1. Introduction
Sensing the Earth´s magnetic field is a widespread ability in the animal kingdom. Behavioral experiments and analyses of data from wild animals have shown that night-migratory songbirds, fish, amphibians, reptiles and insects orientate and navigate using magnetic sensing [
1,
2,
3,
4,
5]. Birds sense the inclination of the magnetic field by a process that involves the absorbance of blue light, indicating the involvement of a photoreceptor protein [
2,
6,
7,
8]. In a seminal hypothesis, Schulten et al. (1978) proposed a radical-pair mechanism as the underlying mechanism for magnetic sensing [
9], and a ubiquitously present class of chromophore proteins called cryptochromes (Cry) seemed to match the required properties for a magnetosensor [
6,
10,
11]. Further research showed that magnetic compass information is detected in the retina and processed in the birds’ visual system [
12]. However, the exact molecular basis of this complex process is currently unknown.
Cry proteins are flavoproteins harboring a flavin adenine dinucleotide (FAD) in a binding pocket situated in a conserved photolyase homology region. The FAD chromophore enables Cry to absorb blue light, which triggers the formation of a radical pair involving a series of three or four neighboring tryptophan (Trp) residues [
8,
13].
Thus far, six different Crys (Cry1a, Cry1b, Cry2a, Cry2b, Cry4a and Cry4b) have been identified to be expressed in the retina of different bird species, but only Cry1a and Cry4a are currently discussed as the primary magnetic field receptors. Cry1a is localized in UV-sensitive cones in the retina of European robins [
14] and a light-dependent immunostaining pattern of Cry1a in UV cones was interpreted to detect a light-triggered conformational change in Cry1a [
14,
15]. However, this light-dependent staining pattern of Cry1a in UV cones was not supported by a more recent study [
16]. Instead, several lines of evidence point towards Cry4a being the magnetoreceptive protein. European robin Cry4a (
ErCry4a) can be purified with a bound FAD chromophore, exhibits a photo-induced electron transfer pathway leading to the formation of a [FAD
●− TrpH
●+] radical pair, and it is magnetically sensitive in vitro [
17,
18,
19]. Furthermore, Xu et al. (2021) found a substantially higher magnetic field effect in
ErCry4a than in chicken and pigeon Cry4 [
19]. In a separate study, Hochstoeger et al. (2020) presented evidence for pigeon Cry4 acting as an ultraviolet-blue photoreceptor that forms photo-induced radical pairs [
20]. These authors localized Cry4 in horizontal cells of the Pigeon retina and proposed a magnetic sensing mechanism by modulation of glutamatergic synapses. The immunohistochemistry of Cry4 in the Pigeon retina differs from results obtained with retinae from European robin, where
ErCry4 expresses in the outer segments of double cones and long-wavelength single cones but not in other retinal cells [
21]. In the same study, Günther et al. (2018) co-localized
ErCry4 and iodopsin (long wavelength opsin) in long-wavelength single cones and in double cones [
21].
A further important step for validating that Cry4 variants indeed operate as magnetoreceptive molecules is to link them to signal transduction processes causing magnetic-field-sensitive changes in the membrane potential. Wu et al. (2020) identified six putative protein interaction partners of
ErCry4a by a yeast two-hybrid screening approach [
22]. All six genes code for retina-specific proteins. These proteins are
GNAT2 coding for the α-subunit of a cone-specific heterotrimeric G protein,
GNG10 coding for the γ-subunit of a cone-specific heterotrimeric G protein,
LWS, also called iodopsin, coding for long-wavelength-sensitive opsin,
KCNV2 coding for potassium voltage-gated channel subfamily V member 2,
RBP1 coding for retinol-binding protein 1 and
RGR coding for retinal G protein-coupled receptor. The identified α- and γ-subunit of the G protein are the cone-specific orthologues of the heterotrimeric G protein transducin (G
t) that mediates phototransduction in mammalian and other vertebrate rod and cone photoreceptor cells. The versatile role of heterotrimeric G proteins in signal transduction pathways makes them primary candidates to test for direct interaction with Cry4 variants. Could the G protein be the long-sought-after first interaction partner of Cry4a in a radical-pair-based magnetoreception signaling pathway?
