Abstract
Interferon regulatory factor 8 (IRF8) is a transcription factor of the IRF protein family. IRF8 was originally identified as an essentialfactor for myeloid cell lineage commitment and differentiation. Deletion of Irf8 leads to massive accumulation of CD11b+Gr1+ immature myeloid cells (IMCs), particularly the CD11b+Ly6Chi/+Ly6G− polymorphonuclear myeloid-derived suppressor cell-like cells (PMN-MDSCs). Under pathological conditions such as cancer, Irf8 is silenced by its promoter DNA hypermethylation, resulting in accumulation of PMN-MDSCs and CD11b+ Ly6G+Ly6Clo monocytic MDSCs (M-MDSCs) in mice. IRF8 is often silenced in MDSCs in human cancer patients. MDSCs are heterogeneous populations of immune suppressive cells that suppress T and NK cell activity to promote tumor immune evasion and produce growth factors to exert direct tumor-promoting activity. Emerging experimental data reveals that IRF8 is also expressed in non-hematopoietic cells. Epithelial cell-expressed IRF8 regulates apoptosis and represses Osteopontin (OPN). Human tumor cells may use the IRF8 promoter DNA methylation as a mechanism to repress IRF8 expression to advance cancer through acquiring apoptosis resistance and OPN up-regulation. Elevated OPN engages CD44 to suppress T cell activation and promote tumor cell stemness to advance cancer. IRF8 thus is a transcription factor that regulates both the immune and non-immune components in human health and diseases.
1. Introduction
Interferon regulatory factor 8 (IRF8), originally termed interferon consensus sequence binding protein (ICSBP), is a member of the IRF transcription factor [1]. IRFs were first identified as a regulator of the type I interferon (IFN-I) response for the activation of IFN-stimulated genes that are essential for immune response to viruses and other pathogens [2,3,4,5], and are now known to be important in turning pathogen associated molecular patterns into chromatin changes and eventually into immune cell activation [6]. IRF8 was first cloned as an IFNγ-inducible nuclear protein-encoding gene that binds to specific IFN-responsive DNA motif in the major histocompatibility complex class I (MHC I) genes [7]. IRF8 has since been determined to be constitutively expressed and IFNγ inducible and plays key roles in the IFN response pathways in immune cell differentiation and function, as well as in non-hematopoietic cell turnover and pathogenesis [8,9,10,11,12,13,14,15,16,17,18,19,20,21].
2. IRF8 Function and Diseases
A 915C> T mutation (R294C) resembles IRF8 KO mice in accumulating immature myeloid cells (IMCs) and causes susceptibility to infection in mice [22,23]. K108E and T80A mutations results in IRF8 loss of function that leads to impaired dendritic cell monocyte development and function in humans or mice [9,24]. A 331C>T [R111* (stop codon)] mutation may cause IRF8 loss of function, resulting in neutrophilia, monocytopenia and decreased CD3+ T cell and CD8+ T cell counts in humans [25]. IRF-8 polymorphisms have been implicated in the development of autoimmune thyroiditis, Behcet’s disease and, increased susceptibility to tuberculosis (TB). The IRF-8 polymorphism, rs17445836, was associated with both development of autoimmune thyroiditis and TB [26,27,28]. Moreover, IRF8 also functions as an apoptosis regulator in hematopoietic tumor cells [29,30,31,32]. IRF8’s intrinsic function in hematopoietic cells thus plays essential roles in myelopoiesis to regulate immune cell activation and immune suppressors under physiological conditions [20,33,34] and in suppression of tumorigenesis [29,30,31,32]. Indeed, IRF8 has been shown to function as a suppressor of acute myeloid leukemia (AML) [35]. However, IRF8 has recently been reported to function as an acute myeloid leukemia (AML)-specific susceptibility factor and promote AML cell proliferation. High IRF8 expression is associated with poorer prognoses in certain AML patients [36]. On the other hand, IRF8 is also expressed in non-hematopoietic cells [8,10,37,38,39,40,41,42,43,44,45]. IRF8 functions as an apoptosis regulator in several types of human tumor cells [38,41,46,47] and mice with IRF8 deletion only in the colon epithelial cells develop significantly more colon tumors than the littermate control mice [41]. Furthermore, IRF8 expression level is positively linked to cancer patient prognosis and response to immunotherapy [18,19]. IRF8 therefore also functions as a tumor suppressor in non-hematopoietic cells. In this review, we discuss the function of IRF8 in hematopoietic and non-hematopoietic cell populations and the role of IRF8 in antitumor immune responses.
3. IRF8 Function as a Transcription Factor That Depends on IAD-Interacting Transcription Factors to Exert Its Activity
Analysis and comparison of the IRF8 protein sequences in the National Center for Biotechnology Information with the Blast protein program [48] revealed that IRF8 proteins are highly conserved in mammals, with an 89.44–89.67% similarity in amino acid sequences between human and mouse IRF8 proteins. Human IRF8 gene is located at chromosome 16 with 23,228 bp that encodes 9 exons and 8 introns (Figure 1). The human IRF8 protein is 426 amino acids long with a DNA-binding domain (DBD) and an IRF association domain (IAD) [23]. The DBD binds to unique consensus sequence motifs at promoters of genes, such as IFN-responsive genes, to regulate transcription. The IAD associates with other transcription factors to direct the IRF8 protein complex binding to specific DNA motif to regulate specific gene transcription (Figure 1). IRF8 has weak DNA-binding affinity and its transcription regulatory activity therefore depends on its IAD association with other transcriptional factors to form a transcription complex which renders IRF8 specific and high affinity to bind to the unique DNA motifs at gene promoters [23,34,49,50]. IRF8 functions as either a transcriptional activator or repressor and it is the IAD-associated transcription factor(s) that dictates IRF8 bindingand transcriptional activity [20,23,33,51,52,53,54,55].
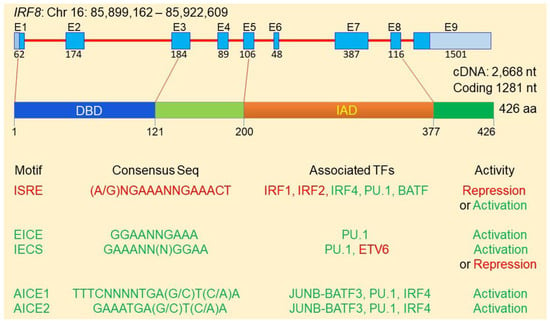
Figure 1.
The IRF8 protein structure and transcription regulatory network.
IRF8 transcription factor protein complexes selectively binds to certain types of DNA consensus sequence motifs to activate or repress gene transcription [33,56,57,58,59]. Heterodimers of IRF8 in association with IRFs (e.g., IRF1 or IRF2) bind to the IFN-stimulated response element (ISRE: [(A/G)NGAAANNGAAACT]. IRF8-IRF1/IRF2 heterodimers generally bind to promoter region ISRE to repress the transcription of the downstream genes (Figure 1) [1]. The IRF8-Ets (Erythroblast Transformation Specific/E-twenty-six) and IRF8-PU1 heterodimers bind to the Ets-RF8 composite element (EICE: GGAANNGAAA) or the IRF8-Ets composite sequence (IECS: GAAANN(N)GGAA) to activate target gene transcription [52,53,56,57,58,60,61,62,63,64,65,66]. In addition, IRF8 has been shown to form a complex with the JUNB/AP-1-BATF3and/or IRF4 and binds to the AP-1-IRFcomposite elements 1 and 2 [AICE1:TTTCNNNNTGA(G/C)T(C/A)A; AICE2: GAAATGA(G/C)T(C/A)A] to activate transcription of genes [11,67,68,69]. For example, IRF8 forms a heterodimer with AP-1 factor BATF and binds to the AICE site to promote gene activation in T helper 17 (Th17) cells, B and dendritic cells [67,69,70]. It has been shown that IRF8 also forms complexes with IRF4, PU.1 and BATF at the ISRE-like sites to activate Il9 and Il21 transcription in Th9 cells [66]. IRF8 can also form a heterodimer with IRF1 or IRF2 [71] that binds to ISRE element to repress transcription of genes induced by IFN and retinoic acid [72,73] or to activate target gene expression [50,70,71]. Although IRF8 binds to IECS to activate gene expression [1,54], IRF8 binding to IECS element at the Asah1 promoter represses acid ceramidase expression [31]. Furthermore, IRF6 interacts with ETV6 to repress Il4 transcription in Th9 cells [66].
4. IRF8 Expression Profiles
The expression of IRF8 was originally detected in hematopoietic cells and its expression has long been thought to be restricted to hematopoietic cells [56]. Although IRF8 is abundantly expressed in hematopoietic cells, particularly in B cells and DCs, it has since been determined that IRF8 is also expressed in epithelial cells in the intestine, colon, ocular lens, skin, lung, liver, and heart [8,10,37,38,39,40,41,42,43,44,45]. Analysis of IRF8 expression from single cell RNA sequencing datasets revealed that IRF8 is indeed expressed in several types of non-hematopoietic cells, including theca cells, melanocytes, enterocytes and intestinal cells in humans (Figure 2).
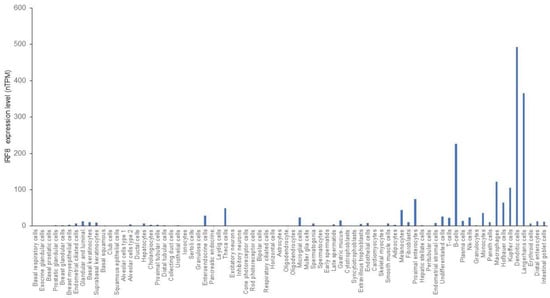
Figure 2.
IRF8 expression profiles in human cells. Single cell RNA sequencing datasets were adapted from the human protein atlas.
5. IRF8 Function in Hematopoietic Cells
In hematopoietic cells, the highest IRF8 expression is in the progenitors and mature cells of the B cell, cDC1, and pDC lineages [1,20,74,75]. Consistent with the high level of IRF8 expression in these cell lineages, IRF8 is essential for the development of monocytes and DCs (Figure 3). IRF8 may also regulate myeloid cell lineage differentiation indirectly through regulating GM-CSF expression in T cells [76]. Loss of IRF8 expression or function often leads to impaired immune response to infection and hematopoietic malignancies [9,22,29,30,31,77,78]. IRF8 deficiency results in a severe immunodeficiency humans as characterized by susceptibility to infections due to loss of DC subsets, CD14+ and CD16+ monocytes and a decreased level of NK cells with reduced activity [70]. A genome-wide ChIP-Seq analysis identified 319 IRF8-binding sites at the gene promoter regions with an EICE-like motif in macrophages stimulated with IFNγ/CpG. Analysis of these IRF8 target genes in Mycobacterium tuberculosis-infected lungs revealed specific enrichment of pathways/genes in antigen processing and presenting, indicating a key role of IRF8 function in myeloid cells in regulating early response to infection [79]. De novo DNA motif analysis using IRF8 ChIP-seq identified 89 genes with an AICE-dependent gene program that requires high levels of IRF8 in cDC1 [75]. These findings indicate that IRF8 directly regulates a large set of genes in myeloid cells.
