
The Role of MiR-181 Family Members in Endothelial Cell Dysfunction and Tumor Angiogenesis


Abstract
:1. Introduction
2. MiR-181 Family Members
3. MiR-181 in EC Differentiation
4. MiR-181 in EC Barriers of the Blood-Brain Barrier (BBB) and Blood-Tumor Barrier (BTB)
5. The Pathophysiological Role of MiR-181 in EC Angiogenesis
6. MiR-181 in Tumor Angiogenesis

{kind=link}
{kind=link}
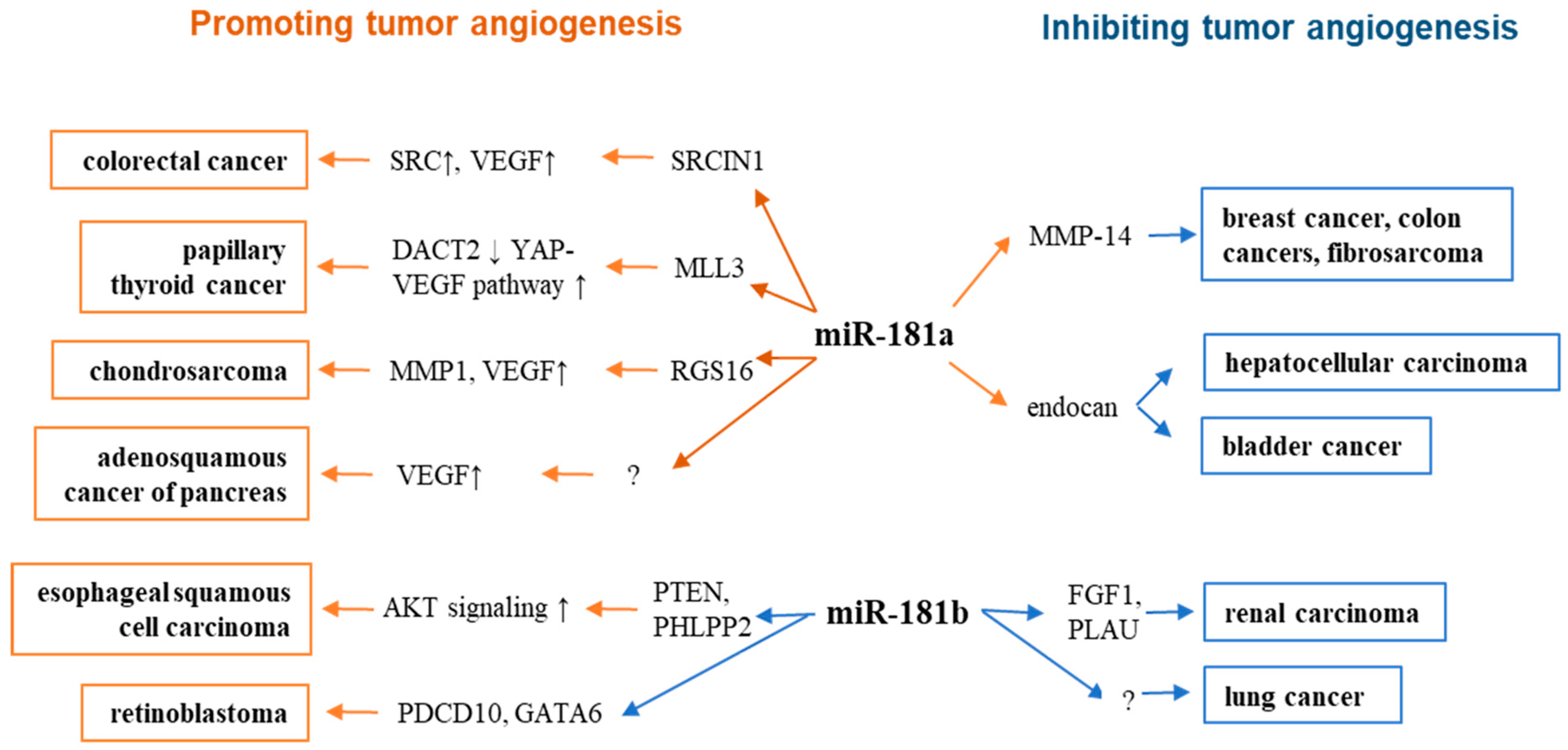
| Activity in Tumor Angiogenesis | MiR-181 | Upstream Events | Targets | Downstream Signaling/Events | Ref |
|---|---|---|---|---|---|
| inhibits angiogenesis in breast cancer, colon cancer, fibrosarcoma | miR-181a-5p | MMP-14 | [85] | ||
| inhibits angiogenesis in bladder cancer and hepatocarcinoma | miR-181a-5p | endocan | VEGF signaling↓ | [86,87] | |
| promotes angiogenesis in ASCP | miR-181a-5p | VEGF signaling↑ | [15] | ||
| promotes angiogenesis in PTC | exosomal miR-181a | hypoxic | MLL3 | DACT2↓, YAP-VEGF signaling↑ | [89] |
| promotes angiogenesis in CRC | miR-181a | SRCIN1 | SRC-VEGF pathway↑ | [92] | |
| promotes angiogenesis in chondrosarcoma | miR-181a | RGS16 | CXCR4 signaling↑, VEGF↑, MMP1↑ | [88,95] | |
| promotes angiogenesis in ESCC | miR-181b-5p | PTEN, PHLPP2 | Akt signaling↑ | [99] | |
| promotes angiogenesis in Rb | miR-181b | hypoxia | PDCD10, GATA6 | [100] | |
| inhibits angiogenesis in renal carcinomas | miR-181b | FGF1, PLAU | FGFR signaling↓ | [101] | |
| inhibits angiogenesis in lung cancer | miR-181b-3p | EC sprouting↓ | [102] |
7. Potential Clinical Application of AngiomiR-Based Cancer Therapy
8. Conclusions
Author Contributions
Funding
Informed Consent Statement
Conflicts of Interest
Abbreviations
| 3′-UTRs | 3′-untranslated region |
| AKT/PKB | protein kinase B |
| AMO | anti-miRNA oligonucleotides |
| Angs | angiopoietins |
| ASCP | adenosquamous carcinoma of the pancreas |
| ASCs | adipose-derived stromal/stem cells |
| BBB | blood-brain barrier |
| BCL2 | B-cell lymphoma 2 |
| BMECs | brain microvascular endothelial cells |
| BM-EPCs | bone marrow-derived endothelial progenitor cells |
| BSCB | blood-spinal cord barrier |
| BTB | blood-tumor barrier |
| CaM | calmodulin |
| CaMKII | calcium/calmodulin-dependent protein kinase II |
| CCN1 | cellular communication network factor 1 |
| CDKN3 | cyclin-dependent kinase inhibitor 3 |
| circRNAs | circular RNAs |
| CNS | central nervous system |
| CRC | colorectal cancer |
| CXCR4 | CXC chemokine receptor 4 |
| DACT2 | disheveled binding antagonist of beta-catenin 2 |
| DM | diabetes mellitus |
| DME | diabetic macular edema |
| ECs | endothelial cells |
| ERK | extracellular signal-regulated kinase |
| EVs | extracellular vesicles |
| FGF | fibroblast growth factor |
| FMRP | fragile X mental retardation protein |
| FoxO1 | forkhead box protein O1 |
| GAS5 | growth arrest-specific 5 |
| GECs | glioma endothelial cells |
| HDL | high-density lipoprotein |
| hESCs | human embryonic stem cells |
| HIF | hypoxia-inducible factor |
| HLSC | human liver stem-like cells |
| HLSC | human liver stem cells |
| HMEC-1 | human microvascular endothelial cell |
| HUVECs | human umbilical vein endothelial cells |
| KLF6 | Kruppel-like factor 6 |
| LIF | leukemia inhibitory factor |
| LMECs | pulmonary capillary endothelial cells |
| lncRNA | long non-coding RNAs |
| MAPK | mitogen-activated protein kinase |
| MDA | malondialdehyde |
| miRNA | microRNAs |
| MLL3 | histone-lysine N-methyltransferase-3 |
| MMPs | matrix metalloproteinases |
| NCAPG | non-SMC condensin I complex subunit G |
| ncRNAs | non-coding RNAs |
| NEAT1 | nuclear paraspeckle assembly transcript 1 |
| NHO-1 | nuclear heme oxygenase-1 |
| NPI | nanopiece |
| NPs | nanoparticles |
| OIR | oxygen-induced retinopathy |
| PDGF | platelet-derived growth factor |
| PDPK1 | 3-phosphoinositide-dependent protein kinase-1 |
| pre-miRNA | precursors miRNA |
| pri-miRNA | primary miRNA |
| Prox 1 | prospero homeobox |
| PTC | papillary thyroid cancer |
| Rb | retinoblastoma |
| RGS16 | regulator of G-protein signaling 16 |
| RISC | ribonucleic acid-induced silencing complex |
| RNV | retinal neovascularization |
| ROP | retinopathy of prematurity |
| ROS | reactive oxygen species |
| SFRP1 | secreted frizzled-related protein-1 |
| SOX5 | sex-determining region Y-box protein 5 |
| T2DM | type 2 diabetes mellitus |
| TGF-β | transforming growth factor-beta |
| TIMP3 | tissue inhibitor of metalloproteinases 3 |
| TRPM7 | transient receptor potential melastatin 7 |
| uPA | urokinase-type plasminogen activator |
| VEGF | vascular endothelial growth factor |
| VHL | von Hippel-Lindau |
| ZO | zonula occludins |
References