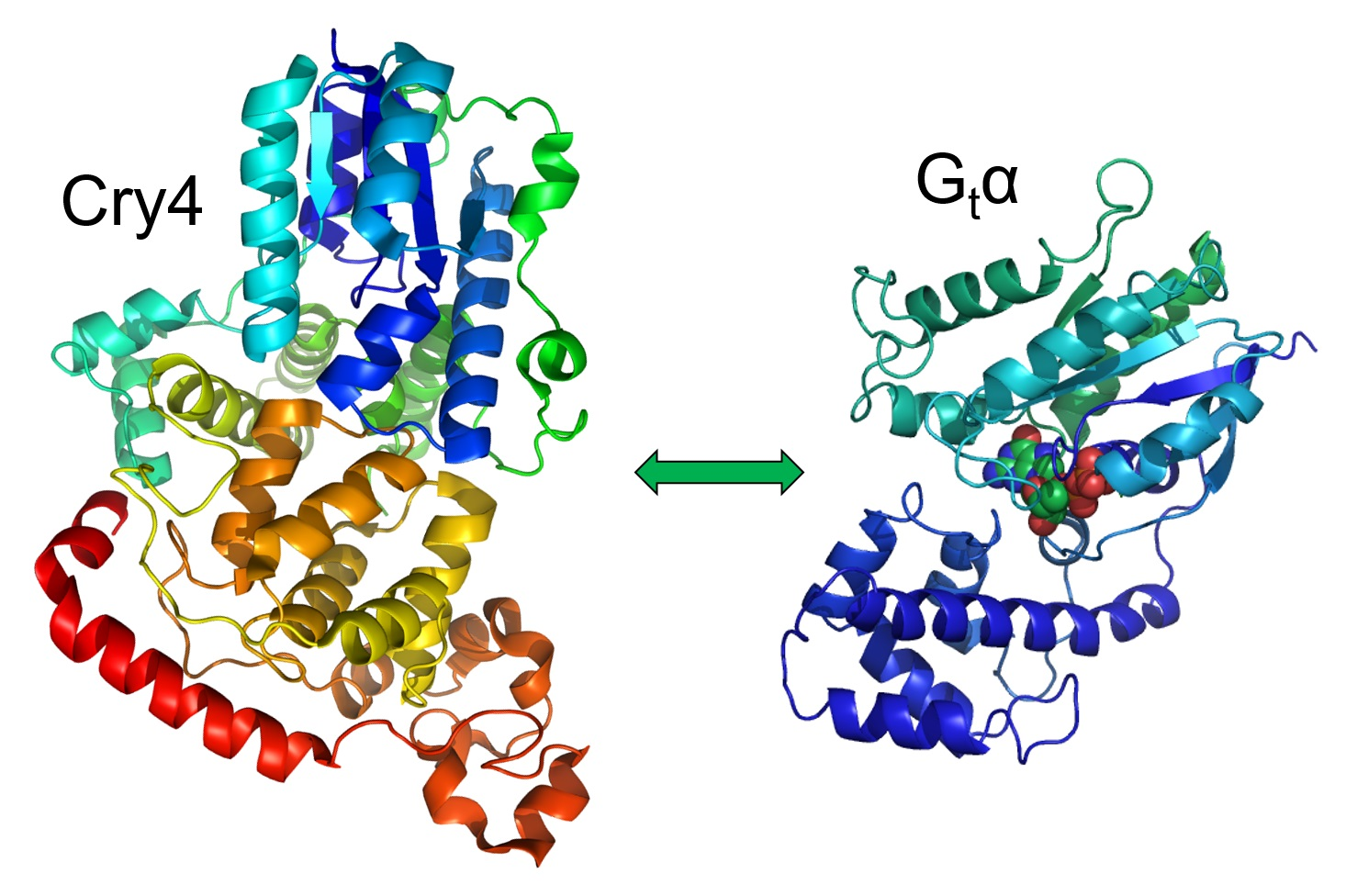
The aim of the present study is to test whether ErCry4a directly interacts with the G protein α-subunit of European robin (ErGtα) on the molecular level. We expressed recombinant variants of both proteins, purified them and verified their functional state by independent functional assays. Using a biosensor approach employing surface plasmon resonance (SPR), we investigated the protein–protein interaction process and analyzed the kinetic parameter of the binding process, which was further corroborated by in vitro pulldown affinity assays. Using acceptor photobleaching Förster resonance energy transfer (FRET), we further validated that the interaction between ErCry4a and ErGtα also occurs in living cells in a neuroretinal bird cell line.
2. Materials and Methods
2.1. General Cloning Strategies
All primers for cloning steps are listed in the
Supplement Materials (Table S1). The coding sequence of
ErGnat2 (coding for cone-specific Gα from European robin; see
Figures S1–S3 for sequence information and alignments) was present in the pGBKT7 vector as described by Wu et al. [
22]. A PCR was performed to amplify the coding sequence and add flanking
SpeI and
XhoI restriction sites. The product was subsequently digested and ligated with digested pCold vector (Takara Bio, Shiga, Japan) to create the pCold
ErGnat2 construct.
To create the chimera, a deletion mutagenesis of amino acids 220–298 of ErGnat2 in the pCold vector was performed, and the corresponding region of bovine inhibitory alpha subunit was amplified from the commercial pCS6 vector containing bGNA1i. The products were then assembled using the Gibson assembly to create the completed pCold ErGnat2 chimera. To create the pET21a ErGnat2 chimera vector, the chimera sequence was amplified from pCold and assembled using the Gibson assembly with NdeI and XhoI digested pET21a. To create the pACE SUMO ErGnat2 chimera vector, the chimera sequence was amplified and combined with the pACE SUMO vector using the Sequence and Ligation Independent Cloning (SLIC) technique. To remove 6 non-native amino acids from the coding sequence, PCR mutagenesis was performed.
2.2. Cloning of the Gtα/Giα Chimera Containing a SUMO Tag
Earlier work on mammalian G
tα showed that expression in
E. coli resulted in misfolded protein [
23], and we obtained the same result by trying to express wildtype
ErG
tα. However, it is common practice to construct, express and purify chimeric mammalian G
tα proteins for functional studies [
23]. The chimeric G-protein α-subunit G
tα/Giα is based on the coding sequence for the
European robin gene
ErGNAT2 [
22] encoding
ErGtα and the
Bovine taurus Giα1 (
Figure S1). The sequence was cloned seamlessly into a T7lac expression vector containing a T7lac promoter and a Small Ubiquitin-Related Modifier (SUMO) protein tag modified with an N-terminal 6x histidine tag (
Figure S4). The initiator methionine of the
ErGNAT2 sequence was removed, and plasmids were confirmed by Sanger sequencing (Eurofins Genomics). Primer pairs utilized and more details about the cloning steps can be found in
Supplement Table S1.
