
Progressive and Coordinated Mobilization of the Skeletal Muscle Niche throughout Tissue Repair Revealed by Single-Cell Proteomic Analysis

 ,
, 
Abstract
:1. Background
2. Methods
2.1. Mouse Strains
2.2. Induction of Muscle Damage
2.3. Collection of Muscle Cells and CyTOF Antibody Staining
2.4. Processing of CyTOF samples
2.5. CyTOF Data Analysis
2.6. Single TA Muscle Dissociation and Immunostaining for Flow Cytometry Analysis
2.7. Statistical Analysis
3. Results
3.1. Establishing a Single Cell Proteomic Panel for CyTOF Analysis of Skeletal Muscle
3.2. Single Cell Proteomic Analysis Reveals Rapid Modifications During Dissociation.
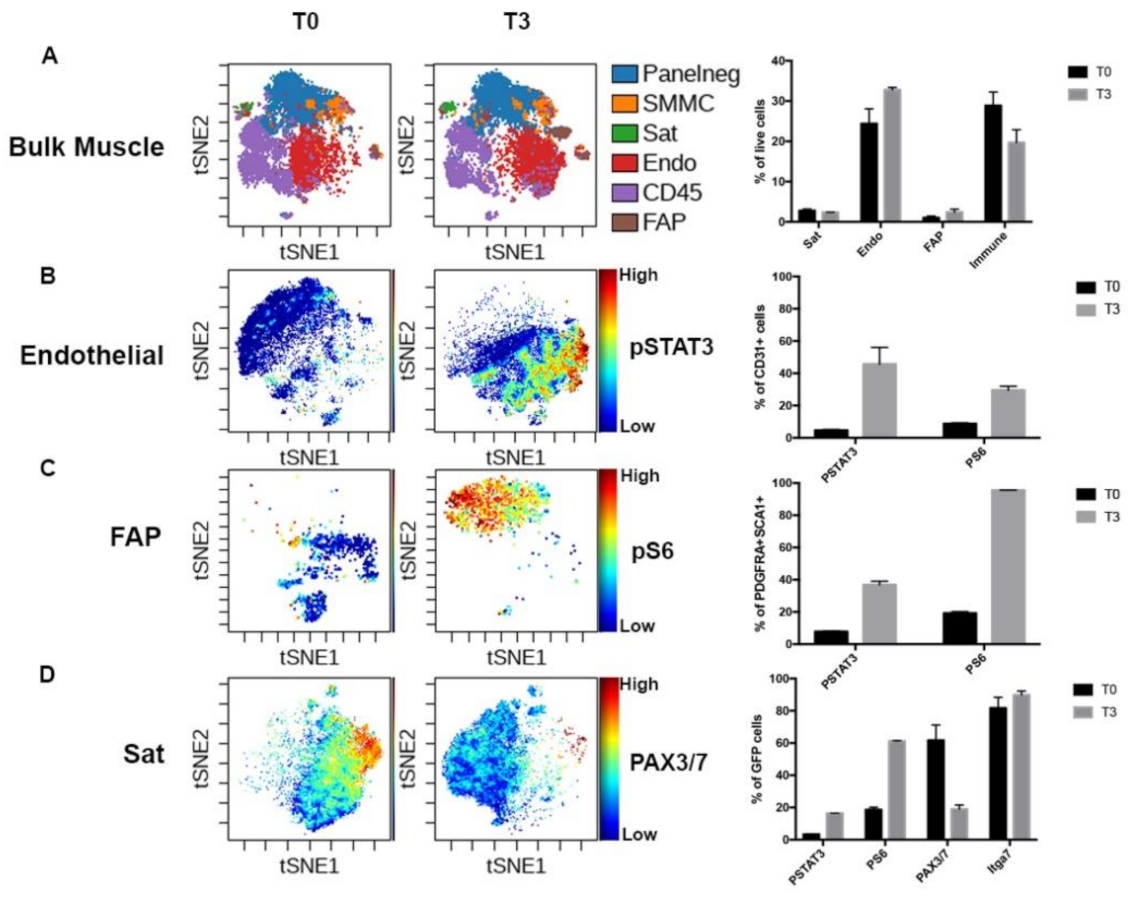
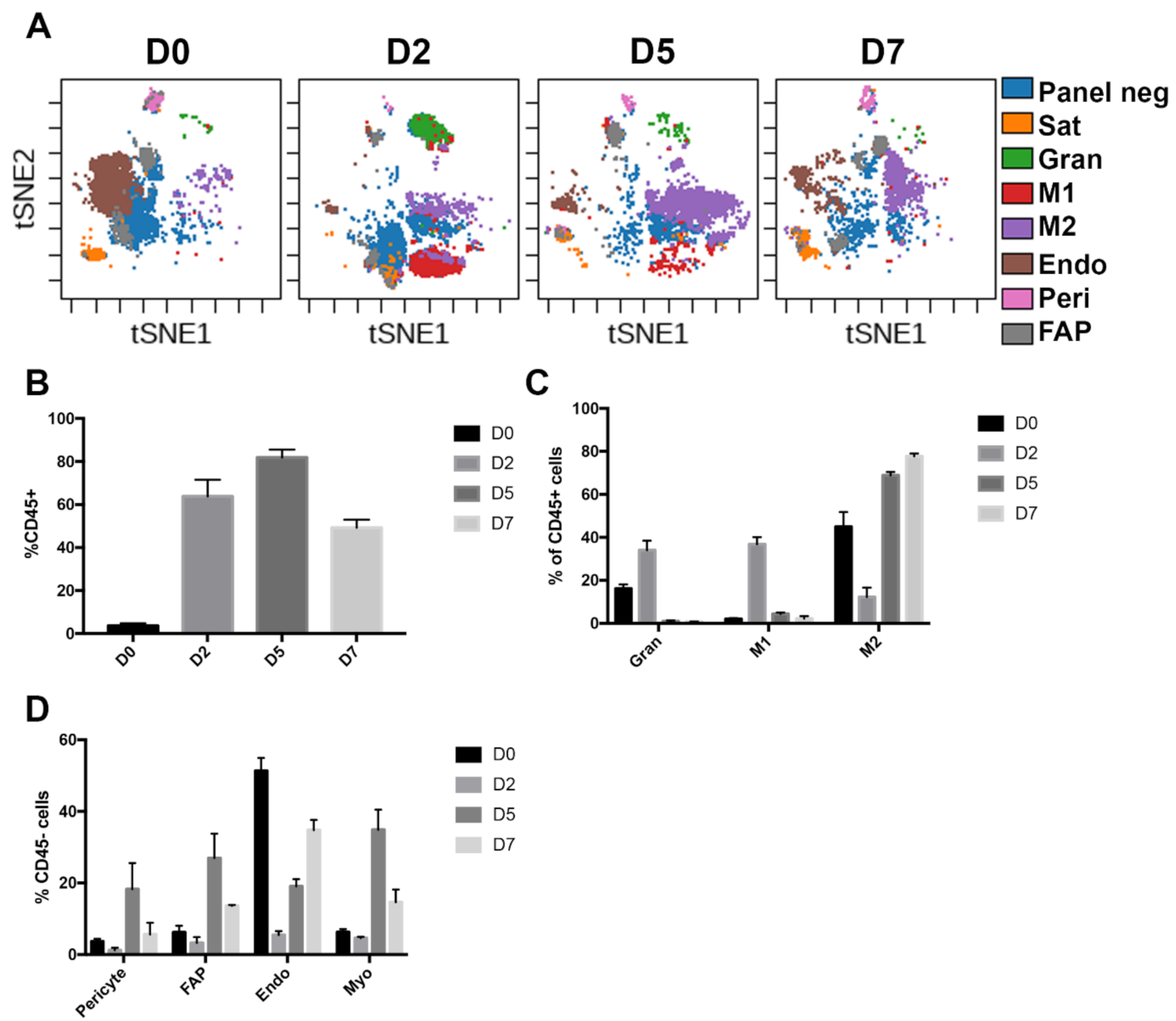
3.3. Muscle Niche Lineages Show Drastic Changes During Skeletal Muscle Regeneration
3.4. Dynamic Modification of the Myogenic Population Proteome During Regeneration
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Baghdadi, M.B.; Tajbakhsh, S. Regulation and phylogeny of skeletal muscle regeneration. Dev. Biol. 2018, 433, 200–209. [Google Scholar] [CrossRef] [PubMed]
- Hardy, D.; Besnard, A.; Latil, M.; Jouvion, G.; Briand, D.; Thépenier, C.; Pascal, Q.; Guguin, A.; Gayraud-Morel, B.; Cavaillon, J.-M.; et al. Comparative Study of Injury Models for Studying Muscle Regeneration in Mice. PLoS ONE 2016, 11, e0147198. [Google Scholar] [CrossRef] [PubMed]
- Collo, G.; Starr, L.; Quaranta, V. A new isoform of the laminin receptor integrin alpha 7 beta 1 is developmentally regulated in skeletal muscle. J. Biol. Chem. 1993, 268, 19019–19024. [Google Scholar] [CrossRef]
- Moore, R.; Walsh, F.S. The cell adhesion molecule M-cadherin is specifically expressed in developing and regenerating, but not denervated skeletal muscle. Development 1993, 117, 1409–1420. [Google Scholar]
- Zammit, P.S.; Golding, J.P.; Nagata, Y.; Hudon, V.; Partridge, T.A.; Beauchamp, J.R. Muscle satellite cells adopt divergent fates: A mechanism for self-renewal? J. Cell Biol. 2004, 166, 347–357. [Google Scholar] [CrossRef]
- Günther, S.; Kim, J.; Kostin, S.; Lepper, C.; Fan, C.-M.; Braun, T. Myf5-Positive Satellite Cells Contribute to Pax7-Dependent Long-Term Maintenance of Adult Muscle Stem Cells. Cell Stem Cell 2013, 13, 590–601. [Google Scholar] [CrossRef] [Green Version]
- Verma, M.; Asakura, Y.; Murakonda, B.S.R.; Pengo, T.; Latroche, C.; Chazaud, B.; McLoon, L.K.; Asakura, A. Muscle Satellite Cell Cross-Talk with a Vascular Niche Maintains Quiescence via VEGF and Notch Signaling. Cell Stem Cell 2018, 23, 530–543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, M.M.; Lawson, J.A.; Mathew, S.J.; Hutcheson, D.A.; Kardon, G. Satellite cells, connective tissue fibroblasts and their interactions are crucial for muscle regeneration. Development 2011, 138, 3625–3637. [Google Scholar] [CrossRef] [Green Version]
