
Dynamics of Myosin II Filaments during Wound Repair in Dividing Cells



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Plasmids and Mutants
2.3. Chamber Preparation
2.4. Wounding and Microscopy
2.5. Measurement of Influx of PI and Ca2+
2.6. Ca–EGTA Buffer
2.7. Inhibitors
2.8. Statistical Analysis
3. Results
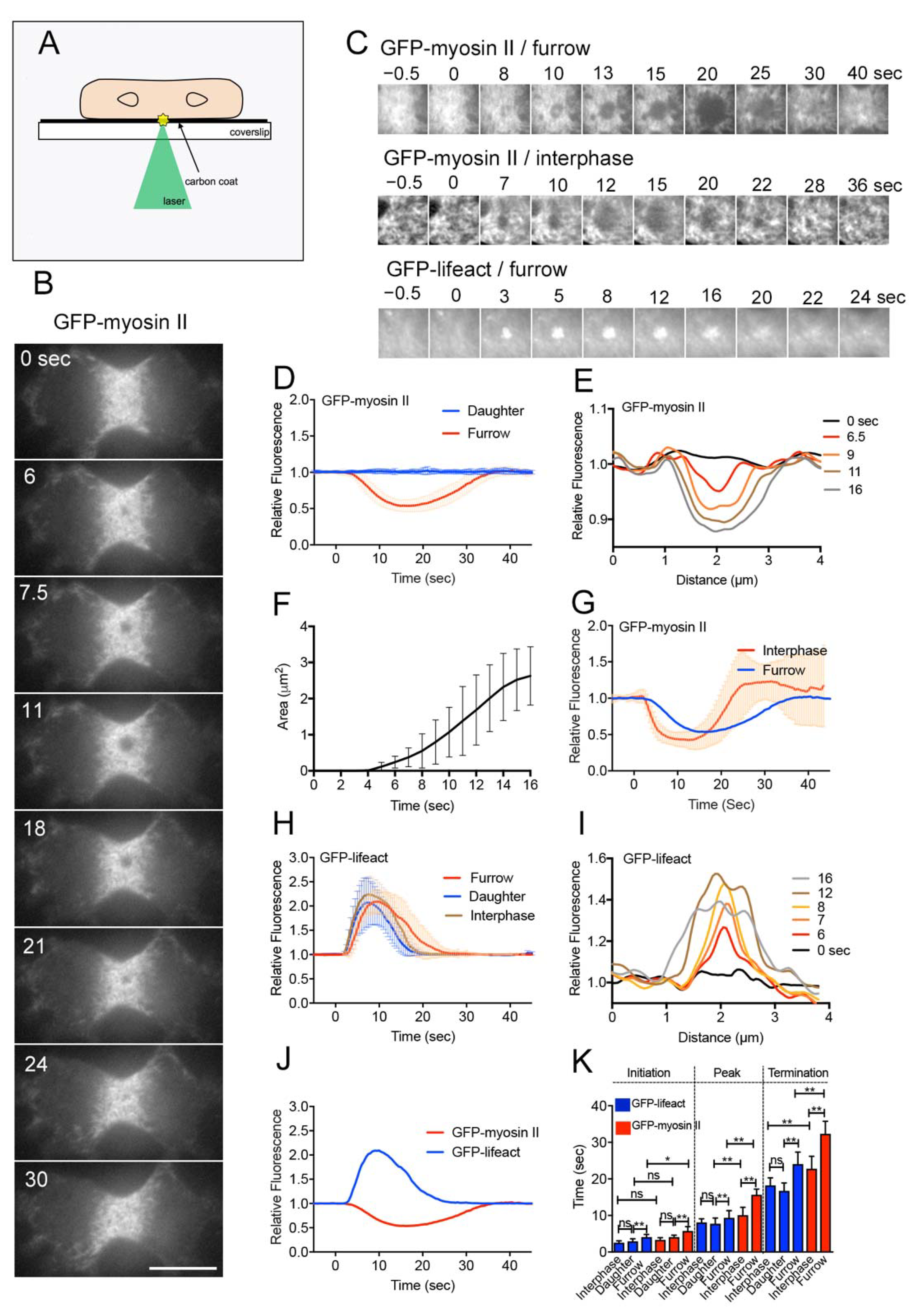
3.1. Myosin II Filaments Transiently Disappear from the Wound Site
3.2. Actin Filaments Transiently Accumulate at the Wound Site
3.3. Both Myosin II and Actin Dynamics Are Regulated by Ca2+ and Calmodulin
3.4. Role of Myosin II in Wound Repair
3.5. Mechanism for Myosin II Dynamics
3.6. Other Proteins Localizing at the Furrow also Disappeared from the Wound Site
4. Discussion
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bansal, D.; Miyake, K.; Vogel, S.S.; Groh, S.; Chen, C.-C.; Williamson, R.A.; McNeil, P.L.; Campbell, K.P. Defective membrane repair in dysferlin-deficient muscular dystrophy. Nat. Cell Biol. 2003, 423, 168–172. [Google Scholar] [CrossRef]
- Chakrabarti, S.; Kobayashi, K.S.; Flavell, R.A.; Marks, C.B.; Miyake, K.; Liston, D.R.; Fowler, K.T.; Gorelick, F.S.; Andrews, N.W. Impaired membrane resealing and autoimmune myositis in synaptotagmin VII–deficient mice. J. Cell Biol. 2003, 162, 543–549. [Google Scholar] [CrossRef]
- Howard, A.C.; McNeil, A.K.; Xiong, F.; Xiong, W.C.; McNeil, P.L. A novel cellular defect in diabetes: Membrane repair failure. Diabetes 2011, 60, 3034–3043. [Google Scholar] [CrossRef] [Green Version]
- Labazi, M.; McNeil, A.K.; Kurtz, T.; Lee, T.C.; Pegg, R.B.; Angeli, J.P.F.; Conrad, M.; McNeil, P.L. The antioxidant requirement for plasma membrane repair in skeletal muscle. Free. Radic. Biol. Med. 2015, 84, 246–253. [Google Scholar] [CrossRef] [Green Version]
- Meng, X.; Yang, Q.; Yu, X.; Zhou, J.; Ren, X.; Zhou, Y.; Xu, S. Actin polymerization and ESCRT trigger recruitment of the fusogens Syntaxin-2 and EFF-1 to promote membrane repair in C. elegans. Dev. Cell 2020, 54, 624–638.e5. [Google Scholar] [CrossRef] [PubMed]
- Waddell, L.B.; Lemckert, F.A.; Zheng, X.F.; Tran, J.; Evesson, F.J.; Hawkes, J.M.; Lek, A.; Street, N.E.; Lin, P.; Clarke, N.F.; et al. Dysferlin, annexin A1, and mitsugumin 53 are upregulated in muscular dystrophy and localize to longitudinal tubules of the T-system with stretch. J. Neuropathol. Exp. Neurol. 2011, 70, 302–313. [Google Scholar] [CrossRef] [Green Version]
- Abreu-Blanco, M.T.; Verboon, J.; Parkhurst, S.M. Cell wound repair in drosophila occurs through three distinct phases of membrane and cytoskeletal remodeling. J. Cell Biol. 2011, 193, 455–464. [Google Scholar] [CrossRef] [Green Version]
- Bi, G.Q.; Alderton, J.M.; Steinhardt, R.A. Calcium-regulated exocytosis is required for cell membrane resealing. J. Cell Biol. 1995, 131, 1747–1758. [Google Scholar] [CrossRef] [Green Version]
- Gingell, D. Contractile responses at the surface of an amphibian egg. Development 1970, 23, 583–609. [Google Scholar] [CrossRef]
