Green Version]
- Kissen, R.; Rossiter, J.; Bones, A. The ‘mustard oil bomb’: Not so easy to assemble?! Localization, expression and distribution of the components of the myrosinase enzyme system. Phytochem. Rev. 2009, 8, 69–86. [Google Scholar] [CrossRef]
- Bones, A.M.; Rossiter, J.T. The enzymic and chemically induced decomposition of glucosinolates. Phytochemistry 2006, 67, 1053–1067. [Google Scholar] [CrossRef] [PubMed]
- Bones, A.M.; Rossiter, J.T. The myrosinase-glucosinolate system, its organisation and biochemistry. Physiol. Plant. 1996, 97, 194–208. [Google Scholar] [CrossRef]
- Andréasson, E.; Jørgensen, L.B.; Höglund, A.S.; Rask, L.; Meijer, J. Different myrosinase and idioblast distribution in Arabidopsis and Brassica napus. Plant Physiol. 2001, 127, 1750–1763. [Google Scholar] [CrossRef]
- Husebye, H.; Chadchawan, S.; Winge, P.; Thangstad, O.P.; Bones, A.M. Guard cell- and phloem idioblast-specific expression of thioglucoside glucohydrolase 1 (myrosinase) in Arabidopsis. Plant Physiol. 2002, 128, 1180–1188. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Sack, F.D. Myrosin idioblast cell fate and development are regulated by the Arabidopsis transcription factor FAMA, the auxin pathway, and vesicular trafficking. Plant Cell 2014, 26, 4053–4066. [Google Scholar] [CrossRef] [Green Version]
- Bones, A.M.; Thangstad, O.P.; Haugen, O.A.; Espevik, T. Fate of myrosin cells—Characterization of monoclonal-antibodies against myrosinase. J. Exp. Bot. 1991, 42, 1541–1549. [Google Scholar] [CrossRef]
- Bones, A.; Iversen, T.H. Myrosin cells and myrosinase. Isr. J. Bot. 1985, 34, 351–375. [Google Scholar]
- Nadeau, J.A.; Sack, F.D. Stomatal development in Arabidopsis. Arab. Book Am. Soc. Plant Biol. 2002, 1, E0066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nadeau, J.A. Stomatal development: New signals and fate determinants. Curr. Opin. Plant Biol. 2009, 12, 29–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blatt, M.R. Cellular signaling and volume control in stomatal movements in plants. Annu. Rev. Cell Dev. Biol. 2000, 16, 221–241. [Google Scholar] [CrossRef] [PubMed]
- Monda, K.; Araki, H.; Kuhara, S.; Ishigaki, G.; Akashi, R.; Negi, J.; Kojima, M.; Sakakibara, H.; Takahashi, S.; Hashimoto-Sugimoto, M.; et al. Enhanced stomatal conductance by a spontaneous Arabidopsis tetraploid, Me-0, results from increased stomatal size and greater stomatal aperture. Plant Physiol. 2016, 170, 1435–1444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blatt, M.R. Toward understanding vesicle traffic and the guard cell model. New Phytol. 2002, 153, 405–413. [Google Scholar] [CrossRef]
- Karnik, R.; Waghmare, S.; Zhang, B.; Larson, E.; Lefoulon, C.; Gonzalez, W.; Blatt, M.R. Commandeering channel voltage sensors for secretion, cell turgor, and volume control. Trends Plant Sci. 2017, 22, 81–95. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Ohashi-Ito, K.; Bergmann, D.C. Orthologs of Arabidopsis thaliana stomatal bHLH genes and regulation of stomatal development in grasses. Development 2009, 136, 2265–2276. [Google Scholar] [CrossRef] [Green Version]
- Rudall, P.J.; Chen, E.D.; Cullen, E. Evolution and development of monocot stomata. Am. J. Bot. 2017, 104, 1122–1141. [Google Scholar] [CrossRef]
- Li, Y.; Beisson, F.; Koo, A.J.K.; Molina, I.; Pollard, M.; Ohlrogge, J. Identification of acyltransferases required for cutin biosynthesis and production of cutin with suberin-like monomers. Proc. Natl. Acad. Sci. USA 2007, 104, 18339–18344. [Google Scholar] [CrossRef] [Green Version]
