
Graphene Oxide–Silver Nanoparticle Nanocomposites Induce Oxidative Stress and Aberrant Methylation in Caprine Fetal Fibroblast Cells

 ,
,  , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Synthesis and Characterization of GO-AgNPs
2.3. Cell Culture
2.4. Cell Viability Assay
2.5. Cell Morphology
2.6. Annexin V–FITC/PI Staining Assay
2.7. Measurement of ROS Production
2.8. Measurement of Total Superoxide Dismutase (SOD) Enzyme Activity
2.9. Measurement of Malondialdehyde (MDA) Production
2.10. Measurement of Lactate Dehydrogenase (LDH) Production
2.11. Measurement of Caspase-3 Activity
2.12. Determination of Global 5-mC
2.13. Quantitative Reverse Transcription PCR (RT-qPCR) Analysis
2.14. Statistical Analysis
3. Results
3.1. Characterization of GO-AgNPs
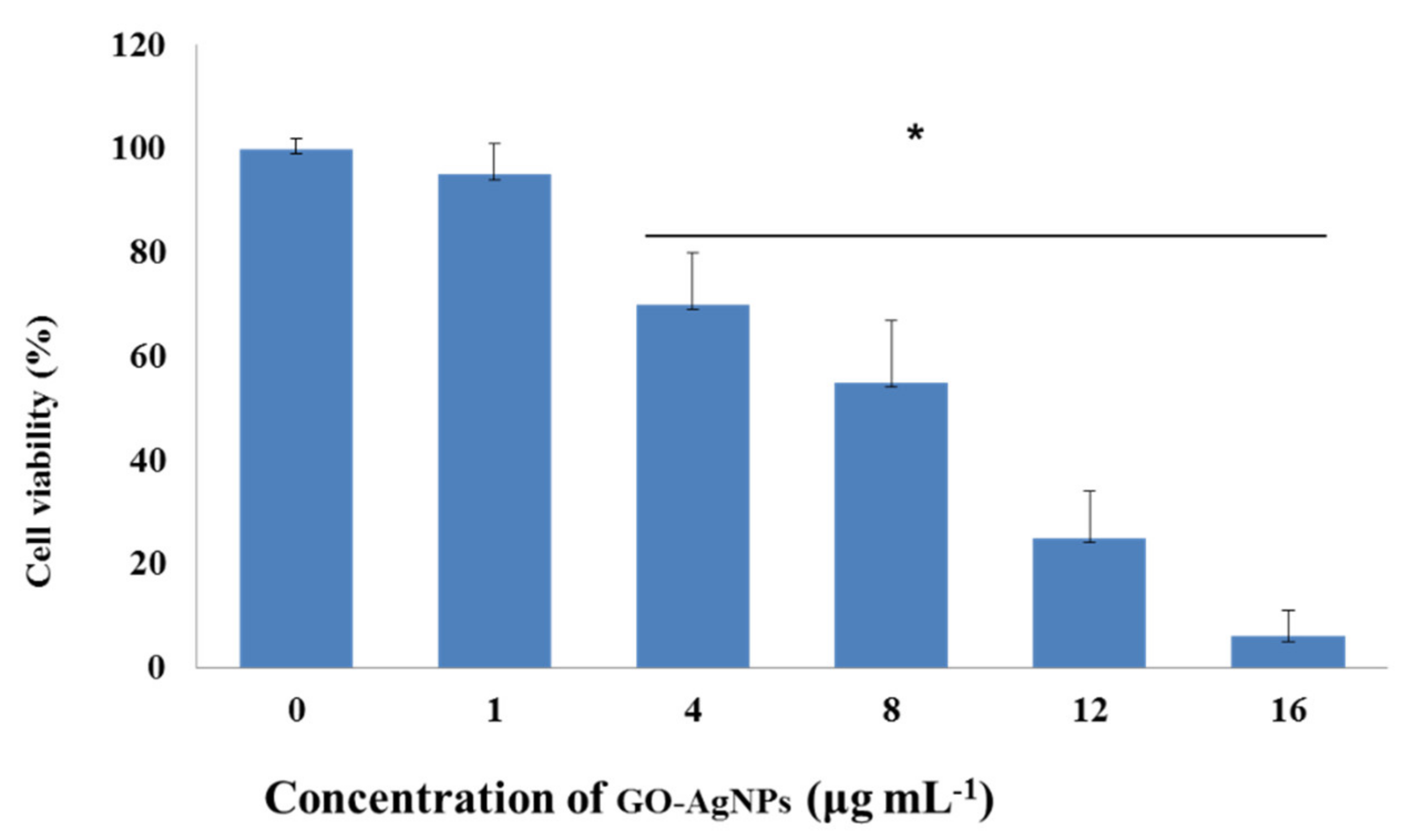
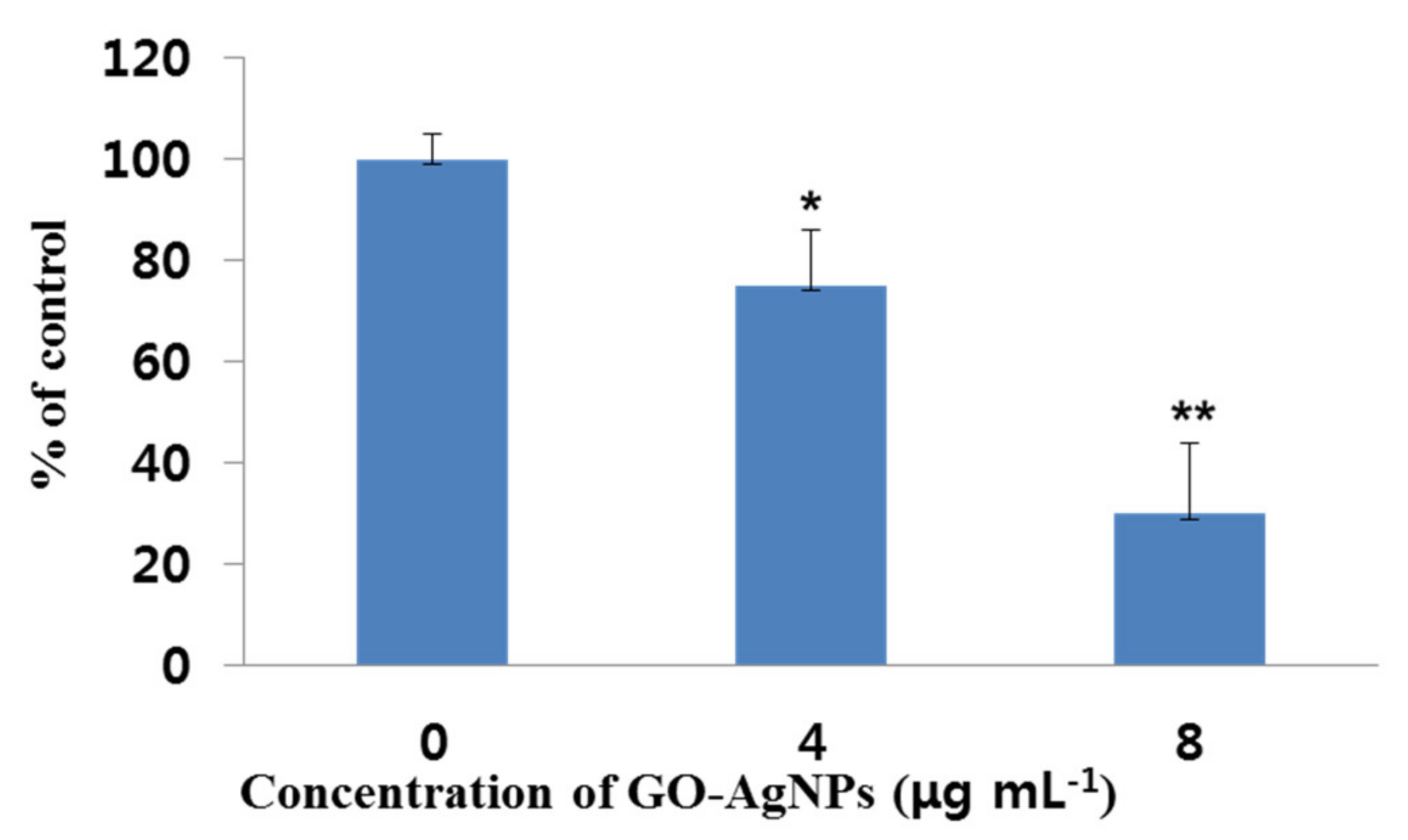
3.2. Effect of GO-AgNPs on Caprine Fetal Fibroblast Cell (CFFC) Viability
3.3. Effect of GO-AgNPs on Cell Morphology
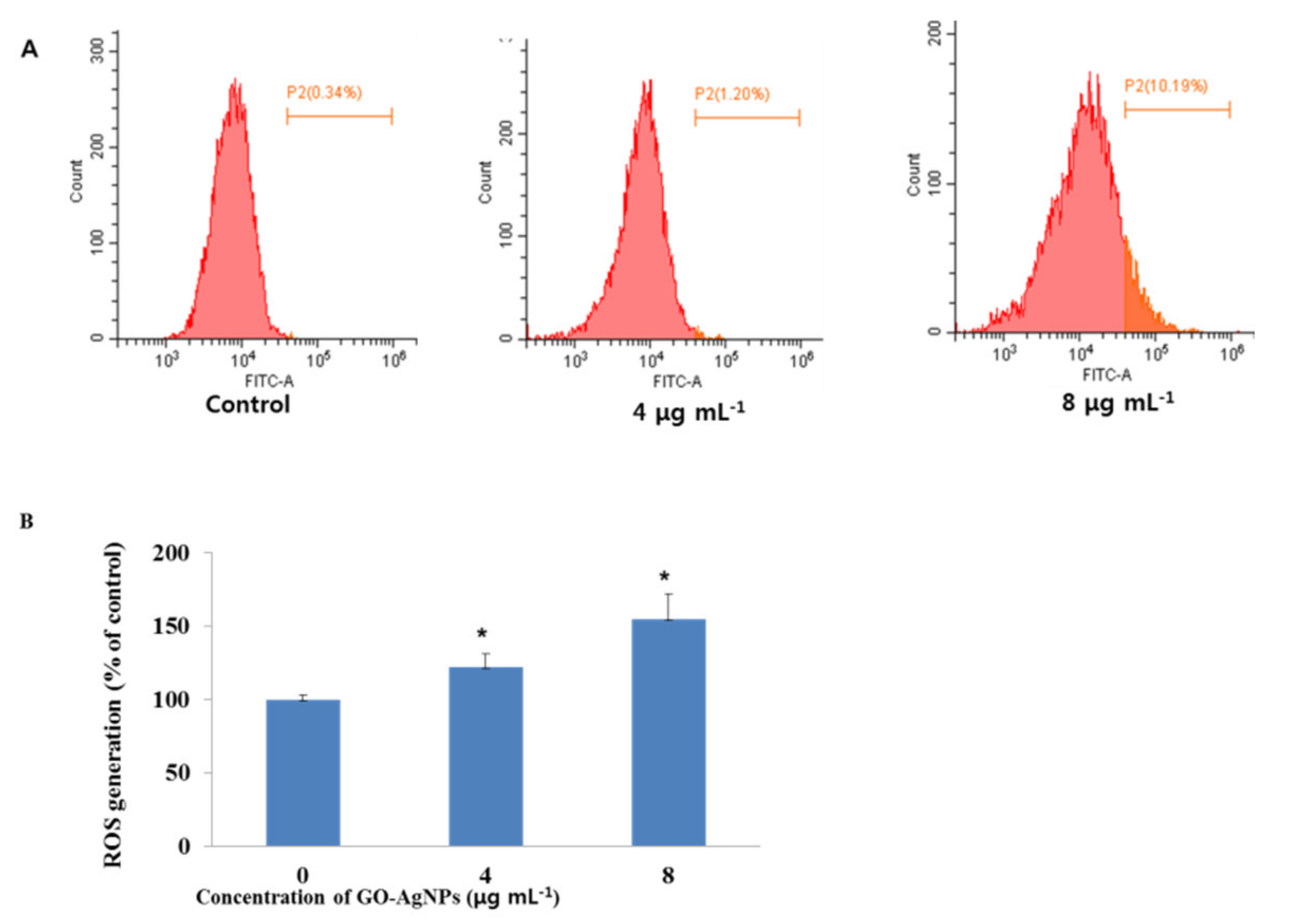
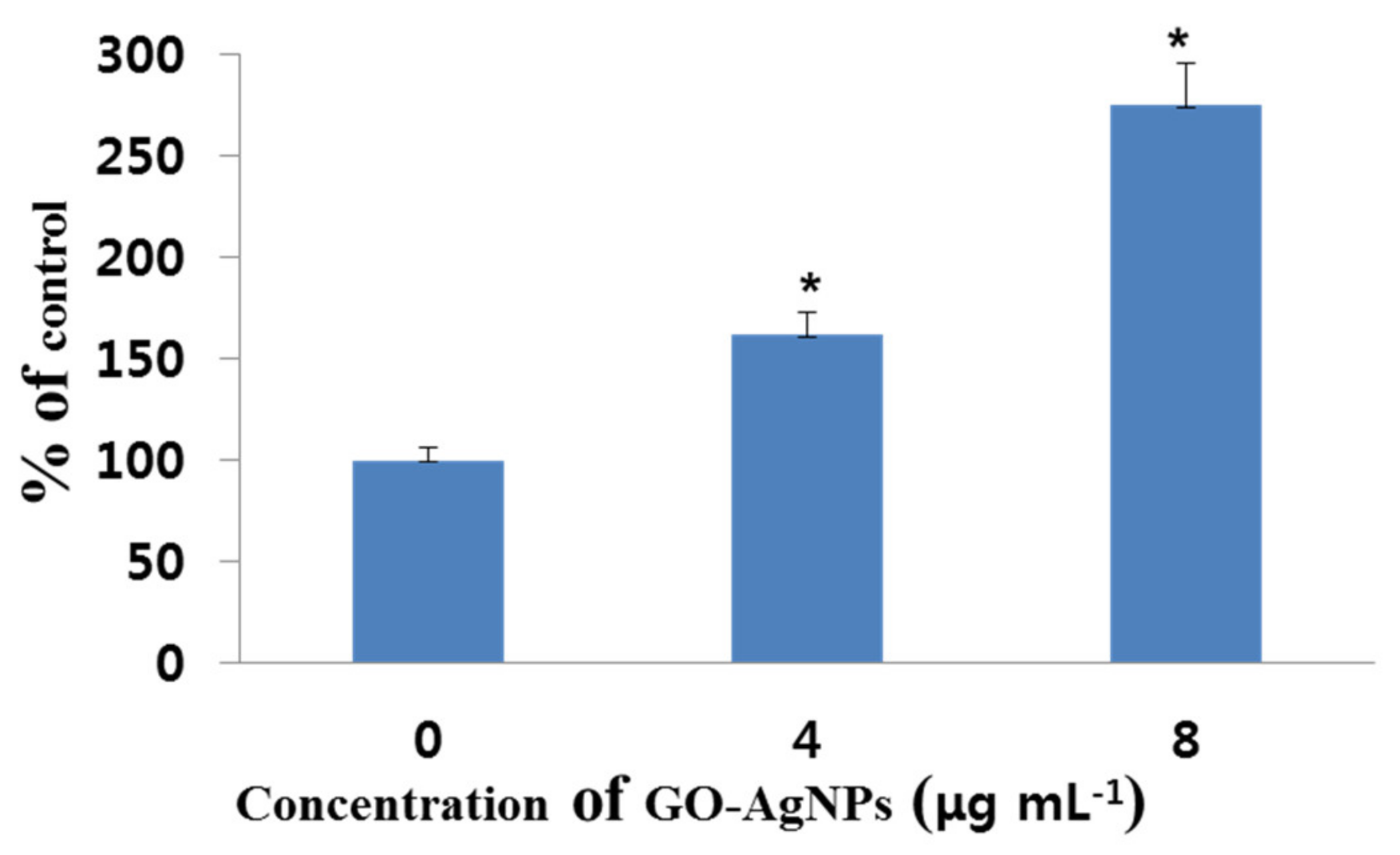
3.4. Effect of GO-AgNPs on Reactive Oxygen Species (ROS) Production
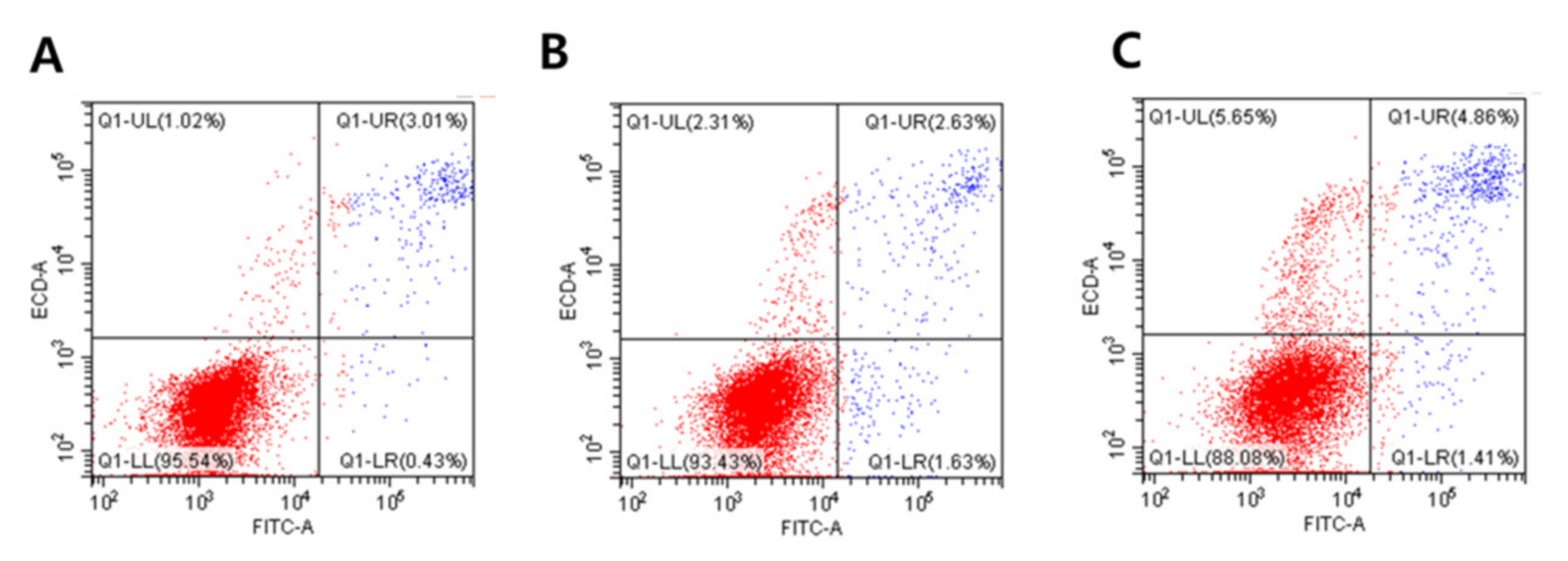
3.5. Effects of GO-AgNPs on Apoptosis
3.6. Effects of GO-AgNPs on SOD Production
3.7. Effects of GO-AgNPs on MDA Production
3.8. Effects of GO-AgNPs on LDH
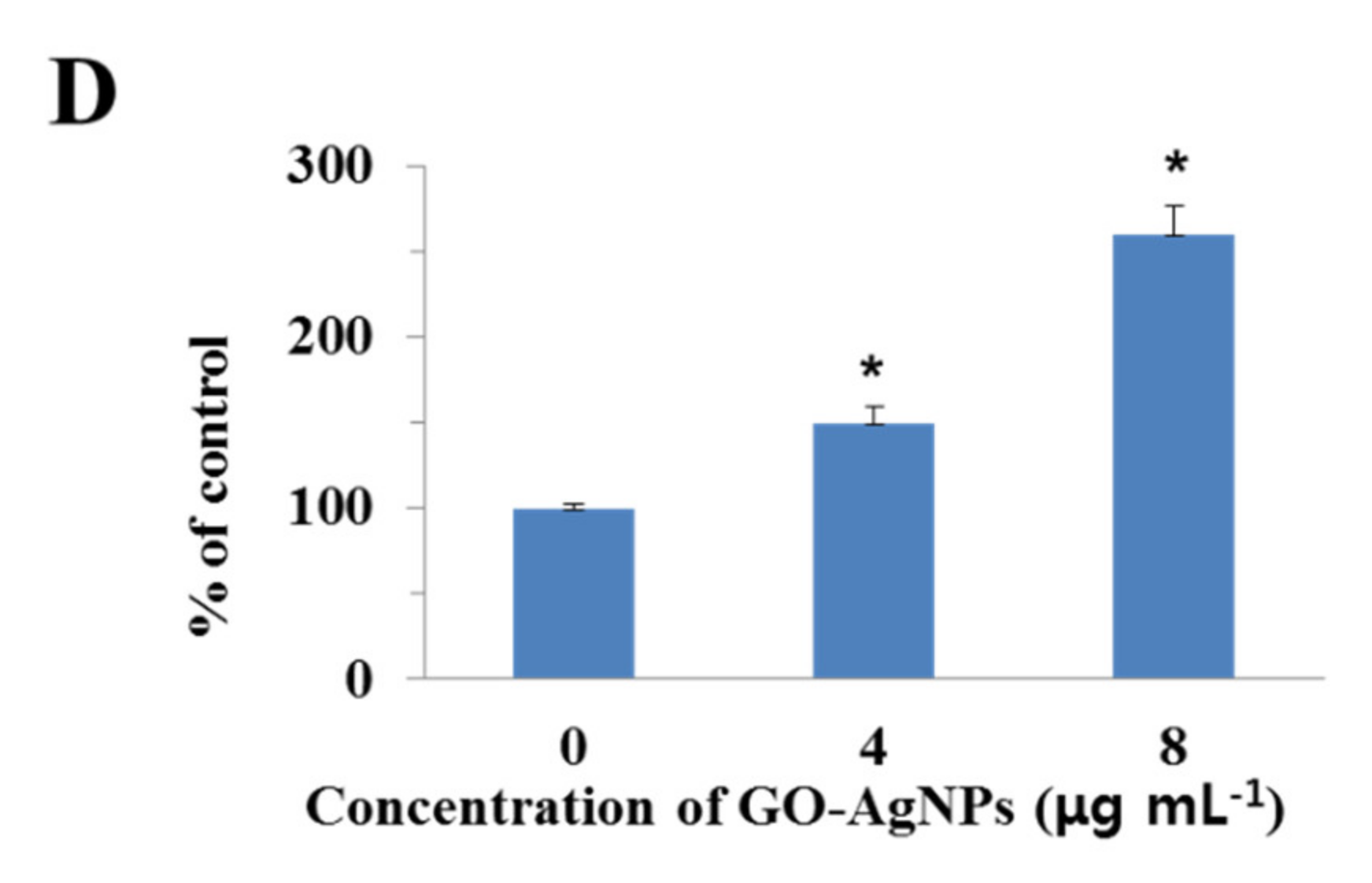
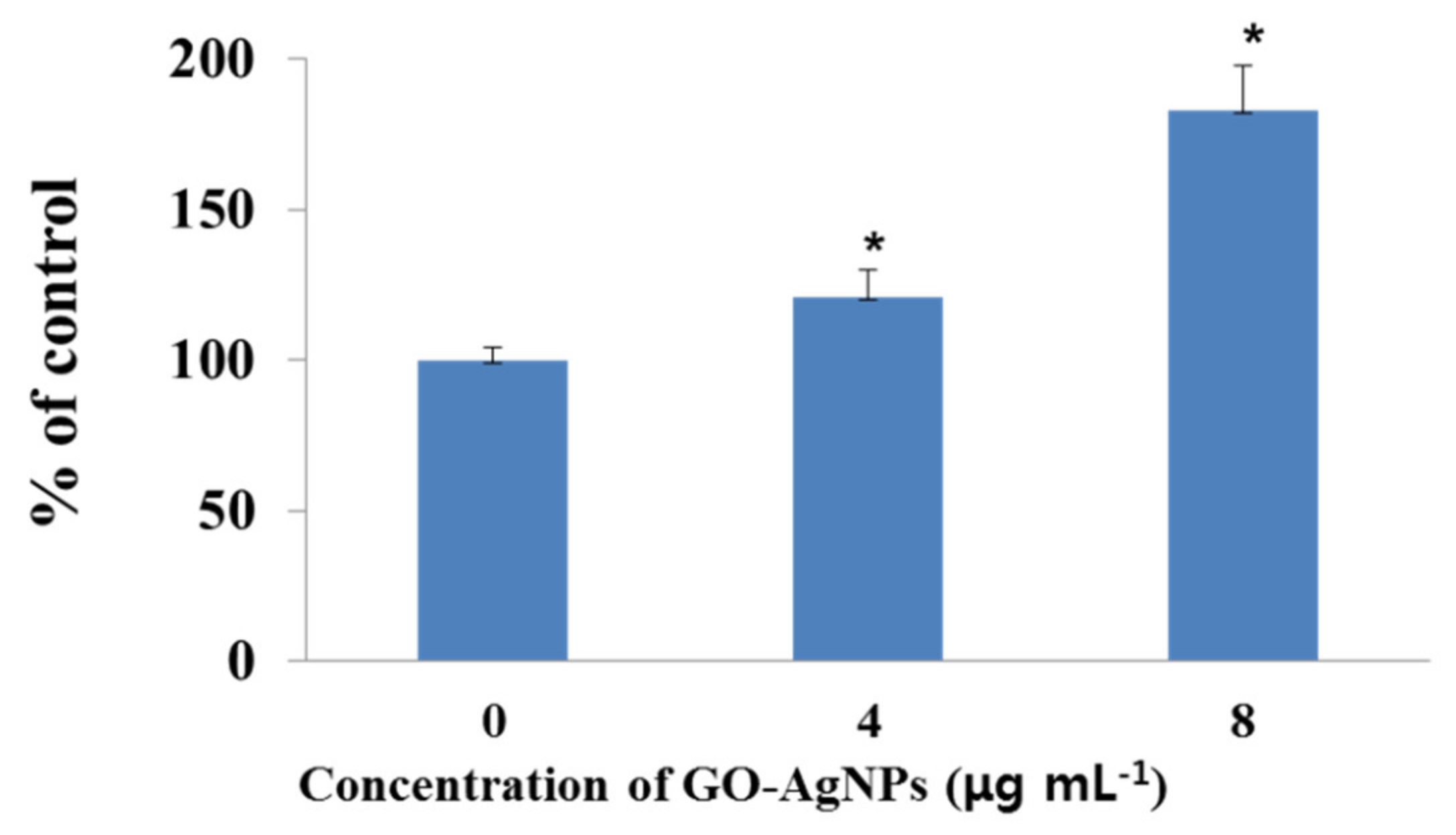
3.9. Effects of GO-AgNPs on the Caspase-3 Activity
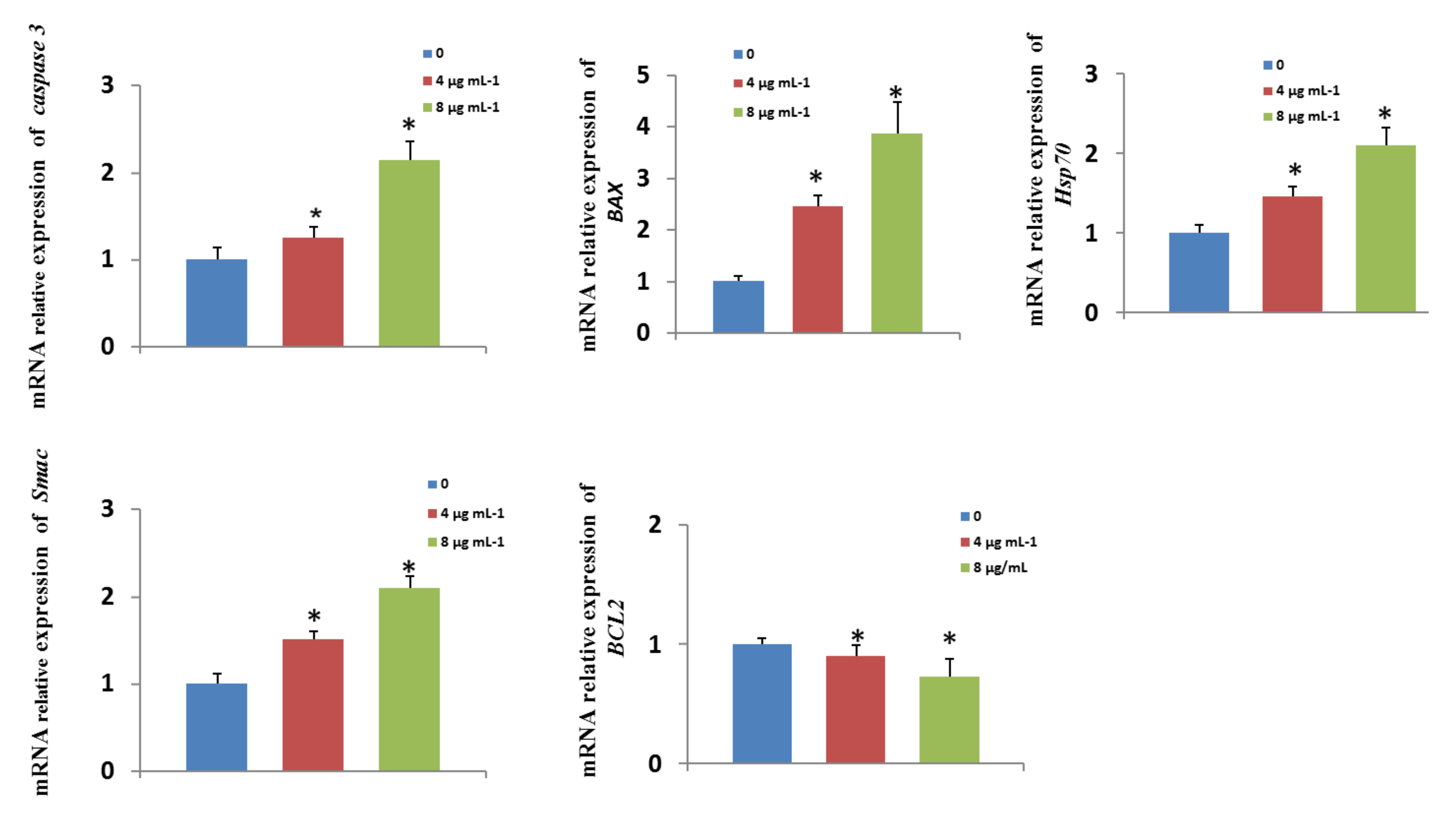
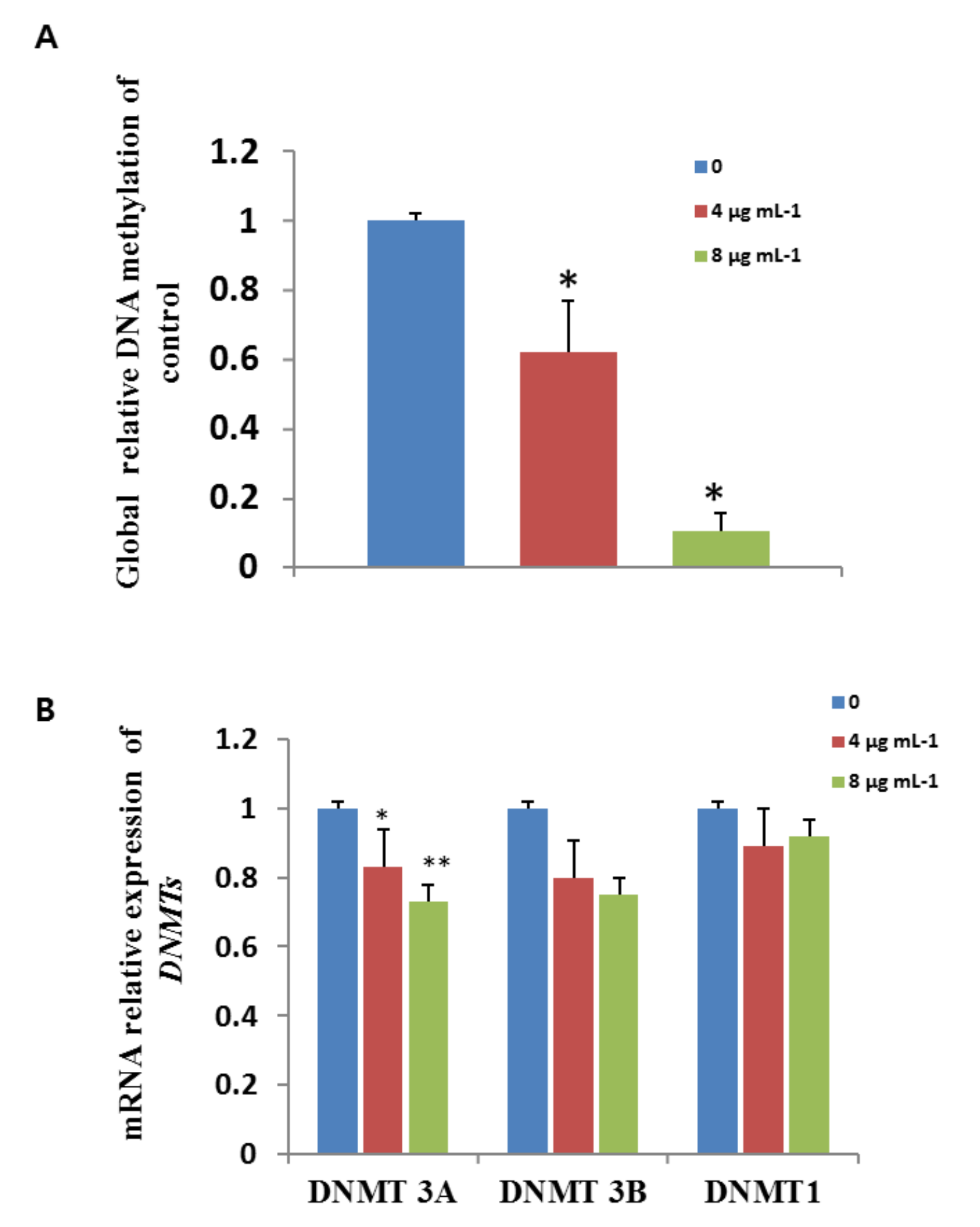
3.10. Effects of GO-AgNPs on Gene Expression
3.11. Effects of GO-AgNPs on Global DNA Methylation
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pinho, A.R.; Martins, F.; Costa, M.; Senos, A.; Silva, O.; Pereira, M.L.; Rebelo, S. In vitro cytotoxicity effects of zinc oxide nanoparticles on spermatogonia cells. Cells 2020, 9, 1081. [Google Scholar] [CrossRef] [PubMed]
- Gurunathan, S.; Kim, J.H. Graphene oxide-silver nanoparticles nanocomposite stimulates differentiation in human neuroblastoma cancer cells (SH-SY5Y). Int. J. Mol. Sci. 2017, 18, 2549. [Google Scholar] [CrossRef] [PubMed]