- Patel-Hett, S.; D’Amore, P.A. Signal transduction in vasculogenesis and developmental angiogenesis. Int. J. Dev. Biol. 2011, 55, 353–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potente, M.; Gerhardt, H.; Carmeliet, P. Basic and Therapeutic Aspects of Angiogenesis. Cell 2011, 146, 873–887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carmeliet, P.; Jain, R.K. Molecular mechanisms and clinical applications of angiogenesis. Nature 2011, 473, 298–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, R.H.; Alitalo, K. Molecular regulation of angiogenesis and lymphangiogenesis. Nat. Rev. Mol. Cell Biol. 2007, 8, 464–478. [Google Scholar] [CrossRef]
- Rust, R.; Gantner, C.; Schwab, M.E. Pro- and antiangiogenic therapies: Current status and clinical implications. FASEB J. 2019, 33, 34–48. [Google Scholar] [CrossRef]
- Hernández-Romero, I.; Guerra-Calderas, L.; Salgado-Albarrán, M.; Maldonado-Huerta, T.; Soto-Reyes, E. The Regulatory Roles of Non-coding RNAs in Angiogenesis and Neovascularization From an Epigenetic Perspective. Front. Oncol. 2019, 9, 1091. [Google Scholar] [CrossRef]
- Yang, W.; Lee, D.Y.; Ben-David, Y. The roles of microRNAs in tumorigenesis and angiogenesis. Int. J. Physiol. Pathophysiol. Pharmacol. 2011, 3, 140–155. [Google Scholar]
- Ebbesen, K.K.; Hansen, T.; Kjems, J. Insights into circular RNA biology. RNA Biol. 2017, 14, 1035–1045. [Google Scholar] [CrossRef]
- Hansen, T.B.; Jensen, T.I.; Clausen, B.H.; Bramsen, J.B.; Finsen, B.; Damgaard, C.K.; Kjems, J. Natural RNA circles function as efficient microRNA sponges. Nature 2013, 495, 384–388. [Google Scholar] [CrossRef]
- Han, J.; Lee, Y.; Yeom, K.-H.; Kim, Y.-K.; Jin, H.; Kim, V.N. The Drosha-DGCR8 complex in primary microRNA processing. Genes Dev. 2004, 18, 3016–3027. [Google Scholar] [CrossRef] [Green Version]
- Bartel, D.P. MicroRNAs: Genomics, Biogenesis, Mechanism, and Function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Kuehbacher, A.; Urbich, C.; Zeiher, A.M.; Dimmeler, S. Role of Dicer and Drosha for Endothelial MicroRNA Expression and Angiogenesis. Circ. Res. 2007, 101, 59–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, W.J.; Yang, D.D.; Na, S.; Sandusky, G.E.; Zhang, Q.; Zhao, G. Dicer Is Required for Embryonic Angiogenesis during Mouse Development. J. Biol. Chem. 2005, 280, 9330–9335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.-Z.; Li, L.; Lodish, H.F.; Bartel, D.P. MicroRNAs Modulate Hematopoietic Lineage Differentiation. Science 2004, 303, 83–86. [Google Scholar] [CrossRef] [Green Version]
- Silvestris, N.; Danza, K.; Longo, V.; Brunetti, O.; Fucci, L.; Argentiero, A.; Calabrese, A.; Cataldo, I.; Tamma, R.; Ribatti, D.; et al. Angiogenesis in adenosquamous cancer of pancreas. Oncotarget 2017, 8, 95773–95779. [Google Scholar] [CrossRef] [Green Version]
- Suárez, Y.; Fernández-Hernando, C.; Yu, J.; Gerber, S.A.; Harrison, K.D.; Pober, J.S.; Iruela-Arispe, M.L.; Merkenschlager, M.; Sessa, W.C. Dicer-dependent endothelial microRNAs are necessary for postnatal angiogenesis. Proc. Natl. Acad. Sci. USA 2008, 105, 14082–14087. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.; Min, H.; Yue, S.; Chen, C.-Z. Pre-miRNA Loop Nucleotides Control the Distinct Activities of mir-181a-1 and mir-181c in Early T Cell Development. PLoS ONE 2008, 3, e3592. [Google Scholar] [CrossRef] [Green Version]
- Ji, J.; Yamashita, T.; Budhu, A.; Forgues, M.; Jia, H.-L.; Li, C.; Deng, C.; Wauthier, E.; Reid, L.M.; Ye, Q.-H.; et al. Identification of microRNA-181 by genome-wide screening as a critical player in EpCAM-positive hepatic cancer stem cells. Hepatology 2009, 50, 472–480. [Google Scholar] [CrossRef] [Green Version]
- Ji, J.; Yamashita, T.; Wang, X.W. Wnt/beta-catenin signaling activates microRNA-181 expression in hepatocellular carcinoma. Cell Biosci. 2011, 1, 4. [Google Scholar] [CrossRef] [Green Version]
- Braicu, C.; Gulei, D.; Raduly, L.; Harangus, A.; Rusu, A.; Berindan-Neagoe, I. Altered expression of miR-181 affects cell fate and targets drug resistance-related mechanisms. Mol. Asp. Med. 2019, 70, 90–105. [Google Scholar] [CrossRef]
- Indrieri, A.; Carrella, S.; Carotenuto, P.; Banfi, S.; Franco, B. The Pervasive Role of the miR-181 Family in Development, Neurodegeneration, and Cancer. Int. J. Mol. Sci. 2020, 21, 2092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pop-Bica, C.; Pintea, S.; Cojocneanu-Petric, R.; Del Sal, G.; Piazza, S.; Wu, Z.-H.; Alencar, A.J.; Lossos, I.S.; Berindan-Neagoe, I.; Calin, G.A. MiR-181 family-specific behavior in different cancers: A meta-analysis view. Cancer Metastasis Rev. 2018, 37, 17–32. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Yang, Q.; Kong, W.-Q.; Liu, T.; Liu, M.; Li, X.; Tang, H. MicroRNA-181b targets cAMP responsive element binding protein 1 in gastric adenocarcinomas. IUBMB Life 2012, 64, 628–635. [Google Scholar] [CrossRef] [PubMed]