- Kono, K.; Saeki, Y.; Yoshida, S.; Tanaka, K.; Pellman, D. Proteasomal degradation resolves competition between cell polarization and cellular wound healing. Cell 2012, 150, 151–164. [Google Scholar] [CrossRef] [Green Version]
- Pervin, M.S.; Itoh, G.; Talukder, S.U.; Fujimoto, K.; Morimoto, Y.V.; Tanaka, M.; Ueda, M.; Yumura, S. A study of wound repair in Dictyostelium cells by using novel laserporation. Sci. Rep. 2018, 8, 7969. [Google Scholar] [CrossRef]
- Szubinska, B. “New membrane” formation in amoeba proteus upon injury of individual cells. J. Cell Biol. 1971, 49, 747–772. [Google Scholar] [CrossRef] [PubMed]
- Togo, T.; Alderton, J.; Bi, G.; Steinhardt, R. The mechanism of facilitated cell membrane resealing. J. Cell Sci. 1999, 112, 719–731. [Google Scholar] [CrossRef]
- Steinhardt, R.; Bi, G.; Alderton, J. Cell membrane resealing by a vesicular mechanism similar to neurotransmitter release. Science 1994, 263, 390–393. [Google Scholar] [CrossRef]
- Bement, W.M.; Mandato, C.A.; Kirsch, M.N. Wound-induced assembly and closure of an actomyosin purse string in Xenopus oocytes. Curr. Biol. 1999, 9, 579–587. [Google Scholar] [CrossRef] [Green Version]
- Togo, T.; Steinhardt, R.A. Nonmuscle myosin IIA and IIB Have distinct functions in the exocytosis-dependent process of cell membrane repair. Mol. Biol. Cell 2004, 15, 688–695. [Google Scholar] [CrossRef] [Green Version]
- Yumura, S.; Hashima, S.; Muranaka, S. Myosin II does not contribute to wound repair in Dictyostelium cells. Biol. Open 2014, 3, 966–973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talukder, S.U.; Pervin, M.S.; Tanvir, I.O.; Fujimoto, K.; Tanaka, M.; Itoh, G.; Yumura, S. Ca2+–Calmodulin dependent wound repair in Dictyostelium cell membrane. Cells 2020, 9, 1058. [Google Scholar] [CrossRef] [Green Version]
- Bouter, A.; Gounou, C.; Bérat, R.; Tan, S.; Gallois, B.; Granier, T.; D’Estaintot, B.L.; Pöschl, E.; Brachvogel, B.; Brisson, A.R. Annexin-A5 assembled into two-dimensional arrays promotes cell membrane repair. Nat. Commun. 2011, 2, 270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaiswal, J.K.; Lauritzen, S.P.; Scheffer, L.; Sakaguchi, M.; Bunkenborg, J.; Simon, S.M.; Kallunki, T.; Jaattela, M.; Nylandsted, J. S100A11 is required for efficient plasma membrane repair and survival of invasive cancer cells. Nat. Commun. 2014, 5, 1–13. [Google Scholar] [CrossRef] [Green Version]
- López-Jiménez, A.T.; Cardenal-Muñoz, E.; Leuba, F.; Gerstenmaier, L.; Barisch, C.; Hagedorn, M.; King, J.S.; Soldati, T. The ESCRT and autophagy machineries cooperate to repair ESX-1-dependent damage at the Mycobacterium-containing vacuole but have opposite impact on containing the infection. PLoS Pathog. 2018, 14, e1007501. [Google Scholar] [CrossRef] [Green Version]
- McNeil, A.K.; Rescher, U.; Gerke, V.; McNeil, P.L. Requirement for annexin A1 in plasma membrane repair. J. Biol. Chem. 2006, 281, 35202–35207. [Google Scholar] [CrossRef] [Green Version]