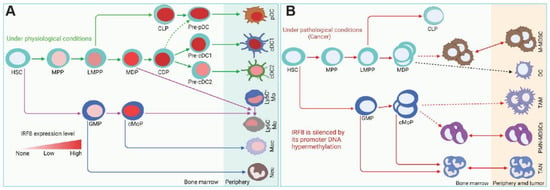
Figure 3.
IRF8 functions under physiological and pathological conditions. (A). IRF8 regulation of myeloid cell lineage differentiation under physiological conditions. (B). Under pathological conditions such as cancer, IRF8 is silenced in myeloid cells, resulting in accumulation of immature myeloid cells and neutrophil. HSC: hematopoietic stem cell, MPP: multipotent progenitors, LMPP: lymphoid-primed multipotent progenitors, MDP: macrophage DC progenitors, CDP: common DC progenitors, CLP: common lymphoid progenitors, pDC: plasmacytoid DC, cDC1: conventional DC1, cDC2: conventional DC2, GMP: granulocyte-monocyte progenitor, cMoP: common monocyte progenitor, M-MDSC: monocytic-MDSC, PMN-MDSC: polymorphonuclear MDSC TAM: tumor-associated macrophage, TAN: tumor-associated neutrophils.
IRF8 also plays a key role in regulating Th1 polarization of early immune response following infection. IRF8-deficient mice exhibit diminished production of IL-12p40, lack of Th1 polarization [1,24,70]. Mice with deletion of the Irf8 gene exhibit a significantly reduced number of pre-pro-B cells. Hematopoietic stem cells of IRF8 KO mice are deficient in differentiation to B220+ B-lineage cells and this defect can be corrected by adoptive transfer of bone marrow cells of WT bone marrow cells [80,81,82]. IRF8 therefore not only plays an essential role in the linage differentiation of innate immune cells but also plays a key role in the development of the adaptive immune cells.
5.1. Dendritic Cells
Dendritic cells (DCs) are found in most tissues and classically function to activate antigen-specific naïve T cells. Discoveries of different transcription factors and surface markers allow for distinction between DCs and other immune cells as well as distinction between different DC subsets. DCs are divided into either conventional (cDC1 and cDC2) or plasmacytoid (pDC). Conventional DCs prime naïve T cells according to their surface markers (cDC1 cells present to CD8+ T cells and cDC2 cells present to CD4+ T cells) while pDCs produce large quantities of Type I interferons [83,84] and their antigen-presenting ability is likely less efficient than that of conventional DCs [85]. Much of the research regarding IRF8 in the context of DCs is directed at understanding its role in differentiation [86,87,88].
In the early stages of DC differentiation, IRF8 is required for the transition from macrophage and DC precursor (MDP) to common DC precursor (CDP) [89,90]. Not all the literature is in agreement about the point at which IRF8 becomes necessary for DC differentiation. Another study found that at an earlier lymphoid primed multipotent progenitor (LMPP) stage, IRF8 expression biased differentiation towards DCs by making DC-lineage genes more accessible [91]. When IRF8 was absent from MDPs, differentiation was skewed in favor of a macrophage phenotype rather than a DC phenotype [90]. Additionally, IRF8 was required for differentiation into cDC1 cells but not required for the differentiation of cDC2 cells from the CDP stage [89]. This indicates that IRF8 is necessary for commitment to the DC lineage but is more involved in cDC1 speciation than cDC2 speciation. As for pDCs, IRF8 was not necessary for their differentiation from CDPs. However, IRF8 KO mice produced less Type I IFN in response to stimuli indicating that IRF8 might play a larger role in the function of pDCs than in their development [89].
Recently there has been increasing evidence for an association between IRF8 and dendritic cell deficiencies leading to diseases such as chronic myelogenous leukemia [92], atherosclerosis [93], and cancers [42,94]. For example, IRF8-deficient mice display a CML- like phenotype dominated by expansion of granulocyte precursors and lack both CD8α+ dendritic cells (DCs) and plasmacytoid DCs (pDCs) [70]. An autosomal recessive IRF8 deficiency, K108E, was associated with a complete lack of dendritic cells and resulted in a severe immunodeficiency [9]. Upon analysis of this patient’s blood, the IRF8-deficient dendritic cells exhibited defective antigen presentation and anergic T cells. IFR8 is responsible for survival and function of terminally differentiated cDC1s. It has been shown that increased IRF8 expression in CD11b−CD103− DN DC progenitor cells promotes differentiation into cDC1s to enhance CAR-T cell efficacy [95]. Because IRF8 is necessary for DC differentiation and adequate functioning, the loss of IRF8 may provide an immunodeficient environment contributing to disease pathogenesis and progression.
5.2. Macrophages
IRF8 is involved in the differentiation of macrophages. Myeloid progenitor cells established from IRF8-deficient mice were able to differentiate into functional macrophages and express macrophage-specific genes upon retroviral transduction of IRF8 [96]. In contrast, transduction of IRF8 resulted in a decrease in granulocytic differentiation and repression of granulocytic differentiation genes. Together, this suggests that IRF8 promotes macrophage differentiation over granulocytic differentiation from myeloid progenitor cells. IRF8’s role in macrophage differentiation was also studied in a zebrafish embryogenesis model [97]. IRF8 knockout models showed a depletion of macrophages, an increase in neutrophils (a subset of granulocytes), but no difference in myeloid progenitors. Upon overexpression of IRF8, macrophage differentiation was promoted while neutrophil differentiation was suppressed. This model further supports the ability of IRF8 to promote macrophage differentiation over granulocyte differentiation.
IRF8 also appears to be involved in the homeostasis and differentiation of tissue macrophages including osteoclasts [13,17,98,99]. In IRF8-deficient mice, macrophage morphology and cell numbers were assessed. IRF8-deficient macrophages from the brain, liver, and kidney exhibited changes in their population sizes; in brain and liver macrophages, cell morphology was changed as well. For macrophages to maintain their functionality, different genes related to immune functioning must be expressed. In IRF8-deficient macrophages, a downregulation of immune-related genes like toll-like receptor 12 and major histocompatibility complex class II was observed. This study indicates that IRF8 plays a role in maintaining the morphology, population size, and gene profile of tissue macrophages [98].
IRF8 regulates macrophage functioning as part of the innate immune system. It promotes transcription of pro-inflammatory cytokines such as IL-12, affects macrophage responses to LPS stimulation, and determines macrophage response to intracellular and engulfed pathogens. In IRF8-deficient mice, increased susceptibility to intracellular infection was due to impaired synthesis of IL-12 [100]. Co-transfection with IRF8 and IRF1 was found to act synergistically to activate the IL-12 promoter as determined by a luciferase reporter gene activity [101]. Additionally, IRF8 and IRF1 were found to act through binding to an IRSE-like sequence within the IL-12 promoter. Taken together, IRF8 (acting synergistically with IRF1) activates the IL-12 promoter through binding with an IRSE-like sequence. Enhanced IL-12 allows for more effective resistance to intracellular infection.
In addition to IL-12 production, other IRF8-mediated macrophage responses to bacterial infection include autophagy and inflammasome activation. IRF8 was found to activate genes involved in autophagosome formation and lysosome fusion [102]. As such, in IRF8-deficient macrophages, activation of autophagy genes and pathogen degradation was defective and growth of the facultative intracellular pathogen, Listeria monocytogenes, was unhindered. Similarly, IRF8 induced Nramp1 (cation transporter involved in phagosome functioning) expression through activation of its promoter [103]. Decreased clearance of intraphagosomal pathogens was observed in macrophages deficient in IRF8 or Nramp1, suggesting that IRF8 induces Nramp1 thereby facilitating phagosome degradation of pathogens. Within macrophages, IRF8 was found to promote NLRC4 inflammasome activation by enhancing the transcription of Naips [104]. Additionally, IRF8 was found to protect against S. typhimurium and B. thailandensis bacterial infection in vivo likely due to inflammasome-dependent cytokines and pyroptosis. These studies highlight the role of IRF8 in the macrophage response to pathogens.
5.3. Monocytes
Differentiation in monocytes is regulated by IRF8. In one study, granulocyte progenitors and monocyte progenitors were able to be separated from their common granulocyte/monocyte progenitor through cell sorting [105]. Higher IRF8 expression was observed in the lineage-committed cell lines rather than in their common progenitor. IRF8 KO in vitro demonstrated that differentiation of granulocyte progenitors and monocyte progenitors from their common granulocyte/monocyte progenitor was not affected by a lack of IRF8, indicating that IRF8 does not work at the level of granulocyte/monocyte progenitors, but is rather involved in lineage-specific differentiation. Another study found that in IRF8 KO mice, mononuclear phagocyte progenitor cells were unable to give rise to monocytes and instead gave rise to neutrophils [34]. Mechanistically, IRF8 was found to interact with transcription factor C/EBPα and block its ability to stimulate transcription of the genes necessary for neutrophil differentiation, thereby suppressing the neutrophil phenotype. Indeed, a previous study indicated that IRF8 KO mice were found to lack inflammatory Ly6C+ monocytes [57]. Similarly, another group found that IRF8-deficient mice infected with West Nile virus exhibited reduced numbers of peripheral Ly6C+ monocytes with diminished migratory capabilities [106]. Taking the results of these studies together, IRF8 appears to promote monocyte differentiation into specific subsets and suppress neutrophil differentiation.
IRF8 expression and its subsequent role in monocyte differentiation appear to be regulated by the signal transducer and activator of transcription (STAT) protein family, specifically STAT1 [107] and STAT5 [108]. When all-trans retinoic acid (ATRA), which promotes myelomonocytic differentiation, was given to human monoblastic cells, up-regulation of IRF8 (and other differentiation transcription factors) was impaired in cells expressing mutant STAT1 and monocytic inhibition was impaired [107]. This indicates that STAT1 is involved in the upregulation of IRF8 in monocytes thereby contributing to differentiation. In a study on the effects of mTOR on monocyte differentiation, blocking STAT5 activity was found to increase IRF8 expression and increase the differentiation in mTOR-deficient granulocyte/monocyte precursors [108]. This study suggests that STAT5 negatively regulates IRF8 expression. While it is known that ATRA induces STAT expression to promote differentiation in myeloid leukemia [109], one such mechanism may be through the regulation of IRF8 expression.
