
Extracellular Vesicles and Their miRNA Content in Amniotic and Tracheal Fluids of Fetuses with Severe Congenital Diaphragmatic Hernia Undergoing Fetal Intervention

 , ,
, ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. EV Enrichment and RNA Extraction
2.3. EV Characterization
2.4. MicroRNA Profiling
2.5. MicroRNA Validation
2.6. Statistical Analysis
2.7. Prediction of Target Genes
3. Results
3.1. Clinical Data
3.2. Size Distribution and Count of Extracellular Vesicles in FETO Survivors and Non-Survivors
3.3. Characterization of Extracellular Vesicles by Flow Cytometry
3.4. miRNA Expression Screening in Amniotic and Tracheal Fluids
3.5. miRNA Validation in Amniotic and Tracheal Fluids
3.6. Prediction of Target Genes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McGivern, M.R.; Best, K.E.; Rankin, J.; Wellesley, D.; Greenlees, R.; Addor, M.C.; Arriola, L.; de Walle, H.; Barisic, I.; Beres, J.; et al. Epidemiology of congenital diaphragmatic hernia in Europe: A register-based study. Arch. Dis. Child. Fetal Neonatal Ed. 2015, 100, F137–F144. [Google Scholar] [CrossRef]
- Russo, F.M.; Cordier, A.G.; De Catte, L.; Saada, J.; Benachi, A.; Deprest, J. Proposal for standardized prenatal ultrasound assessment of the fetus with congenital diaphragmatic hernia by the European reference network on rare inherited and congenital anomalies (ERNICA). Prenat. Diagn. 2018, 38, 629–637. [Google Scholar] [CrossRef] [PubMed]
- Jani, J.C.; Nicolaides, K.H.; Gratacos, E.; Valencia, C.M.; Done, E.; Martinez, J.M.; Gucciardo, L.; Cruz, R.; Deprest, J.A. Severe diaphragmatic hernia treated by fetal endoscopic tracheal occlusion. Ultrasound Obstetr. Gynecol. 2009, 34, 304–310. [Google Scholar] [CrossRef] [PubMed]
- Horn-Oudshoorn, E.J.J.; Knol, R.; Te Pas, A.B.; Hooper, S.B.; Cochius-den Otter, S.C.M.; Wijnen, R.M.H.; Schaible, T.; Reiss, I.K.M.; DeKoninck, P.L.J. Perinatal stabilisation of infants born with congenital diaphragmatic hernia: A review of current concepts. Arch. Dis. Childh. Fetal Neonatal Ed. 2020, 105, 449–454. [Google Scholar] [CrossRef]
- Wahid, F.; Shehzad, A.; Khan, T.; Kim, Y.Y. MicroRNAs: Synthesis, mechanism, function, and recent clinical trials. Biochim. Biophys. Acta 2010, 1803, 1231–1243. [Google Scholar] [CrossRef]
