
Association of Clonal Hematopoiesis of Indeterminate Potential with Inflammatory Gene Expression in Patients with COPD

 , ,
, ,  , , , , ,
, , , , ,  , add
Show full author list
, add
Show full author list
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. DNA Methylation
2.3. Sample Preparation for NGS and High-Throughput Sequencing
2.4. Metabolomics and Cytokine Analysis
2.5. Cell Culture and Gene Expression Analysis by qPCR
2.6. Human COPD Precision Cut Lung Slides (PCLS)
2.7. Western Blotting
2.8. Statistics
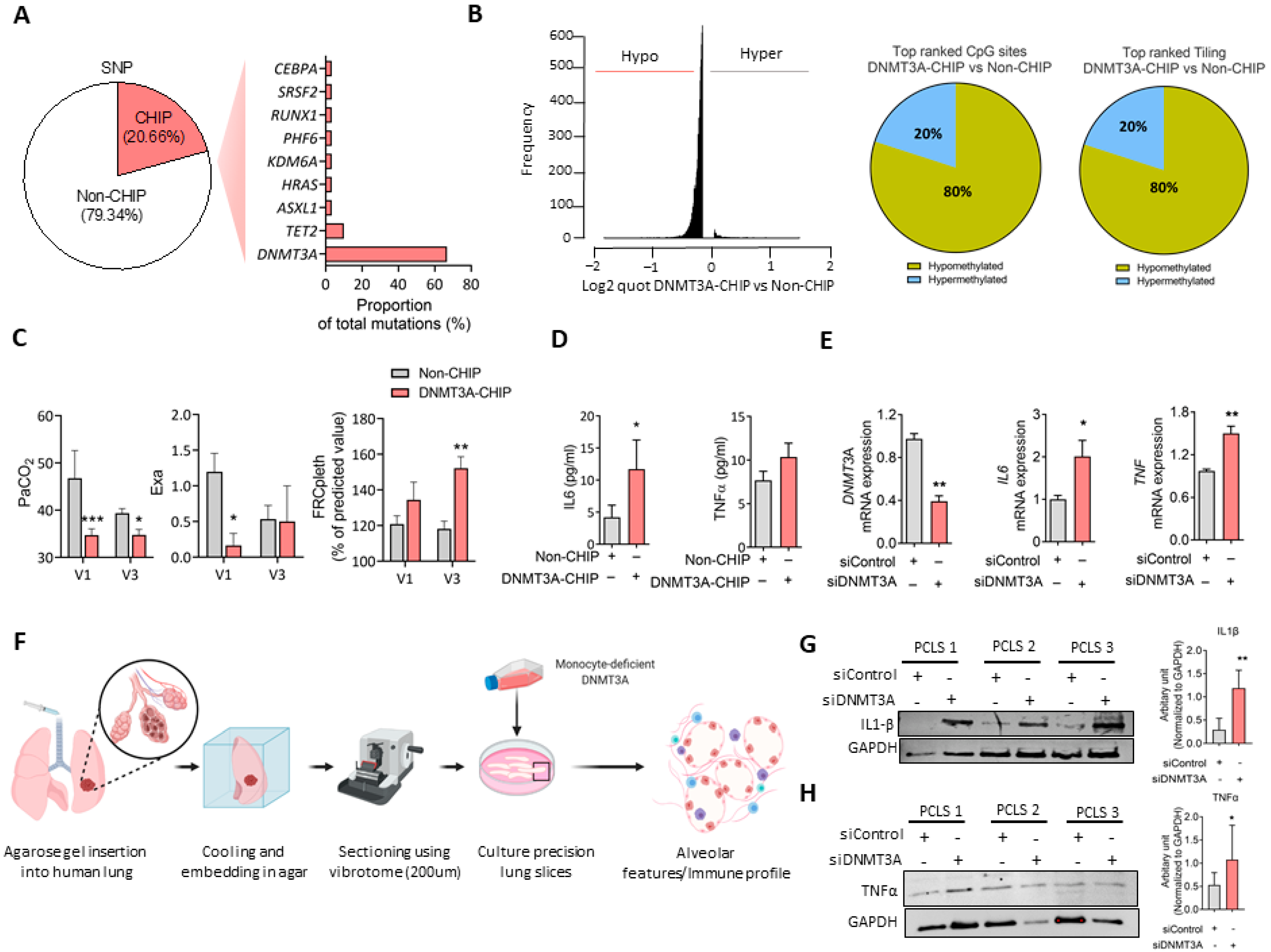
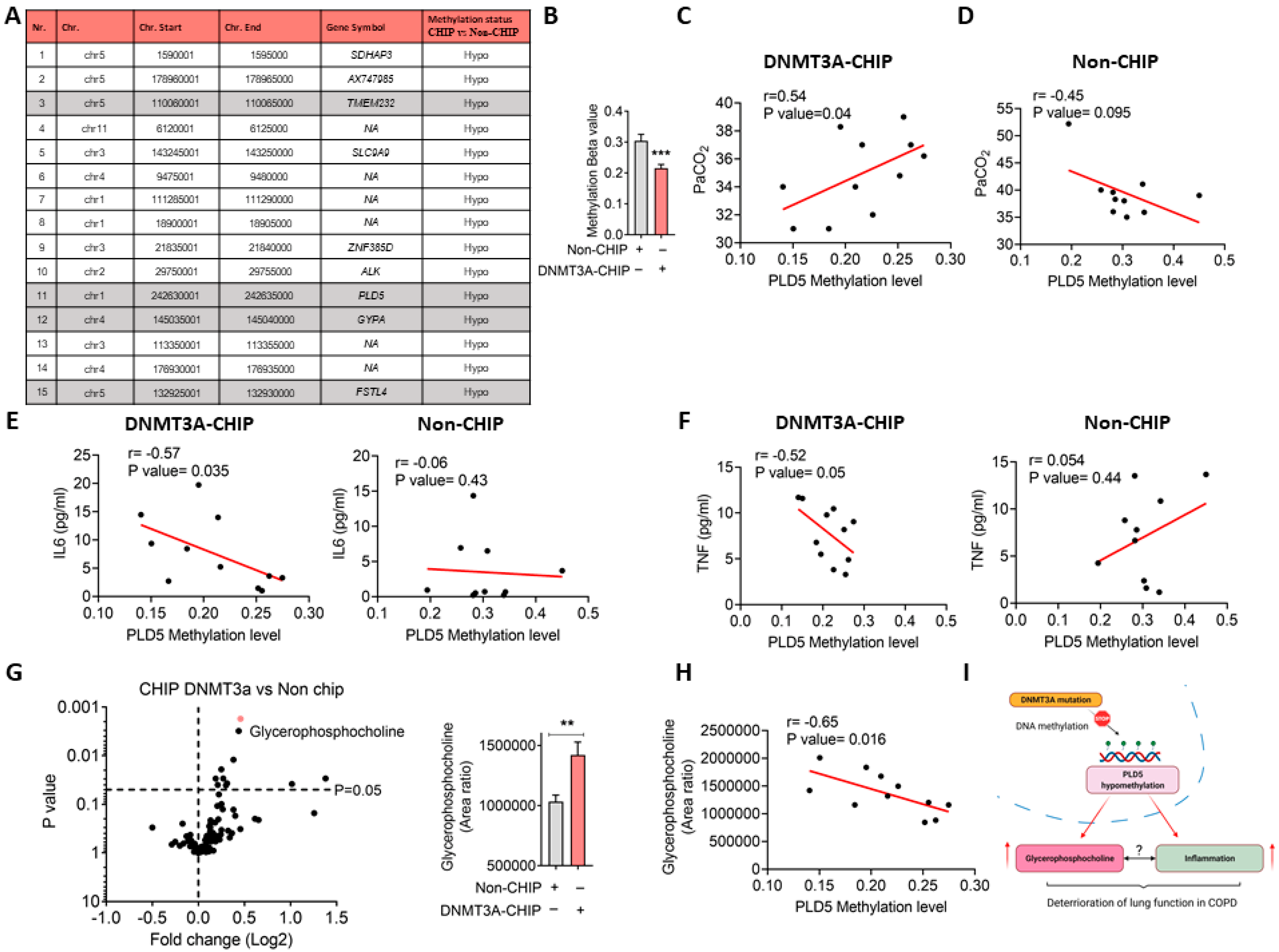
3. Results and Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Jaiswal, S.; Ebert, B.L. Clonal hematopoiesis in human aging and disease. Science 2019, 366, eaan4673. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, S.; Libby, P. Clonal haematopoiesis: Connecting ageing and inflammation in cardiovascular disease. Nat. Rev. Cardiol. 2020, 17, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, S.; Natarajan, P.; Silver, A.J.; Gibson, C.J.; Bick, A.G.; Shvartz, E.; McConkey, M.; Gupta, N.; Gabriel, S.; Ardissino, D.; et al. Clonal Hematopoiesis and Risk of Atherosclerotic Cardiovascular Disease. N. Engl. J. Med. 2017, 377, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Caramori, G.; Adcock, I.M.; Di Stefano, A.; Chung, K.F. Cytokine inhibition in the treatment of COPD. Int. J. Chron. Obstruct. Pulmon. Dis. 2014, 9, 397–412. [Google Scholar] [PubMed] [Green Version]
- Miller, P.G.; Qiao, D.; Rojas-Quintero, J.; Honigberg, M.C.; Sperling, A.S.; Gibson, C.J.; Bick, A.G.; Niroula, A.; McConkey, M.E.; Sandoval, B.; et al. Association of clonal hematopoiesis with chronic obstructive pulmonary disease. Blood 2022, 139, 357–368. [Google Scholar] [CrossRef] [PubMed]
- Quanjer, P.H.; Stanojevic, S.; Cole, T.J.; Baur, X.; Hall, G.L.; Culver, B.H.; Enright, P.L.; Hankinson, J.L.; Ip, M.S.M.; Zheng, J.; et al. Multi-ethnic reference values for spirometry for the 3–95-yr age range: The global lung function 2012 equations. Eur. Respir. J. 2012, 40, 1324–1343. [Google Scholar] [CrossRef] [PubMed]