5.4. IMCs/Myeloid-Derived Suppressor Cells (MDSCs)
A major phenotype of IRF8 knock out (IRF8 KO) mice or mice with IRF8 deficiency is accumulation of CD11b+Gr1+ IMCs [23,110] that phenotypically and functionally resembles tumor-induced CD11b+Gr1+ myeloid-derived suppressor cells (MDSCs) [32,111]. Among the MDSC subtypes, CD11b+Ly6Chi/+Ly6G– polymorphonuclear MDSCs (PMN-MDSCs) is dramatically increased in IRF8 KO mice. Although the percentages of monocytic MDSCs (M-MDSCs) were similar to the WT mice, there is a significant increases in the total numbers of both PMN-MDSCs and M-MDSCs in IRF8 KO mice [111]. In tumor-bearing mice, IRF8 expression level is silenced in both PMN-MDSCs and M-MDSCs [111]. IRF8 is also down-regulated in MDSCs of human cancer patients [32,112]. IRF8 expression can be repressed by its promoter DNA methylation [41]. IRF8 expression is also regulated by micro-RNA, phosphorylation, and ubiquitination [113,114,115]. Under non-pathological conditions, immature myeloid cells differentiate into granulocytes, macrophages, and DCs. However, pathological conditions such as cancer can block their differentiation and increase the expression of immune-suppressive factors. The resulting cells are considered MDSCs [116,117,118,119].
Between the two subsets of MDSCs, PMN-MDSCs exhibit more extensive expansion in cancer when compared to M-MDSCs [120,121], and a decrease in IRF8 has been associated with an increase in MDSC frequency in cancer [111]. Recent research supports IRF8’s role in inhibiting the formation of MDSCs. Tumor growth in mice was associated with the selective expansion of low-IRF8 granulocyte progenitors (GPs). Importantly, tumor-derived granulocyte progenitors exhibited an increased ability to form PMN-MDSCs [122]. When gene expression profiles were compared between tumor-derived GPs and IRF8 KO GPs, shared expression profiles indicated that loss of IRF8 may underlie the formation of GPs and subsequent PMN-MDSC formation in tumors. Further supporting MDSC suppression by IRF8, IRF8-deficient mice generated MDSC-like populations highly homologous to tumor-induced MDSCs while IRF8 overexpression attenuated MDSC accumulation [111].
While IRF8 inhibits the formation of MDSCs, MDSCs (and other factors) also negatively regulate the expression of IRF8. Chronic inflammation, such as in the setting of ulcerative colitis, increases the population of MDSCs. MDSCs express large amounts of IL-10 which activates transcription factor STAT3. Through the activation of DNA methyltransferases, IRF8 is functionally silenced [41]. Taken together, this suggests that MDSC elevation can silence IRF8 expression consequently increasing the expansion of MDSCs. Decreased IRF8 in the context of MDSCs promotes their survival through evasion of cytotoxic T lymphocyte (CTL) elimination. MDSCs can alter the expression of apoptotic proteins Bax and Bcl-xl and deregulate the Fas-mediated apoptosis program utilized by CTLs by suppressing IRF8 [32]. As a result, MDSCs can evade immune clearance [117,123].
5.5. B Cells
Originally IRF8 was found to be expressed in mature mouse B cells and B cell precursors (pro and pre-B cells), while not expressed in tumors composed of mature plasma cells [124]. Further research has supported the differential expression of IRF8 between B cells and terminally differentiated B cells, known as plasma cells, and shown that IRF8 promotes germinal center (GC) formation while altering B cell population [82,125,126,127,128,129]. IRF8 promotes the GC B cell response while inhibiting plasma cell differentiation through the induction of genes that maintain the B cell program and repression of genes that maintain the plasma cell program [130]. Actions that promote the GC B cell response such as dampening of BCR signaling, facilitation of B cell-T cell interactions, and promotion of affinity maturation are regulated by IRF8. In addition to promoting the GC B cell response, IRF8 also antagonizes the IRF4-driven differentiation of plasma cells [81]. Double deletion of both IRF8 and PU.1 led to enhancement in the rate of class switch recombination and plasma cell differentiation further supporting the role of IRF8 in the negative regulation of plasma cell differentiation. It was recently found that mice with deletion of irf8 in B cells have abundance of regulatory B cell (i35-Bregs) [131].
IRF8 has been implicated in diffuse large B cell lymphoma (DLBCL) and pre-B cell acute lymphoblastic leukemia (Pre-B ALL) [132,133,134]. In DLBCL, IRF8 was discovered as an immunoglobulin heavy chain (IGH) fusion partner. Expectedly, the lymphomas with this rearrangement exhibited markedly higher IRF8 expression. IRF8-regulated actions such as induction of genes that promote B cell formation, suppression of plasma cell formation, and promotion of apoptosis resistance likely contribute to the formation of DLBCL [132]. This is in agreement with earlier research suggesting that IRF8 decreases the expression of p53 and p21 facilitating an increase in apoptosis resistance [135]. A proportion of mice double deficient in PU.1 and IRF8 developed Pre-B ALL. It was shown that IRF8, PU.1, and IRF4 bound to the promoter/enhancer regions of tumor suppressor genes, such as Ikaros and Spi-B, and in IRF-deficient samples, the expression of these genes was significantly reduced [133]. While it appears that IRF8 protects against the formation of Pre-B ALL, drastically increased IRF8 (such as in the case of IGH-IRF8 fusion) may promote the formation of DLBCL.
5.6. T Cells
Much of the research on IRF8 function in T cells focuses on its role in T cell differentiation. T Helper 17 (Th17) cells are a subset of CD4+ Helper T cells named for their secretion of cytokine IL-17. IRF8 has been reported to suppress the expansion of Th17 cells [8,136,137] although not all literature is in agreement with this [138]. IRF8 appears to inhibit the expression of RORγt, a crucial transcription factor for Th17 cell differentiation [136,139,140]. Additionally, overexpression of IRF8 suppressed activity at a RORγt-dependent enhancer required for IL-17 expression [137].
IRF8 involvement in the differentiation of other T cell lineages has also been investigated. IRF8 may facilitate the differentiation of T Helper 1 (Th1) cells over T Helper 2 (Th2) cells. IRF8 and IRF1 expression were higher in Th1 polarized cells versus Th2 polarized cells while IRF4 expression was higher in Th2 polarized cells [141]. This implicates the differential expression of IRFs in Helper T cell differentiation with IRF8 being most likely involved in Th1 differentiation. This is congruent with earlier research finding that IRF8 was required for adequate Th1-induced macrophage activation [142]. IL-9 producing Helper T (Th9) cells exert an anti-tumor effect through the secretion of IL-9 and IL-21 [143,144,145]. IRF8 was shown to be required for the differentiation of Th9 cells through binding to Th9 related genes and suppressing the secretion of IL-4 [66].
IRF8 has a relatively unclear role in CD8+ T cells. IRF8 may restrain the expansion of CD8+ T cells. In a mouse model of ocular herpes simplex 1 virus (HSV-1) infection, IRF8 KO mice exhibited marked expansion of HSV-1 specific CD8+ T cells with increased infiltration, inflammation, and viral clearance [40]. This suggests that IRF8 is involved in the suppression of CD8+ T cells and dampening of the associated inflammation. However, a previous study found that depletion of IRF8 decreases differentiation of CD8+ T cells into effector T cells in a mouse model of Graft Versus Host Disease [146].
IRF8 suppresses inflammation and is involved in tumor formation. A decrease in IRF8 increased inflammation in a mouse model of colitis possibly due to the associated increase in Th17 cells [137]. A loss of IRF8 also resulted in increased inflammation in autoimmune uveitis (associated with a significant increase in Th17 cells) and Ocular HSV-1 infection (possibly due to a decreased limitation on CD8+ expansion) [8,40]. Decreased IRF8 expression in CD8+ T cells has been associated with an increase in melanoma growth while an increase in IRF8 has been reported in DLBCL tumor tissues [66,136]. In DLBCL, IRF8-induced reduction of Th17 expansion may contribute to tumor formation.
5.7. Natural Killer Cells
IRF8 is involved in the development and function of Natural Killer (NK) cells [147]. Biallelic mutations in IRF8 resulted in impaired NK cell function, numbers, and maturation [148,149]. It has been shown that T cells and natural killer cells are primary sources of IFN-γ production, but it is unclear whether these two cell types are the main source of IFN-γ in response to CpG oligodeoxynucleotides. It was revealed that IRF8 directly regulates the expression of TLR9 and its deficiency leads to no increased IFNγ production expected with CpG stimulation [147]. Physiologically, IRF8 is highly expressed in NK cells. It was shown that IRF8-deficient mice exhibited a decreased proportion and total count of mature NK cells in the bone marrow. This was due to elevated neutrophil abundance in Irf8−/− bone marrow cavity [147]. In addition, because NK cells are involved in the early immune response against viral pathogens, the resulting NK cell deficiencies resulted in severe viral disease. Similarly, NK IRF8 -/- mice infected with MCMV exhibited poorly controlled viral replication and decreased survival [150]. IRF8 -/- NK cells exhibited decreased virus-driven expansion and impaired proliferation which may account for the poorly controlled viral replication seen in the mice models of IRF8 deficiency [150].
6. IRF8 Expression and Function in Non-Hematopoietic Cancer Cells
While much research has been dedicated to understanding the role of IRF8 in hematopoietic cells, less research has investigated the role of IRF8 in non-hematopoietic cells. In certain types of cancers, IRF8 functions as a tumor suppressor gene [18,19,42,43,44], and its expression is regulated by DNA methylation or micro-RNA interference [113]. In breast and lung cancer, IRF8 is downregulated as a result of promoter hypermethylation [43,151,152], and a higher IRF8 expression level was associated with a better prognosis [19]. Similarly, in gastric, nasopharyngeal, esophageal, colorectal, lung, prostate, and renal cell carcinomas the expression of IRF8 is downregulated [43,44,153,154,155,156,157]. IFN-γ induced expression of IRF8 was increased when gastric cancer cells were treated with a demethylating agent [153]. In other cancer cell lines with a methylated IRF8 promoter, IFN-γ treatment showed decreased ability to induce IRF8 expression [154]. Taken together, these studies provide evidence for promoter methylation as a mechanism for IRF8 silencing in certain types of human cancer cells. Analysis of human lung tumor specimens at the single cell level validate the early finding that IRF8 is silenced in the tumor cells. Except for certain tumor-infiltrating immune cells, including certain B cells, pDC and myeloid cells (Figure 4A), IRF8 is down-regulated in most cell types and is undetectable in human lung cancer cells (Figure 4B) [158].