- Melotto, M.; Underwood, W.; Koczan, J.; Nomura, K.; He, S.Y. Plant stomata function in innate immunity against bacterial invasion. Cell 2006, 126, 969–980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gudesblat, G.E.; Torres, P.S.; Vojnov, A.A. Stomata and pathogens: Warfare at the gates. Plant Signal. Behav. 2009, 4, 1114–1116. [Google Scholar] [CrossRef] [PubMed]
- Shirakawa, M.; Hara-Nishimura, I. Specialized vacuoles of myrosin cells: Chemical defense strategy in Brassicales plants. Plant Cell Physiol. 2018, 59, 1309–1316. [Google Scholar] [CrossRef] [PubMed]
- Ohashi-Ito, K.; Bergmann, D.C. Arabidopsis FAMA controls the final proliferation/differentiation switch during stomatal development. Plant Cell 2006, 18, 2493–2505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chadchawan, S.; Bishop, J.; Thangstad, O.P.; Bones, A.M.; Mitchell-Olds, T.; Bradley, D. Arabidopsis cDNA sequence encoding myrosinase. Plant Physiol. 1993, 103, 671–672. [Google Scholar] [CrossRef] [Green Version]
- Xue, J.; Jørgensen, M.; Pihlgren, U.; Rask, L. The myrosinase gene family in Arabidopsis thaliana: Gene organization, expression and evolution. Plant Mol. Biol. 1995, 27, 911–922. [Google Scholar] [CrossRef]
- Barth, C.; Jander, G. Arabidopsis Myrosinases TGG1 and TGG2 have redundant function in glucosinolate breakdown and insect defense. Plant J. 2006, 46, 549–562. [Google Scholar] [CrossRef]
- Zhao, Z.; Zhang, W.; Stanley, B.A.; Assmann, S.M. Functional proteomics of Arabidopsis thaliana guard cells uncovers new stomatal signaling pathways. Plant Cell 2008, 20, 3210–3226. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Su, H.; Zhou, J.; Liang, W.; Liu, D.; Li, J. Overexpressing the myrosinase gene TGG1 enhances stomatal defense against Pseudomonas syringae and delays flowering in Arabidopsis. Front. Plant Sci. 2019, 10, 1230. [Google Scholar] [CrossRef]
- Islam, M.M.; Tani, C.; Watanabe-Sugimoto, M.; Uraji, M.; Jahan, M.S.; Masuda, C.; Nakamura, Y.; Mori, I.C.; Murata, Y. Myrosinases, TGG1 and TGG2, Redundantly function in ABA and MeJA signaling in Arabidopsis guard cells. Plant Cell Physiol. 2009, 50, 1171–1175. [Google Scholar] [CrossRef]
- Ahuja, I.; De Vos, R.C.H.; Rohloff, J.; Stoopen, G.M.; Halle, K.K.; Ahmad, S.J.N.; Hoang, L.; Hall, R.D.; Bones, A.M. Arabidopsis myrosinases link the glucosinolate-myrosinase system and the cuticle. Sci. Rep. 2016, 6, 38990. [Google Scholar] [CrossRef] [Green Version]
- Hopper, D.W.; Ghan, R.; Cramer, G.R. A rapid dehydration leaf assay reveals stomatal response differences in grapevine genotypes. Hortic. Res. 2014, 1, 2. [Google Scholar] [CrossRef] [Green Version]
- Scarpeci, T.E.; Zanor, M.I.; Valle, E.M. Estimation of stomatal aperture in Arabidopsis thaliana using silicone rubber imprints. Bio-Protocol 2017, 7, E2347. [Google Scholar] [CrossRef]
- Li, C.; Yan, J.-M.; Li, Y.-Z.; Zhang, Z.-C.; Wang, Q.-L.; Liang, Y. Silencing the SpMPK1, SpMPK2, and SpMPKk3 genes in tomato reduces abscisic acid—Mediated drought tolerance. Int. J. Mol. Sci. 2013, 14, 21983–21996. [Google Scholar] [CrossRef] [Green Version]
- Dong, J.; Bergmann, D.C. Stomatal Patterning and Development. In Current Topics in Developmental Biology; Timmermans, M.C.P., Ed.; Academic Press: Cambridge, UK, 2010; Volume 91, Chapter 9; pp. 267–297. [Google Scholar]
- Falbel, T.G.; Koch, L.M.; Nadeau, J.A.; Segui-Simarro, J.M.; Sack, F.D.; Bednarek, S.Y. SCD1 is required for cell cytokinesis and polarized cell expansion in Arabidopsis thaliana. Development 2003, 130, 4011–4024. [Google Scholar] [CrossRef] [Green Version]