The plasmid was transformed into chemically competent BL21 E. coli cells (prepared in-house). Subsequently, the cells were cultured at 37 °C and 180 rpm in LB medium containing 10 g/L NaCl and 100 µg/L ampicillin (Carl Roth, Karlsruhe, Germany). Upon reaching an OD600 of 0.6, the shaking was reduced to 160 rpm and the cells were cooled to 17 °C for 30 min. Next, the cells were induced by adding IPTG (Roth) to 10 µM, incubated for 20–24 h at 17 °C, and then harvested and stored at −20 °C.
Cells from 3.5 L culture were resuspended in 50 mL Ni-NTA binding buffer (20 mM HEPES pH 7.8, 250 mM NaCl, 10 mM MgCl2, 10 mM Imidazole, 10 mM ß-mercaptoethanol, Roche cOmplete™ EDTA-free Protease Inhibitor Cocktail) and lysed by sonication using a Bandelin GM 2200 ultrasonic generator with a UW 2200 converter and MS72 microtip at 45% power. Cell lysates were then centrifuged at 18,000 rpm (~25,000 g avg) in a JA25.5 rotor at 4 °C for 60 min, and the supernatants were applied to equilibrated gravity flow columns containing 2 mL of Ni-NTA resin each. Each column was washed with 80 matrix volumes (160 mL) wash buffer (10 mM HEPES pH 7.8, 250 mM NaCl, 10 mM MgCl2, 20 mM imidazole, 10 mM β-mercaptoethanol), and then eluted twice using 4 mL of elution buffer (20 mM HEPES pH 7.8, 150 mM NaCl, 10 mM MgCl2, 300 mM imidazole, 10 mM β-mercaptoethanol).
Both elution fractions were dialyzed together (SERVAPOR® dialysis tubing, MWCO 12,000–14,000 RC, diameter 21 mm) after adding Ulp1 protease (in-house) against at least 4 L of dialysis buffer (10 mM HEPES pH 7.8, 250 mM NaCl, 10 mM MgCl2) overnight. The digest was then applied to the same equilibrated gravity flow columns containing Ni-NTA matrix to remove digested tag sequences. For this, the columns were washed 3 times with 5 mL wash buffer and eluted with 5 mL elution buffer. The flow-through and wash fractions containing the Gtα/Giα chimera were pooled and concentrated using a Macrosep Advance 10k MWCO centrifugal filter to a concentration of up to 15 mg/mL. Concentrated fractions were applied to house-packed size exclusion chromatography column (Superdex75, volume of 320 mL) pre-equilibrated with size exclusion chromatography buffer (10 mM HEPES pH 7.4, 150 mM NaCl, 10 mM MgCl2, 3.4 mM EDTA). The flow rate was maintained at 1 mL/min, and fractions containing the G-protein were identified with SDS Page, pooled, and concentrated and, then, flash frozen in liquid N2 in 50 or 100 µL aliquots and stored at −80 °C.
2.3. Expression and Purification of the Myristoylated Gtα/Giα Chimera
E. coli BL21 (Codon+) cells, harboring the pET21a G
tα/G
iα chimera plasmid and pBB131 encoding yeast myristoyltransferase, were grown in 5 × 500 mL yeast tryptone (YT) media at 37 °C and 180 rpm in the presence of 100 µg/mL ampicillin (Roth) and 30 µg/mL kanamycin (Carl Roth, Karlsruhe, Germany). At OD
600 of 0.4 myristic acid (Fluka, Buchs, Switzerland) was added to a final concentration of 50 µg/mL, and the incubation was continued until an OD
600 of 0.5–0.6 was reached. Then, the cultures were cooled down to 17 °C at 160 rpm for 30 min. IPTG (Roth) was added to a final concentration of 150 µM, and incubation was continued for 20–24 h at 17 °C and 160 rpm. Afterward, the cell pellets were harvested by centrifugation at 7000 rpm for 7 min at 4 °C. Each cell pellet was resuspended in 10 mL in Ni-NTA binding buffer (20 mM Tris-HCl pH 8.0, 250 mM NaCl, 10 mM imidazole) and a protease inhibitor cocktail (Roche cOmplete™ EDTA-free Protease Inhibitor Cocktail) was added (one tablet dissolved in 2 mL of water, of which 0.2 mL were added to 10 mL of Ni-NTA binding buffer). The cells were lysed by ultrasonication using a Bandelin GM 2200 ultrasonic generator with a UW 2200 converter and MS72 microtip at 45% power and centrifuged at 100,000×
g for 30 min at 4 °C. The supernatant was used for further purification. An empty 20 mL column was packed with 2.5 mL Ni
