
Redox Regulation of Soluble Epoxide Hydrolase—Implications for Cardiovascular Health and Disease


Abstract
:1. Introduction
2. Soluble Epoxide Hydrolase
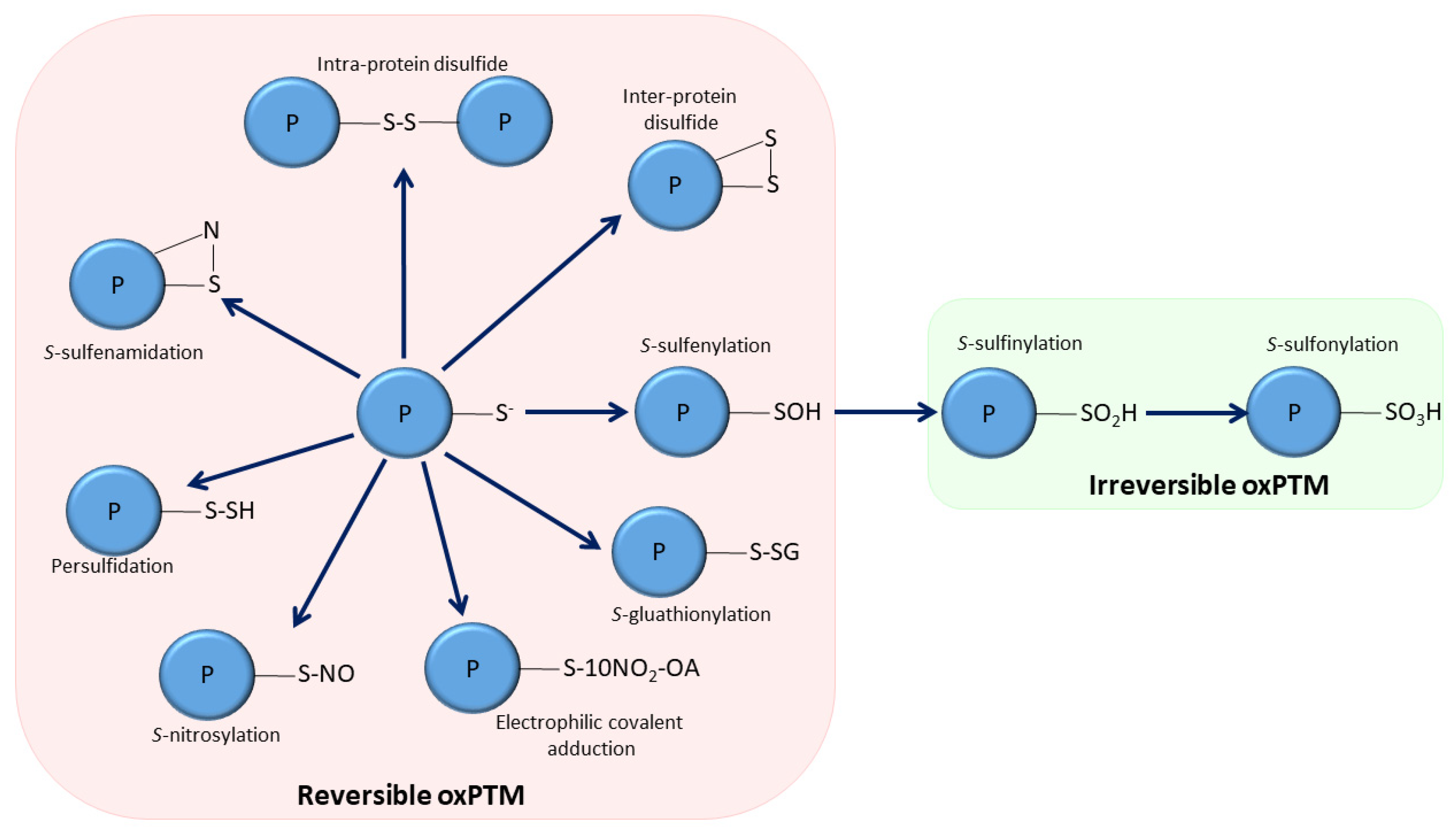
3. Oxidative Post-Translational Modifications of sEH
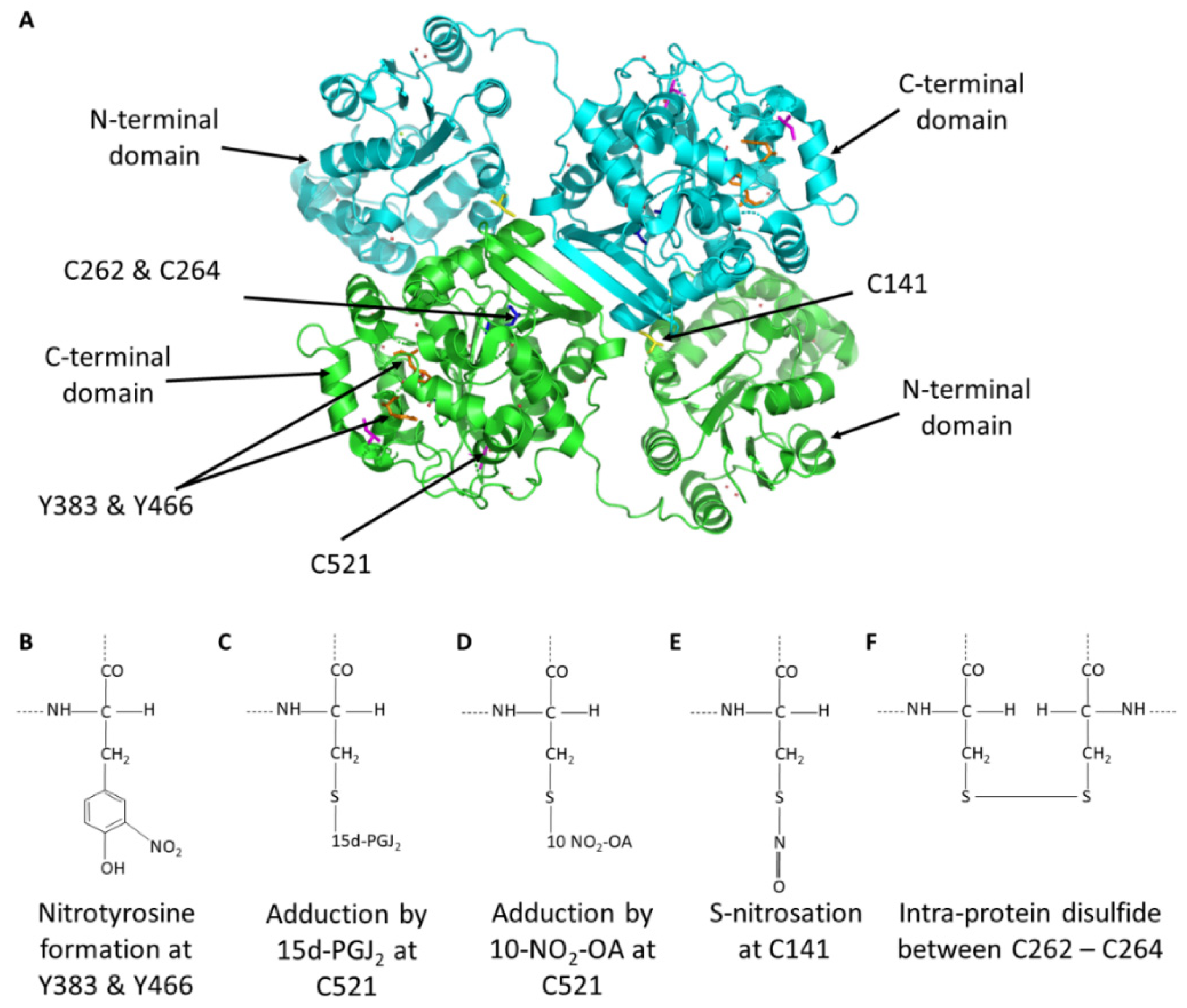
3.1. Tyrosine Nitration
3.2. Adduction of Electrophilic Lipids to sEH
3.3. S-Nitrosation of sEH
3.4. Intra-Disulfide Formation in sEH
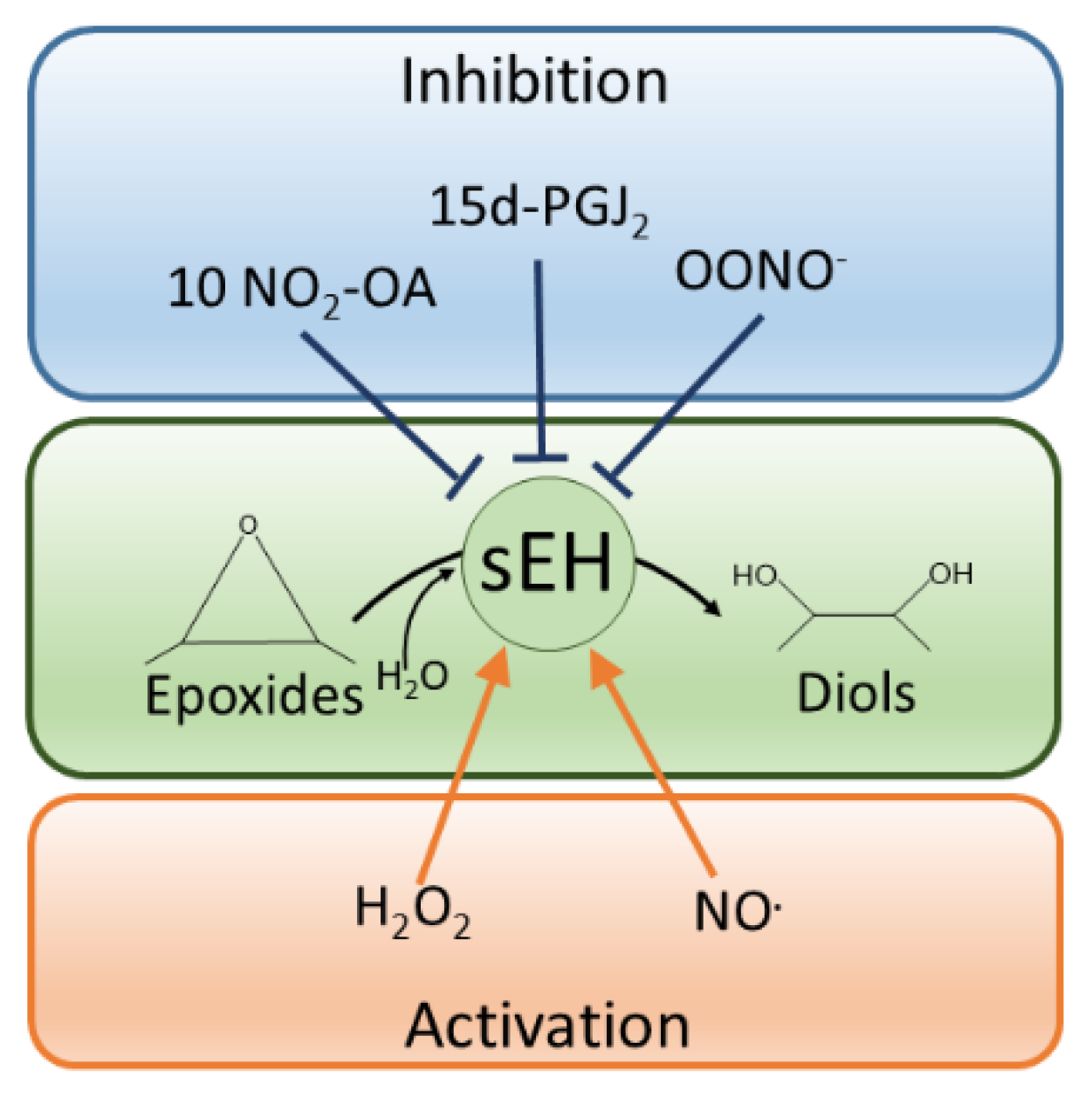
4. Redox Regulation of sEH in Blood Vessels
5. The Role of Redox Regulation of sEH in the Heart
6. Perspectives and Future Directions
7. Closing Thoughts and Considerations
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Davies, M.J. Protein Oxidation and Peroxidation. Biochem. J. 2016, 473, 805–825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radi, R. Protein Tyrosine Nitration: Biochemical Mech anisms and Structural Basis of Functional Effects. Acc. Chem. Res. 2013, 46, 550–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mallozzi, C.; Di Stasi, A.M.M.; Minetti, M. Nitrotyrosine Mimics Phosphotyrosine Binding to the Sh2 Domain of the Src Family Tyrosine Kinase Lyn. FEBS Lett. 2001, 503, 189–195. [Google Scholar] [CrossRef] [Green Version]
- Hoshi, T.; Heinemann, S. Regulation of Cell Function by Methionine Oxidation and Reduction. J. Physiol. 2001, 531 Pt 1, 1–11. [Google Scholar] [CrossRef]
- Reddie, K.G.; Carroll, K.S. Carroll. Expanding the Functional Diversity of Proteins through Cysteine Oxidation. Curr. Opin. Chem. Biol. 2008, 12, 746–754. [Google Scholar] [CrossRef]
- Rudyk, O.; Eaton, P. Biochemical Methods for Monitoring Protein Thiol Redox States in Biological Systems. Redox Biol. 2014, 2, 803–813. [Google Scholar] [CrossRef] [Green Version]
- Bibli, S.I.; Fleming, I. Oxidative Post-Translational Modifications: A Focus on Cysteine S-Sulfhydration and the Regulation of Endothelial Fitness. Antioxid. Redox Signal. 2021, 35, 1494–1514. [Google Scholar] [CrossRef]
- Schopfer, F.J.; Cipollina, C.; Freeman, B.A. Freeman. Formation and Signaling Actions of Electrophilic Lipids. Chem. Rev. 2011, 111, 5997–6021. [Google Scholar] [CrossRef] [Green Version]
- Burgoyne, J.R.; Madhani, M.; Cuello, F.; Charles, R.L.; Brennan, J.P.; Schroder, E.; Browning, D.D.; Eaton, P. Cysteine Redox Sensor in Pkgia Enables Oxidant-Induced Activation. Science 2007, 317, 1393–1397. [Google Scholar] [CrossRef]
- Brennan, J.P.; Bardswell, S.C.; Burgoyne, J.; Fuller, W.; Schröder, E.; Wait, R.; Begum, S.; Kentish, J.C.; Eaton, P. Oxidant-Induced Activation of Type I Protein Kinase a Is Mediated by Ri Subunit Interprotein Disulfide Bond Formation. J. Biol. Chem. 2006, 281, 21827–21836. [Google Scholar] [CrossRef] [Green Version]
- Maller, C.; Schröder, E.; Eaton, P. Glyceraldehyde 3-Phosphate Dehydrogenase Is Unlikely to Mediate Hydrogen Peroxide Signaling: Studies with a Novel Anti-Dimedone Sulfenic Acid Antibody. Antioxid. Redox Signal. 2011, 14, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Salmeen, A.; Andersen, J.N.; Myers, M.P.; Meng, T.-C.; Hinks, J.A.; Tonks, N.K.; Barford, D. Redox Regulation of Protein Tyrosine Phosphatase 1b Involves a Sulphenyl-Amide Intermediate. Nature 2003, 423, 769–773. [Google Scholar] [CrossRef] [PubMed]
- Burgoyne, J.R.; Mongue-Din, H.; Eaton, P.; Shah, A.M. Redox Signaling in Cardiac Physiology and Pathology. Circ. Res. 2012, 111, 1091–1116. [Google Scholar] [CrossRef] [PubMed]
- Charles, R.L.; Eaton, P. Redox Signalling in Cardiovascular Disease. Proteom.-Clin. Appl. 2008, 2, 823–836. [Google Scholar] [CrossRef]
- Benhar, M. Oxidants, Antioxidants and Thiol Redox Switches in the Control of Regulated Cell Death Pathways. Antioxidants 2020, 9, 309. [Google Scholar] [CrossRef] [Green Version]
- Paulsen, C.E.; Carroll, K.S. Cysteine-Mediated Redox Signaling: Chemistry, Biology, and Tools for Discovery. Chem. Rev. 2013, 7, 4633–4679. [Google Scholar] [CrossRef]