- Ahuja, I.; Borgen, B.H.; Hansen, M.; Honne, B.I.; Müller, C.; Rohloff, J.; Rossiter, J.T.; Bones, A.M. Oilseed rape seeds with ablated defence cells of the glucosinolate–myrosinase system. Production and characteristics of double haploid MINELESS plants of Brassica napus L. J. Exp. Bot. 2011, 62, 4975–4993. [Google Scholar] [CrossRef] [Green Version]
- Hunt, L.; Amsbury, S.; Baillie, A.L.; Movahedi, M.; Mitchell, A.; Afsharinafar, M.; Swarup, K.; Denyer, T.; Hobbs, J.; Swarup, R.; et al. Formation of the stomatal outer cuticular ledge requires a guard cell wall proline-rich protein. Plant Physiol. 2017, 174, 689–699. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Nadeau, J.A.; Zhao, L.; Sack, F.D. Characterization of a cytokinesis defective (cyd1) mutant of Arabidopsis. J. Exp. Bot. 1999, 50, 1437–1446. [Google Scholar] [CrossRef]
- Vaughan, J.G.; Gordon, E.; Robinson, D. The identification of myrosinase after the electrophoresis of Brassica and Sinapis seed proteins. Phytochemistry 1968, 7, 1345–1348. [Google Scholar] [CrossRef]
- Gonda, S.; Szűcs, Z.; Plaszkó, T.; Cziáky, Z.; Kiss-Szikszai, A.; Vasas, G.; Hamvas, H.M. A Simple method for on-gel detection of myrosinase activity. Molecules 2018, 23, 2204. [Google Scholar] [CrossRef] [Green Version]
- Borgen, B.H.; Thangstad, O.P.; Ahuja, I.; Rossiter, J.T.; Bones, A.M. Removing the mustard oil bomb from seeds: Transgenic ablation of myrosin cells in oilseed rape (Brassica napus) produces MINELESS seeds. J. Exp. Bot. 2010, 61, 1683–1697. [Google Scholar] [CrossRef]
- Eisele, J.F.; Fäßler, F.; Bürgel, P.F.; Chaban, C. A rapid and simple method for microscopy-based stomata analyses. PLoS ONE 2016, 11, E0164576. [Google Scholar] [CrossRef]
- Cambell, L.M.; Stevenson, D.W. Anatomía vegetativa de Aratitiyopea lopezii (Xyridaceae). Acta Bot. Venez. Online 2005, 28, 395–408. [Google Scholar]
- Lü, S.; Zhao, H.; Des Marais, D.L.; Parsons, E.P.; Wen, X.; Xu, X.; Bangarusamy, D.K.; Wang, G.; Rowland, O.; Juenger, T.; et al. Arabidopsis ECERIFERUM9 involvement in cuticle formation and maintenance of plant water status. Plant Physiol. 2012, 159, 930–944. [Google Scholar] [CrossRef] [Green Version]
- Tang, J.; Yang, X.; Xiao, C.; Li, J.; Chen, Y.; Li, R.; Li, S.; Lü, S.; Hu, H. GDSL lipase occluded stomatal pore 1 is required for wax biosynthesis and stomatal cuticular ledge formation. New Phytol. 2020, 228, 1880–1896. [Google Scholar] [CrossRef]
- Willmer, C.; Fricker, M. Stomata; Springer Science & Business Media: Berlin, Germany, 1996. [Google Scholar]
- Hunt, L.; Gray, J.E. How the stomate got his pore: Very long chain fatty acids and a structural cell wall protein sculpt the guard cell outer cuticular ledge. New Phytol. 2020, 228, 1698–1700. [Google Scholar] [CrossRef]
- Kosma, D.K.; Jenks, M.A. Eco-physiological and molecular-genetic determinants of plant cuticle function in drought and salt stress tolerance. In Advances in Molecular Breeding toward Drought and Salt Tolerant Crops; Jenks, M.A., Hasegawa, P.M., Jain, S.M., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 91–120. [Google Scholar]
- Hoch, H.C.; Staples, R.C.W.B.; Comeau, J.; Wolf, E.D. Signaling for growth orientation and cell differentiation by surface topography in Uromyces. Science 1987, 235, 1659–1662. [Google Scholar] [CrossRef]
- Pautov, A.; Bauer, S.; Ivanova, O.; Krylova, E.; Sapach, Y.; Gussarova, G. Role of the outer stomatal ledges in the mechanics of guard cell movements. Trees 2017, 31, 125–135. [Google Scholar] [CrossRef]