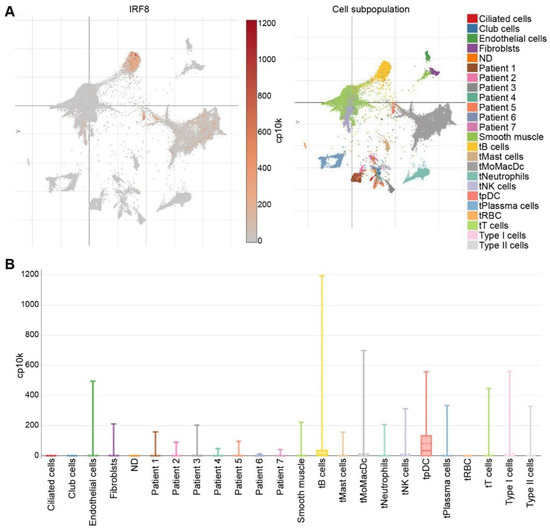
Figure 4.
IRF8 expression profiles in human lung cancer in the single cell level. (A). scRNA-Seq datasets were extracted from the lung cancer scRNA-Seq datasets in the Broad Institute Single Cell Portal. Right panel shows UMAP plot of cell subpopulations from human lung tumor. Cells are color-coded. Left panel shows UMAP plot of IRF8 expression in the respective cells as shown in the right panel. (B). Box plot showing IRF8 expression level in the indicated cells and tumor (Patients 1–7). t: tumor.
Within non-hematopoietic cancer cells, IRF8 plays a protective role against the formation of a metastatic phenotype. In human colon cancer cells, IRF8 protein levels are inversely correlated with the metastatic phenotype [46,47]. IRF8 expression sensitized the metastatic tumor cells to Fas-mediated apoptosis, indicating that loss of IRF8 in cancers could promote earlier metastasis [46]. It has been suggested that colon cancer cells silence IRF8 expression through inhibition of pSTAT1 function at the IRF8 promoter [47]. IRF8 may also function in non-hematopoietic cancers to negatively regulate the expression of MMP3, a matrix metalloproteinase associated with a poor prognosis and metastasis in various solid organ cancers [159,160,161]. In a chemically-induced sarcoma model, an inverse relationship between IRF8 and MMP3 expression was demonstrated; in a mouse model of mammary cancer, loss of MMP3 reduced spontaneous lung metastasis [162]. This suggests that IRF8 may negatively regulate MMP3 expression, protecting against MMP3-induced metastasis in solid organ tumors. While most research supports an anti-metastasis role for IRF8 in non-hematopoietic cancers, one study found that IRF8 promoted EMT-like phenomena, cell motility, and invasion in a human osteosarcoma cell line providing evidence for a possible role in the acquisition of a metastatic-like phenotype [163].
IRF8 appears to impede the progression and formation of cancer by increasing the expression of tumor suppressor genes, such as Caspase 1, p21, p27, and PTEN [44,157,164]. Expression of tumor suppressor genes inhibited lens cell carcinoma growth upon transfection with IRF8 [164]. Similarly, IRF8-induced expression of p27 induced lung cancer cell senescence [44]. Mechanistically, IRF8 was shown to inhibit Akt activity thereby inducing the accumulation of p27 and promoting senescence. While IRF8 increases tumor suppressor genes, one study also showed an IRF8-induced decrease in oncogene (Yap1 and Survivin) expression in renal cell carcinoma cell lines [157]. In this cell line, ectopic expression of IRF8 induced cell cycle G2/M arrest and apoptosis further supporting IRF8’s role in impeding cancer progression.
IRF8 functions as a promoter of Fas-mediated apoptosis. This form of extrinsic apoptosis is key to immune clearance of tumor cells and is targeted in cancer therapy [165,166]. Modification of Fas expression can render immune checkpoint inhibitor (ICI) therapies unsuccessful. Colon cancer cells with low Fas expression exhibited decreased sensitivity to FasL-induced apoptosis (a mechanism utilized by ICIs) [167] and lower Fas expression was correlated with decreased survival in colon cancer patients [168]. One mechanism by which cancer cells may alter the expression of Fas, and thereby reduce sensitivity to apoptosis, is through downregulation of IRF8.
As previously mentioned, IRF8 is downregulated in many cancers. Studies revolving around a loss of IRF8 have elucidated ways in which IRF8 helps facilitate Fas-mediated apoptosis and resulting in tumor immune sensitivity. In soft tissue sarcoma cells, IRF8 was found to be a repressor of FLICE-like protein (FLIP) [169]. Silencing of FLIP and ectopic IRF8 expression each increased susceptibility to FasL-induced apoptosis. This suggests that by repressing FLIP, IRF8 promotes Fas-mediated apoptosis susceptibility. Mechanistically, IRF8 disruption diminishes JAK1 expression and inhibits STAT1 phosphorylation to block IFN-γ induced Fas upregulation in sarcoma cells [170]. IRF8 has also been found to regulate pro-apoptotic and anti-apoptotic factors contributing to Fas-mediated apoptosis. In tumor-induced MDSCs, levels of IRF8 are decreased. IRF8-deficient MDSCs showed an increase in anti-apoptotic Bcl-xL and a decrease in pro-apoptotic Bax, providing a mechanism by which IRF8 downregulation can impede the Fas-mediated apoptosis pathway to evade elimination by T lymphocytes [32]. Another mechanism by which IRF8 can promote Fas-mediated apoptosis is through repression of acid ceramidase (A-CDase). A-CDase upregulation has been implicated in certain cancers such as prostate and breast [171,172,173]. IRF8 induced repression of A-CDase resulted in C16 ceramide accumulation and increased apoptosis sensitivity [31]. This suggests that IRF8 downregulation likely contributes to decreased Fas-mediated apoptosis sensitivity through failure to repress A-CDase. In summary, IRF8 promotes Fas-mediated apoptosis through repression of FLIP and A-CDase and by regulating apoptotic factors such as Bcl-xL and Bax [31,32,169].
IRF8 functions as a link between cancer and the immune system. Poor tumoral homing of DCs and T cells was observed in melanoma-bearing IRF8 KO mice [42]. Additionally, the few DCs that were able to infiltrate the tumor were immature and unable to produce T cell-mediated immune responses. Mechanistically, the decreased tumoral immune infiltration observed in IRF8 KO mice is suggested to occur through the aberrant expression of multiple chemokine receptors and ligands [42]. When IRF8 was induced, immune infiltration was reestablished, and the chemokine expression pattern was reversed. Using a microfluidic device, in which melanoma cells and splenocytes were separated by a microchannel, IRF8 KO splenocytes exhibited a marked decrease in the ability to cross the channel to infiltrate the melanoma cells while melanoma cells exhibited marked propensity to invade the channels suggesting existence of an IRF8-regulated communicating soluble factor [174].
OPN has recently emerged as another immune checkpoint that may compensate PD-L1 function to promote colon tumor immune evasion [175]. IRF8 is expressed in human and mouse gastric and colon epithelial cells [38,41] and binds to the ISRE elements at the Spp1 promoter to repress OPN expression [176]. In human colon and pancreatic cancers, IRF8 expression is down-regulated and OPN is up-regulated [175,176,177,178,179]. scRNA-Seq analysis indicates that OPN is abundantly expressed in tumor cells, MDSCs, and ILCs [175]. Furthermore, scRNA-Seq analysis revealed the tumor-promoting role of SPP1+ cells in cancer [180,181]. OPN protein level is highly elevated in the peripheral blood in human cancer patients and OPN expression is up-regulated in several different types of human cancers [182,183,184,185,186,187,188,189,190]. Silenced IRF8 expression in both tumor cells and tumor-induced myeloid cells by DNA methylation and H3K9me3 deposition at the Irf8 promoter are likely major mechanisms underlying OPN elevation in cancer patients and tumor-bearing mice [41,46,47,154,191]. It was observed that IRF8 KO mice exhibited deficient generation of antigen-specific T cells and this deficiency was due to OPN engagement of CD44 to directly suppress T cell activation [176]. Accordingly, OPN blockade increased activation of tumor-specific T cells and suppressed tumor growth in vivo [175]. OPN is highly expressed in human pancreatic tumor cells and may bind to CD44 on tumor cells via autocrine and paracrine manners to promote pancreatic cancer stemness and progression [177,179]. Therefore, in addition to functioning as a repressor of SPP1 in immune cells such as MDSCs and ILCs, IRF8 not only functions as an intrinsic tumor suppressor [41] but also suppresses tumorigenesis and progression through repressing OPN expression in tumor cells (Figure 5) [176].
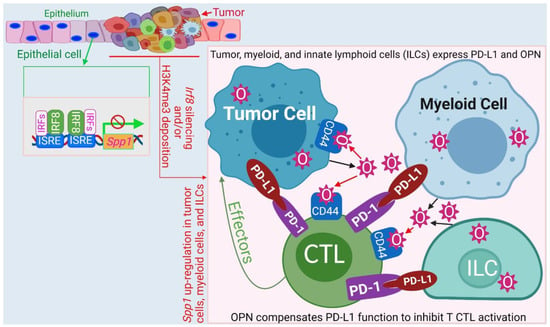
Figure 5.
Model of the IRF8-OPN axis mechanism of action in immune suppression and tumor promotion. IRF8 functions as a repressor that binds to the two ISRE sites at the Spp1 promoter to repress Spp1 transcription in colon epithelial cells. During tumor development, Irf8 is silenced by its promoter DNA methylation and H3K9me3 deposition, resulting in increased OPN expression in tumor cells, MDSCs, and ILCs. OPN binds to CD44 to suppress T cell activation and to CD44 on tumor cells to promote tumor cell stemness and progression. The IRF8-OPN axis thus control tumor growth and progression through its functions in both the immune and non-immune cell components.