- Rühl, S.; Shkarina, K.; Demarco, B.; Heilig, R.; Santos, J.C.; Broz, P. ESCRT-dependent membrane repair negatively regulates pyroptosis downstream of GSDMD activation. Science 2018, 362, 956–960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yumura, S. Myosin II dynamics and cortical flow during contractile ring formation in Dictyostelium cells. J. Cell Biol. 2001, 154, 137–146. [Google Scholar] [CrossRef] [Green Version]
- Yumura, S.; Uyeda, T.Q. Transport of myosin II to the equatorial region without its own motor activity in mitotic Dictyostelium cells. Mol. Biol. Cell 1997, 8, 2089–2099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yumura, S.; Yoshida, M.; Betapudi, V.; Licate, L.S.; Iwadate, Y.; Nagasaki, A.; Uyeda, T.Q.; Egelhoff, T.T. Multiple myosin ii heavy chain kinases: Roles in filament assembly control and proper cytokinesis in Dictyostelium. Mol. Biol. Cell 2005, 16, 4256–4266. [Google Scholar] [CrossRef] [Green Version]
- Chung, C.Y.; Firtel, R.A. Paka, a putative pak family member, is required for cytokinesis and the regulation of the cytoskeleton in Dictyostelium discoideum cells during chemotaxis. J. Cell Biol. 1999, 147, 559–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujimoto, K.; Tanaka, M.; Rana, A.K.M.M.; Jahan, G.S.; Itoh, G.; Tsujioka, M.; Uyeda, T.Q.P.; Miyagishima, S.-Y.; Yumura, S. Dynamin-like protein B of Dictyostelium contributes to cytokinesis cooperatively with other dynamins. Cells 2019, 8, 781. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, M.; Fujimoto, K.; Yumura, S. Regulation of the total cell surface area in dividing Dictyostelium cells. Front. Cell Dev. Biol. 2020, 8, 238. [Google Scholar] [CrossRef]
- Pramanik, M.K.; Iijima, M.; Iwadate, Y.; Yumura, S. PTEN is a mechanosensing signal transducer for myosin II localization in Dictyostelium cells. Genes Cells 2009, 14, 821–834. [Google Scholar] [CrossRef]
- Tanaka, Y.; Jahan, G.S.; Kondo, T.; Nakano, M.; Yumura, S. Cytokinesis D is mediated by cortical flow of dividing cells instead of chemotaxis. Cells 2019, 8, 473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yumura, S.; Matsuzaki, R.; Kitanishi-Yumura, T. Introduction of macromolecules into living Dictyostelium cells by electroporation. Cell Struct. Funct. 1995, 20, 185–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yumura, S. A novel low-power laser-mediated transfer of foreign molecules into cells. Sci. Rep. 2016, 6, 22055. [Google Scholar] [CrossRef] [PubMed]
- Manstein, D.J.; Titus, M.A.; De Lozanne, A.; Spudich, J.A. Gene replacement in Dictyostelium: Generation of myosin null mutants. EMBO J. 1989, 8, 923–932. [Google Scholar] [CrossRef] [PubMed]
- Niswonger, M.L.; O’Halloran, T.J. A novel role for clathrin in cytokinesis. Proc. Natl. Acad. Sci. USA 1997, 94, 8575–8578. [Google Scholar] [CrossRef] [Green Version]