- Müller, F.; Scherer, M.; Assenov, Y.; Lutsik, P.; Walter, J.; Lengauer, T.; Bock, C. RnBeads 2.0: Comprehensive analysis of DNA methylation data. Genome Biol. 2019, 20, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Preussner, J.; Bayer, J.; Kuenne, C.; Looso, M. ADMIRE: Analysis and visualization of differential methylation in genomic re-gions using the Infinium HumanMethylation450 Assay. Epigenetics Chromatin 2015, 8, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dorsheimer, L.; Assmus, B.; Rasper, T.; Ortmann, C.A.; Ecke, A.; Abou-El-Ardat, K.; Schmid, T.; Brüne, B.; Wagner, S.; Serve, H.; et al. Association of Mutations Contributing to Clonal Hematopoiesis With Prognosis in Chronic Ischemic Heart Failure. JAMA Cardiol. 2019, 4, 25–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pullamsetti, S.S.; Kojonazarov, B.; Storn, S.; Gall, H.; Salazar, Y.; Wolf, J.; Weigert, A.; El-Nikhely, N.; Ghofrani, H.A.; Krombach, G.A.; et al. Lung cancer–associated pulmonary hypertension: Role of microenvironmental inflammation based on tumor cell–immune cell cross-talk. Sci. Transl. Med. 2017, 9, eaai9048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Celarain, N.; Tomas-Roig, J. Aberrant DNA methylation profile exacerbates inflammation and neurodegeneration in multi-ple sclerosis patients. J. Neuroinflamm. 2020, 17, 21. [Google Scholar] [CrossRef] [PubMed]
- Colombo, S.; Martín-Sierra, C.; Melo, T.; Laranjeira, P.; Paiva, A.; Domingues, P.; Domingues, M.R. Modulation of the inflammatory response of immune cells in human peripheral blood by oxidized arachidonoyl aminophospholipids. Arch. Biochem. Biophys. 2018, 660, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Pniewska, E.; Pawliczak, R. The Involvement of Phospholipases A2 in Asthma and Chronic Obstructive Pulmonary Disease. Mediat. Inflamm. 