- Wosczyna, M.N.; Konishi, C.T.; Carbajal, E.E.P.; Wang, T.T.; Walsh, R.A.; Gan, Q.; Wagner, M.W.; Rando, T.A. Mesenchymal Stromal Cells Are Required for Regeneration and Homeostatic Maintenance of Skeletal Muscle. Cell Rep. 2019, 27, 2029–2035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kostallari, E.; Baba-Amer, Y.; Alonso-Martin, S.; Ngoh, P.; Relaix, F.; Lafuste, P.; Gherardi, R.K. Pericytes in the myovascular niche promote post-natal myofiber growth and satellite cell quiescence. Development 2015, 142, 1242–1253. [Google Scholar] [CrossRef] [Green Version]
- Arnold, L.; Henry, A.; Poron, F.; Baba-Amer, Y.; Van Rooijen, N.; Plonquet, A.; Gherardi, R.K.; Chazaud, B. Inflammatory monocytes recruited after skeletal muscle injury switch into antiinflammatory macrophages to support myogenesis. J. Exp. Med. 2007, 204, 1057–1069. [Google Scholar] [CrossRef] [Green Version]
- Lemos, D.R.; Babaeijandaghi, F.; Low, M.; Chang, C.-K.; Lee, S.T.; Fiore, D.; Zhang, R.-H.; Natarajan, A.; Nedospasov, S.A.; Rossi, F.M.V. Nilotinib reduces muscle fibrosis in chronic muscle injury by promoting TNF-mediated apoptosis of fibro/adipogenic progenitors. Nat. Med. 2015, 21, 786–794. [Google Scholar] [CrossRef]
- Spada, F.; Fuoco, C.; Pirro, S.; Paoluzi, S.; Castagnoli, L.; Gargioli, C.; Cesareni, G. Characterization by mass cytometry of different methods for the preparation of muscle mononuclear cells. New Biotechnol. 2016, 33, 514–523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porpiglia, E.; Samusik, N.; Van Ho, A.T.; Cosgrove, B.D.; Mai, T.; Davis, K.L.; Jager, A.; Nolan, G.P.; Bendall, S.C.; Fantl, W.J.; et al. High-resolution myogenic lineage mapping by single-cell mass cytometry. Nat. Cell Biol. 2017, 19, 558–567. [Google Scholar] [CrossRef] [Green Version]
- Dell’Orso, S.; Juan, A.H.; Ko, K.; Naz, F.; Perovanovic, J.; Gutierrez-Cruz, G.; Feng, X.; Sartorelli, V. Single cell analysis of adult mouse skeletal muscle stem cells in homeostatic and regenerative conditions. Development 2019, 146, dev174177. [Google Scholar] [CrossRef] [Green Version]
- Giordani, L.; He, G.J.; Negroni, E.; Sakai, H.; Law, J.Y.C.; Siu, M.M.; Wan, R.; Corneau, A.; Tajbakhsh, S.; Cheung, T.H.; et al. High-Dimensional Single-Cell Cartography Reveals Novel Skeletal Muscle Resident Cell Populations. Mol. Cell 2018, 74, 609–621. [Google Scholar] [CrossRef] [PubMed]