- Ueda, H.; Nishiyama, C.; Shimada, T.; Koumoto, Y.; Hayashi, Y.; Kondo, M.; Takahashi, T.; Ohtomo, I.; Nishimura, M.; Hara-Nishimura, I. AtVAM3 is required for normal specification of idioblasts, myrosin cells. Plant Cell Physiol. 2006, 47, 164–175. [Google Scholar] [CrossRef] [Green Version]
- Brembu, T.; Winge, P.; Seem, M.; Bones, A.M. NAPP and PIRP encode subunits of a putative wave regulatory protein complex involved in plant cell morphogenesis. Plant Cell 2004, 16, 2335–2349. [Google Scholar] [CrossRef] [Green Version]
- He, Z.-H.; Cheeseman, I.; He, D.; Kohorn, B.D. A cluster of five cell wall-associated receptor kinase genes, Wak1–5, are expressed in specific organs of Arabidopsis. Plant Mol. Biol. 1999, 39, 1189–1196. [Google Scholar] [CrossRef]
- Saito, C.; Ueda, T.; Abe, H.; Wada, Y.; Kuroiwa, T.; Hisada, A.; Furuya, M.; Nakano, A. A complex and mobile structure forms a distinct subregion within the continuous vacuolar membrane in young cotyledons of Arabidopsis. Plant J. 2002, 29, 245–255. [Google Scholar] [CrossRef]
- Zhao, L.; Sack, F.D. Ultrastructure of stomatal development in Arabidopsis (Brassicaceae) leaves. Am. J. Bot. 1999, 86, 929–939. [Google Scholar] [CrossRef] [Green Version]
- Lascève, G.; Leymarie, J.; Vavasseur, A. Alterations in light-induced stomatal opening in a starch-deficient mutant of Arabidopsis thaliana l. deficient in chloroplast phosphoglucomutase activity. Plant Cell Environ. 1997, 20, 350–358. [Google Scholar] [CrossRef] [Green Version]
- Lai, L.B.; Nadeau, J.A.; Lucas, J.; Lee, E.-K.; Nakagawa, T.; Zhao, L.; Geisler, M.; Sack, F.D. The Arabidopsis R2R3 MYB proteins FOUR LIPS and MYB88 restrict divisions late in the stomatal cell lineage. Plant Cell 2005, 17, 2754–2767. [Google Scholar] [CrossRef] [Green Version]
- Merced, A.; Renzaglia, K.S. Contrasting pectin polymers in guard cell walls of Arabidopsis and the hornwort Phaeoceros reflect physiological differences. Ann. Bot. 2018, 123, 579–585. [Google Scholar] [CrossRef]
- Jørgensen, L.B. Myrosin cells and dilated cisternae of the endoplasmic reticulum in the order Capparales. Nord. J. Bot. 1981, 1, 433–445. [Google Scholar] [CrossRef]
- Gao, X.-Q.; Li, C.-G.; Wei, P.-C.; Zhang, X.-Y.; Chen, J.; Wang, X.-C. The dynamic changes of tonoplasts in guard cells are important for stomatal movement in Vicia faba. Plant Physiol. 2005, 139, 1207–1216. [Google Scholar] [CrossRef] [Green Version]
- Shope, J.C.; Dewald, D.B.; Mott, K.A. Changes in surface area of intact guard cells are correlated with membrane internalization. Plant Physiol. 2003, 133, 1314–1321. [Google Scholar] [CrossRef] [Green Version]
- Couot-Gastelier, J.; Laffray, D.; Louguet, P. Étude comparée de l’ultrastructure des stomates ouverts et fermés chez le Tradescantia virginiana. Can. J. Bot. 1984, 62, 1505–1512. [Google Scholar] [CrossRef]
- Kerstiens, G. Water transport in plant cuticles: An update. J. Exp. Bot. 2006, 57, 2493–2499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schönherr, J.; Bukovac, M.J. Penetration of stomata by liquids: Dependence on surface tension, wettability, and stomatal morphology. Plant Physiol. 1972, 49, 813–819. [Google Scholar] [CrossRef] [PubMed]
- Yan, C.; Shen, H.; Li, Q.; He, Z. A novel ABA-hypersensitive mutant in Arabidopsis defines a genetic locus that confers tolerance to xerothermic stress. Planta 2006, 224, 889–899. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, J.I.; Kwak, J.M.; Allen, G.J. Guard cell abscisic acid signalling and engineering drought hardiness in plants. Nature 2001, 410, 327–330. [Google Scholar] [CrossRef]
- Koh, J.; Chen, G.; Yoo, M.-J.; Zhu, N.; Dufresne, D.; Erickson, J.E.; Shao, H.; Chen, S. Comparative proteomic analysis of Brassica napus in response to drought stress. J. Proteome Res. 2015, 14, 3068–3081. [Google Scholar] [CrossRef]