- Gies, V.; Zou, S. Systematic toxicity investigation of graphene oxide: Evaluation of assay selection, cell type, exposure period and flake size. Toxicol. Res. 2018, 7, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Gurunathan, S.; Han, J.W.; Eppakayala, V.; Dayem, A.A.; Kwon, D.N.; Kim, J.H. Biocompatibility effects of biologically synthesized graphene in primary mouse embryonic fibroblast cells. Nanoscale. Res. Lett. 2013, 8, 393. [Google Scholar] [CrossRef]
- Thapa, R.K.; Kim, J.H.; Jeong, J.H.; Shin, B.S.; Choi, H.G.; Yong, C.S.; Kim, J.O. Silver nanoparticle-embedded graphene oxide-methotrexate for targeted cancer treatment. Colloids. Surf. B. Biointerfaces 2017, 153, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Syama, S.; Paul, W.; Sabareeswaran, A.; Mohanan, P.V. Raman spectroscopy for the detection of organ distribution and clearance of PEGylated reduced graphene oxide and biological consequences. Biomaterials 2017, 131, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Nurunnabi, M.; Khatun, Z.; Huh, K.M.; Park, S.Y.; Lee, D.Y.; Cho, K.J.; Lee, Y.K. In vivo biodistribution and toxicology of carboxylated graphene quantum dots. ACS. Nano 2013, 7, 6858–6867. [Google Scholar] [CrossRef]
- Ivask, A.; Voelcker, N.H.; Seabrook, S.A.; Hor, M.; Kirby, J.K.; Fenech, M.; Davis, T.P.; Ke, P.C. DNA melting and genotoxicity induced by silver nanoparticles and graphene. Chem. Res. Toxicol. 2015, 28, 1023–1035. [Google Scholar] [CrossRef] [PubMed]
- Hyun, J.S.; Lee, B.S.; Ryu, H.Y.; Sung, J.H.; Chung, K.H.; Yu, I.J. Effects of repeated silver nanoparticles exposure on the histological structure and mucins of nasal respiratory mucosa in rats. Toxicol. Lett. 2008, 182, 24–28. [Google Scholar] [CrossRef] [PubMed]
- Rosas-Hernandez, H.; Jimenez-Badillo, S.; Martinez-Cuevas, P.P.; Gracia-Espino, E.; Terrones, H.; Terrones, M.; Hussain, S.M.; Ali, S.F.; González, C. Effects of 45-nm silver nanoparticles on coronary endothelial cells and isolated rat aortic rings. Toxicol. Lett. 2009, 191, 305–313. [Google Scholar] [CrossRef] [PubMed]
- Rezvani, E.; Rafferty, A.; McGuinness, C.; Kennedy, J. Adverse effects of nanosilver on human health and the environment. Acta. Biomater. 2019, 94, 145–159. [Google Scholar] [CrossRef]