7. Conclusions
IRF8 is a transcription factor that was identified as a member of the IFN transcription factor family more than three decades ago. IRF8 is essential for myelopoiesis and loss of IRF8 expression or function results in accumulation of CD11b+Gr1+ IMCs that phenotypically and functionally resemble tumor-induced MDSCs. IRF8 also plays a key role in immune cell homeostasis and turnover to prevent hematopoietic malignancies. IRF8 silencing by its promoter DNA methylation leads to accumulation of MDSCs in both human cancer patients and tumor-bearing mice. Although IRF8 is highly expressed in hematopoietic cells and it was once thought that its expression was restricted to immune cells, IRF8 expression and function has been extended to non-hematopoietic cells. IRF8 is now known to play a key role in epithelial cell turnover and functions as a tumor suppressor in solid tumors. Furthermore, IRF8 has been found to protect against immune evasion of tumor cells via the IRF8-OPN axis to control tumorigenesis and progression. IRF8 therefore is not only a key regulator of host response to infection, but is also a key regulator in host cancer immune surveillance and response to immunotherapy. In the therapeutic front, recent developments have been made in the relationship between IRF8 and immunotherapies. One such study focused on the role of IRF8 in response to anti-CD20 treatments such as rituximab and ofatumumab. IRF8 was necessary for CD20 transcription and anti-CD20 therapy efficacy [192]. Additionally, IRF8 is a transcriptional regulator of CD37 in DLBCL and regulates the efficacy of anti-CD37 pharmacotherapies [55]. Continued investigation on anti-PD-L1 therapy has shown that, in hepatocellular carcinoma, IRF8 enhances the response to treatment and suppressed progression [18]. IRF8 is therefore a therapeutic target in both hematopoietic and non-hematopoietic cancer.
Author Contributions
Conceptualization, H.R.M., Y.R., D.B.P. and K.L.; Formal analysis, D.B.P. and K.L.; Data curation, D.B.P. and K.L.; writing—original draft preparation, H.R.M. and Y.R.; writing—review and editing, K.L.; supervision, K.L.; funding acquisition, K.L. All authors have read and agreed to the published version of the manuscript.
Funding
NCI/NIH CA227433 (K.L.) and US Department of Veterans Affairs CX001364 (K.L.).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Tamura, T.; Yanai, H.; Savitsky, D.; Taniguchi, T. The IRF family transcription factors in immunity and oncogenesis. Annu. Rev. Immunol. 2008, 26, 535–584. [Google Scholar] [CrossRef] [PubMed]
- Harada, H.; Fujita, T.; Miyamoto, M.; Kimura, Y.; Maruyama, M.; Furia, A.; Miyata, T.; Taniguchi, T. Structurally similar but functionally distinct factors, IRF-1 and IRF-2, bind to the same regulatory elements of IFN and IFN-inducible genes. Cell 1989, 58, 729–739. [Google Scholar] [CrossRef]
- Miyamoto, M.; Fujita, T.; Kimura, Y.; Maruyama, M.; Harada, H.; Sudo, Y.; Miyata, T.; Taniguchi, T. Regulated expression of a gene encoding a nuclear factor, IRF-1, that specifically binds to IFN-beta gene regulatory elements. Cell 1988, 54, 903–913. [Google Scholar] [CrossRef]
- Honda, K.; Takaoka, A.; Taniguchi, T. Type I interferon [corrected] gene induction by the interferon regulatory factor family of transcription factors. Immunity 2006, 25, 349–360. [Google Scholar] [CrossRef]
- Das, A.; Chauhan, K.S.; Kumar, H.; Tailor, P. Mutation in Irf8 Gene (Irf8(R294C)) Impairs Type I IFN-Mediated Antiviral Immune Response by Murine pDCs. Front. Immunol. 2021, 12, 758190. [Google Scholar] [CrossRef]
- Ikushima, H.; Negishi, H.; Taniguchi, T. The IRF family transcription factors at the interface of innate and adaptive immune responses. Cold Spring Harb. Lab. Press 2013, 78, 105–116. [Google Scholar] [CrossRef]
- Driggers, P.H.; Ennist, D.L.; Gleason, S.L.; Mak, W.H.; Marks, M.S.; Levi, B.Z.; Flanagan, J.R.; Appella, E.; Ozato, K. An interferon gamma-regulated protein that binds the interferon-inducible enhancer element of major histocompatibility complex class I genes. Proc. Natl. Acad. Sci. USA 1990, 87, 3743–3747. [Google Scholar] [CrossRef]
- Kim, S.H.; Burton, J.; Yu, C.R.; Sun, L.; He, C.; Wang, H.; Morse, H.C., 3rd; Egwuagu, C.E. Dual Function of the IRF8 Transcription Factor in Autoimmune Uveitis: Loss of IRF8 in T Cells Exacerbates Uveitis, Whereas Irf8 Deletion in the Retina Confers Protection. J. Immunol. 2015, 195, 1480–1488. [Google Scholar] [CrossRef]
- Hambleton, S.; Salem, S.; Bustamante, J.; Bigley, V.; Boisson-Dupuis, S.; Azevedo, J.; Fortin, A.; Haniffa, M.; Ceron-Gutierrez, L.; Bacon, C.M.; et al. IRF8 mutations and human dendritic-cell immunodeficiency. N. Engl. J. Med. 2011, 365, 127–138. [Google Scholar] [CrossRef]
- Jiang, D.S.; Wei, X.; Zhang, X.F.; Liu, Y.; Zhang, Y.; Chen, K.; Gao, L.; Zhou, H.; Zhu, X.H.; Liu, P.P.; et al. IRF8 suppresses pathological cardiac remodelling by inhibiting calcineurin signalling. Nat. Commun. 2014, 5, 3303. [Google Scholar] [CrossRef]
- Grajales-Reyes, G.E.; Iwata, A.; Albring, J.; Wu, X.; Tussiwand, R.; Kc, W.; Kretzer, N.M.; Briseno, C.G.; Durai, V.; Bagadia, P.; et al. Batf3 maintains autoactivation of Irf8 for commitment of a CD8alpha(+) conventional DC clonogenic progenitor. Nat. Immunol. 2015, 16, 708–717. [Google Scholar] [CrossRef] [PubMed]
- Lanca, T.; Ungerback, J.; Da Silva, C.; Joeris, T.; Ahmadi, F.; Vandamme, J.; Svensson-Frej, M.; Mowat, A.M.; Kotarsky, K.; Sigvardsson, M.; et al. IRF8 deficiency induces the transcriptional, functional, and epigenetic reprogramming of cDC1 into the cDC2 lineage. Immunity 2022, 55, 1431–1447. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Inoue, K.; Du, Y.; Baker, S.J.; Premkumar Reddy, E.; Greenblatt, M.B.; Zhao, B. TGFbeta reprograms TNF stimulation of macrophages towards a non-canonical pathway driving inflammatory osteoclastogenesis. Nat. Commun. 2022, 13, 3920. [Google Scholar] [CrossRef]
- Liu, T.T.; Kim, S.; Desai, P.; Kim, D.H.; Huang, X.; Ferris, S.T.; Wu, R.; Ou, F.; Egawa, T.; Van Dyken, S.J.; et al. Ablation of cDC2 development by triple mutations within the Zeb2 enhancer. Nature 2022, 607, 142–148. [Google Scholar] [CrossRef]
- Ferris, S.T.; Ohara, R.A.; Ou, F.; Wu, R.; Huang, X.; Kim, S.; Chen, J.; Liu, T.T.; Schreiber, R.D.; Murphy, T.L.; et al. cDC1 vaccines drive tumor rejection by direct presentation independently of host cDC1. Cancer Immunol. Res. 2022, 10, 920–931. [Google Scholar] [CrossRef] [PubMed]
- Onuora, S. SLE risk variant regulates IRF8 expression. Nat. Rev. Rheumatol. 2022, 18, 306. [Google Scholar] [CrossRef]
- Gao, X.; Ge, J.; Zhou, W.; Xu, L.; Geng, D. IL-10 inhibits osteoclast differentiation and osteolysis through MEG3/IRF8 pathway. Cell Signal 2022, 95, 110353. [Google Scholar] [CrossRef]
- Wu, H.; Li, Y.; Shi, G.; Du, S.; Wang, X.; Ye, W.; Zhang, Z.; Chu, Y.; Ma, S.; Wang, D.; et al. Hepatic interferon regulatory factor 8 expression suppresses hepatocellular carcinoma progression and enhances the response to anti-programmed cell death protein-1 therapy. Hepatology 2022. [Google Scholar] [CrossRef]
- Gatti, G.; Betts, C.; Rocha, D.; Nicola, M.; Grupe, V.; Ditada, C.; Nunez, N.G.; Roselli, E.; Araya, P.; Dutto, J.; et al. High IRF8 expression correlates with CD8 T cell infiltration and is a predictive biomarker of therapy response in ER-negative breast cancer. Breast Cancer Res. 2021, 23, 40. [Google Scholar] [CrossRef]
- Murakami, K.; Sasaki, H.; Nishiyama, A.; Kurotaki, D.; Kawase, W.; Ban, T.; Nakabayashi, J.; Kanzaki, S.; Sekita, Y.; Nakajima, H.; et al. A RUNX-CBFbeta-driven enhancer directs the Irf8 dose-dependent lineage choice between DCs and monocytes. Nat. Immunol. 2021, 22, 301–311. [Google Scholar] [CrossRef]
- Nutt, S.L.; Chopin, M. Transcriptional Networks Driving Dendritic Cell Differentiation and Function. Immunity 2020, 52, 942–956. [Google Scholar] [CrossRef] [PubMed]
- Turcotte, K.; Gauthier, S.; Tuite, A.; Mullick, A.; Malo, D.; Gros, P. A mutation in the Icsbp1 gene causes susceptibility to infection and a chronic myeloid leukemia-like syndrome in BXH-2 mice. J. Exp. Med. 2005, 201, 881–890. [Google Scholar] [CrossRef] [PubMed]