- Yanez-Mo, M.; Siljander, P.R.; Andreu, Z.; Zavec, A.B.; Borras, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; et al. Biological properties of extracellular vesicles and their physiological functions. J. Extracell. Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef]
- Johar, D.; Siragam, V.; Mahood, T.H.; Keijzer, R. New insights into lung development and diseases: The role of microRNAs. Biochem. Cell Biol. 2015, 93, 139–148. [Google Scholar] [CrossRef]
- Lee, Y.; El Andaloussi, S.; Wood, M.J. Exosomes and microvesicles: Extracellular vesicles for genetic information transfer and gene therapy. Hum. Mol. Genet. 2012, 21, R125–R134. [Google Scholar] [CrossRef]
- Pitt, J.M.; Kroemer, G.; Zitvogel, L. Extracellular vesicles: Masters of intercellular communication and potential clinical interventions. J. Clin. Investig. 2016, 126, 1139–1143. [Google Scholar] [CrossRef]
- Tkach, M.; Thery, C. Communication by Extracellular Vesicles: Where We Are and Where We Need to Go. Cell 2016, 164, 1226–1232. [Google Scholar] [CrossRef]
- Deprest, J.; Gratacos, E.; Nicolaides, K.H. Fetoscopic tracheal occlusion (FETO) for severe congenital diaphragmatic hernia: Evolution of a technique and preliminary results. Ultrasound Obstet. Gynecol. 2004, 24, 121–126. [Google Scholar] [CrossRef]
- Thery, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef]
- Andersen, C.L.; Jensen, J.L.; Orntoft, T.F. Normalization of real-time quantitative reverse transcription-PCR data: A model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Dweep, H.; Gretz, N.; Sticht, C. miRWalk database for miRNA-target interactions. Methods Mol. Biol. 2014, 1182, 289–305. [Google Scholar] [CrossRef]
- Dweep, H.; Sticht, C.; Pandey, P.; Gretz, N. miRWalk—Database: Prediction of possible miRNA binding sites by “walking” the genes of three genomes. J. Biomed. Inform. 2011, 44, 839–847. [Google Scholar] [CrossRef] [PubMed]
- Russell, M.K.; Longoni, M.; Wells, J.; Maalouf, F.I.; Tracy, A.A.; Loscertales, M.; Ackerman, K.G.; Pober, B.R.; Lage, K.; Bult, C.J.; et al. Congenital diaphragmatic hernia candidate genes derived from embryonic transcriptomes. Proc. Natl. Acad. Sci. USA 2012, 109, 2978–2983. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, D.L.; Church, D.M.; Edgar, R.; Federhen, S.; Helmberg, W.; Madden, T.L.; Pontius, J.U.; Schuler, G.D.; Schriml, L.M.; Sequeira, E.; et al. Database resources of the National Center for Biotechnology Information: Update. Nucleic Acids Res. 2004, 32, D35–D40. [Google Scholar] [CrossRef] [PubMed]