- Jia, P.P.; Sun, T.; Junaid, M.; Yang, L.; Ma, Y.B.; Cui, Z.S.; Wei, D.P.; Shi, H.F.; Pei, D.S. Nanotoxicity of different sizes of graphene (G) and graphene oxide (GO) in vitro and in vivo. Environ. Pollut. 2019, 247, 595–606. [Google Scholar] [CrossRef] [PubMed]
- Seabra, A.B.; Paula, A.J.; de Lima, R.; Alves, O.L.; Durán, N. Nanotoxicity of graphene and graphene oxide. Chem. Res. Toxicol. 2014, 27, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Pogribna, M.; Hammons, G. Epigenetic Effects of nanomaterials and nanoparticles. J. Nanobiotechnol. 2021, 19, 2. [Google Scholar] [CrossRef]
- Uzair, B.; Liaqat, A.; Iqbal, H.; Menaa, B.; Razzaq, A.; Thiripuranathar, G.; Rana, N.F.; Menaa, F. Green and cost-effective synthesis of metallic nanoparticles by algae: Safe methods for translational medicine. Bioengineering 2020, 7, 129. [Google Scholar] [CrossRef]
- Gurunathan, S.; Han, J.W.; Park, J.H.; Kim, E.; Choi, Y.J.; Kwon, D.N.; Kim, J.H. Reduced graphene oxide-silver nanoparticle nanocomposite: A potential anticancer nanotherapy. Int. J. Nanomed. 2015, 10, 6257–6276. [Google Scholar] [CrossRef]
- Gurunathan, S.; Hyun Park, J.; Choi, Y.J.; Woong Han, J.; Kim, J.H. Synthesis of graphene oxide-silver nanoparticle nanocomposites: An efficient novel antibacterial agent. Curr. Nanosci. 2016, 12, 762–773. [Google Scholar] [CrossRef]
- Cobos, M.; De-La-Pinta, I.; Quindós, G.; Fernández, M.J.; Fernández, M.D. Graphene Oxide-Silver Nanoparticle Nanohybrids: Synthesis, Characterization, and Antimicrobial Properties. Nanomaterials 2020, 10, 376. [Google Scholar] [CrossRef] [PubMed]
- Akter, M.; Sikder, M.T.; Rahman, M.M.; Ullah, A.K.M.A.; Hossain, K.F.B.; Banik, S.; Hosokawa, T.; Saito, T.; Ku-rasaki, M. A systematic review on silver nanoparticles-induced cytotoxicity: Physicochemical properties and perspectives. J. Adv. Res. 2017, 9, 1–16. [Google Scholar] [CrossRef]
- Courtois, P.; Rorat, A.; Lemiere, S.; Guyoneaud, R.; Attard, E.; Levard, C.; Vandenbulcke, F. Ecotoxicology of silver nanoparticles and their derivatives introduced in soil with or without sewage sludge: A review of effects on microorganisms, plants and animals. Environ. Pollut. 2019, 253, 578–598. [Google Scholar] [CrossRef] [PubMed]