- Kazenwadel, J.; Michael, M.Z.; Harvey, N. Prox1 expression is negatively regulated by miR-181 in endothelial cells. Blood 2010, 116, 2395–2401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaik, S.; Martin, E.; Hayes, D.J.; Gimble, J.; Devireddy, R. microRNA Sequencing of CD34+ Sorted Adipose Stem Cells Undergoing Endotheliogenesis. Stem Cells Dev. 2021, 30, 265–288. [Google Scholar] [CrossRef]
- Kane, N.M.; Meloni, M.; Spencer, H.L.; Craig, M.A.; Strehl, R.; Milligan, G.; Houslay, M.D.; Mountford, J.C.; Emanueli, C.; Baker, A.H. Derivation of Endothelial Cells From Human Embryonic Stem Cells by Directed Differentiation. Arter. Thromb. Vasc. Biol. 2010, 30, 1389–1397. [Google Scholar] [CrossRef] [Green Version]
- Kane, N.M.; Howard, L.; Descamps, B.; Meloni, M.; McClure, J.; Lu, R.; McCahill, A.; Breen, C.; Mackenzie, R.M.; Delles, C.; et al. Role of MicroRNAs 99b, 181a, and 181b in the Differentiation of Human Embryonic Stem Cells to Vascular Endothelial Cells. Stem Cells 2012, 30, 643–654. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.; Yao, Y.; Wang, P.; Liu, Y.; Zhao, L.; Li, Z.; Li, Z.; Xue, Y. MiR-181a Regulates Blood-Tumor Barrier Permeability by Targeting Krüppel-Like Factor 6. J. Cereb. Blood Flow Metab. 2014, 34, 1826–1836. [Google Scholar] [CrossRef]
- Tominaga, N.; Kosaka, N.; Ono, M.; Katsuda, T.; Yoshioka, Y.; Tamura, K.; Lötvall, J.; Nakagama, H.; Ochiya, T. Brain metastatic cancer cells release microRNA-181c-containing extracellular vesicles capable of destructing blood–brain barrier. Nat. Commun. 2015, 6, 6716. [Google Scholar] [CrossRef] [Green Version]
- Yu, D.; Sun, R.; Shen, D.; Ge, L.; Xue, T.; Cao, Y. Nuclear heme oxygenase-1 improved the hypoxia-mediated dysfunction of blood-spinal cord barrier via the miR-181c-5p/SOX5 signaling pathway. NeuroReport 2021, 32, 112–120. [Google Scholar] [CrossRef]
- Guo, J.; Cai, H.; Zheng, J.; Liu, X.; Liu, Y.; Ma, J.; Que, Z.; Gong, W.; Gao, Y.; Tao, W.; et al. Long non-coding RNA NEAT1 regulates permeability of the blood-tumor barrier via miR-181d-5p-mediated expression changes in ZO-1, occludin, and claudin-5. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2017, 1863, 2240–2254. [Google Scholar] [CrossRef] [PubMed]
- Engelhardt, B.; Sorokin, L. The blood–brain and the blood–cerebrospinal fluid barriers: Function and dysfunction. Semin. Immunopathol. 2009, 31, 497–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czupalla, C.J.; Liebner, S.; Devraj, K. In Vitro Models of the Blood–Brain Barrier. Methods Mol. Biol. 2014, 1135, 415–437. [Google Scholar] [CrossRef] [PubMed]
- Furuse, M. Molecular Basis of the Core Structure of Tight Junctions. Cold Spring Harb. Perspect. Biol. 2010, 2, a002907. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, K.; Skare, I.; Stankewich, M.; Furuse, M.; Tsukita, S.; Rogers, R.; Lynch, R.; Schneeberger, E. Occludin is a functional component of the tight junction. J. Cell Sci. 1996, 109, 2287–2298. [Google Scholar] [CrossRef]
- Daneman, R.; Prat, A. The Blood–Brain Barrier. Cold Spring Harb. Perspect. Biol. 2015, 7, a020412. [Google Scholar] [CrossRef] [Green Version]
- Daneman, R.; Agalliu, D.; Zhou, L.; Kuhnert, F.; Kuo, C.J.; Barres, B.A. Wnt/β-catenin signaling is required for CNS, but not non-CNS, angiogenesis. Proc. Natl. Acad. Sci. USA 2009, 106, 641–646. [Google Scholar] [CrossRef] [Green Version]
- Laksitorini, M.; Yathindranath, V.; Xiong, W.; Hombach-Klonisch, S.; Miller, D.W. Modulation of Wnt/β-catenin signaling promotes blood-brain barrier phenotype in cultured brain endothelial cells. Sci. Rep. 2019, 9, 19718. [Google Scholar] [CrossRef] [Green Version]
- Abbott, N.J.; Patabendige, A.A.K.; Dolman, D.E.M.; Yusof, S.R.; Begley, D.J. Structure and function of the blood-brain barrier. Neurobiol. Dis. 2010, 37, 13–25. [Google Scholar] [CrossRef]
- Paolinelli, R.; Corada, M.; Orsenigo, F.; Dejana, E. The molecular basis of the blood brain barrier differentiation and maintenance. Is it still a mystery? Pharmacol. Res. 2011, 63, 165–171. [Google Scholar] [CrossRef]
- Arvanitis, C.D.; Ferraro, G.B.; Jain, R.K. The blood–brain barrier and blood–tumour barrier in brain tumours and metastases. Nat. Cancer 2020, 20, 26–41. [Google Scholar] [CrossRef] [PubMed]
- Orr, F.W.; Wang, H.H.; Lafrenie, R.; Scherbarth, S.; Nance, D.M. Interactions between cancer cells and the endothelium in metastasis. J. Pathol. 2000, 190, 310–329. [Google Scholar] [CrossRef]
- Das, A.; Fernandez-Zapico, M.E.; Cao, S.; Yao, J.; Fiorucci, S.; Hebbel, R.P.; Urrutia, R.; Shah, V.H. Disruption of an SP2/KLF6 Repression Complex by SHP Is Required for Farnesoid X Receptor-induced Endothelial Cell Migration. J. Biol. Chem. 2006, 281, 39105–39113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ueda, R.; Yoshida, K.; Kawase, T.; Kawakami, Y.; Toda, M. Preferential expression and frequent IgG responses of a tumor antigen, SOX5, in glioma patients. Int. J. Cancer 2007, 120, 1704–1711. [Google Scholar] [CrossRef] [PubMed]
- Suto, A.; Tanaka, S.; Nakajima, H. Sox5 and Th17 cell differentiation. Oncotarget 2015, 6, 19952–19953. [Google Scholar] [CrossRef]
