
The Critical Role of Growth Factors in Gastric Ulcer Healing: The Cellular and Molecular Mechanisms and Potential Clinical Implications


Abstract
:
{kind=link}
{kind=link}
{kind=link}
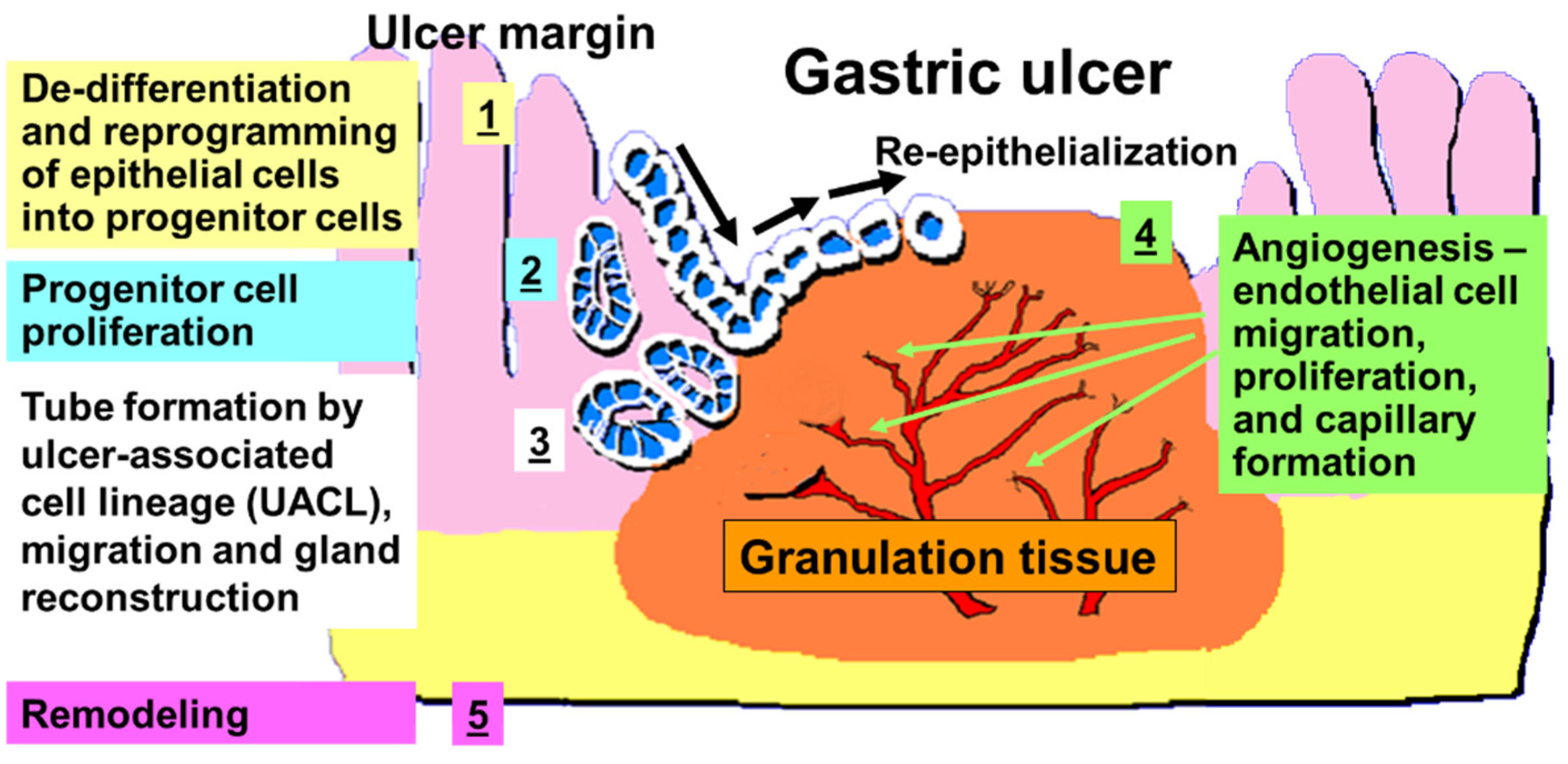{kind=link}
{kind=link}
{kind=link}
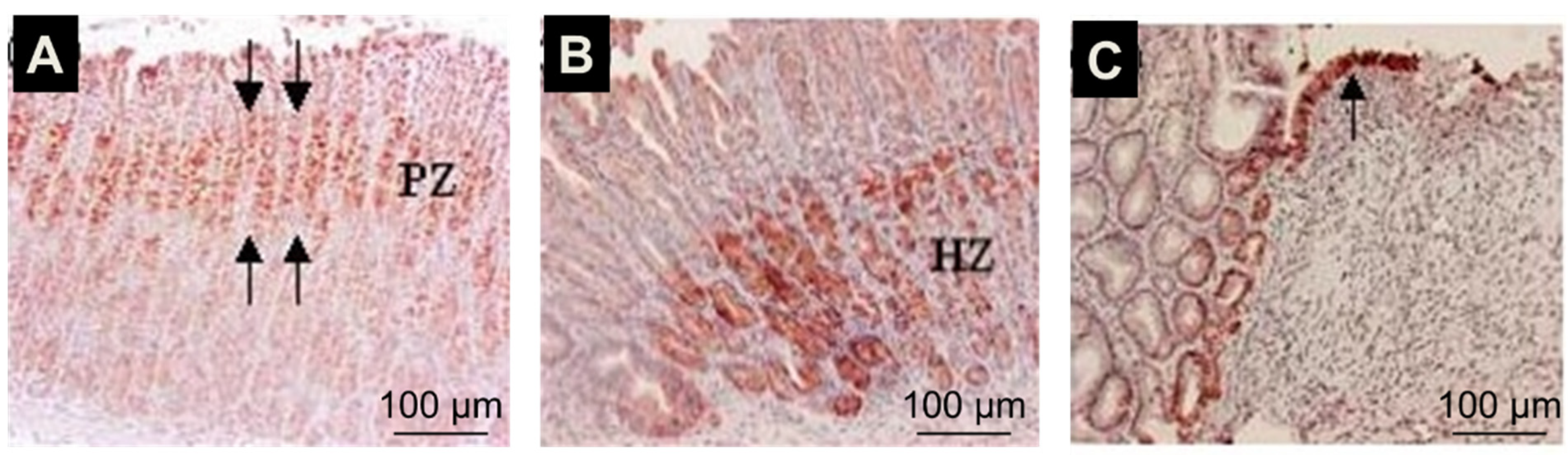{kind=link}
{kind=link}
{kind=link}
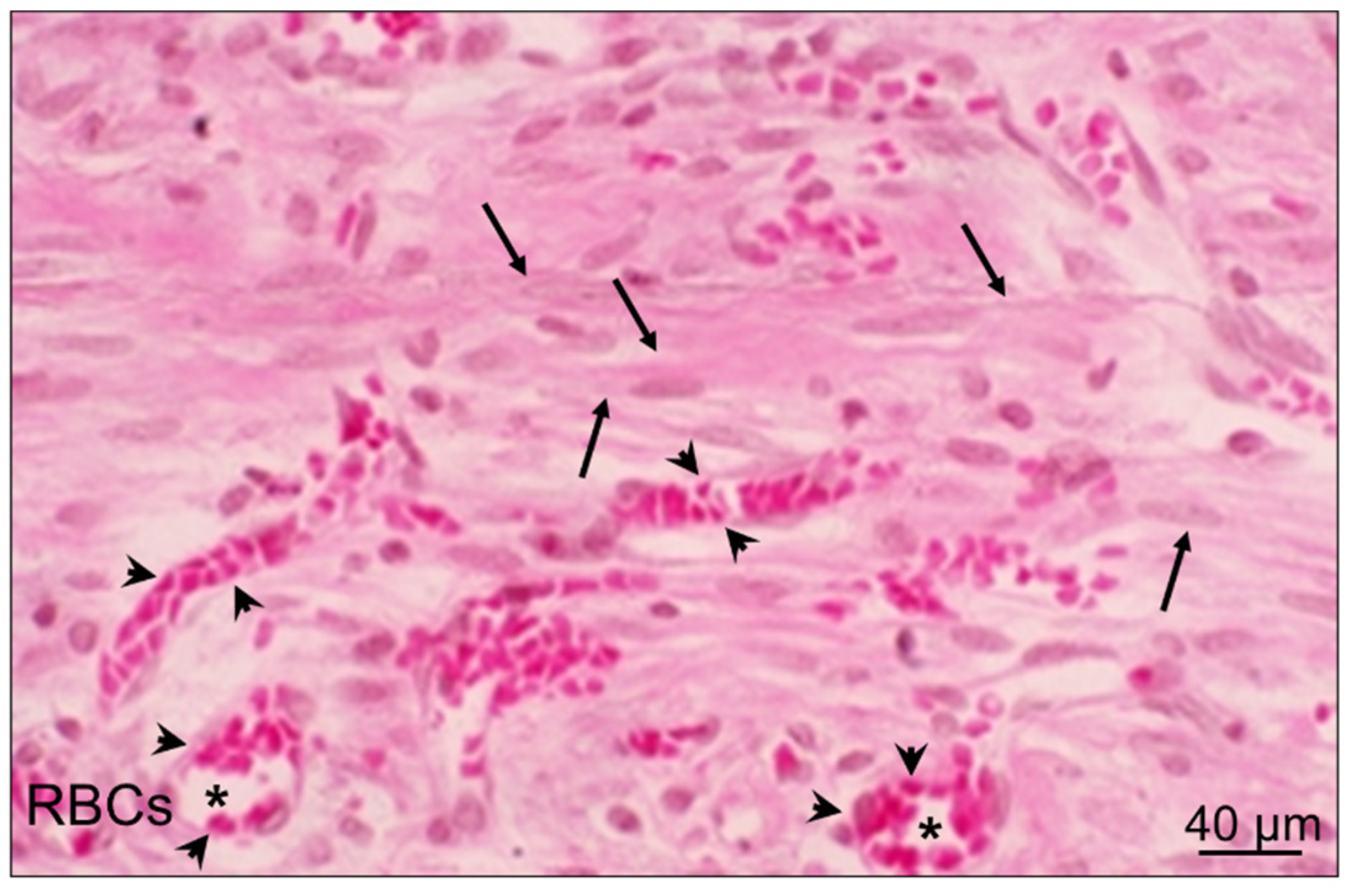{kind=link}
{kind=link}
1. Overview of Gastric Ulcer
2. Cellular and Molecular Events in the Ulcer Margin
3. Growth Factors—Discovery, Brief Historical Background, and Their Role in GU Healing
4. The Role of EGF in GU Healing
5. Trefoil Factor (TFF) Peptides and GU Healing
6. HGF and GU Healing
7. Fibroblast Growth Factors and GU Healing
8. Insulin-Like Growth Factor 1 (IGF-1) and GU Healing
9. Temporal and Spatial Gene Expression during Ulcer Healing. Epithelial/Mesenchymal Interactions and Back-Ups
10. Cellular and Molecular Events in GU Granulation Tissue
11. VEGF and Angiogenesis
12. Novel Roles of SRF in Re-Epithelialization and VEGF Induced Angiogenesis
13. The Role of SRF in GU Healing
14. A Novel Role of Nerve Growth Factor (NGF) in GU Healing
15. Summary and Future Directions
16. Potential Future Directions—Therapy of GU with Local Administration of Growth Factors?
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AKT | serine-threonine protein kinase |
| Ang | angiopoietin |
| AR | Amphiregulin |
| bFGF | basic fibroblast growth factor |
| BMD-EPCs | bone marrow derived endothelial progenitor cells |
| BTC | Betacellulin (BTC) |
| COX-2 | cyclooxygenase 2 |
| ECs | endothelial cells |
| EGF | epidermal growth factor |
| EGF-R | epidermal growth factor receptor |
| EPR | Epiregulin |
| ERK | extracellular regulated kinase |
| FGF | fibroblast growth factor |
| GT | granulation tissue |
| GU | gastric ulcer |
| HB-EGF | heparin-binding EGF-like growth factor |
| HGF | hepatocyte growth factor |
| HIF | hypoxia inducible factor |
| Hp | Helicobacter pylori |
| IGF-1 | Insulin-like growth factor 1 |
| IL-1α | interleukin-1 alpha |
| IL-lβ | interleukin-1 beta |
| KGF | Keratinocyte growth factor |
| MAPK | mitogen activated protein kinase |
| NGF | nerve growth factor |
| NRG1-4 | neuregulins 1-4 |
| NSAIDS | non-steroidal anti-inflammatory drugs |
| PDGF | platelet derived growth factor |
| PGs | prostaglandins |
| PI-3K | Phosphatidylinositol 3-kinase |
| PLCγ | Phospholipase C gamma |
| Rho/Rac | GTPases |
| RT-PCR | reverse transcription polymerase chain reaction |
| SDF-1 | stromal derived factor 1 |
| SRE | serum response element |
| SRF | serum response factor |
| TGFα | transforming growth factor alpha |
| TGFβ | transforming growth factor beta |
| TFF | Trefoil factor |
| TNFα | tumor necrosis factor alpha |
| UACL | ulcer-associated cell lineage |
| VEGF | Vascular endothelial growth factor |
References
- Tarnawski, A.S. Cellular and molecular mechanisms of gastrointestinal ulcer healing. Dig. Dis. Sci. 2005, 50 (Suppl. 1), S24–S33. [Google Scholar] [CrossRef] [PubMed]
- Tarnawski, A.; Hollander, D.; Stachura, J.; Krause, W.J.; Eltorai, M.; Dabros, W.; Gergely, H. Vascular and microvascular changes--key factors in the development of acetic acid-induced gastric ulcers in rats. J. Clin. Gastroenterol. 1990, 12 (Suppl. 1), S148–S157. [Google Scholar] [CrossRef]
- Clark, R.A.F. Wound repair: Overview and general considerations. In The Molecular and Cellular Biology of Wound Repair, 2nd ed.; Clark, R., Ed.; Plenum Press: New York, NY, USA, 1996; pp. 3–35. [Google Scholar]