- Brami, C.; Glover, A.R.; Butt, K.R.; Lowe, C.N. Effects of silver nanoparticles on survival, biomass change and avoidance behaviour of the endogeic earthworm Allolobophora chlorotica. Ecotoxicol. Environ. Saf. 2017, 141, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Ong, C.; Lee, Q.Y.; Cai, Y.; Liu, X.; Ding, J.; Yung, L.Y.; Bay, B.H.; Baeg, G.H. Silver nanoparticles disrupt germline stem cell maintenance in the Drosophila testis. Sci. Rep. 2016, 6, 1–10. [Google Scholar] [CrossRef]
- Vadalasetty, K.P.; Lauridsen, C.; Engberg, R.M.; Vadalasetty, R.; Kutwin, M.; Chwalibog, A.; Sawosz, E. Influence of silver nanoparticles on growth and health of broiler chickens after infection with campylobacter jejuni. BMC Vet. Res. 2018, 14, 1. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.G.; Peng, Q.L.; Gurunathan, S. Effects of Silver Nanoparticles on Multiple Drug-Resistant Strains of Staphylococcus aureus and Pseudomonas aeruginosa from Mastitis-Infected Goats: An Alternative Approach for Antimicrobial Therapy. Int. J. Mol. Sci. 2017, 18, 569. [Google Scholar] [CrossRef]
- Zhang, X.F.; Park, J.H.; Choi, Y.J.; Kang, M.H.; Gurunathan, S.; Kim, J.H. Silver nanoparticles cause complica-tions in pregnant mice. Int. J. Nanomed. 2015, 10, 7057–7071. [Google Scholar]
- Yuan, Y.G.; Wang, Y.H.; Xing, H.H.; Gurunathan, S. Quercetin-mediated synthesis of graphene oxide-silver nanoparticle nanocomposites: A suitable alternative nanotherapy for neuroblastoma. Int. J. Nanomed. 2017, 12, 5819–5839. [Google Scholar] [CrossRef]
- Yuan, Y.G.; Song, S.Z.; Zhu, M.M.; He, Z.Y.; Lu, R.; Zhang, T.; Mi, F.; Wang, J.Y.; Cheng, Y. Human lactoferrin efficiently targeted into caprine beta-lactoglobulin locus with transcription activator-like effector nucleases. Asian-Australas. J. Anim. Sci. 2017, 30, 1175–1182. [Google Scholar]
- Yuan, Y.G.; Zhang, S.; Hwang, J.Y.; Kong, I.K. Silver nanoparticles potentiates cytotoxicity and apoptotic potential of camptothecin in human cervical cancer cells. Oxid. Med. Cell. Longev. 2018, 2018, 6121328. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.J.; Gurunathan, S.; Kim, J.H. Graphene oxide-silver nanocomposite enhances cytotoxic and apoptotic potential of salinomycin in human ovarian cancer stem cells (OvCSCs): A novel approach for cancer therapy. Int. J. Mol. Sci. 2018, 19, 710. [Google Scholar] [CrossRef]