- Zhu, A.; Chu, L.; Ma, Q.; Li, Y. RETRACTED ARTICLE: Long non-coding RNA H19 down-regulates miR-181a to facilitate endothelial angiogenic function. Artif. Cells Nanomed. Biotechnol. 2019, 47, 2698–2705. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.; Sun, L.; Hua, Y.; Yang, C.; Teng, Y. Overexpression of long non-coding RNA H19 protects lung fibroblasts from LPS-induced injury by targeting miR-181a and Runx2 via activation of Notch and JNK pathways. J. Cell Biochem. 2018, 120, 12045–12056. [Google Scholar] [CrossRef]
- Sun, T.; Yin, L.; Kuang, H. miR-181a/b-5p regulates human umbilical vein endothelial cell angiogenesis by targeting PDGFRA. Cell Biochem. Funct. 2020, 38, 222–230. [Google Scholar] [CrossRef]
- Li, Y.; Fan, S.; Xia, W.; Qiao, B.; Huang, K.; Zhou, J.; Liang, M. MiR-181b suppresses angiogenesis by directly targeting cellular communication network factor 1. Lab. Investig. 2021, 101, 1026–1035. [Google Scholar] [CrossRef]
- Cui, Y.; Han, Z.; Hu, Y.; Song, G.; Hao, C.; Xia, H.; Ma, X. MicroRNA-181b and microRNA-9 mediate arsenic-induced angiogenesis via NRP1. J. Cell. Physiol. 2012, 227, 772–783. [Google Scholar] [CrossRef]
- Geretti, E.; Shimizu, A.; Klagsbrun, M. Neuropilin structure governs VEGF and semaphorin binding and regulates angiogenesis. Angiogenesis 2008, 11, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Staton, C.A.; Kumar, I.; Reed, M.; Brown, N.J. Neuropilins in physiological and pathological angiogenesis. J. Pathol. 2007, 212, 237–248. [Google Scholar] [CrossRef] [PubMed]
- Moran, E.P.; Wang, Z.; Chen, J.; Sapieha, P.; Smith, L.E.H.; Ma, J.-X. Neurovascular cross talk in diabetic retinopathy: Pathophysiological roles and therapeutic implications. Am. J. Physiol. Circ. Physiol. 2016, 311, H738–H749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dogra, M.R.; Katoch, D.; Dogra, M. An Update on Retinopathy of Prematurity (ROP). Indian J. Pediatr. 2017, 84, 930–936. [Google Scholar] [CrossRef]
- Karali, M.; Peluso, I.; Marigo, V.; Banfi, S. Identification and Characterization of MicroRNAs Expressed in the Mouse Eye. Investig. Opthalmol. Vis. Sci. 2007, 48, 509–515. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; Tahiri, H.; Cai, C.; Gu, M.; Gagnon, C.; Hardy, P. microRNA-181a inhibits ocular neovascularization by interfering with vascular endothelial growth factor expression. Cardiovasc. Ther. 2018, 36, e12329. [Google Scholar] [CrossRef]
- He, Q.; Zhou, X.; Li, S.; Jin, Y.; Chen, Z.; Chen, D.; Cai, Y.; Liu, Z.; Zhao, T.; Wang, A. MicroRNA-181a suppresses salivary adenoid cystic carcinoma metastasis by targeting MAPK–Snai2 pathway. Biochim. Biophys. Acta (BBA) Gen. Subj. 2013, 1830, 5258–5266. [Google Scholar] [CrossRef]
- Cuevas, A.; Saavedra, N.; Cavalcante, M.F.; Salazar, L.A.; Abdalla, D.S. Identification of microRNAs involved in the modulation of pro-angiogenic factors in atherosclerosis by a polyphenol-rich extract from propolis. Arch. Biochem. Biophys. 2014, 557, 28–35. [Google Scholar] [CrossRef]
- Ouyang, Y.-B.; Lu, Y.; Yue, S.; Giffard, R.G. miR-181 targets multiple Bcl-2 family members and influences apoptosis and mitochondrial function in astrocytes. Mitochondrion 2012, 12, 213–219. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Yao, Y.; Yuan, F.; Xie, B. Overexpression of miR-181a-5p inhibits retinal neovascularization through endocan and the ERK1/2 signaling pathway. J. Cell. Physiol. 2020, 235, 9323–9335. [Google Scholar] [CrossRef]
- Rocha, S.F.; Schiller, M.; Jing, D.; Li, H.; Butz, S.; Vestweber, D.; Biljes, D.; Drexler, H.C.; Nieminen-Kelhä, M.; Vajkoczy, P.; et al. Esm1 Modulates Endothelial Tip Cell Behavior and Vascular Permeability by Enhancing VEGF Bioavailability. Circ. Res. 2014, 115, 581–590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Del Toro, R.; Prahst, C.; Mathivet, T.; Siegfried, G.; Kaminker, J.S.; Larrivee, B.; Breant, C.; Duarte, A.; Takakura, N.; Fukamizu, A.; et al. Identification and functional analysis of endothelial tip cell–enriched genes. Blood 2010, 116, 4025–4033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, T.; Zhong, Y.; Demetriades, A.M.; Shen, J.; Sui, A.; Yao, Y.; Gao, Y.; Zhu, Y.; Shen, X.; Xie, B. Endocan Blockade Suppresses Experimental Ocular Neovascularization in Mice. Investig. Opthalmol. Vis. Sci. 2018, 59, 930–939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.; Wang, Y.; Meng, C.; Fang, N. FMRP regulates endothelial cell proliferation and angiogenesis via the miR-181a-CaM-CaMKII pathway. Cell Biol. Int. 2018, 42, 1432–1444. [Google Scholar] [CrossRef] [PubMed]