- Xiao, H.; Jedrychowski, M.P.; Schweppe, D.K.; Huttlin, E.L.; Yu, Q.; Heppner, D.E.; Li, J.; Long, J.; Mills, E.L.; Szpyt, J.; et al. A Quantitative Tissue-Specific Landscape of Protein Redox Regulation during Aging. Cell 2020, 180, 968–983.e24. [Google Scholar] [CrossRef]
- Revermann, M. Pharmacological inhibition of the soluble epoxide hydrolase—from mouse to man. Curr. Opin. Pharmacol. 2010, 10, 173–178. [Google Scholar] [CrossRef]
- Imig, J.D.; Hammock, B.D. Soluble epoxide hydrolase as a therapeutic target for cardiovascular diseases. Nat. Rev. Drug Discov. 2009, 8, 794–805. [Google Scholar] [CrossRef] [Green Version]
- Chiamvimonvat, N.; Ho, C.-M.; Tsai, H.-J.; Hammock, B.D. The Soluble Epoxide Hydrolase as a Pharmaceutical Target for Hypertension. J. Cardiovasc. Pharmacol. 2007, 50, 225–237. [Google Scholar] [CrossRef] [Green Version]
- Marino, J.P., Jr. Soluble Epoxide Hydrolase, a Target with Multiple Opportunities for Cardiovascular Drug Discovery. Curr. Top. Med. Chem. 2009, 9, 452–463. [Google Scholar] [CrossRef] [PubMed]
- Imig, J.D. Cardiovascular Therapeutic Aspects of Soluble Epoxide Hydrolase Inhibitors. Cardiovasc. Drug Rev. 2006, 24, 169–188. [Google Scholar] [CrossRef] [PubMed]
- Imig, J.D.; Zhao, X.; Zaharis, C.Z.; Olearczyk, J.J.; Pollock, D.M.; Newman, J.; Kim, I.-H.; Watanabe, T.; Hammock, B.D. An Orally Active Epoxide Hydrolase Inhibitor Lowers Blood Pressure and Provides Renal Protection in Salt-Sensitive Hypertension. Hypertension 2005, 46, 975–981. [Google Scholar] [CrossRef] [Green Version]
- Gross, G.J.; Nithipatikom, K. Soluble epoxide hydrolase: A new target for cardioprotection. Curr. Opin. Investig. Drugs 2009, 10, 253–258. [Google Scholar] [PubMed]
- Fang, X. Soluble Epoxide Hydrolase: A Novel Target for the Treatment of Hypertension. Recent Patents Cardiovasc. Drug Discov. 2006, 1, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Monti, J.; Fischer, J.; Paskas, S.; Heinig, M.; Schulz, H.; Gosele, C.; Heuser, A.; Fischer, R.; Schmidt, C.; Schirdewan, A.; et al. Soluble epoxide hydrolase is a susceptibility factor for heart failure in a rat model of human disease. Nat. Genet. 2008, 40, 529–537. [Google Scholar] [CrossRef] [PubMed]
- Davis, B.B.; Morisseau, C.; Newman, J.W.; Pedersen, T.L.; Hammock, B.D.; Weiss, R.H. Attenuation of Vascular Smooth Muscle Cell Proliferation by 1-Cyclohexyl-3-dodecyl Urea Is Independent of Soluble Epoxide Hydrolase Inhibition. J. Pharmacol. Exp. Ther. 2006, 316, 815–821. [Google Scholar] [CrossRef] [Green Version]
- Ulu, A.; Davis, B.B.; Tsai, H.-J.; Kim, I.-H.; Morisseau, C.; Inceoglu, B.; Fiehn, O.; Hammock, B.D.; Weiss, R.H. Soluble Epoxide Hydrolase Inhibitors Reduce the Development of Atherosclerosis in Apolipoprotein E-Knockout Mouse Model. J. Cardiovasc. Pharmacol. 2009, 52, 314–323. [Google Scholar] [CrossRef] [Green Version]
- Olearczyk, J.J.; Quigley, J.E.; Mitchell, B.C.; Yamamoto, T.; Kim, I.-H.; Newman, J.W.; Luria, A.; Hammock, B.D.; Imig, J.D. Administration of a substituted adamantyl urea inhibitor of soluble epoxide hydrolase protects the kidney from damage in hypertensive Goto–Kakizaki rats. Clin. Sci. 2008, 116, 61–70. [Google Scholar] [CrossRef] [Green Version]
- Jung, O.; Brandes, R.P.; Kim, I.-H.; Schweda, F.; Schmidt, R.; Hammock, B.D.; Busse, R.; Fleming, I. Soluble Epoxide Hydrolase Is a Main Effector of Angiotensin II–Induced Hypertension. Hypertension 2005, 45, 759–765. [Google Scholar] [CrossRef] [Green Version]
- Imig, J.D.; Zhao, X.; Capdevila, J.H.; Morisseau, C.; Hammock, B.D. Soluble epoxide hydrolase inhibition lowers arterial blood pressure in angiotensin II hypertension. Hypertension 2002, 39, 690–694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, T.R.; Li, N.; Nipavan, C.; Hammock, B.D. The Potential of Soluble Epoxide Hydrolase Inhibition in the Treatment of Cardiac Hypertrophy. Congest. Hear. Fail. 2008, 14, 219–224. [Google Scholar] [CrossRef] [PubMed]