- Gurunathan, S.; Kim, J.H. Synthesis, toxicity, biocompatibility, and biomedical applications of graphene and graphene-related materials. Int. J. Nanomed. 2016, 11, 1927–1945. [Google Scholar] [CrossRef]
- He, K.; Zeng, Z.; Chen, A.; Zeng, G.; Xiao, R.; Xu, P.; Huang, Z.; Shi, J.; Hu, L.; Chen, G. Advancement of ag-graphene based nanocomposites: An overview of synthesis and its applications. Small 2018, 14, e1800871. [Google Scholar] [CrossRef] [PubMed]
- Ali, D.; Alarifi, S.; Alkahtani, S.; Almeer, R.S. Silver-doped graphene oxide nanocomposite triggers cytotoxicity and apoptosis in human hepatic normal and carcinoma cells. Int. J. Nanomed. 2018, 13, 5685–5699. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.F.; Choi, Y.J.; Han, J.W.; Park, J.H.; Gurunathan, S.; Kim, J.H. Differential nanoreprotoxicity of silver nanoparticles in male somatic cells and spermatogonial stem cells. Int. J. Nanomed. 2015, 10, 1335–1357. [Google Scholar]
- Xu, F.; Piett, C.; Farkas, S.; Qazzaz, M.; Syed, N.I. Silver nanoparticles (AgNPs) cause degeneration of cytoskeleton and disrupt synaptic machinery of cultured cortical neurons. Mol. Brain. 2013, 6, 29. [Google Scholar] [CrossRef] [PubMed]
- Hondroulis, E.; Liu, C.; Li, C.Z. Whole cell based electrical impedance sensing approach for a rapid nanotoxicity assay. Nanotechnology 2010, 21, 315103. [Google Scholar] [CrossRef]
- Abdel-Mohsen, A.M.; Abdel-Rahman, R.M.; Fouda, M.M.; Vojtova, L.; Uhrova, L.; Hassan, A.F.; Al-Deyab, S.S.; El-Shamy, I.E.; Jancar, J. Preparation, characterization and cytotoxicity of schizophyllan/silver nanoparticle composite. Carbohydr. Polym. 2014, 102, 238–245. [Google Scholar] [CrossRef]
- Sahu, S.C.; Zheng, J.W.; Graham, L.; Chen, L.; Ihrie, J.; Yourick, J.J.; Sprando, R.L. Comparative cytotoxicity of nanosilver in human liver HepG2 and colon Caco2 cells in culture. J. Appl. Toxicol. 2014, 34, 1155–1166. [Google Scholar] [CrossRef]