- Tailor, P.; Tamura, T.; Morse, H.C., 3rd; Ozato, K. The BXH2 mutation in IRF8 differentially impairs dendritic cell subset development in the mouse. Blood 2008, 111, 1942–1945. [Google Scholar] [CrossRef]
- Salem, S.; Langlais, D.; Lefebvre, F.; Bourque, G.; Bigley, V.; Haniffa, M.; Casanova, J.L.; Burk, D.; Berghuis, A.; Butler, K.M.; et al. Functional characterization of the human dendritic cell immunodeficiency associated with the IRF8K108E mutation. Blood 2014, 124, 1894–1904. [Google Scholar] [CrossRef]
- Dang, D.; Liu, Y.; Zhou, Q.; Li, H.; Wang, Y.; Wu, H. Identification of a novel IRF8 homozygous mutation causing neutrophilia, monocytopenia and fatal infection in a female neonate. Infect. Genet. Evol. 2021, 96, 105121. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.D.; Wang, Y.H.; Liu, C.H.; Lin, Y.C.; Lin, J.A.; Lin, Y.F.; Tang, K.T.; Cheng, C.W. Association of IRF8 gene polymorphisms with autoimmune thyroid disease. Eur. J. Clin. Investig. 2015, 45, 711–719. [Google Scholar] [CrossRef]
- Ding, S.; Jiang, T.; He, J.; Qin, B.; Lin, S.; Li, L. Tagging single nucleotide polymorphisms in the IRF1 and IRF8 genes and tuberculosis susceptibility. PLoS ONE 2012, 7, e42104. [Google Scholar] [CrossRef]
- Jiang, Y.; Wang, H.; Yu, H.; Li, L.; Xu, D.; Hou, S.; Kijlstra, A.; Yang, P. Two Genetic Variations in the IRF8 region are associated with Behcet’s disease in Han Chinese. Sci. Rep. 2016, 6, 19651. [Google Scholar] [CrossRef]
- Gabriele, L.; Phung, J.; Fukumoto, J.; Segal, D.; Wang, I.M.; Giannakakou, P.; Giese, N.A.; Ozato, K.; Morse, H.C. Regulation of apoptosis in myeloid cells by interferon consensus sequence-binding protein. J. Exp. Med. 1999, 190, 411–421. [Google Scholar] [CrossRef]
- Burchert, A.; Cai, D.; Hofbauer, L.C.; Samuelsson, M.K.; Slater, E.P.; Duyster, J.; Ritter, M.; Hochhaus, A.; Muller, R.; Eilers, M.; et al. Interferon consensus sequence binding protein (ICSBP.; IRF-8) antagonizes BCR/ABL and down-regulates bcl-2. Blood 2004, 103, 3480–3489. [Google Scholar] [CrossRef]
- Hu, X.; Yang, D.; Zimmerman, M.; Liu, F.; Yang, J.; Kannan, S.; Burchert, A.; Szulc, Z.; Bielawska, A.; Ozato, K.; et al. IRF8 regulates acid ceramidase expression to mediate apoptosis and suppresses myelogeneous leukemia. Cancer Res. 2011, 71, 2882–2891. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Bardhan, K.; Paschall, A.V.; Yang, D.; Waller, J.L.; Park, M.A.; Nayak-Kapoor, A.; Samuel, T.A.; Abrams, S.I.; Liu, K. Deregulation of apoptotic factors Bcl-xL and Bax confers apoptotic resistance to myeloid-derived suppressor cells and contributes to their persistence in cancer. J. Biol. Chem. 2013, 288, 19103–19115. [Google Scholar] [CrossRef] [PubMed]
- Tamura, T.; Ozato, K. ICSBP/IRF-8: Its regulatory roles in the development of myeloid cells. J. Interferon Cytokine Res. 2002, 22, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Kurotaki, D.; Yamamoto, M.; Nishiyama, A.; Uno, K.; Ban, T.; Ichino, M.; Sasaki, H.; Matsunaga, S.; Yoshinari, M.; Ryo, A.; et al. IRF8 inhibits C/EBPalpha activity to restrain mononuclear phagocyte progenitors from differentiating into neutrophils. Nat. Commun. 2014, 5, 4978. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Yun, H.; Jyotsana, N.; Chaturvedi, A.; Schwarzer, A.; Yung, E.; Lai, C.K.; Kuchenbauer, F.; Argiropoulos, B.; Gorlich, K.; et al. Constitutive IRF8 expression inhibits AML by activation of repressed immune response signaling. Leukemia 2015, 29, 157–168. [Google Scholar] [CrossRef] [PubMed]
- Liss, F.; Frech, M.; Wang, Y.; Giel, G.; Fischer, S.; Simon, C.; Weber, L.M.; Nist, A.; Stiewe, T.; Neubauer, A.; et al. IRF8 Is an AML-Specific Susceptibility Factor That Regulates Signaling Pathways and Proliferation of AML Cells. Cancers 2021, 13, 764. [Google Scholar] [CrossRef]
- Wang, H.; Yan, M.; Sun, J.; Jain, S.; Yoshimi, R.; Abolfath, S.M.; Ozato, K.; Coleman, W.G., Jr.; Ng, A.P.; Metcalf, D.; et al. A Reporter Mouse Reveals Lineage-Specific and Heterogeneous Expression of IRF8 during Lymphoid and Myeloid Cell Differentiation. J. Immunol. 2014, 193, 1766–1777. [Google Scholar] [CrossRef]
- Yan, M.; Wang, H.; Sun, J.; Liao, W.; Li, P.; Zhu, Y.; Xu, C.; Joo, J.; Sun, Y.; Abbasi, S.; et al. Cutting Edge: Expression of IRF8 in Gastric Epithelial Cells Confers Protective Innate Immunity against Helicobacter pylori Infection. J. Immunol. 2016, 196, 1999–2003. [Google Scholar] [CrossRef]
- Li, W.; Nagineni, C.N.; Ge, H.; Efiok, B.; Chepelinsky, A.B.; Egwuagu, C.E. Interferon consensus sequence-binding protein is constitutively expressed and differentially regulated in the ocular lens. J. Biol. Chem. 1999, 274, 9686–9691. [Google Scholar] [CrossRef]
- Sun, L.; St Leger, A.J.; Yu, C.R.; He, C.; Mahdi, R.M.; Chan, C.C.; Wang, H.; Morse, H.C., 3rd; Egwuagu, C.E. Interferon Regulator Factor 8 (IRF8) Limits Ocular Pathology during HSV-1 Infection by Restraining the Activation and Expansion of CD8+ T Cells. PLoS ONE 2016, 11, e0155420. [Google Scholar] [CrossRef]
- Ibrahim, M.L.; Klement, J.D.; Lu, C.; Redd, P.S.; Xiao, W.; Yang, D.; Browning, D.D.; Savage, N.M.; Buckhaults, P.J.; Morse, H.C., 3rd; et al. Myeloid-Derived Suppressor Cells Produce IL-10 to Elicit DNMT3b-Dependent IRF8 Silencing to Promote Colitis-Associated Colon Tumorigenesis. Cell Rep. 2018, 25, 3036–3046.e6. [Google Scholar] [CrossRef] [PubMed]
- Mattei, F.; Schiavoni, G.; Sestili, P.; Spadaro, F.; Fragale, A.; Sistigu, A.; Lucarini, V.; Spada, M.; Sanchez, M.; Scala, S.; et al. IRF-8 controls melanoma progression by regulating the cross talk between cancer and immune cells within the tumor microenvironment. Neoplasia 2012, 14, 1223–1235. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.; Ikeda, K.; Shiraishi, K.; Eguchi, A.; Mori, T.; Yoshimoto, K.; Shibata, H.; Ito, T.; Baba, Y.; Baba, H. Aberrant methylation and silencing of IRF8 expression in non-small cell lung cancer. Oncol. Lett. 2014, 8, 1025–1030. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Lu, F.; Li, B.; Liu, L.; Zeng, G.; Zhou, Q.; Chen, L. IRF8 induces senescence of lung cancer cells to exert its tumor suppressive function. Cell Cycle 2019, 18, 3300–3312. [Google Scholar] [CrossRef]
- Kesper, C.; Viestenz, A.; Wiese-Rischke, C.; Scheller, M.; Hammer, T. Impact of the transcription factor IRF8 on limbal epithelial progenitor cells in a mouse model. Exp. Eye Res. 2022, 218, 108985. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Thangaraju, M.; Greeneltch, K.; Browning, D.D.; Schoenlein, P.V.; Tamura, T.; Ozato, K.; Ganapathy, V.; Abrams, S.I.; Liu, K. Repression of IFN regulatory factor 8 by DNA methylation is a molecular determinant of apoptotic resistance and metastatic phenotype in metastatic tumor cells. Cancer Res. 2007, 67, 3301–3309. [Google Scholar] [CrossRef]
- McGough, J.M.; Yang, D.; Huang, S.; Georgi, D.; Hewitt, S.M.; Rocken, C.; Tanzer, M.; Ebert, M.P.; Liu, K. DNA methylation represses IFN-gamma-induced and signal transducer and activator of transcription 1-mediated IFN regulatory factor 8 activation in colon carcinoma cells. Mol. Cancer Res. 2008, 6, 1841–1851. [Google Scholar] [CrossRef]
- Lipman, D.J.; Pearson, W.R. Rapid and sensitive protein similarity searches. Science 1985, 227, 1435–1441. [Google Scholar] [CrossRef]
- Escalante, C.R.; Brass, A.L.; Pongubala, J.M.; Shatova, E.; Shen, L.; Singh, H.; Aggarwal, A.K. Crystal structure of PU.1/IRF-4/DNA ternary complex. Mol. Cell 2002, 10, 1097–1105. [Google Scholar] [CrossRef]
- Langlais, D.; Barreiro, L.B.; Gros, P. The macrophage IRF8/IRF1 regulome is required for protection against infections and is associated with chronic inflammation. J. Exp. Med. 2016, 213, 585–603. [Google Scholar] [CrossRef]
- Tamura, T.; Kong, H.J.; Tunyaplin, C.; Tsujimura, H.; Calame, K.; Ozato, K. ICSBP/IRF-8 inhibits mitogenic activity of p210 Bcr/Abl in differentiating myeloid progenitor cells. Blood 2003, 102, 4547–4554. [Google Scholar] [CrossRef] [PubMed]
- Tamura, T.; Thotakura, P.; Tanaka, T.S.; Ko, M.S.; Ozato, K. Identification of target genes and a unique cis element regulated by IRF-8 in developing macrophages. Blood 2005, 106, 1938–1947. [Google Scholar] [CrossRef] [PubMed]