- Ai, D.; Pang, W.; Li, N.; Xu, M.; Jones, P.D.; Yang, J.; Zhang, Y.; Chiamvimonvat, N.; Shyy, J.Y.-J.; Hammock, B.D.; et al. Soluble epoxide hydrolase plays an essential role in angiotensin II-induced cardiac hypertrophy. Proc. Natl. Acad. Sci. USA 2009, 106, 564–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sirish, P.; Li, N.; Liu, J.-Y.; Lee, K.S.S.; Hwang, S.H.; Qiu, H.; Zhao, C.; Ma, S.M.; López, J.E.; Hammock, B.D.; et al. Unique mechanistic insights into the beneficial effects of soluble epoxide hydrolase inhibitors in the prevention of cardiac fibrosis. Proc. Natl. Acad. Sci. USA 2013, 110, 5618–5623. [Google Scholar] [CrossRef] [Green Version]
- Morisseau, C.; Schebb, N.H.; Dong, H.; Ulu, A.; Aronov, P.A.; Hammock, B.D. Role of soluble epoxide hydrolase phosphatase activity in the metabolism of lysophosphatidic acids. Biochem. Biophys. Res. Commun. 2012, 419, 796–800. [Google Scholar] [CrossRef] [Green Version]
- Oguro, A.; Imaoka, S. Lysophosphatidic acids are new substrates for the phosphatase domain of soluble epoxide hydrolase. J. Lipid Res. 2012, 53, 505–512. [Google Scholar] [CrossRef] [Green Version]
- Arand, M.; Cronin, A.; Adamska, M.; Oesch, F. Epoxide Hydrolases: Structure, Function, Mechanism, and Assay. Methods Enzym. 2005, 400, 569–588. [Google Scholar]
- Newman, J.W.; Morisseau, C.; Hammock, B.D. Epoxide hydrolases: Their roles and interactions with lipid metabolism. Prog. Lipid Res. 2005, 44, 1–51. [Google Scholar] [CrossRef]
- Argiriadi, M.A.; Morisseau, C.; Goodrow, M.H.; Dowdy, D.L.; Hammock, B.D.; Christianson, D.W. Binding of Alkylurea Inhibitors to Epoxide Hydrolase Implicates Active Site Tyrosines in Substrate Activation. J. Biol. Chem. 2000, 275, 15265–15270. [Google Scholar] [CrossRef] [Green Version]
- Gomez, G.A.; Morisseau, C.; Hammock, A.B.D.; Christianson, D.W. Structure of Human Epoxide Hydrolase Reveals Mechanistic Inferences on Bifunctional Catalysis in Epoxide and Phosphate Ester Hydrolysis. Biochemistry 2004, 43, 4716–4723. [Google Scholar] [CrossRef]
- Gomez, G.A.; Morisseau, C.; Hammock, B.D.; Christianson, D.W. Human Soluble Epoxide Hydrolase: Structural Basis of Inhibition by 4-(3-Cyclohexylureido)-Carboxylic Acids. Protein. Sci. 2006, 15, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Campbell, W.B.; Deeter, C.; Gauthier, K.M.; Ingraham, R.H.; Falck, J.R.; Li, P.L. 14,15-Dihydroxyeicosatrienoic Acid Relaxes Bovine Coronary Arteries by Activation of K(Ca) Channels. Am. J. Physiol. Heart Circ. Physiol. 2002, 282, H1656–H1664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, W.B.; Gebremedhin, D.; Pratt, P.F.; Harder, D.R. Identification of Epoxyeicosatrienoic Acids as Endothelium-Derived Hyperpolarizing Factors. Circ. Res. 1996, 78, 415–423. [Google Scholar] [CrossRef] [PubMed]
- Campbell, W.B.; Harder, D.R. Endothelium-derived hyperpolarizing factors and vascular cytochrome P450 metabolites of arachidonic acid in the regulation of tone. Circ. Res. 1999, 84, 484–488. [Google Scholar] [CrossRef] [Green Version]
- Fleming, I. Discreet Regulators of Homeostasis: Epoxyeicosatrienoic Acids, Cytochrome P450 Epoxygenases and Vascular Inflammation. Trends Pharmacol. Sci. 2007, 28, 448–452. [Google Scholar] [CrossRef]
- Luria, A.; Weldon, S.M.; Kabcenell, A.K.; Ingraham, R.H.; Matera, D.; Jiang, H.; Gill, R.; Morisseau, C.; Newman, J.; Hammock, B.D. Compensatory Mechanism for Homeostatic Blood Pressure Regulation in Ephx2 Gene-disrupted Mice. J. Biol. Chem. 2007, 282, 2891–2898. [Google Scholar] [CrossRef] [Green Version]