- de Micheli, A.J.; Fraczek, P.; Soueid-Baumgarten, S.; Ravichandran, H.; de Vlaminck, I.; Elemento, O. Single-Cell Analysis of the Muscle Stem Cell Hierarchy Identifies Heterotypic Communication Signals Involved in Skeletal Muscle Regeneration. Cell Rep. 2020, 30, 3583–3595. [Google Scholar] [CrossRef] [Green Version]
- Petrilli, L.L.; Spada, F.; Palma, A.; Reggio, A.; Rosina, M.; Gargioli, C.; Castagnoli, L.; Fuoco, C.; Cesareni, G. High-Dimensional Single-Cell Quantitative Profiling of Skeletal Muscle Cell Population Dynamics during Regeneration. Cells 2020, 9, 1723. [Google Scholar] [CrossRef]
- Machado, L.; de Lima, J.E.; Fabre, O.; Proux, C.; Legendre, R.; Szegedi, A.; Varet, H.; Ingerslev, L.R.; Barrès, R.; Relaix, F.; et al. In Situ Fixation Redefines Quiescence and Early Activation of Skeletal Muscle Stem Cells. Cell Rep. 2017, 21, 1982–1993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machado, L.; Geara, P.; Camps, J.; Dos Santos, M.; Teixeira-Clerc, F.; Van Herck, J.; Varet, H.; Legendre, R.; Pawlotsky, J.-M.; Sampaolesi, M.; et al. Tissue damage induces a conserved stress response that initiates quiescent muscle stem cell activation. Cell Stem Cell 2021, 1934. [Google Scholar] [CrossRef]
- van Velthoven, C.T.J.; de Morree, A.; Egner, I.M.; Brett, J.O.; Rando, T.A. Transcriptional Profiling of Quiescent Muscle Stem Cells In Vivo. Cell Rep. 2017, 21, 1994–2004. [Google Scholar] [CrossRef] [Green Version]
- Sambasivan, R.; Gayraud-Morel, B.; Dumas, G.; Cimper, C.; Paisant, S.; Kelly, R.G.; Tajbakhsh, S. Distinct regulatory cascades govern extraocular and pharyngeal arch muscle progenitor cell fates. Dev. Cell 2009, 16, 810–821. [Google Scholar] [CrossRef] [Green Version]
- Gattazzo, F.; Laurent, B.; Relaix, F.; Rouard, H.; Didier, N. Distinct Phases of Postnatal Skeletal Muscle Growth Govern the Progressive Establishment of Muscle Stem Cell Quiescence. Stem Cell Rep. 2020, 15, 597–611. [Google Scholar] [CrossRef]
- Dutzmann, J.; Daniel, J.-M.; Bauersachs, J.; Hilfiker-Kleiner, D.; Sedding, D.G. Emerging translational approaches to target STAT3 signalling and its impact on vascular disease. Cardiovasc. Res. 2015, 106, 365–374. [Google Scholar] [CrossRef] [Green Version]
- Trenerry, M.K.; Carey, K.A.; Ward, A.C.; Cameron-Smith, D. STAT3 signaling is activated in human skeletal muscle following acute resistance exercise. J. Appl. Physiol. 2007, 102, 1483–1489. [Google Scholar] [CrossRef] [Green Version]