2013, 2013, 793505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tombor, L.S.; John, D.; Glaser, S.F.; Luxán, G.; Forte, E.; Furtado, M.; Rosenthal, N.; Baumgarten, N.; Schulz, M.H.; Wittig, J.; et al. Single cell sequencing reveals endothelial plasticity with transient mesenchymal activation after myocardial infarction. Nat. Commun. 2021, 12, 681. [Google Scholar] [CrossRef]
- Mas-Peiro, S.; Hoffmann, J.; Fichtlscherer, S.; Dorsheimer, L.; Rieger, M.A.; Dimmeler, S.; Vasa-Nicotera, M.; Zeiher, A.M. Clonal haematopoiesis in patients with degenerative aortic valve stenosis undergoing transcatheter aortic valve implantation. Eur. Heart J. 2020, 41, 933–939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ortmann, C.A.; Dorsheimer, L.; Abou-El-Ardat, K.; Hoffrichter, J.; Assmus, B.; Bonig, H.; Scholz, A.; Pfeifer, H.; Martin, H.; Schmid, T.; et al. Functional Dominance of CHIP-Mutated Hematopoietic Stem Cells in Patients Undergoing Autologous Transplantation. Cell Rep. 2019, 27, 2022–2028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarode, P.; Zheng, X.; Giotopoulou, G.A.; Weigert, A.; Kuenne, C.; Günther, S.; Friedrich, A.; Gattenlöhner, S.; Stiewe, T.; Brüne, B.; et al. Reprogramming of tumor-associated macrophages by targeting β-catenin/FOSL2/ARID5A signaling: A potential treatment of lung cancer. Sci. Adv. 2020, 6, 6105. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Weigert, A.; Reu, S.; Guenther, S.; Mansouri, S.; Bassaly, B.; Gattenlöhner, S.; Grimminger, F.; Pullamsetti, S.; Seeger, W.; et al. Spatial Density and Distribution of Tumor-Associated Macrophages Predict Survival in Non-Small Cell Lung Carcinoma. Cancer Res. 2020, 80, 4414–4425. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Subset | GOLD 0 Patients | GOLD I–III Patients | GOLD III/IV Patients | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| CHIP | All | Negative | Positive | All | Negative | Positive | All | Negative | Positive | |
| Number | 26 | 20 | 6 | 90 | 71 | 19 | 36 | 26 | 10 | |