- Imig, J.D. Epoxides and Soluble Epoxide Hydrolase in Cardiovascular Physiology. Physiol. Rev. 2012, 92, 101–130. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Mäki-Petäjä, K.; Cheriyan, J.; McEniery, C.; Wilkinson, I.B. The role of epoxyeicosatrienoic acids in the cardiovascular system. Br. J. Clin. Pharmacol. 2015, 80, 28–44. [Google Scholar] [CrossRef] [Green Version]
- Node, K.; Ruan, X.-L.; Dai, J.; Yang, S.-X.; Graham, L.; Zeldin, D.C.; Liao, J.K. Activation of Galpha S Mediates Induction of Tissue-Type Plasminogen Activator Gene Transcription by Epoxyeicosatrienoic Acids. J. Biol. Chem. 2001, 276, 15983–15989. [Google Scholar] [CrossRef] [Green Version]
- Inceoglu, B.; Jinks, S.L.; Ulu, A.; Hegedus, C.M.; Georgi, K.; Schmelzer, K.R.; Wagner, K.; Jones, P.D.; Morisseau, C.; Hammock, B.D. Soluble epoxide hydrolase and epoxyeicosatrienoic acids modulate two distinct analgesic pathways. Proc. Natl. Acad. Sci. USA 2008, 105, 18901–18906. [Google Scholar] [CrossRef] [Green Version]
- Imig, J.D. Epoxide hydrolase and epoxygenase metabolites as therapeutic targets for renal diseases. Am. J. Physiol. Physiol. 2005, 289, F496–F503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, T.R.; Hammock, B.D. Soluble Epoxide Hydrolase: Gene Structure, Expression and Deletion. Gene 2013, 526, 61–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbosa-Sicard, E.; Frömel, T.; Keserü, B.; Brandes, R.P.; Morisseau, C.; Hammock, B.D.; Braun, T.; Krüger, M.; Fleming, I. Inhibition of the Soluble Epoxide Hydrolase by Tyrosine Nitration. J. Biol. Chem. 2009, 284, 28156–28163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charles, R.L.; Burgoyne, J.R.; Mayr, M.; Weldon, S.M.; Hubner, N.; Dong, H.; Morisseau, C.; Hammock, B.D.; Landar, A.; Eaton, P. Redox Regulation of Soluble Epoxide Hydrolase by 15-Deoxy-Delta-Prostaglandin J2 Controls Coronary Hypoxic Vasodilation. Circ. Res. 2011, 108, 324–334. [Google Scholar] [CrossRef] [Green Version]
- Shibata, T. 15-Deoxy-Delta(1)(2),(1)(4)-Prostaglandin J(2) as an Electrophilic Mediator. Biosci. Biotechnol. Biochem. 2015, 79, 1044–1049. [Google Scholar] [CrossRef] [Green Version]
- Charles, R.L.; Rudyk, O.; Prysyazhna, O.; Kamynina, A.; Yang, J.; Morisseau, C.; Hammock, B.D.; Freeman, B.A.; Eaton, P. Protection from hypertension in mice by the Mediterranean diet is mediated by nitro fatty acid inhibition of soluble epoxide hydrolase. Proc. Natl. Acad. Sci. USA 2014, 111, 8167–8172. [Google Scholar] [CrossRef] [Green Version]
- Abis, G.; Charles, R.L.; Kopec, J.; Yue, W.W.; Atkinson, R.A.; Bui, T.T.T.; Lynham, S.; Popova, S.; Sun, Y.-B.; Fraternali, F.; et al. 15-deoxy-Δ12,14-Prostaglandin J2 inhibits human soluble epoxide hydrolase by a dual orthosteric and allosteric mechanism. Commun. Biol. 2019, 2, 188. [Google Scholar] [CrossRef]
- Rudolph, V.; Rudolph, T.K.; Schopfer, F.J.; Bonacci, G.; Woodcock, S.R.; Cole, M.P.; Baker, P.R.; Ramani, R.; Freeman, B.A. Endogenous generation and protective effects of nitro-fatty acids in a murine model of focal cardiac ischaemia and reperfusion. Cardiovasc. Res. 2010, 85, 155–166. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Luis, V.; Lin, C.; Fan, Z.; Milton, H.; Zhu, T.; Chen, S.; Chen, M.; Cole, P.; Francisco, J.; et al. Nitro-Oleic Acid Inhibits Angiotensin Ii–Induced Hypertension. Circ. Res. 2010, 107, 540–548. [Google Scholar] [CrossRef] [Green Version]
- Coles, B.; Bloodsworth, A.; Clark, S.R.; Lewis, M.J.; Cross, A.R.; Freeman, B.A.; O’Donnell, V.B. Nitrolinoleate Inhibits Superoxide Generation, Degranulation, and Integrin Expression by Human Neutrophils: Novel Antiinflammatory Properties of Nitric Oxide–Derived Reactive Species in Vascular Cells. Circ. Res. 2002, 91, 375–381. [Google Scholar] [CrossRef] [Green Version]