- Laine, L.; Takeuchi, K.; Tarnawski, A. Gastric mucosal defense and cytoprotection: Bench to bedside. Gastroenterology 2008, 135, 41–60. [Google Scholar] [CrossRef]
- Tarnawski, A. Molecular mechanisms of ulcer healing. Drug News Perspect. 2000, 13, 158–168. [Google Scholar] [CrossRef] [PubMed]
- Cotran, R.; Kumar, V.; Robbins, S. Gastric ulceration. In Robbins Pathologic Basis of Disease, 5th ed.; Cotran, R., Kumar, V., Robbins, S., Eds.; Saunder: Philadelphia, PA, USA, 1999. [Google Scholar]
- Tarnawski, A. Cellular mechanisms of gastric ulcer healing. In The Stomach; Domschke, W., Konturek, S., Eds.; Springer: Berlin, Germany; New York, NY, USA, 1993; pp. 177–192. [Google Scholar]
- Baatar, D.; Kawanaka, H.; Szabo, I.L.; Pai, R.; Jones, M.K.; Kitano, S.; Tarnawski, A.S. Esophageal ulceration activates keratinocyte growth factor and its receptor in rats: Implications for ulcer healing. Gastroenterology 2002, 122, 458–468. [Google Scholar] [CrossRef] [PubMed]
- Tarnawski, A.; Stachura, J.; Krause, W.J.; Douglass, T.G.; Gergely, H. Quality of gastric ulcer healing: A new, emerging concept. J. Clin. Gastroenterol. 1991, 13 (Suppl. 1), S42–S47. [Google Scholar] [CrossRef]
- Vanwijck, R. Surgical biology of wound healing. Bull. Mem. Acad. R. Med. Belg. 2001, 156, 175–184; discussion 185. [Google Scholar]
- Tarnawski, A.; Hollander, D.; Krause, W.J.; Dabros, W.; Stachura, J.; Gergely, H. “Healed” experimental gastric ulcers remain histologically and ultrastructurally abnormal. J. Clin. Gastroenterol. 1990, 12 (Suppl. 1), S139–S147. [Google Scholar] [CrossRef]
- Taupin, D.; Wu, D.C.; Jeon, W.K.; Devaney, K.; Wang, T.C.; Podolsky, D.K. The trefoil gene family are coordinately expressed immediate-early genes: EGF receptor- and MAP kinase-dependent interregulation. J. Clin. Investig. 1999, 103, R31–R38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, W.M.; Playford, R.J.; Wright, N.A. Peptide gene expression in gastrointestinal mucosal ulceration: Ordered sequence or redundancy? Gut 2000, 46, 286–292. [Google Scholar] [CrossRef] [Green Version]
- Tarnawski, A.S.; Jones, M.K.; Ahluwalia, A. Dedifferentiation and reprogramming of epithelial cells during gastric ulcer healing is triggered by hypoxia and a well-coordinated, sequential activation of egfr signaling: Cross talk with IGF1 and COX2. Gastroenterology 2021, 160, S77–S78. [Google Scholar] [CrossRef]
- Tarnawski, A.; Stachura, J.; Durbin, T.; Sarfeh, I.J.; Gergely, H. Increased expression of epidermal growth factor receptor during gastric ulcer healing in rats. Gastroenterology 1992, 102, 695–698. [Google Scholar] [CrossRef]
- Wright, N.A.; Pike, C.; Elia, G. Induction of a novel epidermal growth factor-secreting cell lineage by mucosal ulceration in human gastrointestinal stem cells. Nature 1990, 343, 82–85. [Google Scholar] [CrossRef]
- Tarnawski, A.S.; Jones, M.K. The role of epidermal growth factor (EGF) and its receptor in mucosal protection, adaptation to injury, and ulcer healing: Involvement of EGF-R signal transduction pathways. J. Clin. Gastroenterol. 1998, 27 (Suppl. 1), S12–S20. [Google Scholar] [CrossRef]
- Okamoto, R.; Yajima, T.; Yamazaki, M.; Kanai, T.; Mukai, M.; Okamoto, S.; Ikeda, Y.; Hibi, T.; Inazawa, J.; Watanabe, M. Damaged epithelia regenerated by bone marrow-derived cells in the human gastrointestinal tract. Nat. Med. 2002, 8, 1011–1017. [Google Scholar] [CrossRef]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gurdon, J.B. The developmental capacity of nuclei taken from intestinal epithelium cells of feeding tadpoles. J. Embryol. Exp. Morphol. 1962, 10, 622–640. [Google Scholar] [PubMed]