- Letsiou, E.; Sammani, S.; Zhang, W.; Zhou, T.; Quijada, H.; Moreno-Vinasco, L.; Dudek, S.M.; Garcia, J.G. Pathologic mechanical stress and endotoxin exposure increases lung endothelial microparticle shedding. Am. J. Respir. Cell Mol. Biol. 2015, 52, 193–204. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Zhang, D.; Zhu, Z.; Dela Cruz, C.S.; Jin, Y. Epithelial cell-derived microvesicles activate macrophages and promote inflammation via microvesicle-containing microRNAs. Sci. Rep. 2016, 6, 35250. [Google Scholar] [CrossRef]
- Moon, H.G.; Cao, Y.; Yang, J.; Lee, J.H.; Choi, H.S.; Jin, Y. Lung epithelial cell-derived extracellular vesicles activate macrophage-mediated inflammatory responses via ROCK1 pathway. Cell Death Dis. 2015, 6, e2016. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Sun, M.; Ramchandran, R.; Raj, J.U. IGF-1 signaling in neonatal hypoxia-induced pulmonary hypertension: Role of epigenetic regulation. Vasc. Pharmacol. 2015, 73, 20–31. [Google Scholar] [CrossRef]
- Ambalavanan, N.; Bulger, A.; Murphy-Ullrich, J.; Oparil, S.; Chen, Y.F. Endothelin-A receptor blockade prevents and partially reverses neonatal hypoxic pulmonary vascular remodeling. Pediatr. Res. 2005, 57, 631–636. [Google Scholar] [CrossRef]
- Schiffrin, E.L. Endothelin: Potential role in hypertension and vascular hypertrophy. Hypertension 1995, 25, 1135–1143. [Google Scholar] [CrossRef]
- Wang, K.; Song, Y.; Chen, D.B.; Zheng, J. Protein phosphatase 3 differentially modulates vascular endothelial growth factor- and fibroblast growth factor 2-stimulated cell proliferation and signaling in ovine fetoplacental artery endothelial cells. Biol. Reprod. 2008, 79, 704–710. [Google Scholar] [CrossRef]
- Feng, L.; Liao, W.X.; Luo, Q.; Zhang, H.H.; Wang, W.; Zheng, J.; Chen, D.B. Caveolin-1 orchestrates fibroblast growth factor 2 signaling control of angiogenesis in placental artery endothelial cell caveolae. J. Cell. Physiol 2012, 227, 2480–2491. [Google Scholar] [CrossRef]