- Coles, B.; Bloodsworth, A.; Eiserich, J.P.; Coffey, M.J.; McLoughlin, R.; Giddings, J.C.; Lewis, M.J.; Haslam, R.J.; Freeman, B.A.; O’Donnell, V.B. Nitrolinoleate Inhibits Platelet Activation by Attenuating Calcium Mobilization and Inducing Phosphorylation of Vasodilator-stimulated Phosphoprotein through Elevation of cAMP. J. Biol. Chem. 2002, 277, 5832–5840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Covas, M.-I. Olive oil and the cardiovascular system. Pharmacol. Res. 2007, 55, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Toledo, E.; Hu, F.B.; Estruch, R.; Buil-Cosiales, P.; Corella, D.; Salas-Salvad, Ó.J.; Covas, M.; Arós, F.; Gómez-Gracia, E.; Fiol, M.; et al. Effect of the Mediterranean Diet on Blood Pressure in the Predimed Trial: Results from a Randomized Controlled Trial. BMC Med. 2013, 11, 207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Napolitano, A.; Panzella, L.; Savarese, M.; Sacchi, R.; Giudicianni, I.; Paolillo, L.; D’Ischia, M. Acid-Induced Structural Modifications of Unsaturated Fatty Acids and Phenolic Olive Oil Constituents by Nitrite Ions: A Chemical Assessment. Chem. Res. Toxicol. 2004, 17, 1329–1337. [Google Scholar] [CrossRef] [PubMed]
- Villacorta, L.; Chang, L.; Salvatore, S.; Ichikawa, T.; Zhang, J.; Petrovic-Djergovic, D.; Jia, L.; Carlsen, H.; Schopfer, F.J.; Freeman, B.A.; et al. Electrophilic nitro-fatty acids inhibit vascular inflammation by disrupting LPS-dependent TLR4 signalling in lipid rafts. Cardiovasc. Res. 2013, 98, 116–124. [Google Scholar] [CrossRef]
- Weitzberg, E.; Lundberg, J.O. Novel Aspects of Dietary Nitrate and Human Health. Annu. Rev. Nutr. 2013, 33, 129–159. [Google Scholar] [CrossRef]
- Ding, Y.; Li, Y.; Zhang, X.; He, J.; Lu, D.; Fang, X.; Wang, Y.; Wang, J.; Zhang, Y.; Qiao, X.; et al. Soluble epoxide hydrolase activation by S -nitrosation contributes to cardiac ischemia–reperfusion injury. J. Mol. Cell. Cardiol. 2017, 110, 70–79. [Google Scholar] [CrossRef]
- Charles, R.L.; Abis, G.; Fernandez, B.F.; Guttzeit, S.; Buccafusca, R.; Conte, M.R.; Eaton, P. A thiol redox sensor in soluble epoxide hydrolase enables oxidative activation by intra-protein disulfide bond formation. Redox Biol. 2021, 46, 102107. [Google Scholar] [CrossRef]
- Wolhuter, K.; Whitwell, H.J.; Switzer, C.H.; Burgoyne, J.R.; Timms, J.F.; Eaton, P. Evidence against Stable Protein S-Nitrosylation as a Widespread Mechanism of Post-translational Regulation. Mol. Cell 2018, 69, 438–450.e5. [Google Scholar] [CrossRef]
- Heizer, M.L.; McKinney, J.S.; Ellis, E.F. 14,15-Epoxyeicosatrienoic acid inhibits platelet aggregation in mouse cerebral arterioles. Stroke 1991, 22, 1389–1393. [Google Scholar] [CrossRef] [Green Version]
- Fitzpatrick, F.A.; Ennis, M.D.; Baze, M.E.; Wynalda, M.A.; McGee, J.E.; Liggett, W.F. Inhibition of Cyclooxygenase Activity and Platelet Aggregation by Epoxyeicosatrienoic Acids. Influence of Stereochemistry. J. Biol. Chem. 1986, 261, 15334–15338. [Google Scholar] [CrossRef]
- Potente, M.; Fisslthaler, B.; Busse, R.; Fleming, I. 11,12-Epoxyeicosatrienoic Acid-Induced Inhibition of Foxo Factors Promotes Endothelial Proliferation by Down-Regulating P27kip1. J. Biol. Chem. 2003, 278, 29619–29625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potente, M.; Michaelis, U.R.; Fisslthaler, B.; Busse, R.; Fleming, I. Cytochrome P450 2c9-Induced Endothelial Cell Proliferation Involves Induction of Mitogen-Activated Protein (Map) Kinase Phosphatase-1, Inhibition of the C-Jun N-Terminal Kinase, and up-Regulation of Cyclin D1. J. Biol. Chem. 2002, 277, 15671–15676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bir, S.C.; Kolluru, G.K.; Fang, K.; Kevil, C.G. Redox balance dynamically regulates vascular growth and remodeling. Semin. Cell Dev. Biol. 2012, 23, 745–757. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Brewer, A.C.; Schröder, K.; Santos, C.X.C.; Grieve, D.J.; Wang, M.; Anilkumar, N.; Yu, B.; Dong, X.; Walker, S.J.; et al. NADPH oxidase-4 mediates protection against chronic load-induced stress in mouse hearts by enhancing angiogenesis. Proc. Natl. Acad. Sci. USA 2010, 107, 18121–18126. [Google Scholar] [CrossRef] [Green Version]