- Lieskovska, J.; Guo, D.; Derman, E. Growth impairment in IL-6-overexpressing transgenic mice is associated with induction of SOCS3 mRNA. Growth Horm. IGF Res. 2003, 13, 26–35. [Google Scholar] [CrossRef]
- Dogra, C.; Srivastava, D.S.; Kumar, A. Protein–DNA array-based identification of transcription factor activities differentially regulated in skeletal muscle of normal and dystrophin-deficient mdx mice. Mol. Cell. Biochem. 2008, 312, 17–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srikuea, R.; Esser, K.A.; Pholpramool, C. Leukaemia inhibitory factor is expressed in rat gastrocnemius muscle after contusion and increases proliferation of rat L6 myoblasts via c-Myc signalling. Clin. Exp. Pharmacol. Physiol. 2011, 38, 501–509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodgers, J.T.; King, K.Y.; Brett, J.O.; Cromie, M.J.; Charville, G.W.; Maguire, K.K.; Brunson, C.; Mastey, N.; Liu, L.; Tsai, C.; et al. mTORC1 controls the adaptive transition of quiescent stem cells from G0 to G(Alert). Nature 2014, 510, 393–396. [Google Scholar] [CrossRef]
- der Vartanian, A.; Quétin, M.; Michineau, S.; Auradé, F.; Hayashi, S.; Dubois, C.; Rocancourt, D.; Drayton-Libotte, B.; Sze-gedi, A.; Buckingham, M.; et al. PAX3 Confers Functional Heterogeneity in Skeletal Muscle Stem Cell Responses to Environmental Stress. Cell Stem Cell 2019, 24, 958–973. [Google Scholar] [CrossRef]
- Bjornson, C.R.; Cheung, T.H.T.; Liu, L.; Tripathi, P.V.; Steeper, K.M.; Rando, T.A. Notch Signaling Is Necessary to Maintain Quiescence in Adult Muscle Stem Cells. Stem Cells 2011, 30, 232–242. [Google Scholar] [CrossRef] [Green Version]
- Mourikis, P.; Sambasivan, R.; Castel, D.; Rocheteau, P.; Bizzarro, V.; Tajbakhsh, S. A Critical Requirement for Notch Signaling in Maintenance of the Quiescent Skeletal Muscle Stem Cell State. Stem Cells 2012, 30, 243–252. [Google Scholar] [CrossRef]
- Mademtzoglou, D.; Asakura, Y.; Borok, M.J.; Alonso-Martin, S.; Mourikis, P.; Kodaka, Y.; Mohan, A.; Asakura, A.; Relaix, F. Author response: Cellular localization of the cell cycle inhibitor Cdkn1c controls growth arrest of adult skeletal muscle stem cells. eLife 2018, 7, e33337. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Wong, C.; Liu, D.; Finegold, M.; Harper, J.W.; Elledge, S.J. p21CIP1 and p57KIP2 control muscle differentiation at the myogenin step. Genes Dev. 1999, 13, 213–224. [Google Scholar] [CrossRef] [PubMed]