- He, S.; Liu, N.; Bayliss, G.; Zhuang, S. EGFR activity is required for renal tubular cell dedifferentiation and proliferation in a murine model of folic acid-induced acute kidney injury. Am. J. Physiol. Renal. Physiol. 2013, 304, F356–F366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang-Panesso, M.; Kadyrov, F.F.; Lalli, M.; Wu, H.; Ikeda, S.; Kefaloyianni, E.; Abdelmageed, M.M.; Herrlich, A.; Kobayashi, A.; Humphreys, B.D. FOXM1 drives proximal tubule proliferation during repair from acute ischemic kidney injury. J. Clin. Investig. 2019, 129, 5501–5517. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, G.; Cohen, S. Epidermal growth factor. Annu. Rev. Biochem. 1979, 48, 193–216. [Google Scholar] [CrossRef]
- Cohen, S. Nobel lecture. Epidermal growth factor. Biosci. Rep. 1986, 6, 1017–1028. [Google Scholar] [CrossRef]
- Cohen, S. Epidermal growth factor and its receptor. Eur. J. Gastroenterol. Hepatol. 1993, 5, S3–S4. [Google Scholar]
- Cohen, S.; Levi-Montalcini, R. A Nerve Growth-Stimulating Factor Isolated from Snake Venom. Proc. Natl. Acad. Sci. USA 1956, 42, 571–574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basson, M.D.; Modlin, I.M.; Turowski, G.; Madri, J.A. Enterocyte-matrix interactions in the healing of mucosal injury. Eur. J. Gastroenterol. Hepatol. 1993, 5, S21–S29. [Google Scholar]
- Tamawski, A.; Santos, A.M.; Ichikawa, Y.; Stachura, J.; Krause, W.J. Development, evolution and healing of experimental gastric ulcers: An overview. Eur. J. Gastroenterol. Hepatol. 1993, 5, S5–S12. [Google Scholar]
- Sabbah, D.A.; Hajjo, R.; Sweidan, K. Review on Epidermal Growth Factor Receptor (EGFR) Structure, Signaling Pathways, Interactions, and Recent Updates of EGFR Inhibitors. Curr. Top. Med. Chem. 2020, 20, 815–834. [Google Scholar] [CrossRef]
- Singh, B.; Carpenter, G.; Coffey, R.J. EGF receptor ligands: Recent advances. F1000Research 2016, 5. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.X.; Yang, J.J.; Zhang, S.A.; Zhang, S.M.; Wang, J.X.; Xu, Z.Y.; Lin, R.Y. HIF-1alpha promotes inflammatory response of chronic obstructive pulmonary disease by activating EGFR/PI3K/AKT pathway. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 6077–6084. [Google Scholar] [CrossRef] [PubMed]
- Basson, M.D.; Modlin, I.M.; Madri, J.A. Human enterocyte (Caco-2) migration is modulated in vitro by extracellular matrix composition and epidermal growth factor. J. Clin. Investig. 1992, 90, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Brzozowski, T.; Konturek, P.C.; Konturek, S.J.; Schuppan, D.; Drozdowicz, D.; Kwiecien, S.; Majka, J.; Nakamura, T.; Hahn, E. Effect of local application of growth factors on gastric ulcer healing and mucosal expression of cyclooxygenase-1 and -2. Digestion 2001, 64, 15–29. [Google Scholar] [CrossRef] [PubMed]
- Pai, R.; Soreghan, B.; Szabo, I.L.; Pavelka, M.; Baatar, D.; Tarnawski, A.S. Prostaglandin E2 transactivates EGF receptor: A novel mechanism for promoting colon cancer growth and gastrointestinal hypertrophy. Nat. Med. 2002, 8, 289–293. [Google Scholar] [CrossRef]
- Pai, R.; Ohta, M.; Itani, R.M.; Sarfeh, I.J.; Tarnawski, A.S. Induction of mitogen-activated protein kinase signal transduction pathway during gastric ulcer healing in rats. Gastroenterology 1998, 114, 706–713. [Google Scholar] [CrossRef]