- Cho, H.J.; Switzer, C.H.; Kamynina, A.; Charles, R.; Rudyk, O.; Ng, T.; Burgoyne, J.R.; Eaton, P. Complex Interrelationships between Nitro-Alkene-Dependent Inhibition of Soluble Epoxide Hydrolase, Inflammation and Tumor Growth. Redox Biol. 2020, 29, 101405. [Google Scholar] [CrossRef]
- Frömel, T.; Jungblut, B.; Hu, J.; Trouvain, C.; Barbosa-Sicard, E.; Popp, R.; Liebner, S.; Dimmeler, S.; Hammock, B.D.; Fleming, I. Soluble epoxide hydrolase regulates hematopoietic progenitor cell function via generation of fatty acid diols. Proc. Natl. Acad. Sci. USA 2012, 109, 9995–10000. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Popp, R.; Fromel, T.; Ehling, M.; Awwad, K.; Adams, R.H.; Hammes, H.P.; Fleming, I. Muller Glia Cells Regulate Notch Signaling and Retinal Angiogenesis Via the Generation of 19,20-Dihydroxydocosapentaenoic Acid. J. Exp. Med. 2014, 211, 281–295. [Google Scholar] [CrossRef] [Green Version]
- Node, K.; Huo, Y.; Ruan, X.; Yang, B.; Spiecker, M.; Ley, K.; Zeldin, D.C.; Liao, J.K. Anti-inflammatory Properties of Cytochrome P450 Epoxygenase-Derived Eicosanoids. Science 1999, 285, 1276–1279. [Google Scholar] [CrossRef] [Green Version]
- Inceoglu, B.; Jinks, S.L.; Schmelzer, K.R.; Waite, T.; Kim, I.H.; Hammock, B.D. Inhibition of soluble epoxide hydrolase reduces LPS-induced thermal hyperalgesia and mechanical allodynia in a rat model of inflammatory pain. Life Sci. 2006, 79, 2311–2319. [Google Scholar] [CrossRef] [Green Version]
- Manhiani, M.; Quigley, J.E.; Knight, S.F.; Tasoobshirazi, S.; Moore, T.; Brands, M.W.; Hammock, B.D.; Imig, J.D. Soluble epoxide hydrolase gene deletion attenuates renal injury and inflammation with DOCA-salt hypertension. Am. J. Physiol. Physiol. 2009, 297, F740–F748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.; Yamamoto, T.; Newman, J.W.; Kim, I.-H.; Watanabe, T.; Hammock, B.D.; Stewart, J.; Pollock, J.S.; Pollock, D.M.; Imig, J.D. Soluble epoxide hydrolase inhibition protects the kidney from hypertension-induced damage. J. Am. Soc. Nephrol. 2004, 15, 1244–1253. [Google Scholar] [PubMed]
- Mittal, M.; Siddiqui, M.R.; Tran, K.; Reddy, S.P.; Malik, A.B. Reactive Oxygen Species in Inflammation and Tissue Injury. Antioxid. Redox Signal. 2014, 20, 1126–1167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sudhahar, V.; Shaw, S.; Imig, J.D. Epoxyeicosatrienoic Acid Analogs and Vascular Function. Curr. Med. Chem. 2010, 17, 1181–1190. [Google Scholar] [CrossRef] [PubMed]
- Fleming, I.; Rueben, A.; Popp, R.; Fisslthaler, B.; Schrodt, S.; Sander, A.; Haendeler, J.; Falck, J.R.; Morisseau, C.; Hammock, B.D.; et al. Epoxyeicosatrienoic Acids Regulate Trp Channel—Dependent Ca2+ Signaling and Hyperpolarization in Endothelial Cells. Arter. Thromb. Vasc. Biol. 2007, 27, 2612–2618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, H.; Vriens, J.; Prenen, J.; Droogmans, G.; Voets, T.; Nilius, B. Anandamide and arachidonic acid use epoxyeicosatrienoic acids to activate TRPV4 channels. Nature 2003, 424, 434–438. [Google Scholar] [CrossRef]
- Makita, K.; Takahashi, K.; Karara, A.; Jacobson, H.R.; Falck, J.; Capdevila, J.H. Experimental and/or genetically controlled alterations of the renal microsomal cytochrome P450 epoxygenase induce hypertension in rats fed a high salt diet. J. Clin. Investig. 1994, 94, 2414–2420. [Google Scholar] [CrossRef]
- Loch, D.; Hoey, A.; Morisseau, C.; Hammock, B.O.; Brown, L. Prevention of Hypertension in Doca-Salt Rats by an Inhibitor of Soluble Epoxide Hydrolase. Cell Biochem. Biophys. 2007, 47, 87–98. [Google Scholar] [CrossRef]
- Welch, W.J. Angiotensin Ii-Dependent Superoxide: Effects on Hypertension and Vascular Dysfunction. Hypertension 2008, 52, 51–56. [Google Scholar] [CrossRef] [Green Version]