- Zheng, J.; Wen, Y.; Song, Y.; Wang, K.; Chen, D.B.; Magness, R.R. Activation of multiple signaling pathways is critical for fibroblast growth factor 2- and vascular endothelial growth factor-stimulated ovine fetoplacental endothelial cell proliferation. Biol. Reprod. 2008, 78, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Kang, Y.; Kojima, Y.; Lighthouse, J.K.; Hu, X.; Aldred, M.A.; McLean, D.L.; Park, H.; Comhair, S.A.; Greif, D.M.; et al. An endothelial apelin-FGF link mediated by miR-424 and miR-503 is disrupted in pulmonary arterial hypertension. Nat. Med. 2013, 19, 74–82. [Google Scholar] [CrossRef]
- Zeng, Y.; Zhang, X.; Kang, K.; Chen, J.; Wu, Z.; Huang, J.; Lu, W.; Chen, Y.; Zhang, J.; Wang, Z.; et al. MicroRNA-223 Attenuates Hypoxia-induced Vascular Remodeling by Targeting RhoB/MLC2 in Pulmonary Arterial Smooth Muscle Cells. Sci. Rep. 2016, 6, 24900. [Google Scholar] [CrossRef]
- Liu, A.; Liu, Y.; Li, B.; Yang, M.; Su, J. Role of miR-223-3p in pulmonary arterial hypertension via targeting ITGB3 in the ECM pathway. Cell Prolif. 2019, 52, e12550. [Google Scholar] [CrossRef] [PubMed]
- Pereira-Terra, P.; Deprest, J.A.; Kholdebarin, R.; Khoshgoo, N.; DeKoninck, P.; Munck, A.A.; Wang, J.; Zhu, F.; Rottier, R.J.; Iwasiow, B.M.; et al. Unique Tracheal Fluid MicroRNA Signature Predicts Response to FETO in Patients With Congenital Diaphragmatic Hernia. Ann. Surg. 2015, 262, 1130–1140. [Google Scholar] [CrossRef] [PubMed]




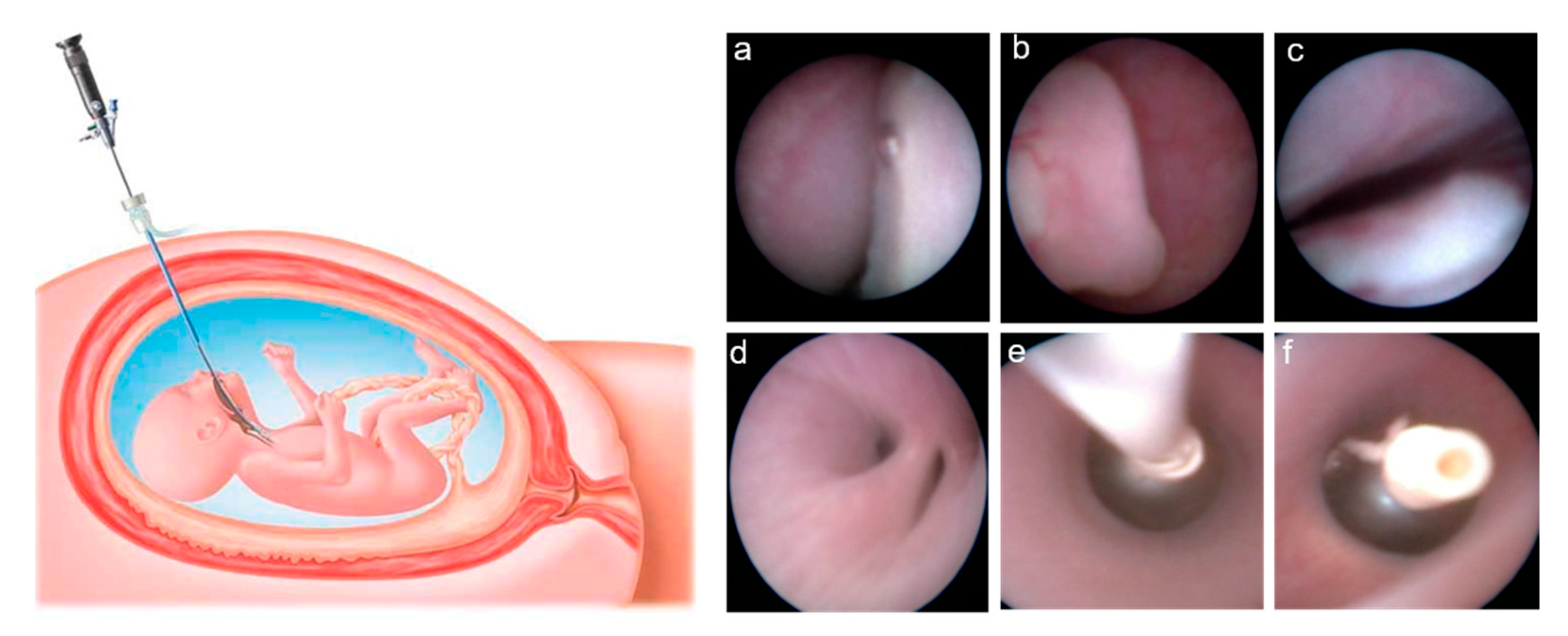{kind=link}
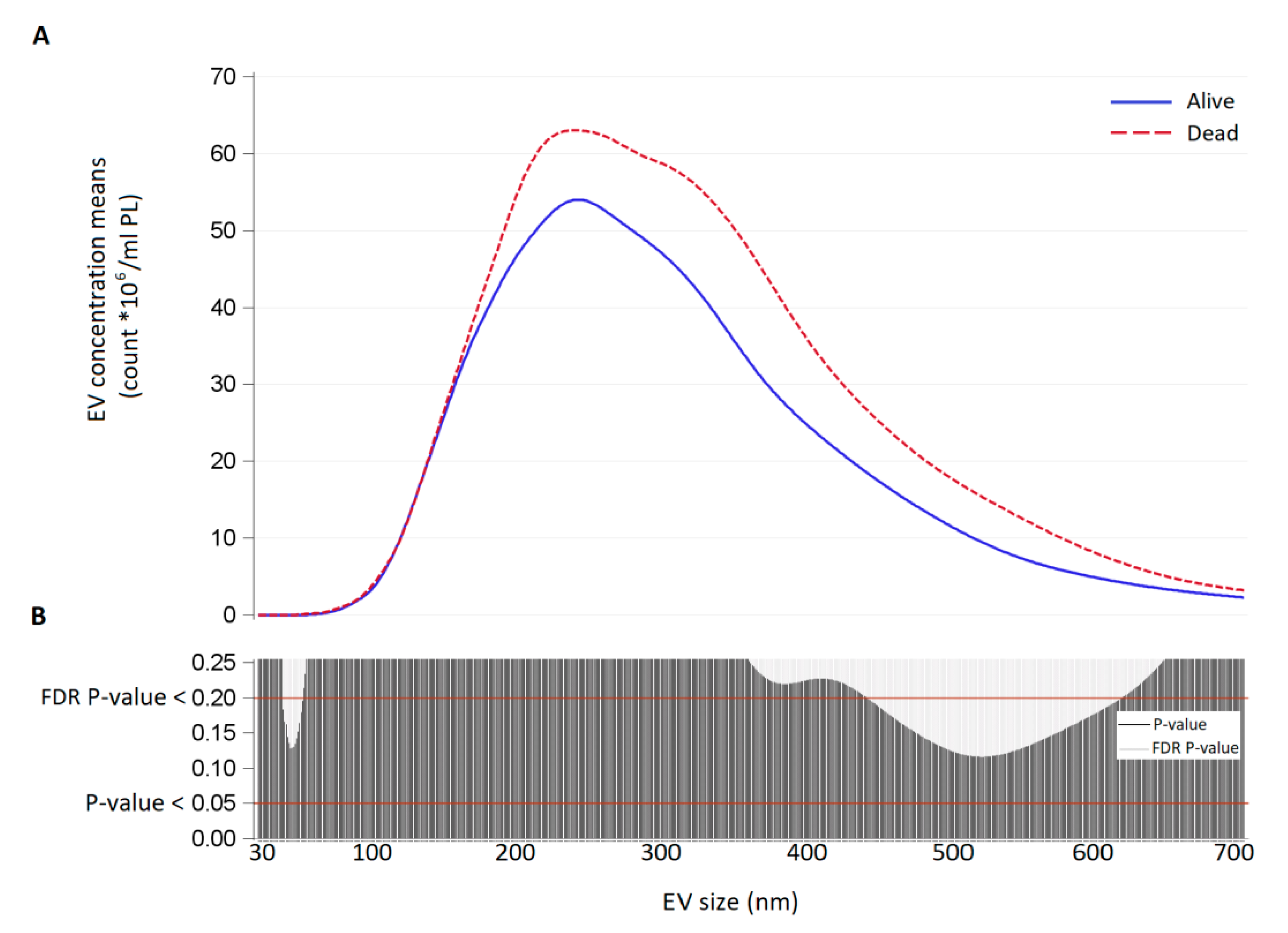{kind=link}
{kind=link}
| Survivors n = 7 | Non-Survivors n = 8 | p-Value | |
|---|---|---|---|
| Obstetric Factors | |||
| GA at FETO, weeks median (q1–q3) | 29.3 (28–29.9) | 28.0 (27.7–29.2) | 0.2699 |
| Duration of TO *, days median (q1–q3) | 42 (37–44) | 37 (35–42.5) | 0.4154 |
| GA at delivery, weeks median (q1–q3) | 38.3 (36.6–39) | 33.3 (32.5–36.5) | 0.0127 |
| Fetal Factors | |||
| Birth weight, g median (q1–q3) | 3015 (2500–3440) | 2160 (1740–2480) | 0.0205 |
| Female gender, n (%) | 4 (57.1) | 5 (62.5) | 0.8327 |
| Side of the defect | |||
| Left | 4 (57.1) | 6 (75.0) | 0.4126 * |
| Right | 3 (42.8) | 1 (12.5) | |
| Bilateral | 0 (0) | 1 (12.5) | |
| O/E LHR pre-FETO, % median (q1–q3) | 33 (22.6–40.4) | 25.3 (21.2–28) | 0.1476 |
| O/E LHR pre-removal, % median (q1–q3) | 67 (59.3–76) | 39.3 (37–50.8) | 0.0015 |
| Difference between pre-balloon removal and pre-FETO O/E LHR, % median (q1–q3) | 35.8 (26.4–44.4) | 16.2 (13.8–24.4) | 0.0177 |
| Amniotic Fluid | Tracheal Fluid | |||
|---|---|---|---|---|
| Ratio Non-Survivor/Survivor | p-Value * | Ratio Non-Survivor/Survivor | p-Value * | |