- Weber, I.; Gerisch, G.; Heizer, C.; Murphy, J.; Badelt, K.; Stock, A.; Schwartz, J.; Faix, J. Cytokinesis mediated through the recruitment of cortexillins into the cleavage furrow. EMBO J. 1999, 18, 586–594. [Google Scholar] [CrossRef] [PubMed]
- Adachi, H.; Takahashi, Y.; Hasebe, T.; Shirouzu, M.; Yokoyama, S.; Sutoh, K. Dictyostelium IQGAP-related protein specifically involved in the completion of cytokinesis. J. Cell Biol. 1997, 137, 891–898. [Google Scholar] [CrossRef] [Green Version]
- Janetopoulos, C.; Borleis, J.; Vazquez, F.; Iijima, M.; Devreotes, P. Temporal and spatial regulation of phosphoinositide signaling mediates cytokinesis. Dev. Cell 2005, 8, 467–477. [Google Scholar] [CrossRef] [Green Version]
- Pervin, M.S.; Yumura, S. Manipulation of cell migration by laserporation-induced local wounding. Sci. Rep. 2019, 9, 4291. [Google Scholar] [CrossRef]
- Yumura, S.; Itoh, G.; Kikuta, Y.; Kikuchi, T.; Kitanishi-Yumura, T.; Tsujioka, M. Cell-scale dynamic recycling and cortical flow of the actin–myosin cytoskeleton for rapid cell migration. Biol. Open 2013, 2, 200–209. [Google Scholar] [CrossRef] [Green Version]
- Yumura, S.; Mori, H.; Fukui, Y. Localization of actin and myosin for the study of ameboid movement in Dictyostelium using improved immunofluorescence. J. Cell Biol. 1984, 99, 894–899. [Google Scholar] [CrossRef] [PubMed]
- Yumura, S.; Ueda, M.; Sako, Y.; Kitanishi-Yumura, T.; Yanagida, T. Multiple mechanisms for accumulation of myosin ii filaments at the equator during cytokinesis. Traffic 2008, 9, 2089–2099. [Google Scholar] [CrossRef] [PubMed]
- Egelhoff, T.T.; Lee, R.J.; Spudich, J.A. Dictyostelium myosin heavy chain phosphorylation sites regulate myosin filament assembly and localization in vivo. Cell 1993, 75, 363–371. [Google Scholar] [CrossRef]
- Yumura, S.; Uyeda, T.Q. Myosins and cell dynamics in cellular slime molds. Adv. Appl. Microbiol. 2003, 224, 173–225. [Google Scholar] [CrossRef]
- Yumura, S.; Kitanishi-Yumura, T. A mechanism for the intracellular localization of myosin II filaments in the Dictyostelium amoeba. J. Cell Sci. 1993, 105, 233–242. [Google Scholar] [CrossRef]
- Yumura, S.; Kitanishi-Yumura, T. Release of myosin II from the membrane-cytoskeleton of Dictyostelium discoideum mediated by heavy-chain phosphorylation at the foci within the cortical actin network. J. Cell Biol. 1992, 117, 1231–1239. [Google Scholar] [CrossRef] [PubMed]
- Faix, J.; Weber, I.; Mintert, U.; Köhler, J.; Lottspeich, F.; Marriott, G. Recruitment of cortexillin into the cleavage furrow is con-trolled by Rac1 and IQGAP-related proteins. EMBO J. 2001, 20, 3705–3715. [Google Scholar] [CrossRef]
- Iijima, M.; Huang, Y.E.; Luo, H.R.; Vazquez, F.; Devreotes, P.N. Novel mechanism of PTEN regulation by its phosphatidylinositol 4,5-bisphosphate binding motif is critical for chemotaxis. J. Biol. Chem. 2004, 279, 16606–16613. [Google Scholar] [CrossRef] [Green Version]