2+-nitrilotriacetic acid-agarose resin (HisPurTM Ni-NTA) and equilibrated with 2 × column volumes of Ni-NTA binding buffer. The supernatant was loaded on the Ni-NTA column by gravity flow and washed with 50 × matrix volume of Ni-NTA washing buffer (20 mM Tris-HCl pH 8.0, 150 mM NaCl, 20 mM imidazole, 10 mM β-mercaptoethanol). The protein was eluted with 2 × matrix volume of Ni-NTA elution buffer (20 mM Tris-HCl pH 8.0, 150 mM NaCl, 300 mM imidazole, 10 mM β-mercaptoethanol). Next, an anion exchange chromatography (AEC) was performed. The elution fraction from the NTA-affinity chromatography was diluted 1:3 in AEC buffer A (20 mM Tris-HCl pH 7.0 and 1 mM DTT). A HiTrap
TM 5 mL Q Sepharose High Performance (QHP, Cytiva, Uppsala, Sweden) column was equilibrated with 5% AEC buffer B (20 mM Tris-HCl pH 7.0, 1 M NaCl and 1 mM DTT) and 95% AEC buffer A. The protein sample was loaded on the column and washed with 5% AEC buffer B and a flow rate of 1 mL/min until a stable UV signal was reached. The protein was eluted with a salt gradient from 5 to 50% AEC buffer B over 112.5 mL at a flow rate of 1 mL/min. Fractions containing the G
tα/G
iα chimera were pooled, and to remove all remaining contamination, a size exclusion chromatography (SEC) was performed. A Superdex 200 16/600 column with a volume of 120 mL was equilibrated using SPR buffer (10 mM Hepes pH 7.4, 150 mM NaCl, 10 mM MgCl
2, 3.4 mM EDTA), and the pooled AEC fractions were concentrated using a Macrosep Advance 10k MWCO centrifugal filter. Using a flow rate of 0.5 mL/min, the sample was applied to the column, and the SEC run started. Fractionation was started after 0.4 CV using 3 mL fraction sizes. Desired fractions were pooled, concentrated, and flash frozen in liquid N
2 for storage at −80 °C. Myristoylation was analyzed and verified by reversed phase analytical HPLC as described previously for myristoylated neuronal calcium sensor proteins [
24], yielding 80% myristoylation. SDS-PAGE analyses of Gtα/Giα chimera variants did not show any heterogeneity indicating higher myristoylation rates (
Figure S5).
2.4. Cloning, Expression, and Purification of ErCry Variants
Basic steps of cloning, expression and purification of ErCy4a have been described previously [
19]. The following modifications were applied: the LB media for expression contained 10 g/L yeast extract instead of 5 g/L. The C-terminally truncated mutant
ErCry4-497 was produced by mutagenesis using forward and reverse primers as listed in the
Supplementary Materials (Table S1) on the WT
ErCry4a pCold plasmid as described [
19].
ErCRY4-497 was expressed and purified as
ErCry4a, except that the expression time was extended from 22 to 44 h.
ErCry1a and
ErCry1b, as well as the chimeric G-protein α-subunit Gtα/Giα were cloned into the pFastBacHT B vector (Thermo Fisher, Waltham, MA, USA) using
BamHI and
XhoI restriction sites and primers listed in
Supplement Table S1. Mouse-codon optimized
ErCRY4 in the pFast vector was a gift from Joseph S. Takahashi (University of Texas Southwestern Medical Center, Dallas, TX, USA).
Baculovirus was produced in SF9 cells (Thermo Fisher) using the Bac-to-Bac Baculovirus expression system (ThermoFisher), while Tni cells (BioTrend, Cologne, Germany) were used for protein expression. Cells from 1 L culture were sedimented by centrifugation at 3745× g and resuspended in 30 mL homogenization buffer (50 mM Tris, pH 8.8, 300 mM NaCl, 15 mM imidazole, 10 mM β-mercaptoethanol, Roche cOmplete™ EDTA-free Protease Inhibitor Cocktail) per 10 g cell pellet. Cells were lysed with a Potter-Elvehjem-homogenizer and clarified by centrifugation at 48,384× g. Clarified cell lysates were applied to Ni-NTA agarose columns (Qiagen, Hilden, Germany), pre-equilibrated with homogenization buffer. Bound proteins were eluted with elution buffer (50 mM Tris, 300 mM NaCl, 400 mM Imidazole and 10 mM β-mercaptoethanol; pH 8.0 for ErCry4 and Gtα/Giα, and pH 8.8 for ErCry1a and ErCry1b). The proteins were diluted 1:10 in 20 mM Tris and further purified on anion-exchange 5 mL Hitrap Q columns (Cytiva, Uppsala, Sweden), after equilibration with buffer A (20 mM Tris, 30 mM NaCl, and 10 mM β-mercaptoethanol; pH 8 or 8.8). Proteins were eluted with a gradient increasing NaCl to 0.5 M at a flow rate of 1 mL/min. While fractions containing ErCry4 were yellow in color, due to the absorption of bound FAD, those of ErCry1a and ErCry1b were colorless.