| Sex | Male | 42% | 30% | 83% | 68% | 69% | 63% | 61% | 58% | 70% |
| Female | 58% | 70% | 17% | 32% | 31% | 37% | 39% | 42% | 30% | |
| Age | 60 (5) | 59 (4.7) | 62 (5.6) | 62 (5.5) | 62 (5.8) | 62 (4.4) | 62 (5.7) | 62 (6.1) | 62 (4.5) | |
| BMI | 27 (4.8) | 26 (5.4) | 28 (2.1) | 27 (5.8) | 27 (6.2) | 26 (4.1) | 26 (4.7) | 27 (5.2) | 25 (3.2) | |
| Smoking | Packyears | 37 (19) | 34 (17) | 46 (26) | 42 (31) | 41 (31) | 46 (29) | 48 (29) | 48 (28) | 50 (33) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuhnert, S.; Mansouri, S.; Rieger, M.A.; Savai, R.; Avci, E.; Díaz-Piña, G.; Padmasekar, M.; Looso, M.; Hadzic, S.; Acker, T.; et al. Association of Clonal Hematopoiesis of Indeterminate Potential with Inflammatory Gene Expression in Patients with COPD. Cells 2022, 11, 2121. https://doi.org/10.3390/cells11132121
Kuhnert S, Mansouri S, Rieger MA, Savai R, Avci E, Díaz-Piña G, Padmasekar M, Looso M, Hadzic S, Acker T, et al. Association of Clonal Hematopoiesis of Indeterminate Potential with Inflammatory Gene Expression in Patients with COPD. Cells. 2022; 11(13):2121. https://doi.org/10.3390/cells11132121
Chicago/Turabian StyleKuhnert, Stefan, Siavash Mansouri, Michael A. Rieger, Rajkumar Savai, Edibe Avci, Gabriela Díaz-Piña, Manju Padmasekar, Mario Looso, Stefan Hadzic, Till Acker, and et al. 2022. "Association of Clonal Hematopoiesis of Indeterminate Potential with Inflammatory Gene Expression in Patients with COPD" Cells 11, no. 13: 2121. https://doi.org/10.3390/cells11132121
APA StyleKuhnert, S., Mansouri, S., Rieger, M. A., Savai, R., Avci, E., Díaz-Piña, G., Padmasekar, M., Looso, M., Hadzic, S., Acker, T., Klatt, S., Wilhelm, J., Fleming, I., Sommer, N., Weissmann, N., Vogelmeier, C., Bals, R., Zeiher, A., Dimmeler, S., ... Pullamsetti, S. S. (2022). Association of Clonal Hematopoiesis of Indeterminate Potential with Inflammatory Gene Expression in Patients with COPD. Cells, 11(13), 2121. https://doi.org/10.3390/cells11132121