- Sambasivan, R.; Yao, R.; Kissenpfennig, A.; van Wittenberghe, L.; Paldi, A.; Gayraud-Morel, B.; Guenou, H.; Malissen, B.; Tajbakhsh, S.; Galy, A. Pax7-expressing satellite cells are indispensable for adult skeletal muscle regeneration. Development 2011, 138, 3647–3656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bustos, F.; De La Vega, E.; Cabezas, F.; Thompson, J.; Cornelison, D.; Olwin, B.B.; Yates, J.R.; Olguín, H.C. NEDD4 Regulates PAX7 Levels Promoting Activation of the Differentiation Program in Skeletal Muscle Precursors. STEM CELLS 2015, 33, 3138–3151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scott, R.W.; Arostegui, M.; Schweitzer, R.; Rossi, F.M.; Underhill, T.M. Hic1 Defines Quiescent Mesenchymal Progenitor Subpopulations with Distinct Functions and Fates in Skeletal Muscle Regeneration. Cell Stem Cell 2019, 25, 797–813. [Google Scholar] [CrossRef] [Green Version]
- Lepper, C.; Partridge, T.A.; Fan, C.-M. An absolute requirement for Pax7-positive satellite cells in acute injury-induced skeletal muscle regeneration. Development 2011, 138, 3639–3646. [Google Scholar] [CrossRef] [Green Version]
- McCarthy, J.J.; Mula, J.; Miyazaki, M.; Erfani, R.; Garrison, K.; Farooqui, A.B.; Srikuea, R.; Lawson, B.A.; Grimes, B.; Keller, C.; et al. Effective fiber hypertrophy in satellite cell-depleted skeletal muscle. Development 2011, 138, 3657–3666. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antigen | Localization | Metal Isotope | Company | Experiment |
|---|---|---|---|---|
| CD45 | Surface | 89Y | Fluidigm | Native and regenerating |
| LY6G | Surface | 141Pr | Fluidigm | Native and regenerating |
| CASP3 | Intracellular | 142Pr | Fluidigm | Native and regenerating |
| P57 | Intracellular | 143Nd | Santa Cruz | Native and regenerating |
| PDGFRB | Surface | 144Nd | Cell Signaling | Native and regenerating |
| F4/80 | Surface | 146Nd | Fluidigm | Native and regenerating |
| SMA | Intracellular | 147Sm | Sigma | Native and regenerating |
| CD34 | Surface | 148Nd | BD Biosciences | Regenerating |
| PDGFRA | Surface | 148Nd | Fluidigm | Native |
| p4EBP1 | Intracellular | 149Sm | Fluidigm | Native and regenerating |
| LY-6C | Surface | 150Nd | Fluidigm | Native and regenerating |
| NESTIN | Intracellular | 151Eu | Fluidigm | Native and regenerating |
| pAKT | Intracellular | 152Sm | Fluidigm | Regenerating |
| CD3E | Surface | 152Sm | Fluidigm | Native |
| CYCLINB1 | Intracellular | 153Eu | Fluidigm | Native and regenerating |
| SYNDECAN4 | Surface | 154Sm | Santa Cruz | Regenerating |
| ITGA7 | Surface | 154Sm | R&D Systems | Native |
| TCF4 | Surface | 155Gd | Fisher Sci | Native and regenerating |
| pSMAD1/5 | Intracellular | 156Gd | Cell Signaling | Native and regenerating |
| pSTAT3 | Intracellular | 158Gd | Fluidigm | Native and regenerating |