| A—Nanoparticle Tracking Analysis | ||||
| Total EV | 1.28 | <0.0001 | 3.12 | <0.0001 |
| Exosomes (<130 nm) | 1.06 | 0.0965 | 1.94 | <0.0001 |
| Microvesicles (>130 nm) | 1.29 | <0.0001 | 3.17 | <0.0001 |
| B—Flow Cytometry | ||||
| CD66+ (neutrophils) | 1.41 | <0.0001 | 0.97 | 0.6873 |
| CD14+ (monocytes/macrophages) | 1.42 | <0.0001 | 1.33 | 0.0005 |
| CD105+ (endothelium) | n/a | n/a | 1.5 | <0.0001 |
| EpCAM+ (epithelium) | 1.55 | <0.0001 | 1.32 | 0.0010 |
| HLAG+ (placenta) | 1.43 | <0.0001 | n/a | n/a |
| HERV-W+ (placenta) | 1.13 | 0.0008 | n/a | n/a |
| Amniotic Fluids | Tracheal Fluids | |||
|---|---|---|---|---|
| miRNA | Fold Change | p-Value * | Fold Change | p-Value * |
| mir-200a-5p | 0.450 | 0.2569 | 2.087 | 0.0005 |
| mir-17-3p | Not expressed | Not expressed | 0.268 | 0.0084 |
| mir-200b-5p | 0.247 | 0.2630 | 0.004 | 0.0104 |
| mir-548d-5p | 1.926 | 0.1540 | 3.381 | 0.0148 |
| mir-503-5p | 1.883 | 0.1116 | 2.129 | 0.0154 |
| mir-505-5p | 1.287 | 0.9787 | 0.191 | 0.0169 |
| mir-223-3p | 1.314 | 0.3905 | 3.506 | 0.0226 |
| mir-29b-3p | 1.261 | 0.5726 | 2.685 | 0.0241 |
| mir-379-5p | 2.425 | 0.0378 | 0.614 | 0.3493 |
| mir-190-5p | 2.546 | 0.0478 | 1.728 | 0.1021 |
| mir-889-3p | 2.094 | 0.0571 | 0.659 | 0.5155 |
| miRNA | Ratio Non-Survivor/Survivor | p-Value * | |
|---|---|---|---|
| Amniotic Fluids | mir-889-3p | 2.02 | 0.03 |
| mir-379-5p | 2.32 | 0.03 | |
| mir-190-5p | 1.75 | 0.10 | |
| Tracheal Fluids | mir-223-3p | 2.58 | 0.03 |
| mir-503-5p | 1.64 | 0.05 | |
| mir-548d-5p | 1.63 | 0.13 | |
| mir-29b-3p | 1.46 | 0.26 | |
| mir-17-3p | 1.72 | 0.3 | |
| mir-200a-5p | 1.25 | 0.5 | |
| mir-505-5p | 0.80 | 0.55 | |
| mir-200b-5p | 1.15 | 0.73 |
| miRNA | Target |
|---|---|
| mir-223-3p | Fibroblast growth factor receptor 2, GLI family zinc finger3, Paired box protein 7, Protein-lisine 6-ossidase, Low-density lipoprotein-related protein 2, Nuclear receptor subfamily 2 group 2 member 2/Transcription factor 2 COUP, Interleukin enhancer binding factor 3, Matrix metallopeptidase 14, Platelet-derived growth factor receptor A, Transcription factor 21 |
| mir-503-5p | Low-density lipoprotein-related protein 2, Fibrillin 1, SRY-Box 7, Ephrin B1, Glypican-3, H 2.0 Like Homeobox, Homeobox protein Hox-B4, Willms tumor 1, Musculin, Platelet-derived growth factor receptor A, Transcription factor 21, Slit Guidance Ligand 3, Collagen type III Alpha 1 Chain |