- Wan, R.-P.; Zhou, L.-T.; Yang, H.-X.; Zhou, Y.-T.; Ye, S.-H.; Zhao, Q.-H.; Gao, M.-M.; Liao, W.-P.; Yi, Y.-H.; Long, Y.-S. Involvement of FMRP in Primary MicroRNA Processing via Enhancing Drosha Translation. Mol. Neurobiol. 2017, 54, 2585–2594. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Li, B.; Dong, Q.; Qian, C.; Cheng, J.; Wang, Y. Repetitive Transient Ischemia-Induced Cardiac Angiogenesis is Mediated by Camkii Activation. Cell. Physiol. Biochem. 2018, 47, 914–924. [Google Scholar] [CrossRef]
- Xiong, Y.; Chang, L.-L.; Tran, B.; Dai, T.; Zhong, R.; Mao, Y.-C.; Zhu, Y.-Z. ZYZ-803, a novel hydrogen sulfide-nitric oxide conjugated donor, promotes angiogenesis via cross-talk between STAT3 and CaMKII. Acta Pharmacol. Sin. 2020, 41, 218–228. [Google Scholar] [CrossRef]
- Yang, Y.; Cai, Y.; Zhang, Y.; Liu, J.; Xu, Z. Exosomes Secreted by Adipose-Derived Stem Cells Contribute to Angiogenesis of Brain Microvascular Endothelial Cells Following Oxygen–Glucose Deprivation In Vitro Through MicroRNA-181b/TRPM7 Axis. J. Mol. Neurosci. 2018, 65, 74–83. [Google Scholar] [CrossRef]
- Wang, B.; Hsu, S.-H.; Majumder, S.; Kutay, H.; Huang, W.; Jacob, S.T.; Ghoshal, K. TGFβ-mediated upregulation of hepatic miR-181b promotes hepatocarcinogenesis by targeting TIMP3. Oncogene 2010, 29, 1787–1797. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Z.; Inoue, K.; Sun, H.; Leng, T.; Feng, X.; Zhu, L.; Xiong, Z.-G. TRPM7 regulates vascular endothelial cell adhesion and tube formation. Am. J. Physiol. Cell Physiol. 2015, 308, C308–C318. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Rong, P.-Z.; Song, M.-S.; Shi, Z.-W.; Feng, G.; Chen, X.-J.; Shi, L.; Wang, C.-H.; Pang, Q.-J. lncRNA SNHG1 induced by SP1 regulates bone remodeling and angiogenesis via sponging miR-181c-5p and modulating SFRP1/Wnt signaling pathway. Mol. Med. 2021, 27, 141. [Google Scholar] [CrossRef] [PubMed]
- Häusler, K.D.; Horwood, N.J.; Chuman, Y.; Fisher, J.L.; Ellis, J.; Martin, T.J.; Rubin, J.S.; Gillespie, M.T. Secreted Frizzled-Related Protein-1 Inhibits RANKL-Dependent Osteoclast Formation. J. Bone Miner. Res. 2004, 19, 1873–1881. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Sun, Y.; Liu, X.; Zhu, Y.; Bao, B.; Gao, T.; Chai, Y.; Xu, J.; Zheng, X. EGFL6 regulates angiogenesis and osteogenesis in distraction osteogenesis via Wnt/β-catenin signaling. Stem Cell Res. Ther. 2021, 12, 415. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Li, S.; Li, S.; Yu, C.; Huang, D.; Chen, H.; Yin, X. CircPDE4B inhibits retinal pathological angiogenesis via promoting degradation of HIF -1α though targeting miR-181c. IUBMB Life 2020, 72, 1920–1929. [Google Scholar] [CrossRef]
- Sun, L.-F.; Zhang, B.; Chen, X.-J.; Wang, X.-Y.; Zhang, B.-W.; Ji, Y.-Y.; Wu, K.-C.; Wu, J.-Y.; Jin, Z.-B. Circular RNAs in human and vertebrate neural retinas. RNA Biol. 2019, 16, 821–829. [Google Scholar] [CrossRef]
- Schönenberger, D.; Rajski, M.; Harlander, S.; Frew, I.J. Vhl deletion in renal epithelia causes HIF-1α-dependent, HIF-2α-independent angiogenesis and constitutive diuresis. Oncotarget 2016, 7, 60971–60985. [Google Scholar] [CrossRef] [Green Version]
- Song, G.; Yu, M.; Mi, D.; Jiang, Y.; Tian, G.; Geng, C.; Yuan, J.; Miao, S.; Wu, X. Lower growth arrest-specific 5 level in endometrium is related to endometriosis via promoting cell proliferation and angiogenesis. Kaohsiung J. Med Sci. 2021, 37, 776–783. [Google Scholar] [CrossRef]
- Taylor, R.N.; Yu, J.; Torres, P.B.; Schickedanz, A.C.; Park, J.K.; Mueller, M.D.; Sidell, N. Mechanistic and Therapeutic Implications of Angiogenesis in Endometriosis. Reprod. Sci. 2009, 16, 140–146. [Google Scholar] [CrossRef] [Green Version]
- Takawale, A.; Zhang, P.; Azad, A.; Wang, W.; Wang, X.; Murray, A.G.; Kassiri, Z. Myocardial overexpression of TIMP3 after myocardial infarction exerts beneficial effects by promoting angiogenesis and suppressing early proteolysis. Am. J. Physiol. Circ. Physiol. 2017, 313, H224–H236. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Wang, X.; Yao, J.; Qiu, Z. Long non-coding RNA NEAT1 inhibits oxidative stress-induced vascular endothelial cell injury by activating the miR-181d-5p/CDKN3 axis. Artif. Cells Nanomed. Biotechnol. 2019, 47, 3129–3137. [Google Scholar] [CrossRef] [Green Version]
- Teleanu, R.I.; Chircov, C.; Grumezescu, A.M.; Teleanu, D.M. Tumor Angiogenesis and Anti-Angiogenic Strategies for Cancer Treatment. J. Clin. Med. 2019, 9, 84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiarelli, C.; Kozarekar, P.; Adler, H.L.; Cao, J. Membrane type 1-matrix metalloproteinase promotes human prostate cancer invasion and metastasis. Thromb. Haemost. 2005, 93, 770–778. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.G.; Davies, G.; Martin, T.A.; Parr, C.; Watkins, G.; Mason, M.D.; Mansel, R.E. Expression of membrane type-1 matrix metalloproteinase, MT1-MMP in human breast cancer and its impact on invasiveness of breast cancer cells. Int. J. Mol. Med. 2006, 17, 583–590. [Google Scholar] [CrossRef]