| P21 | Intracellular | 159Tb | Fluidigm | Regenerating |
| JAG1 | Surface | 159Tb | Santa Cruz | Native |
| PDGFRA | Surface | 160Gd | Fluidigm | Regenerating |
| PAX3/7 | Intracellular | 161Dy | Santa Cruz | Native and regenerating |
| KI67 | Intracellular | 162Dy | Fluidigm | Native and regenerating |
| NG2 | Surface | 163Dy | Cell Signaling | Native and regenerating |
| LY-6A_E | Surface | 164Dy | Fluidigm | Native and regenerating |
| CD31 | Surface | 165Ho | Fluidigm | Native and regenerating |
| pRB | Intracellular | 166Er | Fluidigm | Native and regenerating |
| P27 | Intracellular | 167Er | Santa Cruz | Native and regenerating |
| MCADHERIN | Surface | 168Er | Fisher Sci | Regenerating |
| CD8A | Surface | 168Er | Fluidigm | Native |
| GFP | Intracellular | 169Tm | Fluidigm | Native and regenerating |
| MYOGENIN | Intracellular | 170Er | Santa Cruz | Native and regenerating |
| pERK1/2 | Intracellular | 171Yb | Fluidigm | Native and regenerating |
| CD11B | Surface | 172Yb | Fluidigm | Regenerating |
| pS6 | Intracellular | 172Yb | Fluidigm | Native |
| VCAM | Surface | 173Yb | BD Biosciences | Native and regenerating |
| MYF5 | Intracellular | 174Yb | Santa Cruz | Regenerating |
| NRF2 | Intracellular | 174Yb | Cell Signaling | Native |
| pHH3 | Intracellular | 175Lu | Fluidigm | Native and regenerating |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Borok, M.; Didier, N.; Gattazzo, F.; Ozturk, T.; Corneau, A.; Rouard, H.; Relaix, F. Progressive and Coordinated Mobilization of the Skeletal Muscle Niche throughout Tissue Repair Revealed by Single-Cell Proteomic Analysis. Cells 2021, 10, 744. https://doi.org/10.3390/cells10040744
Borok M, Didier N, Gattazzo F, Ozturk T, Corneau A, Rouard H, Relaix F. Progressive and Coordinated Mobilization of the Skeletal Muscle Niche throughout Tissue Repair Revealed by Single-Cell Proteomic Analysis. Cells. 2021; 10(4):744. https://doi.org/10.3390/cells10040744
Chicago/Turabian StyleBorok, Matthew, Nathalie Didier, Francesca Gattazzo, Teoman Ozturk, Aurelien Corneau, Helene Rouard, and Frederic Relaix. 2021. "Progressive and Coordinated Mobilization of the Skeletal Muscle Niche throughout Tissue Repair Revealed by Single-Cell Proteomic Analysis" Cells 10, no. 4: 744. https://doi.org/10.3390/cells10040744
APA StyleBorok, M., Didier, N., Gattazzo, F., Ozturk, T., Corneau, A., Rouard, H., & Relaix, F. (2021). Progressive and Coordinated Mobilization of the Skeletal Muscle Niche throughout Tissue Repair Revealed by Single-Cell Proteomic Analysis. Cells, 10(4), 744. https://doi.org/10.3390/cells10040744