- Kuwata, T.; Gongora, C.; Kanno, Y.; Sakaguchi, K.; Tamura, T.; Kanno, T.; Basrur, V.; Martinez, R.; Appella, E.; Golub, T.; et al. Gamma interferon triggers interaction between ICSBP (IRF-8) and TEL, recruiting the histone deacetylase HDAC3 to the interferon-responsive element. Mol. Cell Biol. 2002, 22, 7439–7448. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, M.; Kato, T.; Hotta, C.; Nishiyama, A.; Kurotaki, D.; Yoshinari, M.; Takami, M.; Ichino, M.; Nakazawa, M.; Matsuyama, T.; et al. Shared and distinct functions of the transcription factors IRF4 and IRF8 in myeloid cell development. PLoS ONE 2011, 6, e25812. [Google Scholar] [CrossRef] [PubMed]
- Elfrink, S.; Ter Beest, M.; Janssen, L.; Baltissen, M.P.; Jansen, P.; Kenyon, A.N.; Steen, R.M.; de Windt, D.; Hagemann, P.M.; Hess, C.; et al. IRF8 is a transcriptional activator of CD37 expression in diffuse large B-cell lymphoma. Blood Adv. 2022, 6, 2254–2266. [Google Scholar] [CrossRef]
- Kanno, Y.; Levi, B.Z.; Tamura, T.; Ozato, K. Immune cell-specific amplification of interferon signaling by the IRF-4/8-PU.1 complex. J. Interferon Cytokine Res. 2005, 25, 770–779. [Google Scholar] [CrossRef]
- Kurotaki, D.; Osato, N.; Nishiyama, A.; Yamamoto, M.; Ban, T.; Sato, H.; Nakabayashi, J.; Umehara, M.; Miyake, N.; Matsumoto, N.; et al. Essential role of the IRF8-KLF4 transcription factor cascade in murine monocyte differentiation. Blood 2013, 121, 1839–1849. [Google Scholar] [CrossRef]
- Kurotaki, D.; Tamura, T. Transcriptional and Epigenetic Regulation of Innate Immune Cell Development by the Transcription Factor, Interferon Regulatory Factor-8. J. Interferon Cytokine Res. 2016, 36, 433–441. [Google Scholar] [CrossRef]
- Laricchia-Robbio, L.; Tamura, T.; Karpova, T.; Sprague, B.L.; McNally, J.G.; Ozato, K. Partner-regulated interaction of IFN regulatory factor 8 with chromatin visualized in live macrophages. Proc. Natl. Acad. Sci. USA 2005, 102, 14368–14373. [Google Scholar] [CrossRef]
- Brass, A.L.; Kehrli, E.; Eisenbeis, C.F.; Storb, U.; Singh, H. Pip, a lymphoid-restricted IRF, contains a regulatory domain that is important for autoinhibition and ternary complex formation with the Ets factor PU.1. Genes Dev. 1996, 10, 2335–2347. [Google Scholar] [CrossRef]
- Brass, A.L.; Zhu, A.Q.; Singh, H. Assembly requirements of PU.1-Pip (IRF-4) activator complexes: Inhibiting function in vivo using fused dimers. EMBO J. 1999, 18, 977–991. [Google Scholar] [CrossRef]
- Ozato, K.; Tsujimura, H.; Tamura, T. Toll-like receptor signaling and regulation of cytokine gene expression in the immune system. Biotechniques 2002, 33, S66–S75. [Google Scholar] [CrossRef]
- Schmidt, M.; Bies, J.; Tamura, T.; Ozato, K.; Wolff, L. The interferon regulatory factor ICSBP/IRF-8 in combination with PU.1 up-regulates expression of tumor suppressor p15(Ink4b) in murine myeloid cells. Blood 2004, 103, 4142–4149. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Tsujimura, H.; Tamura, T.; Gongora, C.; Aliberti, J.; Reis e Sousa, C.; Sher, A.; Ozato, K. ICSBP/IRF-8 retrovirus transduction rescues dendritic cell development in vitro. Blood 2003, 101, 961–969. [Google Scholar] [CrossRef]
- Dror, N.; Rave-Harel, N.; Burchert, A.; Azriel, A.; Tamura, T.; Tailor, P.; Neubauer, A.; Ozato, K.; Levi, B.Z. Interferon Regulatory Factor-8 Is Indispensable for the Expression of Promyelocytic Leukemia and the Formation of Nuclear Bodies in Myeloid Cells. J. Biol. Chem. 2007, 282, 5633–5640. [Google Scholar] [CrossRef] [PubMed]
- Humblin, E.; Thibaudin, M.; Chalmin, F.; Derangere, V.; Limagne, E.; Richard, C.; Flavell, R.A.; Chevrier, S.; Ladoire, S.; Berger, H.; et al. IRF8-dependent molecular complexes control the Th9 transcriptional program. Nat. Commun. 2017, 8, 2085. [Google Scholar] [CrossRef] [PubMed]
- Glasmacher, E.; Agrawal, S.; Chang, A.B.; Murphy, T.L.; Zeng, W.; Vander Lugt, B.; Khan, A.A.; Ciofani, M.; Spooner, C.J.; Rutz, S.; et al. A genomic regulatory element that directs assembly and function of immune-specific AP-1-IRF complexes. Science 2012, 338, 975–980. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Spolski, R.; Liao, W.; Wang, L.; Murphy, T.L.; Murphy, K.M.; Leonard, W.J. BATF-JUN is critical for IRF4-mediated transcription in T cells. Nature 2012, 490, 543–546. [Google Scholar] [CrossRef]
- Tussiwand, R.; Lee, W.L.; Murphy, T.L.; Mashayekhi, M.; Kc, W.; Albring, J.C.; Satpathy, A.T.; Rotondo, J.A.; Edelson, B.T.; Kretzer, N.M.; et al. Compensatory dendritic cell development mediated by BATF-IRF interactions. Nature 2012, 490, 502–507. [Google Scholar] [CrossRef]
- Salem, S.; Salem, D.; Gros, P. Role of IRF8 in immune cells functions, protection against infections, and susceptibility to inflammatory diseases. Hum. Genet. 2020, 139, 707–721. [Google Scholar] [CrossRef]
- Bovolenta, C.; Driggers, P.H.; Marks, M.S.; Medin, J.A.; Politis, A.D.; Vogel, S.N.; Levy, D.E.; Sakaguchi, K.; Appella, E.; Coligan, J.E.; et al. Molecular interactions between interferon consensus sequence binding protein and members of the interferon regulatory factor family. Proc. Natl. Acad. Sci. USA 1994, 91, 5046–5050. [Google Scholar] [CrossRef] [PubMed]
- Nelson, N.; Marks, M.S.; Driggers, P.H.; Ozato, K. Interferon consensus sequence-binding protein, a member of the interferon regulatory factor family, suppresses interferon-induced gene transcription. Mol. Cell Biol. 1993, 13, 588–599. [Google Scholar] [CrossRef] [PubMed]
- Rosenbauer, F.; Waring, J.F.; Foerster, J.; Wietstruk, M.; Philipp, D.; Horak, I. Interferon consensus sequence binding protein and interferon regulatory factor-4/Pip form a complex that represses the expression of the interferon-stimulated gene-15 in macrophages. Blood 1999, 94, 4274–4281. [Google Scholar] [CrossRef] [PubMed]
- Cytlak, U.; Resteu, A.; Pagan, S.; Green, K.; Milne, P.; Maisuria, S.; McDonald, D.; Hulme, G.; Filby, A.; Carpenter, B.; et al. Differential IRF8 Transcription Factor Requirement Defines Two Pathways of Dendritic Cell Development in Humans. Immunity 2020, 53, 353–370.e8. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Bagadia, P.; Anderson, D.A., 3rd; Liu, T.T.; Huang, X.; Theisen, D.J.; O’Connor, K.W.; Ohara, R.A.; Iwata, A.; Murphy, T.L.; et al. High Amount of Transcription Factor IRF8 Engages AP1-IRF Composite Elements in Enhancers to Direct Type 1 Conventional Dendritic Cell Identity. Immunity 2020, 53, 759–774.e9. [Google Scholar] [CrossRef]
- Paschall, A.V.; Zhang, R.; Qi, C.F.; Bardhan, K.; Peng, L.; Lu, G.; Yang, J.; Merad, M.; McGaha, T.; Zhou, G.; et al. IFN Regulatory Factor 8 Represses GM-CSF Expression in T Cells To Affect Myeloid Cell Lineage Differentiation. J. Immunol. 2015, 194, 2369–2379. [Google Scholar] [CrossRef]
- Schwieger, M.; Lohler, J.; Friel, J.; Scheller, M.; Horak, I.; Stocking, C. AML1-ETO inhibits maturation of multiple lymphohematopoietic lineages and induces myeloblast transformation in synergy with ICSBP deficiency. J. Exp. Med. 2002, 196, 1227–1240. [Google Scholar] [CrossRef]
- Mastio, J.; Condamine, T.; Dominguez, G.; Kossenkov, A.V.; Donthireddy, L.; Veglia, F.; Lin, C.; Wang, F.; Fu, S.; Zhou, J.; et al. Identification of monocyte-like precursors of granulocytes in cancer as a mechanism for accumulation of PMN-MDSCs. J. Exp. Med. 2019, 216, 2150–2169. [Google Scholar] [CrossRef]
- Marquis, J.F.; Kapoustina, O.; Langlais, D.; Ruddy, R.; Dufour, C.R.; Kim, B.H.; MacMicking, J.D.; Giguere, V.; Gros, P. Interferon regulatory factor 8 regulates pathways for antigen presentation in myeloid cells and during tuberculosis. PLoS Genet. 2011, 7, e1002097. [Google Scholar] [CrossRef]
- Wang, H.; Lee, C.H.; Qi, C.; Tailor, P.; Feng, J.; Abbasi, S.; Atsumi, T.; Morse, H.C., 3rd. IRF8 regulates B-cell lineage specification, commitment, and differentiation. Blood 2008, 112, 4028–4038. [Google Scholar] [CrossRef]
- Xu, H.; Chaudhri, V.K.; Wu, Z.; Biliouris, K.; Dienger-Stambaugh, K.; Rochman, Y.; Singh, H. Regulation of bifurcating B cell trajectories by mutual antagonism between transcription factors IRF4 and IRF8. Nat. Immunol. 2015, 16, 1274–1281. [Google Scholar] [CrossRef]
- Wang, H.; Jain, S.; Li, P.; Lin, J.X.; Oh, J.; Qi, C.; Gao, Y.; Sun, J.; Sakai, T.; Naghashfar, Z.; et al. Transcription factors IRF8 and PU.1 are required for follicular B cell development and BCL6-driven germinal center responses. Proc. Natl. Acad. Sci. USA 2019, 116, 9511–9520. [Google Scholar] [CrossRef]