- Li, Y.; Kuscu, C.; Banach, A.; Zhang, Q.; Pulkoski-Gross, A.; Kim, D.; Liu, J.; Roth, E.; Li, E.; Shroyer, K.R.; et al. miR-181a-5p Inhibits Cancer Cell Migration and Angiogenesis via Downregulation of Matrix Metalloproteinase-14. Cancer Res. 2015, 75, 2674–2685. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.-Y.; Liu, X.; Wang, S.-L.; Qin, C.-Y. Over-Expression of the Endocan Gene in Endothelial Cells from Hepatocellular Carcinoma is Associated with Angiogenesis and Tumour Invasion. J. Int. Med Res. 2010, 38, 498–510. [Google Scholar] [CrossRef] [Green Version]
- Roudnicky, F.; Poyet, C.; Wild, P.; Krampitz, S.; Negrini, F.; Huggenberger, R.; Rogler, A.; Stöhr, R.; Hartmann, A.; Provenzano, M.; et al. Endocan Is Upregulated on Tumor Vessels in Invasive Bladder Cancer Where It Mediates VEGF-A–Induced Angiogenesis. Cancer Res. 2013, 73, 1097–1106. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Wei, L.; Chen, Q.; Terek, R.M. MicroRNA Regulates Vascular Endothelial Growth Factor Expression in Chondrosarcoma Cells. Clin. Orthop. Relat. Res. 2015, 473, 907–913. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Cen, A.; Yang, Y.; Ye, H.; Li, J.; Liu, S.; Zhao, L. miR-181a, delivered by hypoxic PTC-secreted exosomes, inhibits DACT2 by downregulating MLL3, leading to YAP-VEGF-mediated angiogenesis. Mol. Ther. Nucleic Acids 2021, 24, 610–621. [Google Scholar] [CrossRef]
- Zhang, X.; Yang, Y.; Liu, X.; Herman, J.G.; Brock, M.V.; Licchesi, J.D.; Yue, W.; Pei, X.; Guo, M. Epigenetic regulation of the Wnt signaling inhibitorDACT2in human hepatocellular carcinoma. Epigenetics 2013, 8, 373–382. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.; Herman, J.G.; Brock, M.V.; Sheng, J.; Zhang, M.; Liu, B.; Guo, M. Methylation of DACT2 Promotes Papillary Thyroid Cancer Metastasis by Activating Wnt Signaling. PLoS ONE 2014, 9, e112336. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Wang, X.; Li, J.; You, C.; Lu, P.; Feng, H.; Kong, Y.; Zhang, H.; Liu, Y.; Jiao, R.; et al. MicroRNA-181a promotes angiogenesis in colorectal cancer by targeting SRCIN1 to promote the SRC/VEGF signaling pathway. Cell Death Dis. 2018, 9, 439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irby, R.B.; Yeatman, T.J. Role of Src expression and activation in human cancer. Oncogene 2000, 19, 5636–5642. [Google Scholar] [CrossRef] [Green Version]
- Di Stefano, P.; Damiano, L.; Cabodi, S.; Aramu, S.; Tordella, L.; Praduroux, A.; Piva, R.; Cavallo, F.; Forni, G.; Silengo, L.; et al. p140Cap protein suppresses tumour cell properties, regulating Csk and Src kinase activity. EMBO J. 2007, 26, 2843–2855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.; Charbonneau, C.; Wei, L.; Chen, Q.; Terek, R.M. miR-181a Targets RGS16 to Promote Chondrosarcoma Growth, Angiogenesis, and Metastasis. Mol. Cancer Res. 2015, 13, 1347–1357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berthebaud, M.; Rivière, C.; Jarrier, P.; Foudi, A.; Zhang, Y.; Compagno, D.; Galy, A.; Vainchenker, W.; Louache, F. RGS16 is a negative regulator of SDF-1–CXCR4 signaling in megakaryocytes. Blood 2005, 106, 2962–2968. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Charbonneau, C.; Wei, L.; Yang, W.; Chen, Q.; Terek, R.M. CXCR4-Targeted Therapy Inhibits VEGF Expression and Chondrosarcoma Angiogenesis and Metastasis. Mol. Cancer Ther. 2013, 12, 1163–1170. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Wei, L.; Chen, Q.; Terek, R.M. CXCR4/SDF1 mediate hypoxia induced chondrosarcoma cell invasion through ERK signaling and increased MMP1 expression. Mol. Cancer 2010, 9, 17. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Lu, J.; Chen, L.; Bian, H.; Hu, J.; Li, D.; Xia, C.; Xu, H. Tumor-Derived EV-Encapsulated miR-181b-5p Induces Angiogenesis to Foster Tumorigenesis and Metastasis of ESCC. Mol. Ther. Nucleic Acids 2020, 20, 421–437. [Google Scholar] [CrossRef]
- Xu, X.; Ge, S.; Jia, R.; Zhou, Y.; Song, X.; Zhang, H.; Fan, X. Hypoxia-induced miR-181b enhances angiogenesis of retinoblastoma cells by targeting PDCD10 and GATA6. Oncol. Rep. 2015, 33, 2789–2796. [Google Scholar] [CrossRef] [Green Version]
- Lopatina, T.; Grange, C.; Fonsato, V.; Tapparo, M.; Brossa, A.; Fallo, S.; Pitino, A.; Herrera-Sanchez, M.B.; Kholia, S.; Camussi, G.; et al. Extracellular vesicles from human liver stem cells inhibit tumor angiogenesis. Int. J. Cancer 2018, 144, 322–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Green, C.; Clarke, J.; Bicknell, R.; Turner, A. Pulmonary MicroRNA Changes Alter Angiogenesis in Chronic Obstructive Pulmonary Disease and Lung Cancer. Biomedicines 2021, 9, 830. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Xu, X.; Jia, R.; Song, X.; Wang, J.; Qian, G.; Ge, S.; Fan, X. Microarray-based analysis: Identification of hypoxia-regulated microRNAs in retinoblastoma cells. Int. J. Oncol. 2011, 38, 1385–1393. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Zhang, H.; Yu, L.; Gunel, M.; Boggon, T.J.; Chen, H.; Min, W. Stabilization of VEGFR2 Signaling by Cerebral Cavernous Malformation 3 Is Critical for Vascular Development. Sci. Signal. 2010, 3, ra26. [Google Scholar] [CrossRef] [Green Version]