- Pai, R.; Jones, M.K.; Tomikawa, M.; Tarnawski, A.S. Activation of Raf-1 during experimental gastric ulcer healing is Ras-mediated and protein kinase C-independent. Am. J. Pathol. 1999, 155, 1759–1766. [Google Scholar] [CrossRef] [Green Version]
- Yetkin, G.; Celebi, N.; Ozer, C.; Gonul, B.; Ozogul, C. The healing effect of TGF-alpha on gastric ulcer induced by acetylsalicylic acid in rats. Int. J. Pharm. 2004, 277, 163–172. [Google Scholar] [CrossRef]
- Aihara, E.; Engevik, K.A.; Montrose, M.H. Trefoil Factor Peptides and Gastrointestinal Function. Annu. Rev. Physiol. 2017, 79, 357–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Busch, M.; Dunker, N. Trefoil factor family peptides--friends or foes? Biomol. Concepts 2015, 6, 343–359. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, W. Trefoil factors TFF (trefoil factor family) peptide-triggered signals promoting mucosal restitution. Cell. Mol. Life Sci. 2005, 62, 2932–2938. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, C.; Santamatilde, E.; McCreath, K.J.; Cervera, A.M.; Diez, I.; Ortiz-Masia, D.; Martinez, N.; Calatayud, S.; Esplugues, J.V.; Barrachina, M.D. Induction of trefoil factor (TFF)1, TFF2 and TFF3 by hypoxia is mediated by hypoxia inducible factor-1: Implications for gastric mucosal healing. Br. J. Pharmacol. 2009, 156, 262–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farrell, J.J.; Taupin, D.; Koh, T.J.; Chen, D.; Zhao, C.M.; Podolsky, D.K.; Wang, T.C. TFF2/SP-deficient mice show decreased gastric proliferation, increased acid secretion, and increased susceptibility to NSAID injury. J. Clin. Investig. 2002, 109, 193–204. [Google Scholar] [CrossRef]
- Buda, A.; Jepson, M.A.; Pignatelli, M. Regulatory function of trefoil peptides (TFF) on intestinal cell junctional complexes. Cell Commun. Adhes 2012, 19, 63–68. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; el-Hariry, I.; Karayiannakis, A.J.; Wilding, J.; Chinery, R.; Kmiot, W.; McCrea, P.D.; Gullick, W.J.; Pignatelli, M. Phosphorylation of beta-catenin and epidermal growth factor receptor by intestinal trefoil factor. Lab. Investig. 1997, 77, 557–563. [Google Scholar]
- Nakamura, T.; Mizuno, S. The discovery of hepatocyte growth factor (HGF) and its significance for cell biology, life sciences and clinical medicine. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2010, 86, 588–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, M.K.; Sasaki, E.; Halter, F.; Pai, R.; Nakamura, T.; Arakawa, T.; Kuroki, T.; Tarnawski, A.S. HGF triggers activation of the COX-2 gene in rat gastric epithelial cells: Action mediated through the ERK2 signaling pathway. FASEB J. 1999, 13, 2186–2194. [Google Scholar] [CrossRef] [Green Version]
- Beenken, A.; Mohammadi, M. The FGF family: Biology, pathophysiology and therapy. Nat. Rev. Drug Discov. 2009, 8, 235–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, N.; Grose, R. Fibroblast growth factor signalling: From development to cancer. Nat. Rev. Cancer 2010, 10, 116–129. [Google Scholar] [CrossRef] [PubMed]
- Szabo, S.; Folkman, J.; Vattay, P.; Morales, R.E.; Pinkus, G.S.; Kato, K. Accelerated healing of duodenal ulcers by oral administration of a mutein of basic fibroblast growth factor in rats. Gastroenterology 1994, 106, 1106–1111. [Google Scholar] [CrossRef]
- Ernst, H.; Konturek, P.C.; Hahn, E.G.; Stosiek, H.P.; Brzozowski, T.; Konturek, S.J. Effect of local injection with basic fibroblast growth factor (bFGF) and neutralizing antibody to bFGF on gastric ulcer healing, gastric secretion, angiogenesis and gastric blood flow. J. Physiol. Pharmacol. 2001, 52, 377–390. [Google Scholar]