- Griendling, K.K.; Sorescu, D.; Ushio-Fukai, M. Nad(P)H Oxidase: Role in Cardiovascular Biology and Disease. Circ. Res. 2000, 86, 494–501. [Google Scholar] [CrossRef] [Green Version]
- Gauthier, K.M.; Edwards, E.M.; Falck, J.R.; Reddy, D.S.; Campbell, W.B. 14,15-Epoxyeicosatrienoic Acid Represents a Transferable Endothelium-Dependent Relaxing Factor in Bovine Coronary Arteries. Hypertension 2005, 45, 666–671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moffat, M.P.; Ward, C.A.; Bend, J.R.; Mock, T.; Farhangkhoee, P.; Karmazyn, M. Effects of epoxyeicosatrienoic acids on isolated hearts and ventricular myocytes. Am. J. Physiol. Circ. Physiol. 1993, 264 Pt 2, H1154–H1160. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Lu, T.; Weintraub, N.; VanRollins, M.; Spector, A.A.; Shibata, E. Effects of epoxyeicosatrienoic acids on the cardiac sodium channels in isolated rat ventricular myocytes. J. Physiol. 1999, 519, 153–168. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.F.; Huang, L.; Morgan, J.P. Cytochrome P450: A novel system modulating Ca2+ channels and contraction in mammalian heart cells. J. Physiol. 1998, 508, 777–792. [Google Scholar] [CrossRef]
- Lu, T.; Ye, D.; Wang, X.; Seubert, J.M.; Graves, J.P.; Bradbury, J.A.; Zeldin, D.C.; Lee, H.C. Cardiac and Vascular Katp Channels in Rats Are Activated by Endogenous Epoxyeicosatrienoic Acids through Different Mechanisms. J. Physiol. 2006, 575 Pt 2, 627–644. [Google Scholar] [CrossRef]
- Xu, D.; Li, N.; He, Y.; Timofeyev, V.; Lu, L.; Tsai, H.-J.; Kim, I.-H.; Tuteja, D.; Mateo, R.K.P.; Singapuri, A.; et al. Prevention and reversal of cardiac hypertrophy by soluble epoxide hydrolase inhibitors. Proc. Natl. Acad. Sci. USA 2006, 103, 18733–18738. [Google Scholar] [CrossRef] [Green Version]
- Nadtochiy, S.M.; Zhu, Q.; Urciuoli, W.; Rafikov, R.; Black, S.M.; Brookes, P.S. Nitroalkenes Confer Acute Cardioprotection via Adenine Nucleotide Translocase 1. J. Biol. Chem. 2012, 287, 3573–3580. [Google Scholar] [CrossRef] [Green Version]
- Sun, W.-J.; Xu, D.-G.; Hu, H.-L.; Zou, M.-J.; Du, J.-F.; Wang, J.-F.; Cai, X.; Wang, J.-X.; Ai, H.-S. Construction and characterization of soluble HLA-A*0201-PR1 complex. Zhongguo Shi Yan Xue Ye Xue Za Zhi 2007, 15, 352–356. [Google Scholar]
- Jamieson, K.; Darwesh, A.; Sosnowski, D.; Zhang, H.; Shah, S.; Zhabyeyev, P.; Yang, J.; Hammock, B.; Edin, M.; Zeldin, D.; et al. Soluble Epoxide Hydrolase in Aged Female Mice and Human Explanted Hearts Following Ischemic Injury. Int. J. Mol. Sci. 2021, 22, 1691. [Google Scholar] [CrossRef]
- Nishino, T.; Okamoto, K.; Eger, B.T.; Pai, E.F.; Nishino, T. Mammalian xanthine oxidoreductase-mechanism of transition from xanthine dehydrogenase to xanthine oxidase. FEBS J. 2008, 275, 3278–3289. [Google Scholar] [CrossRef]
- Li, J.-M.; Gall, N.P.; Grieve, D.J.; Chen, M.; Shah, A.M. Activation of NADPH Oxidase During Progression of Cardiac Hypertrophy to Failure. Hypertension 2002, 40, 477–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benhar, M.; Forrester, M.T.; Hess, D.T.; Stamler, J.S. Regulated Protein Denitrosylation by Cytosolic and Mitochondrial Thioredoxins. Science 2008, 320, 1050–1054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irie, Y.; Saeki, M.; Kamisaki, Y.; Martin, E.; Murad, F. Histone H1.2 is a substrate for denitrase, an activity that reduces nitrotyrosine immunoreactivity in proteins. Proc. Natl. Acad. Sci. USA 2003, 100, 5634–5639. [Google Scholar] [CrossRef] [Green Version]
- Batthyány, C.; Schopfer, F.J.; Baker, P.R.S.; Durán, R.; Baker, L.M.S.; Huang, Y.; Cerveñansky, C.; Branchaud, B.P.; Freeman, B.A. Reversible Post-translational Modification of Proteins by Nitrated Fatty Acids in Vivo. J. Biol. Chem. 2006, 281, 20450–20463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abis, G.S.A.; Stockmann, H.B.A.C.; Bonjer, H.J.; van Veenendaal, N.; van Doorn-Schepens, M.L.M.; Budding, A.; Wilschut, J.; van Egmond, M.; Oosterling, S.J.; de Lange, E.S.M.; et al. Randomized clinical trial of selective decontamination of the digestive tract in elective colorectal cancer surgery (SELECT trial). Br. J. Surg. 2019, 106, 355–363. [Google Scholar] [CrossRef] [PubMed]