2.5. Limited Proteolysis of the Gtα/Giα Chimera
The proteolytic digestion was performed under conditions such as those previously described [
25,
26] for tryptophan fluorescence analysis. Purified and frozen G
tα/G
iα chimera samples were thawed. Two reaction mixtures were prepared, both containing 50 mM Tris/HCl at pH 7.4, 50 mM NaCl, 2 mM MgCl
2, 10 µM GDP, and 10 µM G
tα/G
iα chimera. To one reaction, 50 µM AlCl
3 and 10 mM NaF were added. The reactions were started by adding chymotrypsin (Roth), from a frozen stock solution containing 0.5 mg/mL chymotrypsin in 0.5 mM HCl and 50% glycerol, to 12.5 µg/mL and a final volume of 300 µL. Upon addition of protease, the reactions were incubated at 37 °C. Digestion was stopped after 0.5, 2, 5, 10, 20, 40, 60, and 90 min by removing 30 µL of the reaction mixture each, mixing with 10 µL of 4× Laemmli sample buffer, heating to 95 °C for 3 min, and storage at −20 °C. Additionally, a sample was prepared before the addition of protease. Analysis was performed by loading identical volumes of each sample onto 10% Bis-tris SDS-PAGE gels. Gels were stained using Coomassie Brilliant Blue R250 and decolorized using 10% acetic acid and 40% ethanol. Visualization was performed using an Azure c400 Gel Imaging System by Azure Biosystems, Dublin, CA, USA.
2.6. Trp Fluorescence Emission
The intrinsic Trp fluorescence assay for measuring the activation-dependent conformational change in G protein α-subunits is a widely used assay to test their functional intactness. Details of the experimental setup are described [
26]. We recorded relative fluorescence emission on a spectrofluorimeter from Photon Technology International. Purified G
tα/Giα sample was thawed and diluted in fluorescence buffer (50 mM Tris pH 7.4, 50 mM NaCl, 10 µM GDP and varying concentrations of MgCl
2) at a final concentration of 1 µM. Trp fluorescence emission was recorded at 340 nm after excitation at 290 nm. Changes in emission rate were triggered by injection of 50 µM AlF
4− (premix of 50 µM AlCl
3 and 10 mM NaF). Recording time was 500 s.
2.7. Pulldown Experiments
Purification of ErCry4 and Gtα/Giα chimera—Both ErCry4a and non-myristoylated Gtα/Giα chimera were expressed and purified essentially as described above, except that all steps were carried out under far-red light (750+ nm). For the Gtα/Giα chimera, concentrated fractions after affinity purification were not digested to remove the tag but directly purified by size exclusion chromatography using a Superdex200 increase 10/300 column pre-equilibrated with a running buffer (20 mM HEPES, pH 7.4, 150 mM NaCl, 10 mM MgCl2). The flow rate was maintained at 0.17 mL/min, and fractions containing the G-protein were identified with SDS-PAGE and used immediately for pulldown experiments. For ErCry4a, after affinity purification, elution fractions were dialyzed together (Spectra/Por dialysis tubing, MWCO 6000–8000) after adding TEV protease (in-house) to an approximate molar ratio of 1:20 against at least 3 L of dialysis buffer (20 mM HEPES, pH 7.4, 200 mM NaCl, 10 mM MgCl2) overnight. The digest was then applied to the gravity flow columns (Bio-Rad Laboratories GmbH, Feldkirchen, Germany containing Super Ni-NTA agarose resin (Anatrace) equilibrated with dialysis buffer to remove His-tag and protease. The flow-through containing ErCry4a was concentrated using a Vivaspin Turbo 15 10k MWCO centrifugal filter and further purified by size exclusion chromatography using a Superdex200 increase 10/300 column (Cytiva, Uppsala, Sweden) pre-equilibrated with a running buffer (20 mM HEPES, pH 7.4, 150 mM NaCl, 10 mM MgCl2). The flow rate was maintained at 0.17 mL/min, and fractions containing ErCry4a were identified with SDS-PAGE and used immediately for pulldown experiments.
Pulldown—All steps until SDS-PAGE gel electrophoresis were carried out under far-red light (750+ nm). For protein pulldown experiments, 1.47 nmol each of ErCry4a and His-Gtα/Giα were mixed and incubated overnight at 6 °C with gentle agitation. For the negative controls, His-Gtα/Giα was replaced with a running buffer. Next, 50 µL Super Ni-NTA agarose resin (Anatrace, Maumee, OH, USA) was added, and the sample was incubated for an additional 1.5 h at 6 °C with gentle agitation. The slurry was transferred to Micro Bio-SpinTM Columns (Bio-Rad Laboratories GmbH, Feldkirchen, Germany), washed five times with 1 mL of wash buffer (20 mM HEPES, pH 7.4, 150 mM NaCl, 10 mM MgCl2, 20 mM imidazole), then incubated with 70 µL elution buffer (20 mM HEPES, pH 7.4, 150 mM NaCl, 10 mM MgCl2, 500 mM imidazole) for 30 min on ice. For elution, columns were centrifuged for 1 min at 4 °C and 500 g. SDS-PAGE samples were prepared as follows. For the input controls, 2.5 µg of either ErCry4a or His-Gtα/Giα were mixed with SDS-sample buffer, boiled for 5 min at 95 °C, and then placed on ice until loaded on the gel. For the other samples, 20 µL of the elution fraction were mixed with 5 µL SDS-sample buffer, boiled for 5 min at 95 °C, then placed on ice until loaded on the gel. Samples were run on 4–20% Tris SDS-PAGE gradient gels. Gels were stained using Coomassie Brilliant Blue R250 (Sigma-Aldrich, Taufkirchen, Germany and decolorized using 10% acetic acid and 40% ethanol. Visualization was performed using a Fusion FX6 Edge (Vilber, Eberhardzell, Germany) imaging system.