- You, C.; Sandalcioglu, I.E.; Dammann, P.; Felbor, U.; Sure, U.; Zhu, Y. Loss of CCM3 impairs DLL4-Notch signalling: Implication in endothelial angiogenesis and in inherited cerebral cavernous malformations. J. Cell. Mol. Med. 2013, 17, 407–418. [Google Scholar] [CrossRef]
- Perlman, H.; Suzuki, E.; Simonson, M.; Smith, R.C.; Walsh, K. GATA-6 Induces p21Cip1 Expression and G1Cell Cycle Arrest. J. Biol. Chem. 1998, 273, 13713–13718. [Google Scholar] [CrossRef] [Green Version]
- Raju, R.; Palapetta, S.M.; Sandhya, V.K.; Sahu, A.; Alipoor, A.; Balakrishnan, L.; Advani, J.; George, B.; Kini, K.R.; Geetha, N.P.; et al. A Network Map of FGF-1/FGFR Signaling System. J. Signal Transduct. 2014, 2014, 962962. [Google Scholar] [CrossRef] [Green Version]
- Mori, S.; Tran, V.; Nishikawa, K.; Kaneda, T.; Hamada, Y.; Kawaguchi, N.; Fujita, M.; Takada, Y.K.; Matsuura, N.; Zhao, M.; et al. A Dominant-Negative FGF1 Mutant (the R50E Mutant) Suppresses Tumorigenesis and Angiogenesis. PLoS ONE 2013, 8, e57927. [Google Scholar] [CrossRef] [Green Version]
- Tang, L.; Han, X. The urokinase plasminogen activator system in breast cancer invasion and metastasis. Biomed. Pharmacother. 2013, 67, 179–182. [Google Scholar] [CrossRef]
- Dörks, M.; Jobski, K.; Herget-Rosenthal, S.; Hoffmann, F.; Douros, A. Tyrosine kinase inhibitors targeting vascular endothelial growth factor and the risk of aortic dissection—A pharmacovigilance analysis. Pharmacol. Res. Perspect. 2021, 9, e00707. [Google Scholar] [CrossRef]
- Abdollahi, A.; Folkman, J. Evading tumor evasion: Current concepts and perspectives of anti-angiogenic cancer therapy. Drug Resist. Updat. 2010, 13, 16–28. [Google Scholar] [CrossRef] [PubMed]
- Lima, J.F.; Cerqueira, L.; Figueiredo, C.; Oliveira, C.; Azevedo, N.F. Anti-miRNA oligonucleotides: A comprehensive guide for design. RNA Biol. 2018, 15, 338–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebert, M.S.; Sharp, P.A. MicroRNA sponges: Progress and possibilities. RNA 2010, 16, 2043–2050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aquino-Jarquin, G. Emerging Role of CRISPR/Cas9 Technology for MicroRNAs Editing in Cancer Research. Cancer Res. 2017, 77, 6812–6817. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; Wang, R.; Hardy, P. Potential of miRNA-Based Nanotherapeutics for Uveal Melanoma. Cancers 2021, 13, 5192. [Google Scholar] [CrossRef]
- Guzman-Villanueva, D.; El-Sherbiny, I.M.; Herrera-Ruiz, D.; Vlassov, A.V.; Smyth, H.D. Formulation Approaches to Short Interfering RNA and MicroRNA: Challenges and Implications. J. Pharm. Sci. 2012, 101, 4046–4066. [Google Scholar] [CrossRef]
- Reda El Sayed, S.; Cristante, J.; Guyon, L.; Denis, J.; Chabre, O.; Cherradi, N. MicroRNA Therapeutics in Cancer: Current Advances and Challenges. Cancers 2021, 13, 2680. [Google Scholar] [CrossRef]
- Fu, Y.; Chen, J.; Huang, Z. Recent progress in microRNA-based delivery systems for the treatment of human disease. ExRNA 2019, 1, 24. [Google Scholar] [CrossRef] [Green Version]
- Kulkarni, J.; Cullis, P.R.; van der Meel, R. Lipid Nanoparticles Enabling Gene Therapies: From Concepts to Clinical Utility. Nucleic Acid Ther. 2018, 28, 146–157. [Google Scholar] [CrossRef] [Green Version]
- Whitehead, K.A.; Langer, R.; Anderson, D.G. Knocking down barriers: Advances in siRNA delivery. Nat. Rev. Drug Discov. 2009, 8, 129–138. [Google Scholar] [CrossRef]
- Cheng, Q.; Wei, T.; Farbiak, L.; Johnson, L.T.; Dilliard, S.A.; Siegwart, D.J. Selective organ targeting (SORT) nanoparticles for tissue-specific mRNA delivery and CRISPR–Cas gene editing. Nat. Nanotechnol. 2020, 15, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Akinc, A.; Maier, M.A.; Manoharan, M.; Fitzgerald, K.; Jayaraman, M.; Barros, S.; Ansell, S.; Du, X.; Hope, M.J.; Madden, T.D.; et al. The Onpattro story and the clinical translation of nanomedicines containing nucleic acid-based drugs. Nat. Nanotechnol. 2019, 14, 1084–1087. [Google Scholar] [CrossRef] [PubMed]
- Telford, B.J.; Yahyanejad, S.; de Gunst, T.; Boer, H.C.D.; Vos, R.M.; Stegink, M.; Bosch, M.T.v.D.; Alemdehy, M.F.; van Pinxteren, L.A.; Schaapveld, R.Q.; et al. Multi-modal effects of 1B3, a novel synthetic miR-193a-3p mimic, support strong potential for therapeutic intervention in oncology. Oncotarget 2021, 12, 422–439. [Google Scholar] [CrossRef] [PubMed]
- Rosenblum, D.; Joshi, N.; Tao, W.; Karp, J.M.; Peer, D. Progress and challenges towards targeted delivery of cancer therapeutics. Nat. Commun. 2018, 9, 1410. [Google Scholar] [CrossRef] [Green Version]
- Du, J.-Z.; Lane, L.A.; Nie, S. Stimuli-responsive nanoparticles for targeting the tumor microenvironment. J. Control. Release 2015, 219, 205–214. [Google Scholar] [CrossRef] [Green Version]