- Folkman, J.; Szabo, S.; Stovroff, M.; McNeil, P.; Li, W.; Shing, Y. Duodenal ulcer. Discovery of a new mechanism and development of angiogenic therapy that accelerates healing. Ann. Surg. 1991, 214, 414–425. [Google Scholar] [CrossRef]
- Szabo, S.; Vincze, A. Growth factors in ulcer healing: Lessons from recent studies. J. Physiol. Paris 2000, 94, 77–81. [Google Scholar] [CrossRef]
- Florkiewicz, R.Z.; Ahluwalia, A.; Sandor, Z.; Szabo, S.; Tarnawski, A.S. Gastric mucosal injury activates bFGF gene expression and triggers preferential translation of high molecular weight bFGF isoforms through CUG-initiated, non-canonical codons. Biochem. Biophys. Res. Commun. 2011, 409, 494–499. [Google Scholar] [CrossRef]
- Rothstein, H. Regulation of the cell cycle by somatomedins. Int. Rev. Cytol. 1982, 78, 127–232. [Google Scholar] [CrossRef] [PubMed]
- LeRoith, D.; Werner, H.; Beitner-Johnson, D.; Roberts, C.T., Jr. Molecular and cellular aspects of the insulin-like growth factor I receptor. Endocr. Rev. 1995, 16, 143–163. [Google Scholar] [CrossRef]
- Rabinovsky, E.D. The multifunctional role of IGF-1 in peripheral nerve regeneration. Neurol. Res. 2004, 26, 204–210. [Google Scholar] [CrossRef] [PubMed]
- Laburthe, M.; Rouyer-Fessard, C.; Gammeltoft, S. Receptors for insulin-like growth factors I and II in rat gastrointestinal epithelium. Am. J. Physiol. 1988, 254, G457–G462. [Google Scholar] [CrossRef] [PubMed]
- Costigan, D.C.; Guyda, H.J.; Posner, B.I. Free insulin-like growth factor I (IGF-I) and IGF-II in human saliva. J. Clin. Endocrinol. Metab. 1988, 66, 1014–1018. [Google Scholar] [CrossRef] [Green Version]
- Chaurasia, O.P.; Marcuard, S.P.; Seidel, E.R. Insulin-like growth factor I in human gastrointestinal exocrine secretions. Regul. Pept. 1994, 50, 113–119. [Google Scholar] [CrossRef]
- Nguyen, T.; Chai, J.; Li, A.; Akahoshi, T.; Tanigawa, T.; Tarnawski, A.S. Novel roles of local insulin-like growth factor-1 activation in gastric ulcer healing: Promotes actin polymerization, cell proliferation, re-epithelialization, and induces cyclooxygenase-2 in a phosphatidylinositol 3-kinase-dependent manner. Am. J. Pathol. 2007, 170, 1219–1228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrara, N. VEGF as a therapeutic target in cancer. Oncology 2005, 69 (Suppl. 3), 11–16. [Google Scholar] [CrossRef] [PubMed]
- Nico, B.; Mangieri, D.; Benagiano, V.; Crivellato, E.; Ribatti, D. Nerve growth factor as an angiogenic factor. Microvasc. Res. 2008, 75, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Carmeliet, P. Angiogenesis in health and disease. Nat. Med. 2003, 9, 653–660. [Google Scholar] [CrossRef]
- Ahluwalia, A.; Brzozowski, T.; Jones, M.K.; Ichikawa, Y.; Tarnawski, A.S. Formation of new blood vessels during gastric ulcer healing. Role of bone marrow derived endothelial progenitor cells. J. Physiol. Pharmacol. 2017, 68, 585–589. [Google Scholar]
- Ahluwalia, A.; Jones, M.K.; Brzozowska, I.; Tarnawski, A.S. In vitro model of vasculo-angiogenesis: Demonstration that bone marrow derived endothelial progenitor cells form new hybrid capillary blood vessels jointly with gastric endothelial cells. J. Physiol. Pharmacol. 2017, 68, 841–846. [Google Scholar]
- Tarnawski, A.S.; Ahluwalia, A. Molecular mechanisms of epithelial regeneration and neovascularization during healing of gastric and esophageal ulcers. Curr. Med. Chem. 2012, 19, 16–27. [Google Scholar] [CrossRef]