Densiometric analysis—Analysis was carried out in the Fiji software package using the SDS-PAGE analysis function allowing for the integration of grey values with background correction. For each experiment, the ErCry4a input band was quantified. Then the same region of interest was quantified for negative control and pulldown lanes. The bar diagram visualizes the average percentage of input ErCry4a recovered in negative control and pulldown, respectively, with error bars indicating the standard deviation. Individual experimental results are indicated by circles.
2.8. Surface Plasmon Resonance
Surface plasmon resonance (SPR) measurements were performed on a Biacore 3000 (GE Healthcare now Cytiva). The general operation principle, including the immobilization procedures and quantitative data analysis, has been described before [
27,
28]. We used CM5 sensor chips (GE Healthcare) for all applications. The carboxy-methyl dextran coated sensor chip surface of CM5 sensor chips was activated by carbodiimide/N-hydroxy-succinimide chemistry (Biacore Immobilization Kit, Cytiva, Uppsala, Sweden, allowing subsequent covalent coupling of proteins via free NH
2-groups. In preliminary tests, we compared different immobilization geometries, immobilization densities and regeneration protocols. Immobilization densities of myristoylated Gtα/Giα were 3.6–3.7 ng/mm
2 and of non-myristoylated Gtα/Giα were 2.6–3.9 ng/mm
2. Interaction processes were studied by injection of
ErCry variants at different concentrations at a flow rate of either 5 or 20 µL/min. Some recordings were also performed at higher flow rates of 30 and 50 µL/min. SPR running buffer was 10 mM HEPES/NaOH, pH 7.4, 150 mM NaCl, 10 mM MgCl
2, 0.005% Tween-20, 3.4 mM EDTA. Control surfaces were coated with Ulp1 (in-house made) at a density of 2.4–3.6 ng/mm
2 or by an amino coupling activation/deactivation cycle (SPR recordings with
ErCry1a).
For the evaluation of sensorgrams, we used nonlinear curve fitting implemented in the BIAevaluation software 4.1 (GE Healthcare, Boston, MA, USA) by applying the global fitting approach. Sensorgrams obeying a mono-exponential Langmuir binding process (A + B ↔ AB) yielded association and dissociation rate constants and apparent KD values. SPR sensorgrams that did not show mono-exponential binding curves were evaluated by a two-state-reaction model according to A + B ↔ AB ↔AB* (* indicates a different protein conformation). The latter describes the binding process of A + B by a forward rate constant ka1 and a backward rate constant kd1 leading to complex formation AB and a conformational change of AB to AB* (BIAevaluation software 4.1). We calculated the apparent KD from the ratio of kd1/ka1.
2.9. FRET Measurements
Cloning of FRET constructs—cDNA production: RNA was extracted from the retina of one European robin, which was wild-caught in the vicinity of the university campus using mist nets. The animal was sacrificed by decapitation, and the eyes were immediately removed. The retina, free of vitreous, was shock-frozen in liquid nitrogen and stored at −80 °C until RNA extraction. RNA was extracted using the NucleoSpin RNA XS kit (Macherey Nagel, Düren, Germany), and a cDNA library was generated using the Make Your Own “Mate & PlateTM” Library System (Takara Bio, Shiga, Japan).
All FRET constructs were based on the pKan-CMV-mClover3-mRuby3 vector, a gift from Michael Lin (Addgene plasmid #74252;
http://n2t.net/addgene (accessed on 26 February 2020): 74252; RRID: Addgene_74252) [
29].
Table 1 gives an overview of FRET plasmids and expressed proteins. pKan-CMV-mClover3-
ErCRY4 was generated by first linearizing pKanCMV-mClover3-mRuby3 with primers 1 and 2 (
Table S1) using PrimeSTAR Polymerase (Takara Bio, Shiga, Japan), and amplifying the
ErCRY4 cDNA with primer 3, introducing a
BamHI and
AscI restriction site and a Ser-Gly-Ser-Ser-Gly-Ser-Ser-Gly linker between mClover3 and
ErCRY4, and the reverse primer 4, introducing a
XhoI restriction site and a stop codon after
ErCRY4. The linearized vector and gene product were then recombined using In-Fusion (Takara Bio).
pKan-CMV-mClover3 was generated from pKan-CMV-mClover3-ErCRY4 by deletion of ErCRY4 and mRuby3 by using the Q5 site-directed mutagenesis kit (New England Biolabs) with primers 5 and 6, introducing a XhoI restriction site and a stop codon after the mClover3 plus linker. pKan-CMV-mClover3-ErCRY4-497 was generated by amplifying the ErCRY4 cDNA with primers 7 and 8 and In-Fusion recombination with pKan-CMV-mClover3 digested with AscI and XhoI. pKan-CMV-mRuby3 was generated by deletion of mClover3 from pKan-CMV-mClover3-mRuby3 using the Q5 site-directed mutagenesis kit (New England Biolabs, Ipswich, MA, USA) and primers 9 and 10. pKan-CMV-ErGNAT2-mRuby3, with GNAT2 being the corresponding gene to the wildtype ErGtα protein, was generated by first linearizing pKan-CMV-mClover3-mRuby3 with primers 11 and 12 using PrimeSTAR Polymerase, and then amplifying the GNAT2 gene with primers 13, introducing an AscI restriction site, and 14, introducing a XhoI restriction site. Both PCR products were then recombined using In-Fusion, resulting in the deletion of mClover3 and a fusion construct of GNAT2-mRuby3. The GNAT2 sequence was compared to the genome sequence (NCBI Sequence ID LR812129.1).