- Marinović, M.; Mijanović, L.; Šoštar, M.; Vizovišek, M.; Junemann, A.; Fonović, M.; Turk, B.; Weber, I.; Faix, J.; Filić, V. IQGAP-related protein IqgC suppresses ras signaling during large-scale endocytosis. Proc. Natl. Acad. Sci. USA 2019, 116, 1289–1298. [Google Scholar] [CrossRef] [Green Version]
- Pelikan-Conchaudron, A.; Le Clainche, C.; Didry, D.; Carlier, M.-F. The IQGAP1 protein is a calmodulin-regulated barbed end capper of actin filaments: Possible implications in its function in cell migration*. J. Biol. Chem. 2011, 286, 35119–35128. [Google Scholar] [CrossRef] [Green Version]
- Mondal, S.; Burgute, B.; Rieger, D.; Müller, R.; Rivero, F.; Faix, J.; Schleicher, M.; Noegel, A.A. Regulation of the actin cytoskeleton by an interaction of IQGAP related protein GAPA with filamin and cortexillin I. PLoS ONE 2010, 5, e15440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kothari, P.; Srivastava, V.; Aggarwal, V.; Tchernyshyov, I.; Van Eyk, J.E.; Ha, T.; Robinson, D.N. Contractility kits promote assembly of the mechanoresponsive cytoskeletal network. J. Cell Sci. 2019, 132, jcs.226704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Day, D.H.; Mathavarajah, S.; Myre, M.A.; Huber, R.J. Calmodulin-mediated events during the life cycle of the amoebozoan Dictyostelium discoideum. Biol. Rev. 2020, 95, 472–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Condeelis, J.; Vahey, M.; Carboni, J.M.; Demey, J.; Ogihara, S. Properties of the 120,000- and 95,000-dalton actin-binding proteins from Dictyostelium discoideum and their possible functions in assembling the cytoplasmic matrix. J. Cell Biol. 1984, 99, 119s–126s. [Google Scholar] [CrossRef] [Green Version]
- Chaigne, A.; Campillo, C.; Gov, N.S.; Voituriez, R.; Azoury, J.; Umaña-Diaz, C.; Almonacid, M.; Queguiner, I.; Nassoy, P.; Sykes, C.; et al. A soft cortex is essential for asymmetric spindle positioning in mouse oocytes. Nat. Cell Biol. 2013, 15, 958–966. [Google Scholar] [CrossRef]
- Nagasaki, A.; Itoh, G.; Yumura, S.; Uyeda, T.Q. Novel myosin heavy chain kinase involved in disassembly of myosin II filaments and efficient cleavage in mitotic Dictyostelium cells. Mol. Biol. Cell 2002, 13, 4333–4342. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tanvir, M.I.O.; Itoh, G.; Adachi, H.; Yumura, S. Dynamics of Myosin II Filaments during Wound Repair in Dividing Cells. Cells 2021, 10, 1229. https://doi.org/10.3390/cells10051229
Tanvir MIO, Itoh G, Adachi H, Yumura S. Dynamics of Myosin II Filaments during Wound Repair in Dividing Cells. Cells. 2021; 10(5):1229. https://doi.org/10.3390/cells10051229
Chicago/Turabian StyleTanvir, Md. Istiaq Obaidi, Go Itoh, Hiroyuki Adachi, and Shigehiko Yumura. 2021. "Dynamics of Myosin II Filaments during Wound Repair in Dividing Cells" Cells 10, no. 5: 1229. https://doi.org/10.3390/cells10051229
APA StyleTanvir, M. I. O., Itoh, G., Adachi, H., & Yumura, S. (2021). Dynamics of Myosin II Filaments during Wound Repair in Dividing Cells. Cells, 10(5), 1229. https://doi.org/10.3390/cells10051229