- Tarnawski, A.S.; Ahluwalia, A.; Jones, M.K. Angiogenesis in gastric mucosa: An important component of gastric erosion and ulcer healing and its impairment in aging. J. Gastroenterol. Hepatol. 2014, 29 (Suppl. 4), 112–123. [Google Scholar] [CrossRef]
- Risau, W. Mechanisms of angiogenesis. Nature 1997, 386, 671–674. [Google Scholar] [CrossRef] [PubMed]
- Risau, W.; Flamme, I. Vasculogenesis. Annu. Rev. Cell Dev. Biol. 1995, 11, 73–91. [Google Scholar] [CrossRef]
- Carmeliet, P.; Ferreira, V.; Breier, G.; Pollefeyt, S.; Kieckens, L.; Gertsenstein, M.; Fahrig, M.; Vandenhoeck, A.; Harpal, K.; Eberhardt, C.; et al. Abnormal blood vessel development and lethality in embryos lacking a single VEGF allele. Nature 1996, 380, 435–439. [Google Scholar] [CrossRef] [PubMed]
- Ferrara, N.; Carver-Moore, K.; Chen, H.; Dowd, M.; Lu, L.; O’Shea, K.S.; Powell-Braxton, L.; Hillan, K.J.; Moore, M.W. Heterozygous embryonic lethality induced by targeted inactivation of the VEGF gene. Nature 1996, 380, 439–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asahara, T.; Kawamoto, A. Endothelial progenitor cells for postnatal vasculogenesis. Am. J. Physiol. Cell Physiol. 2004, 287, C572–C579. [Google Scholar] [CrossRef] [Green Version]
- Balaji, S.; King, A.; Crombleholme, T.M.; Keswani, S.G. The Role of Endothelial Progenitor Cells in Postnatal Vasculogenesis: Implications for Therapeutic Neovascularization and Wound Healing. Adv. Wound Care 2013, 2, 283–295. [Google Scholar] [CrossRef] [Green Version]
- Matta, A.; Nader, V.; Galinier, M.; Roncalli, J. Transplantation of CD34+ cells for myocardial ischemia. World J. Transplant. 2021, 11, 138–146. [Google Scholar] [CrossRef]
- Naito, H.; Iba, T.; Takakura, N. Mechanisms of new blood-vessel formation and proliferative heterogeneity of endothelial cells. Int. Immunol. 2020, 32, 295–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, X.; Szabo, S.; Chen, L.; Paunovic, B.; Khomenko, T.; Tolstanova, G.; Tarnawski, A.S.; Jones, M.K.; Sandor, Z. New cell therapy using bone marrow-derived stem cells/endothelial progenitor cells to accelerate neovascularization in healing of experimental ulcerative colitis. Curr. Pharm. Des. 2011, 17, 1643–1651. [Google Scholar] [CrossRef] [PubMed]
- Ferrara, N. The role of VEGF in the regulation of physiological and pathological angiogenesis. Mech. Angiogenesis 2005, 209–231. [Google Scholar] [CrossRef]
- Ferrara, N. Vascular endothelial growth factor: Basic science and clinical progress. Endocr. Rev. 2004, 25, 581–611. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.K.; Kawanaka, H.; Baatar, D.; Szabo, I.L.; Tsugawa, K.; Pai, R.; Koh, G.Y.; Kim, I.; Sarfeh, I.J.; Tarnawski, A.S. Gene therapy for gastric ulcers with single local injection of naked DNA encoding VEGF and angiopoietin-1. Gastroenterology 2001, 121, 1040–1047. [Google Scholar] [CrossRef]
- Chai, J.; Baatar, D.; Tarnawski, A. Serum response factor promotes re-epithelialization and muscular structure restoration during gastric ulcer healing. Gastroenterology 2004, 126, 1809–1818. [Google Scholar] [CrossRef]
- Treisman, R. Identification of a protein-binding site that mediates transcriptional response of the c-fos gene to serum factors. Cell 1986, 46, 567–574. [Google Scholar] [CrossRef]
- Chai, J.; Baatar, D.; Moon, W.; Tarnawski, A. Expression of serum response factor in normal rat gastric mucosa. J. Physiol. Pharmacol. 2002, 53, 289–294. [Google Scholar]