- Bradshaw, J.M.; McFarland, J.M.; Paavilainen, V.O.; Bisconte, A.; Tam, D.; Phan, V.T.; Romanov, S.; Finkle, D.; Shu, J.; Patel, V.; et al. Prolonged and tunable residence time using reversible covalent kinase inhibitors. Nat. Chem. Biol. 2015, 11, 525–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez-Bello, C. Designing Irreversible Inhibitors—Worth the Effort? Chem. Med. Chem. 2016, 11, 22–30. [Google Scholar] [CrossRef]
- Singh, J.; Petter, R.C.; Baillie, T.A.; Whitty, A. The resurgence of covalent drugs. Nat. Rev. Drug Discov. 2011, 10, 307–317. [Google Scholar] [CrossRef]
- Smith, A.J.T.; Zhang, X.; Leach, A.G.; Houk, K.N. Beyond Picomolar Affinities: Quantitative Aspects of Noncovalent and Covalent Binding of Drugs to Proteins. J. Med. Chem. 2009, 52, 225–233. [Google Scholar] [CrossRef] [Green Version]
- Steinhubl, S.R. Why Have Antioxidants Failed in Clinical Trials? Am. J. Cardiol. 2008, 101, S14–S19. [Google Scholar] [CrossRef]
- Vivekananthan, D.P.; Penn, M.S.; Sapp, S.K.; Hsu, A.; Topol, E.J. Use of Antioxidant Vitamins for the Prevention of Cardiovascular Disease: Meta-Analysis of Randomised Trials. Lancet 2003, 361, 2017–2023. [Google Scholar] [CrossRef]
- Prysyazhna, O.; Wolhuter, K.; Switzer, C.; Santos, C.; Yang, X.; Lynham, S.; Shah, A.M.; Eaton, P.; Burgoyne, J.R. Blood Pressure-Lowering by the Antioxidant Resveratrol Is Counterintuitively Mediated by Oxidation of Cgmp-Dependent Protein Kinase. Circulation 2019, 140, 126–137. [Google Scholar] [CrossRef] [PubMed]
- Cronin, A.; Sherry, M.; Heike, D.; Shirli, H.; Ingrid, F.; Beate, F.; Franz, O.; Michael, A. The N-Terminal Domain of Mammalian Soluble Epoxide Hydrolase Is a Phosphatase. Proc. Natl. Acad. Sci. USA 2003, 100, 1552–1557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newman, J.W.; Morisseau, C.; Harris, T.R.; Hammock, B.D. The soluble epoxide hydrolase encoded by EPXH2 is a bifunctional enzyme with novel lipid phosphate phosphatase activity. Proc. Natl. Acad. Sci. USA 2003, 100, 1558–1563. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
{kind=link}
{kind=link}
| oxPTM | sEH Activity | Potential and Known Effects on Cardiovascular Function |
|---|---|---|
| Tryosine nitration | Inhibited | Angiogenic Anti-inflammatory Cardioprotective Anti-hypertensive Coronary vasodilatory Increase myocyte contraction |
| Electrophilic lipid adduction | Inhibited | Angiogenic Anti-inflammatory Cardioprotective Anti-hypertensive Coronary vasodilatory Increase myocyte contraction |
| S-nitrosation | Activated | Anti-angiogenic Vasoconstriction Hypertrophic Increased ischemic & reperfusion injury |
| Intra-disulfide Formation | Activated | Anti-angiogenic Vasoconstriction Hypertrophic Increased ischemic & reperfusion injury |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Charles, R.; Eaton, P. Redox Regulation of Soluble Epoxide Hydrolase—Implications for Cardiovascular Health and Disease. Cells 2022, 11, 1932. https://doi.org/10.3390/cells11121932
Charles R, Eaton P. Redox Regulation of Soluble Epoxide Hydrolase—Implications for Cardiovascular Health and Disease. Cells. 2022; 11(12):1932. https://doi.org/10.3390/cells11121932
Chicago/Turabian StyleCharles, Rebecca, and Philip Eaton. 2022. "Redox Regulation of Soluble Epoxide Hydrolase—Implications for Cardiovascular Health and Disease" Cells 11, no. 12: 1932. https://doi.org/10.3390/cells11121932
APA StyleCharles, R., & Eaton, P. (2022). Redox Regulation of Soluble Epoxide Hydrolase—Implications for Cardiovascular Health and Disease. Cells, 11(12), 1932. https://doi.org/10.3390/cells11121932