- Betzer, O.; Shilo, M.; Opochinsky, R.; Barnoy, E.; Motiei, M.; Okun, E.; Yadid, G.; Popovtzer, R. The effect of nanoparticle size on the ability to cross the blood–brain barrier: An in vivo study. Nanomedicine 2017, 12, 1533–1546. [Google Scholar] [CrossRef]
- Fröhlich, E. The role of surface charge in cellular uptake and cytotoxicity of medical nanoparticles. Int. J. Nanomed. 2012, 7, 5577–5591. [Google Scholar] [CrossRef] [Green Version]
- van Zandwijk, N.; Pavlakis, N.; Kao, S.C.; Linton, A.; Boyer, M.; Clarke, S.; Huynh, Y.; Chrzanowska, A.; Fulham, M.; Bailey, D.L.; et al. Safety and activity of microRNA-loaded minicells in patients with recurrent malignant pleural mesothelioma: A first-in-man, phase 1, open-label, dose-escalation study. Lancet Oncol. 2017, 18, 1386–1396. [Google Scholar] [CrossRef]
- Vader, P.; Mol, E.A.; Pasterkamp, G.; Schiffelers, R.M. Extracellular vesicles for drug delivery. Adv. Drug Deliv. Rev. 2016, 106, 148–156. [Google Scholar] [CrossRef]
- Katakowski, M.; Buller, B.; Zheng, X.; Lu, Y.; Rogers, T.; Osobamiro, O.; Shu, W.; Jiang, F.; Chopp, M. Exosomes from marrow stromal cells expressing miR-146b inhibit glioma growth. Cancer Lett. 2013, 335, 201–204. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Chen, Y.; Yu, H.; Machan, J.T.; Alladin, A.; Ramirez, J.; Taliano, R.; Hart, J.; Chen, Q.; Terek, R.M. Anti-miRNA Oligonucleotide Therapy for Chondrosarcoma. Mol. Cancer Ther. 2019, 18, 2021–2029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabatabaei, S.N.; Derbali, R.M.; Yang, C.; Superstein, R.; Hamel, P.; Chain, J.L.; Hardy, P. Co-delivery of miR-181a and melphalan by lipid nanoparticles for treatment of seeded retinoblastoma. J. Control. Release 2019, 298, 177–185. [Google Scholar] [CrossRef] [PubMed]
| Function | MiR-181 Members | Upstream Events | Target | Downstream Signaling/Events | Ref |
|---|---|---|---|---|---|
| EC differentiation | |||||
| promotes lymphangiogenesis toward vascular EC development | miR-181a | Prox1 | ERK1/2 pathway | [24] | |
| induces endotheliogenesis in ASCs | miR-181a-5p | VEGF, VWF, CD31 | [25] | ||
| enhances hESCs differentiation to vascular ECs | miR-181a/b | Pecam1, nitric oxide VE-Cadherin | [27] | ||
| BBB/BTB | |||||
| increases BTB permeability | miR-181a | KLF6 | tight junction proteins | [28] | |
| increases BBB permeability | miR-181c | PDPK1 | cofilin phosphorylation | [29] | |
| increases blood-spinal cord barrier (BSCB) permeability | miR-181c-5p | NHO-1 | SOX5 | tight junction proteins | [30] |
| increases BTB permeability | miR-181d-5p | NEAT1 | SOX5 | tight junction proteins | [31] |
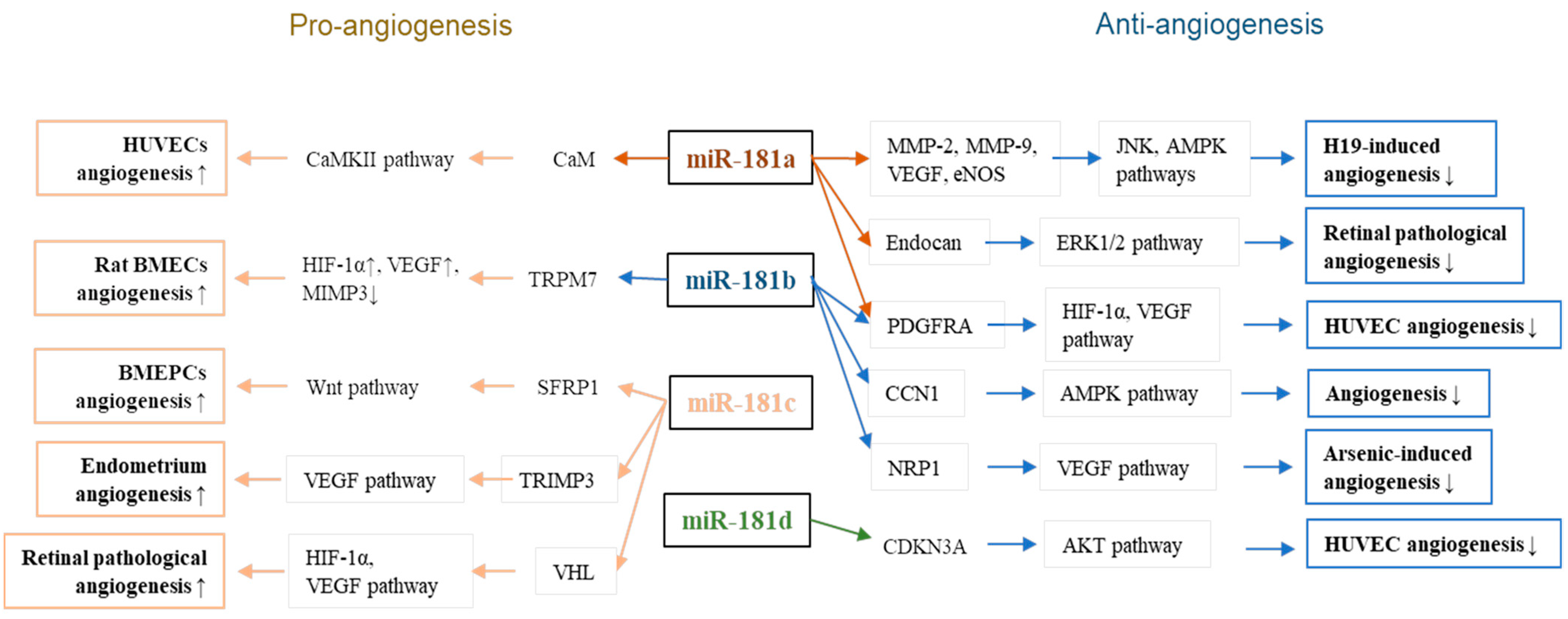
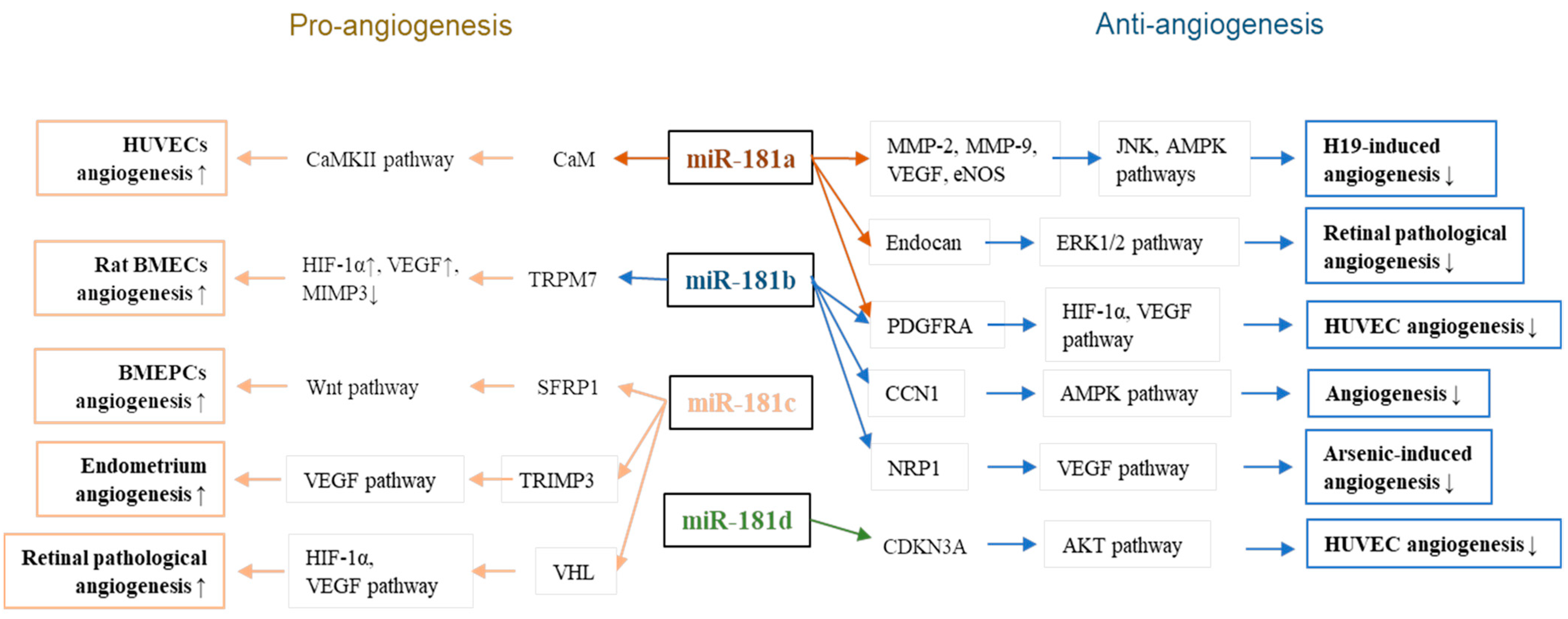
| Function/Effects | MiR-181 | Upstream | Target | Downstream Events/Signaling | Ref |
|---|---|---|---|---|---|
| suppress HMEC-1 angiogenic function | miR-181a | H19 | MMP-2, MMP-9, | VRGF↓, eNOS↓, H19-activated JNK, and AMPK signaling↓ | [46] |
| inhibits ocular neovascularization | miR-181a-5p | Bcl2, MAPK1 | VEGF signaling↓ | [56] | |
| inhibits retinal neovascularization | miR-181a-5p | endocan | ERK signaling↓ | [60] | |
| promotes HUVEC angiogenesis | miR-181a | CaM | CaM-CaMKII pathway↓ | [65] | |
| suppress HUVEC angiogenesis | miR-181a/b-5p | PDGFRA | [48] | ||
| suppresses HUVEC angiogenesis | miR-181b | CCN1 | AMPK signaling↓ | [49] | |
| suppresses arsenic-induced angiogenesis | miR-181b | NRP1 | [50] | ||
| promotes rat BMEC angiogenesis | miR-181b-5p | TRPM7, TIMP3 | HIF-1α↑, VEGF↑, | [68] | |
| promotes BM-EPC angiogenesis | miR-181c-5p | SNHG1 | SFRP1 | Wnt3a/β-catenin signaling↑ | [71] |
| promotes retinal pathological angiogenesis | miR-181c | CircPDE4B | VHL | HIF-1α, VEGF signaling↑ | [74] |
| promotes angiogenesis in the endometrium | miR-181c | GAS5 | TIMP3 | VEGF signaling↑ | [77] |
| suppresses HUVEC angiogenesis | miR-181d-5p | NEAT1 | CDKN3 | Akt signaling ↓ | [80] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, C.; Passos Gibson, V.; Hardy, P. The Role of MiR-181 Family Members in Endothelial Cell Dysfunction and Tumor Angiogenesis. Cells 2022, 11, 1670. https://doi.org/10.3390/cells11101670
Yang C, Passos Gibson V, Hardy P. The Role of MiR-181 Family Members in Endothelial Cell Dysfunction and Tumor Angiogenesis. Cells. 2022; 11(10):1670. https://doi.org/10.3390/cells11101670
Chicago/Turabian StyleYang, Chun, Victor Passos Gibson, and Pierre Hardy. 2022. "The Role of MiR-181 Family Members in Endothelial Cell Dysfunction and Tumor Angiogenesis" Cells 11, no. 10: 1670. https://doi.org/10.3390/cells11101670
APA StyleYang, C., Passos Gibson, V., & Hardy, P. (2022). The Role of MiR-181 Family Members in Endothelial Cell Dysfunction and Tumor Angiogenesis. Cells, 11(10), 1670. https://doi.org/10.3390/cells11101670