- Schratt, G.; Philippar, U.; Berger, J.; Schwarz, H.; Heidenreich, O.; Nordheim, A. Serum response factor is crucial for actin cytoskeletal organization and focal adhesion assembly in embryonic stem cells. J. Cell Biol. 2002, 156, 737–750. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Zhe, X.; Phan, S.H.; Ullenbruch, M.; Schuger, L. Involvement of serum response factor isoforms in myofibroblast differentiation during bleomycin-induced lung injury. Am. J. Respir. Cell Mol. Biol. 2003, 29, 583–590. [Google Scholar] [CrossRef]
- Iyer, V.R.; Eisen, M.B.; Ross, D.T.; Schuler, G.; Moore, T.; Lee, J.C.; Trent, J.M.; Staudt, L.M.; Hudson, J., Jr.; Boguski, M.S.; et al. The transcriptional program in the response of human fibroblasts to serum. Science 1999, 283, 83–87. [Google Scholar] [CrossRef]
- Chai, J.; Jones, M.K.; Tarnawski, A.S. Serum response factor is a critical requirement for VEGF signaling in endothelial cells and VEGF-induced angiogenesis. FASEB J. 2004, 18, 1264–1266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levi-Montalcini, R. The nerve growth factor 35 years later. Science 1987, 237, 1154–1162. [Google Scholar] [CrossRef]
- Levi-Montalcini, R. The nerve growth factor and the neuroscience chess board. Prog. Brain Res. 2004, 146, 525–527. [Google Scholar] [PubMed]
- Shooter, E.M. Early days of the nerve growth factor proteins. Annu. Rev. Neurosci. 2001, 24, 601–629. [Google Scholar] [CrossRef] [PubMed]
- Hosang, M.; Shooter, E.M. The internalization of nerve growth factor by high-affinity receptors on pheochromocytoma PC12 cells. EMBO J. 1987, 6, 1197–1202. [Google Scholar] [CrossRef] [PubMed]
- Misko, T.P.; Radeke, M.J.; Shooter, E.M. Nerve growth factor in neuronal development and maintenance. J. Exp. Biol. 1987, 132, 177–190. [Google Scholar] [CrossRef]
- Madduri, S.; Papaloizos, M.; Gander, B. Synergistic effect of GDNF and NGF on axonal branching and elongation in vitro. Neurosci. Res. 2009, 65, 88–97. [Google Scholar] [CrossRef]
- Freeman, R.S.; Burch, R.L.; Crowder, R.J.; Lomb, D.J.; Schoell, M.C.; Straub, J.A.; Xie, L. NGF deprivation-induced gene expression: After ten years, where do we stand? Prog. Brain Res. 2004, 146, 111–126. [Google Scholar]
- Ahluwalia, A.; Jones, M.K.; Brzozowski, T.; Tarnawski, A.S. Nerve growth factor is critical requirement for in vitro angiogenesis in gastric endothelial cells. Am. J. Physiol. Gastrointest Liver Physiol. 2016, 311, G981–G987. [Google Scholar] [CrossRef] [Green Version]
- Tanigawa, T.; Ahluwalia, A.; Watanabe, T.; Arakawa, T.; Tarnawski, A.S. Nerve growth factor injected into the gastric ulcer base incorporates into endothelial, neuronal, glial and epithelial cells: Implications for angiogenesis, mucosal regeneration and ulcer healing. J. Physiol. Pharmacol. 2015, 66, 617–621. [Google Scholar]
- Ahluwalia, A.; Jones, M.K.; Hoa, N.; Zhu, E.; Brzozowski, T.; Tarnawski, A.S. Reduced NGF in gastric endothelial cells is one of the main causes of impaired angiogenesis in aging gastric mucosa. Cell Mol. Gastroenterol. Hepatol. 2018, 6, 199–213. [Google Scholar] [CrossRef]
- Tarnawski, A.; Douglass, T.G.; Stachura, J.; Krause, W.J. Quality of gastric ulcer healing: Histological and ultrastructural assessment. Aliment. Pharmacol. Ther. 1991, 5 (Suppl. 1), 79–90. [Google Scholar] [CrossRef] [PubMed]
- Aihara, E.; Matthis, A.L.; Karns, R.A.; Engevik, K.A.; Jiang, P.; Wang, J.; Yacyshyn, B.R.; Montrose, M.H. Epithelial Regeneration After Gastric Ulceration Causes Prolonged Cell-Type Alterations. Cell Mol. Gastroenterol. Hepatol. 2016, 2, 625–647. [Google Scholar] [CrossRef] [Green Version]