Cell Culture and Expression—The QNR/K2 Neuroretina Quail cell line was purchased from ATCC (CRL-2533) and cultured in DMEM + GlutaMAX (Gibco, Waltham, MA, USA) supplemented with 10% fetal bovine serum (Gibco) at 39 °C and 5% CO2. QNR/K2 cells were (co-)transfected with the FRET constructs using Lipofectamine 2000 (Invitrogen) according to manufacturer’s instructions, after which cells were handled in dim red light until fixed. Forty-eight hours after transfection, cells were washed twice with PBS (Gibco), fixed in 4% (wt/v) paraformaldehyde for 30 min, and washed again twice with PBS and kept in PBS for imaging. Transfection, fixation and imaging were all performed in µ-Slides 8 Well with a #1.5 polymer coverslip bottom (Ibidi).
Acceptor photobleaching FRET—Imaging of transiently transfected QNR/K2 cells was performed with an inverted Leica TCS SP5 II confocal microscope. A 63×/water objective with 1.2 numerical aperture was used (PL APO, corr CS). A white light laser was used as an excitation light source. The excitation wavelengths were set to 488 nm for mClover3 fluorescent protein and 560 nm for mRuby3. Photomultiplier tubes were used as detectors. The detection range was set to 500–550 nm for mClover3 emission and 570–644 nm for mRuby3 emission using AOTF (acousto-optical tunable filter). To determine the FRET efficiency, the acceptor photobleaching protocol was used. Acceptor photobleaching was performed with the FRET AB wizard in the LAS AF software on selected regions of interest (ROIs), generally one to two per cell in a region where both fluorophores showed expression. Acceptor bleaching was achieved by setting four parallel laser lines of white light laser (560, 568, 576, and 584 nm) to 100% intensity. Donor bleaching was found to be negligible under these conditions. Prebleach and postbleach images were acquired. FRET for the ROIs was observed by an increase in donor (mClover3) fluorescence intensity following the acceptor (mRuby3) photobleaching. FRET efficiency was measured in percent and calculated automatically by Leica LAS AF software as (Dpost − Dpre)/Dpost, where Dpost (Dpre) is the fluorescence intensity of the donor after (before) photobleaching. For each condition, at least three individual experiments from freshly transfected cells were performed.
Statistics—Since the FRET efficiency read-outs were reported as percentages, we opted for a binomial generalized linear model (GLM) followed by an analysis of variance, type III from the ‘car’ package in reference [
30] of the resulting models to compare interactions and negative controls. The data were analyzed with a custom-written R-script [
31].
4. Discussion
In this manuscript, we have identified the α-subunit of the cone-specific heterotrimeric G protein as a protein–protein interaction partner of migratory bird
ErCry4a. Since G proteins are essential components in many signaling pathways, we suggest that this interaction most likely represents the first step in the biochemical reaction cascade underlying light-dependent magnetoreception in night-migratory songbirds. The present investigation is based on two previous studies showing that recombinant
ErCry4a can be purified with a bound FAD chromophore, it exhibits a photo-induced electron transfer pathway, and it is magnetically sensitive in vitro [
19]. Using a yeast two-hybrid screening, Wu et al. [
22] identified six putative protein interaction partners of
ErCry4a, including the α-subunit of the cone-specific G protein. Heterotrimeric G proteins are key proteins in classical signaling cascades [
38,
39], but a direct connection to a migratory bird cryptochrome has never been observed before. We discuss our findings that support the interaction of
ErCry4a with
ErGtα being the primary signaling step in magnetoreception.
First, we show a direct binding of
ErCry4a and
ErGtα on the molecular level and investigated the kinetics of the binding process using SPR. The affinity of myristoylated Gtα/Giα binding to ErCry4a was moderate to high with an apparent K
D = 0.29 ± 0.08 µM (e.g.,
Figure 1A), but the affinity of non-myristoylated Gtα/Giα was about eight-fold higher in the nanomolar range (
Figure 1B). Thus, the binding process occurs with moderate to high affinity and is similar to previous findings for the G protein transducin interacting with rhodopsin [
40,
41,
42,
43]. The results further showed that the myristoyl group in Gtα/Giα decreased the affinity for
ErCry4a. Differences in the observed rate constants point to a plausible explanation for this observation. Dissociation rate constants differ by factors between 9 and 16, but the association rate of
ErCry4a to non-myristoylated Gtα/Giα was 70 to 100-fold higher (compare k
a1 and k
d1 values in the legend of
Figure 1A,B). In a photoreceptor cell, native myristoylated Gtα is fixed to the plane of the membrane by integrating the myristoyl group into the lipid bilayer, leaving the apo-part of the protein freely accessible [
44]. However, our experimental setting does not have a lipid platform to anchor Gtα/Giα via a myristoyl group. It would require a completely different experimental design, which is not realizable in our laboratory at present. Due to the lack of an anchoring lipid platform, we assume that the myristoyl group remains rather flexible near the protein surface in the experiments reported here. This orientation could interfere with the association process leading to a lower association rate constant. In the case of non-myristoylated Gtα/Giα, such a barrier did not exist and access of
ErCry4a was not hindered, resulting in higher association rates. Our results obtained with non-myristoylated Gtα/Giα resemble, therefore, the cellular situation and indicate a high-affinity interaction with
ErCry4a.