| mir-379-5p | Deoxyribonuclease 2, Fibrillin 1, MET Proto-Oncogene receptor tyrosine kinase, Willms tumor 1, Retinoic acid receptor beta, Transcription Factor 21, GLI family zinc finger 3, Platelet-derived growth factor receptor A |
| mir-889-3p | Retinoic acid receptor beta, Transcription Factor 21, C-Terminal Binding Protein 2, Fibroblast growth factor receptor 2, Zinc Finger Protein FOG family member 2, GRB2-associated binding protein, Nuclear receptor subfamily 2 group 2 member 2/Transcription factor 2 COUP, SRY-Box 7, Collagen type III Alpha 1 Chain, Lysyl Oxidase, LDL Receptor Related Protein 2, Paired box protein 3, GLI family zinc finger 3, Platelet-derived growth factor receptor A, Fibrillin 1, Slit Guidance Ligand 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fabietti, I.; Nardi, T.; Favero, C.; Dioni, L.; Cantone, L.; Pergoli, L.; Hoxha, M.; Pinatel, E.; Mosca, F.; Bollati, V.; et al. Extracellular Vesicles and Their miRNA Content in Amniotic and Tracheal Fluids of Fetuses with Severe Congenital Diaphragmatic Hernia Undergoing Fetal Intervention. Cells 2021, 10, 1493. https://doi.org/10.3390/cells10061493
Fabietti I, Nardi T, Favero C, Dioni L, Cantone L, Pergoli L, Hoxha M, Pinatel E, Mosca F, Bollati V, et al. Extracellular Vesicles and Their miRNA Content in Amniotic and Tracheal Fluids of Fetuses with Severe Congenital Diaphragmatic Hernia Undergoing Fetal Intervention. Cells. 2021; 10(6):1493. https://doi.org/10.3390/cells10061493
Chicago/Turabian StyleFabietti, Isabella, Tiago Nardi, Chiara Favero, Laura Dioni, Laura Cantone, Laura Pergoli, Mirjam Hoxha, Eva Pinatel, Fabio Mosca, Valentina Bollati, and et al. 2021. "Extracellular Vesicles and Their miRNA Content in Amniotic and Tracheal Fluids of Fetuses with Severe Congenital Diaphragmatic Hernia Undergoing Fetal Intervention" Cells 10, no. 6: 1493. https://doi.org/10.3390/cells10061493
APA StyleFabietti, I., Nardi, T., Favero, C., Dioni, L., Cantone, L., Pergoli, L., Hoxha, M., Pinatel, E., Mosca, F., Bollati, V., & Persico, N. (2021). Extracellular Vesicles and Their miRNA Content in Amniotic and Tracheal Fluids of Fetuses with Severe Congenital Diaphragmatic Hernia Undergoing Fetal Intervention. Cells, 10(6), 1493. https://doi.org/10.3390/cells10061493