- Lopes, I.M.D.; de Oliveira, I.M.; Bargi-Souza, P.; Cavallin, M.D.; Kolc, C.S.M.; Khalil, N.M.; Quináia, S.P.; Romano, M.A.; Romano, R.M. Effects of silver nanoparticle exposure to the testicular antioxidant system during the prepubertal rat stage. Chem. Res. Toxicol. 2019, 32, 986–994. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Wang, J.; Huang, Y.; Gao, X.; Zhang, T.; Tang, M.; Tang, M. Comparative cytotoxicity and apoptotic pathways induced by nanosilver in human liver HepG2 and L02 cells. Hum. Exp. Toxicol. 2018, 37, 1293–1309. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.G.; Peng, Q.L.; Gurunathan, S. Silver nanoparticles enhance the apoptotic potential of gemcitabine in human ovarian cancer cells: Combination therapy for effective cancer treatment. Int. J. Nanomed. 2017, 12, 6487–6502. [Google Scholar] [CrossRef]
- Lee, Y.H.; Cheng, F.Y.; Chiu, H.W.; Tsai, J.C.; Fang, C.Y.; Chen, C.W.; Wang, Y.J. Cytotoxicity, oxidative stress, apoptosis and the autophagic effects of silver nanoparticles in mouse embryonic fibroblasts. Biomaterials 2014, 35, 4706–4715. [Google Scholar] [CrossRef] [PubMed]
- De Luna, L.A.; de Moraes, A.C.; Consonni, S.R.; Pereira, C.D.; Cadore, S.; Giorgio, S.; Alves, O.L. Comparative in vitro toxicity of a graphene oxide-silver nanocomposite and the pristine counterparts toward macrophages. J. Nanobiotechnol. 2016, 14, 12. [Google Scholar] [CrossRef]
- El-Sonbaty, S.M. Fungus-mediated synthesis of silver nanoparticles and evaluation of antitumor activity. Cancer Nanotechnol. 2013, 4, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Liao, K.H.; Lin, Y.S.; Macosko, C.W.; Haynes, C.L. Cytotoxicity of graphene oxide and graphene in human erythrocytes and skin fibroblasts. ACS Appl. Mater. Interfaces 2011, 3, 2607–2615. [Google Scholar] [CrossRef]
- Yuan, Y.G.; Gurunathan, S. Combination of graphene oxide-silver nanoparticle nanocomposites and cisplatin enhances apoptosis and autophagy in human cervical cancer cells. Int. J. Nanomed. 2017, 12, 6537–6558. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.F.; Huang, F.H.; Zhang, G.L.; Bai, D.P.; Massimo, D.F.; Huang, Y.F.; Gurunathan, S. Novel biomolecule lycopene-reduced graphene oxide-silver nanoparticle enhances apoptotic potential of trichostatin A in human ovarian cancer cells (SKOV3). Int. J. Nanomed. 