- Santelia, D.; Lunn, J.E. Transitory starch metabolism in guard cells: Unique features for a unique function. Plant Physiol. 2017, 174, 539–549. [Google Scholar] [CrossRef] [Green Version]
- Horrer, D.; Flütsch, S.; Pazmino, D.; Matthews, J.S.; Thalmann, M.; Nigro, A.; Leonhardt, N.; Lawson, T.; Santelia, D. Blue light induces a distinct starch degradation pathway in guard cells for stomatal opening. Curr. Biol. 2016, 26, 362–370. [Google Scholar] [CrossRef] [Green Version]
- Mouravieff, J. Action de la lumiere sur la cellule vegetale. I Production du mouvement d’ouverture stomatique par la lumiere de differentes regions du spectre. Bull. De La Soc. Bot. De Fr. 1958, 105, 467. [Google Scholar] [CrossRef]
- Yamada, K.; Hasegawa, T.; Minami, E.; Shibuya, N.; Kosemura, S.; Yamamura, S.; Hasegawa, K. Induction f myrosinase gene expression and myrosinase activity in radish hypocotyls by phototropic stimulation. J. Plant Physiol. 2003, 160, 255–259. [Google Scholar] [CrossRef]
- Hasegawa, T.; Yamada, K.; Kosemura, S.; Yamamura, S.; Hasegawa, K. Phototropic stimulation induces the conversion of glucosinolate to phototropism-regulating substances of radish hypocotyls. Phytochemistry 2000, 54, 275–279. [Google Scholar] [CrossRef]
- Chhajed, S.; Misra, B.B.; Tello, N.; Chen, S. Chemodiversity of the glucosinolate-myrosinase system at the single cell type resolution. Front. Plant Sci. 2019, 10, 618. [Google Scholar] [CrossRef] [PubMed]
- Munemasa, S.; Hauser, F.; Park, J.; Waadt, R.; Brandt, B.; Schroeder, J.I. Mechanisms of abscisic acid-mediated control of stomatal aperture. Curr. Opin. Plant Biol. 2015, 28, 154–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rhaman, M.S.; Nakamura, T.; Nakamura, Y.; Munemasa, S.; Murata, Y. The myrosinases TGG1 and TGG2 function redundantly in reactive carbonyl species signaling in Arabidopsis guard cells. Plant Cell Physiol. 2020, 61, 967–977. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Assmann, S.M. Metabolic signatures in response to abscisic acid (ABA) treatment in Brassica napus guard cells revealed by metabolomics. Sci. Rep. 2017, 7, 12875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khokon, M.A.R.; Jahan, M.S.; Rahman, T.; Hossain, M.A.; Muroyama, D.; Minami, I.; Munemasa, S.; Mori, I.C.; Nakamura, Y.; Murata, Y. Allyl Isothiocyanate (AITC) induces stomatal closure in Arabidopsis. Plant Cell Environ. 2011, 34, 1900–1906. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahuja, I.; Kissen, R.; Hoang, L.; Sporsheim, B.; Halle, K.K.; Wolff, S.A.; Ahmad, S.J.N.; Ahmad, J.N.; Bones, A.M. The Imaging of Guard Cells of thioglucosidase (tgg) Mutants of Arabidopsis Further Links Plant Chemical Defence Systems with Physical Defence Barriers. Cells 2021, 10, 227. https://doi.org/10.3390/cells10020227
Ahuja I, Kissen R, Hoang L, Sporsheim B, Halle KK, Wolff SA, Ahmad SJN, Ahmad JN, Bones AM. The Imaging of Guard Cells of thioglucosidase (tgg) Mutants of Arabidopsis Further Links Plant Chemical Defence Systems with Physical Defence Barriers. Cells. 2021; 10(2):227. https://doi.org/10.3390/cells10020227
Chicago/Turabian StyleAhuja, Ishita, Ralph Kissen, Linh Hoang, Bjørnar Sporsheim, Kari K. Halle, Silje Aase Wolff, Samina Jam Nazeer Ahmad, Jam Nazeer Ahmad, and Atle M. Bones. 2021. "The Imaging of Guard Cells of thioglucosidase (tgg) Mutants of Arabidopsis Further Links Plant Chemical Defence Systems with Physical Defence Barriers" Cells 10, no. 2: 227. https://doi.org/10.3390/cells10020227