- Matthis, A.L.; Kaji, I.; Engevik, K.A.; Akiba, Y.; Kaunitz, J.D.; Montrose, M.H.; Aihara, E. Deficient Active Transport Activity in Healing Mucosa After Mild Gastric Epithelial Damage. Dig. Dis. Sci. 2020, 65, 119–131. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, A.C.; Briquez, P.S.; Hubbell, J.A.; Cochran, J.R. Engineering growth factors for regenerative medicine applications. Acta Biomater. 2016, 30, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Oncel, S.; Gupta, R.; Wang, Q.; Basson, M.D. ZINC40099027 Promotes Gastric Mucosal Repair in Ongoing Aspirin-Associated Gastric Injury by Activating Focal Adhesion Kinase. Cells 2021, 10, 908. [Google Scholar] [CrossRef]
- More, S.K.; Wang, Q.; Vomhof-DeKrey, E.E.; Porter, J.E.; Basson, M.D. ZINC40099027 activates human focal adhesion kinase by accelerating the enzymatic activity of the FAK kinase domain. Pharmacol. Res. Perspect. 2021, 9, e00737. [Google Scholar] [CrossRef]
- Wang, Q.; More, S.K.; Vomhof-DeKrey, E.E.; Golovko, M.Y.; Basson, M.D. Small molecule FAK activator promotes human intestinal epithelial monolayer wound closure and mouse ulcer healing. Sci. Rep. 2019, 9, 14669. [Google Scholar] [CrossRef] [Green Version]
- Bruggeman, K.F.; Wang, Y.; Maclean, F.L.; Parish, C.L.; Williams, R.J.; Nisbet, D.R. Temporally controlled growth factor delivery from a self-assembling peptide hydrogel and electrospun nanofibre composite scaffold. Nanoscale 2017, 9, 13661–13669. [Google Scholar] [CrossRef] [PubMed]
- Yamakawa, S.; Hayashida, K. Advances in surgical applications of growth factors for wound healing. Burns Trauma 2019, 7, 10. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Geng, Z.; Ma, K.; Sun, X.; Fu, X. Efficacy of Topical Recombinant Human Epidermal Growth Factor for Treatment of Diabetic Foot Ulcer: A Systematic Review and Meta-Analysis. Int. J. Low Extrem. Wounds 2016, 15, 120–125. [Google Scholar] [CrossRef] [PubMed]
- Zubair, M.; Ahmad, J. Role of growth factors and cytokines in diabetic foot ulcer healing: A detailed review. Rev. Endocr. Metab. Disord. 2019, 20, 207–217. [Google Scholar] [CrossRef] [PubMed]
- Barrientos, S.; Brem, H.; Stojadinovic, O.; Tomic-Canic, M. Clinical application of growth factors and cytokines in wound healing. Wound Repair. Regen. 2014, 22, 569–578. [Google Scholar] [CrossRef] [Green Version]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tarnawski, A.S.; Ahluwalia, A. The Critical Role of Growth Factors in Gastric Ulcer Healing: The Cellular and Molecular Mechanisms and Potential Clinical Implications. Cells 2021, 10, 1964. https://doi.org/10.3390/cells10081964
Tarnawski AS, Ahluwalia A. The Critical Role of Growth Factors in Gastric Ulcer Healing: The Cellular and Molecular Mechanisms and Potential Clinical Implications. Cells. 2021; 10(8):1964. https://doi.org/10.3390/cells10081964
Chicago/Turabian StyleTarnawski, Andrzej S., and Amrita Ahluwalia. 2021. "The Critical Role of Growth Factors in Gastric Ulcer Healing: The Cellular and Molecular Mechanisms and Potential Clinical Implications" Cells 10, no. 8: 1964. https://doi.org/10.3390/cells10081964
APA StyleTarnawski, A. S., & Ahluwalia, A. (2021). The Critical Role of Growth Factors in Gastric Ulcer Healing: The Cellular and Molecular Mechanisms and Potential Clinical Implications. Cells, 10(8), 1964. https://doi.org/10.3390/cells10081964