2017, 12, 7551–7575. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.G.; Xu, L.; Zhang, S.; Mesalam, A.; Lee, K.L.; Liu, H.; Joo, M.D.; Idrees, M.; Kong, I.K. Polydatin and I-CBP112 protects early bovine embryo against nicotinamide-induced mitochondrial dysfunction. Theriogenology 2019, 134, 1–10. [Google Scholar] [CrossRef]
- Fang, W.; Chi, Z.; Li, W.; Zhang, X.; Zhang, Q. Comparative study on the toxic mechanisms of medical nanosilver and silver ions on the antioxidant system of erythrocytes: From the aspects of antioxidant enzyme activities and molecular interaction mechanisms. J. Nanobiotechnol. 2019, 17, 66. [Google Scholar] [CrossRef] [PubMed]
- Mesalam, A.; Khan, I.; Lee, K.L.; Song, S.H.; Chowdhury, M.M.R.; Uddin, Z.; Park, K.H.; Kong, I.K. 2-methoxystypandrone improves in vitro-produced bovine embryo quality through inhibition of IKBKB. Theriogenology 2017, 99, 10–20. [Google Scholar] [CrossRef]
- Feder, M.E.; Hofmann, G.E. Heat-shock proteins, molecular chaperones, and the stress response: Evolutionary and ecological physiology. Annu. Rev. Physiol. 1999, 6, 243–282. [Google Scholar] [CrossRef]
- Zhang, W.; Liu, S.; Han, D.; He, Z. Engineered nanoparticle-induced epigenetic changes: An important consideration in nanomedicine. Acta Biomater. 2020, 117, 93–107. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, S.R.; Ordaz, J.; Lo, C.L.; Damayanti, N.P.; Zhou, F.; Irudayaraj, J. From the Cover: Zinc oxide Nanoparticles-Induced Reactive Oxygen Species Promotes Multimodal Cyto- and Epigenetic Toxicity. Toxicol. Sci. 2017, 156, 261–274. [Google Scholar] [PubMed]
- Zhang, X.F.; Gurunathan, S.; Kim, J.H. Effects of silver nanoparticles on neonatal testis development in mice. Int. J. Nanomed. 2015, 10, 6243–6256. [Google Scholar]
- Mytych, J.; Zebrowski, J.; Lewinska, A.; Wnuk, M. Prolonged Effects of Silver Nanoparticles on p53/p21 Pathway-Mediated Proliferation, DNA Damage Response, and Methylation Parameters in HT22 Hippocampal Neuronal Cells. Mol. Neurobiol. 2017, 54, 1285–1300. [Google Scholar] [CrossRef] [PubMed]