Second, we studied the interaction of
ErCry4a and
ErGtα in a cellular environment using transfection of a neuroretinal quail cell line with fluorescently labeled constructs of
ErCry4a and
ErGtα in a FRET analysis. The results in
Figure 4A,B not only confirm the SPR binding studies employing purified proteins but they also further demonstrate that
ErCry4a and
ErGtα interact in living cells. Since the
ErGtα fluorescence constructs are based on the
ErGtα wildtype amino acid sequence and not on the chimeric sequences used for the heterologous expression of
ErGtα in
E. coli, we can also exclude any interferences or false-positive results caused by the insertion in Gtα/Giα. However, we cannot exclude that the interaction kinetics differ in living cells or in vivo.
Third, how does the binding process of Gtα/Giα to
ErCry4a fit into a physiological context? To examine this question, we need to compare our results with the canonical binding of G proteins to opsin receptor molecules. Although no information about binding of European robin G protein to iodopsin or any other opsin is available so far, we refer to information from the well-studied bovine transducin and rhodopsin system. Light-activated rhodopsin shows strong binding to transducin with apparent K
D values in the nanomolar to lower micromolar range [
40,
41,
42,
43]. However, Dell’Orco and Koch (2011) reported in their SPR study a K
D of 0.36 µM for the binding of transducin to dark-adapted rhodopsin [
43], which is similar to the affinity that we observed for the binding of myristoylated Gtα/Giα to
ErCry4a, but lower than the binding of non-myristoylated Gtα/Giα to
ErCry4a. If we assume similar affinities for the interaction of European robin G protein to iodopsin and consider the binding process of
ErCry4a and non-myristoylated Gtα/Giα to reflect the cellular situation in a cone photoreceptor cell, the high affinity of the binding process could well compete with dark-adapted iodopsin for binding to
ErGtα. However, once iodopsin is activated by light, the binding affinity for the G protein very likely increases as this was observed for the binding of bovine transducin to light-activated rhodopsin reaching K
D-values in the lower nanomolar range [
43].
Fourth, are there structural conditions that agree with previous observations? Wu et al. (2020) also reported that the C-terminus of
ErCry4a is not essential for the G
tα/Giα interaction process [
22]. Conducting SPR experiments with a truncated variant of
ErCry4a (
ErCry4a-497) confirmed this result and showed further that G
tα/Giα bound to
ErCry4a-497 with similar affinity (K
D = 0.2 µM,
Figure 1C) in comparison to
ErCry4a wildtype. Interestingly, non-myristoylated G
tα/Giα bound to
ErCry4a-497 with lower affinity (K
D = 0.36 µM) compared to wildtype
ErCry4a. For this, we do not have a clear interpretation but point out that purified
ErCry4a-497 was less stable compared to the wildtype protein leaving some ambiguities about the quantitative kinetic analysis of this variant. However, similar FRET efficiencies between wildtype Gtα and
ErCry4 either with or without the C-terminus in a transiently transfected bird neuroretinal cell line confirm the in vitro data and suggest that these two proteins also interact in living cells and that the C-terminal tail of
ErCry4 is not involved in the interaction (
Figure 4).
Finally, we extended the interaction analysis of myristoylated G
tα/Giα to the Cry forms
ErCry1a and
ErCry1b, which are also expressed in retinal cell types of migratory birds, pointing to putative roles in magnetic sensing [
10,
16,
45,
46]. SPR sensorgrams showed robust binding signals in a concentration-dependent manner (
Figure 5). Interaction analysis revealed affinities with apparent K
D values of 0.4 and 1.53 µM for
ErCry1a and
ErCry1b, respectively. These values are similar to those determined for
ErCry4a and did not disclose a preference of G
tα/Giα for either of them. However, the shape of the SPR sensorgrams indicates differences in the association and dissociation rate constants, which was confirmed by nonlinear curve fitting (main text and legends of
Figure 1 and
Figure 5). Association rate and dissociation rates of
ErCry4a with G
tα/Giα were up to ten-fold lower compared to the rates determined for
ErCry1a and
ErCry1b. One might argue that association rates are different in a cellular environment, when the native myristoylated Gtα attaches to the membrane, allowing diffusion only in a two-dimensional plane and a faster collision with the target. However, once a complex of Gtα with an
ErCry variant forms, the dissociation rate is much lower in the case of
ErCry4a. In other words, a complex of
ErCry4a with G
tα seems more stable compared to a complex of
ErCry1a/b with G
tα. Taking the rate constants into consideration might explain the apparent discrepancy of the present findings with the yeast two-hybrid experiments by Wu et al. (2020), who found no evidence for
GNAT2 (G
tα) interacting with
ErCry1a [
22]. An apparent positive interaction of
ErCry1b with G
tα was observed, but reporter gene expression signals indicating a binding process did not differ from control incubations. Thus, reduced stability of complexes involving
ErCry1a and
ErCry1b could account for the differences in the yeast two-hybrid experiments. Furthermore, binding of G
tα/Giα to different
ErCry isoforms seems to be a new variation of a common theme since G proteins exhibit a multiplicity of G protein-coupling and targeting [
38,
39]. For example, transducin binds to the mammalian rod and cone guanylate cyclase type 1 (GC-E) and to glyceraldehyde-3-phosphate dehydrogenase [
47,
48], two rather different enzymes in different physiological settings.



 ,
, 








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}