- Tajima, S.; Suetake, I.; Takeshita, K.; Nakagawa, A.; Kimura, H. Domain Structure of the Dnmt1, Dnmt3a, and Dnmt3b DNA Methyltransferases. Adv. Exp. Med. Biol. 2016, 945, 63–86. [Google Scholar] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer sequence (5‘–3‘) | Product Size (bp) |
|---|---|---|
| caspase-3 | F: CCATGGTGAAGAAGGAATCATTT R: TCCCCTCTGAAGAAACTTGCTAA | 78 |
| BAX | F: GCATCCACCAAGAAGCTGAG R: CCGCCACTCGGAAAAAGAC | 120 |
| Smac | F: TGTTCCAGTCGTGGCTAACTT R: AAAGACACAGCCCTCCTCATT | 171 |
| BCL2 | F: ATGTGTGTGGAGAGCGTCA R: AGAGACAGCCAGGAGAAATC | 113 |
| Hsp70 | F: TCAGGACTCAATCTGCATCG R: ATCCGCATTTCTGGTTATCA | 210 |
| DNMT3A DNMT3B DNMT1 β-actin | F: CTTGGAGAAGCGGAGTGAGC R: GTGCAGCAGCCATTCTCTACAG F: AGCCCCTACCTCACCATC R: CTGATACTCGGTGCTGTCTGC F: GAGGAGGCTGCCAAGGACT R: CAAACACCGCATACGACACAC F: TCACGGAGCGTGGCTACAG R: CCTTGATGTCACGGACGATTT | 138 156 134 127 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, Y.-G.; Cai, H.-Q.; Wang, J.-L.; Mesalam, A.; Md Talimur Reza, A.M.; Li, L.; Chen, L.; Qian, C. Graphene Oxide–Silver Nanoparticle Nanocomposites Induce Oxidative Stress and Aberrant Methylation in Caprine Fetal Fibroblast Cells. Cells 2021, 10, 682. https://doi.org/10.3390/cells10030682
Yuan Y-G, Cai H-Q, Wang J-L, Mesalam A, Md Talimur Reza AM, Li L, Chen L, Qian C. Graphene Oxide–Silver Nanoparticle Nanocomposites Induce Oxidative Stress and Aberrant Methylation in Caprine Fetal Fibroblast Cells. Cells. 2021; 10(3):682. https://doi.org/10.3390/cells10030682
Chicago/Turabian StyleYuan, Yu-Guo, He-Qing Cai, Jia-Lin Wang, Ayman Mesalam, Abu Musa Md Talimur Reza, Ling Li, Li Chen, and Chen Qian. 2021. "Graphene Oxide–Silver Nanoparticle Nanocomposites Induce Oxidative Stress and Aberrant Methylation in Caprine Fetal Fibroblast Cells" Cells 10, no. 3: 682. https://doi.org/10.3390/cells10030682
APA StyleYuan, Y.-G., Cai, H.-Q., Wang, J.-L., Mesalam, A., Md Talimur Reza, A. M., Li, L., Chen, L., & Qian, C. (2021). Graphene Oxide–Silver Nanoparticle Nanocomposites Induce Oxidative Stress and Aberrant Methylation in Caprine Fetal Fibroblast Cells. Cells, 10(3), 682. https://doi.org/10.3390/cells10030682