- Siegal, F.P.; Kadowaki, N.; Shodell, M.; Fitzgerald-Bocarsly, P.A.; Shah, K.; Ho, S.; Antonenko, S.; Liu, Y.-J. The nature of the principal type 1 interferon-producing cells in human blood. Science 1999, 284, 1835–1837. [Google Scholar] [CrossRef] [PubMed]
- Cella, M.; Jarrossay, D.; Facchetti, F.; Alebardi, O.; Nakajima, H.; Lanzavecchia, A.; Colonna, M. Plasmacytoid monocytes migrate to inflamed lymph nodes and produce large amounts of type I interferon. Nat. Med. 1999, 5, 919–923. [Google Scholar] [CrossRef] [PubMed]
- Reizis, B.; Colonna, M.; Trinchieri, G.; Barrat, F.; Gilliet, M. Plasmacytoid dendritic cells: One-trick ponies or workhorses of the immune system? Nat. Rev. Immunol. 2011, 11, 558–565. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, K.S.; Das, A.; Jaiswal, H.; Saha, I.; Kaushik, M.; Patel, V.K.; Tailor, P. IRF8 and BATF3 interaction enhances the cDC1 specific Pfkfb3 gene expression. Cell Immunol. 2022, 371, 104468. [Google Scholar] [CrossRef]
- Anderson, D.A.; Ou, F.; Kim, S.; Murphy, T.L.; Murphy, K.M. Transition from cMyc to L-Myc during dendritic cell development coordinated by rising levels of IRF8. J. Exp. Med. 2022, 219, e20211483. [Google Scholar] [CrossRef]
- Zhang, S.; Coughlan, H.D.; Ashayeripanah, M.; Seizova, S.; Kueh, A.J.; Brown, D.V.; Cao, W.; Jacquelot, N.; D’Amico, A.; Lew, A.M.; et al. Type 1 conventional dendritic cell fate and function are controlled by DC-SCRIPT. Sci. Immunol. 2021, 6, eabf4432. [Google Scholar] [CrossRef]
- Sichien, D.; Scott, C.L.; Martens, L.; Vanderkerken, M.; Van Gassen, S.; Plantinga, M.; Joeris, T.; De Prijck, S.; Vanhoutte, L.; Vanheerswynghels, M.; et al. IRF8 Transcription Factor Controls Survival and Function of Terminally Differentiated Conventional and Plasmacytoid Dendritic Cells, Respectively. Immunity 2016, 45, 626–640. [Google Scholar] [CrossRef]
- Schönheit, J.; Kuhl, C.; Gebhardt Marie, L.; Klett Francisco, F.; Riemke, P.; Scheller, M.; Huang, G.; Naumann, R.; Leutz, A.; Stocking, C.; et al. PU.1 Level-Directed Chromatin Structure Remodeling at the Irf8 Gene Drives Dendritic Cell Commitment. Cell Rep. 2013, 3, 1617–1628. [Google Scholar] [CrossRef]
- Kurotaki, D.; Kawase, W.; Sasaki, H.; Nakabayashi, J.; Nishiyama, A.; Morse, H.C., III; Ozato, K.; Suzuki, Y.; Tamura, T. Epigenetic control of early dendritic cell lineage specification by the transcription factor IRF8 in mice. Blood 2019, 133, 1803–1813. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, T.; Hotta, C.; Koizumi, S.-I.; Miyashita, K.; Nakabayashi, J.; Kurotaki, D.; Sato, G.R.; Yamamoto, M.; Nakazawa, M.; Fujita, H.; et al. The Transcription Factor IRF8 Counteracts BCR-ABL to Rescue Dendritic Cell Development in Chronic Myelogenous Leukemia. Cancer Res. 2013, 73, 6642–6653. [Google Scholar] [CrossRef] [PubMed]
- Clément, M.; Haddad, Y.; Raffort, J.; Lareyre, F.; Newland, S.A.; Master, L.; Harrison, J.; Ozsvar-Kozma, M.; Bruneval, P.; Binder, C.J.; et al. Deletion of IRF8 (Interferon Regulatory Factor 8)-Dependent Dendritic Cells Abrogates Proatherogenic Adaptive Immunity. Circ. Res. 2018, 122, 813–820. [Google Scholar] [CrossRef] [PubMed]
- Meyer, M.A.; Baer, J.M.; Knolhoff, B.L.; Nywening, T.M.; Panni, R.Z.; Su, X.; Weilbaecher, K.N.; Hawkins, W.G.; Ma, C.; Fields, R.C.; et al. Breast and pancreatic cancer interrupt IRF8-dependent dendritic cell development to overcome immune surveillance. Nat. Commun. 2018, 9, 1250. [Google Scholar] [CrossRef]
- Kuhn, N.F.; Lopez, A.V.; Li, X.; Cai, W.; Daniyan, A.F.; Brentjens, R.J. CD103(+) cDC1 and endogenous CD8(+) T cells are necessary for improved CD40L-overexpressing CAR T cell antitumor function. Nat. Commun. 2020, 11, 6171. [Google Scholar] [CrossRef]
- Tamura, T.; Nagamura-Inoue, T.; Shmeltzer, Z.; Kuwata, T.; Ozato, K. ICSBP directs bipotential myeloid progenitor cells to differentiate into mature macrophages. Immunity 2000, 13, 155–165. [Google Scholar] [CrossRef]
- Li, L.; Jin, H.; Xu, J.; Shi, Y.; Wen, Z. Irf8 regulates macrophage versus neutrophil fate during zebrafish primitive myelopoiesis. Blood 2011, 117, 1359–1369. [Google Scholar] [CrossRef]
- Hagemeyer, N.; Kierdorf, K.; Frenzel, K.; Xue, J.; Ringelhan, M.; Abdullah, Z.; Godin, I.; Wieghofer, P.; Costa Jordão, M.J.; Ulas, T.; et al. Transcriptome-based profiling of yolk sac-derived macrophages reveals a role for Irf8 in macrophage maturation. EMBO J. 2016, 35, 1730–1744. [Google Scholar] [CrossRef]
- Yao, Z.; Getting, S.J.; Locke, I.C. Regulation of TNF-Induced Osteoclast Differentiation. Cells 2021, 11, 132. [Google Scholar] [CrossRef]
- Scharton-Kersten, T.; Contursi, C.; Masumi, A.; Sher, A.; Ozato, K. Interferon Consensus Sequence Binding Protein–deficient Mice Display Impaired Resistance to Intracellular Infection Due to a Primary Defect in Interleukin 12 p40 Induction. J. Exp. Med. 1997, 186, 1523–1534. [Google Scholar] [CrossRef]
- Masumi, A.; Tamaoki, S.; Wang, I.-M.; Ozato, K.; Komuro, K. IRF-8/ICSBP and IRF-1 cooperatively stimulate mouse IL-12 promoter activity in macrophages. FEBS Lett. 2002, 531, 348–353. [Google Scholar] [CrossRef]
- Gupta, M.; Shin, D.-M.; Ramakrishna, L.; Goussetis, D.J.; Platanias, L.C.; Xiong, H.; Morse Iii, H.C.; Ozato, K. IRF8 directs stress-induced autophagy in macrophages and promotes clearance of Listeria monocytogenes. Nat. Commun. 2015, 6, 6379. [Google Scholar] [CrossRef]
- Alter-Koltunoff, M.; Goren, S.; Nousbeck, J.; Feng, C.G.; Sher, A.; Ozato, K.; Azriel, A.; Levi, B.-Z. Innate Immunity to Intraphagosomal Pathogens Is Mediated by Interferon Regulatory Factor 8 (IRF-8) That Stimulates the Expression of Macrophage-specific Nramp1 through Antagonizing Repression by c-Myc. J. Biol. Chem. 2008, 283, 2724–2733. [Google Scholar] [CrossRef] [PubMed]
- Karki, R.; Lee, E.; Place, D.; Samir, P.; Mavuluri, J.; Sharma, B.R.; Balakrishnan, A.; Malireddi, R.K.S.; Geiger, R.; Zhu, Q.; et al. IRF8 Regulates Transcription of Naips for NLRC4 Inflammasome Activation. Cell 2018, 173, 920–933.e13. [Google Scholar] [CrossRef]
- Yáñez, A.; Ng, M.Y.; Hassanzadeh-Kiabi, N.; Goodridge, H.S. IRF8 acts in lineage-committed rather than oligopotent progenitors to control neutrophil vs monocyte production. Blood 2015, 125, 1452–1459. [Google Scholar] [CrossRef]
- Terry, R.L.; Deffrasnes, C.; Getts, D.R.; Minten, C.; Van Vreden, C.; Ashhurst, T.M.; Getts, M.T.; Xie, R.D.V.; Campbell, I.L.; King, N.J. Defective inflammatory monocyte development in IRF8-deficient mice abrogates migration to the West Nile virus-infected brain. J. Innate Immun. 2015, 7, 102–112. [Google Scholar] [CrossRef] [PubMed]
- Dimberg, A.; Kårehed, K.; Nilsson, K.; Öberg, F. Inhibition of Monocytic Differentiation by Phosphorylation-deficient Stat1 is Associated with Impaired Expression of Stat2, ICSBP/IRF8 and C/EBPɛ. Scand. J. Immunol. 2006, 64, 271–279. [Google Scholar] [CrossRef]
- Zhao, Y.; Shen, X.; Na, N.; Chu, Z.; Su, H.; Chao, S.; Shi, L.; Xu, Y.; Zhang, L.; Shi, B.; et al. mTOR masters monocyte development in bone marrow by decreasing the inhibition of STAT5 on IRF8. Blood 2018, 131, 1587–1599. [Google Scholar] [CrossRef]
- Matikainen, S.; Ronni, T.; Lehtonen, A.; Sareneva, T.; Melén, K.; Nordling, S.; Levy, D.E.; Julkunen, I. Retinoic acid induces signal transducer and activator of transcription (STAT) 1, STAT2, and p48 expression in myeloid leukemia cells and enhances their responsiveness to interferons. Cell Growth Differ.-Publ. Am. Assoc. Cancer Res. 1997, 8, 687–698. [Google Scholar]
- Holtschke, T.; Lohler, J.; Kanno, Y.; Fehr, T.; Giese, N.; Rosenbauer, F.; Lou, J.; Knobeloch, K.P.; Gabriele, L.; Waring, J.F.; et al. Immunodeficiency and chronic myelogenous leukemia-like syndrome in mice with a targeted mutation of the ICSBP gene. Cell 1996, 87, 307–317. [Google Scholar] [CrossRef]
- Waight, J.D.; Netherby, C.; Hensen, M.L.; Miller, A.; Hu, Q.; Liu, S.; Bogner, P.N.; Farren, M.R.; Lee, K.P.; Liu, K.; et al. Myeloid-derived suppressor cell development is regulated by a STAT/IRF-8 axis. J. Clin. Investig. 2013, 123, 4464–4478. [Google Scholar] [CrossRef] [PubMed]
- Redd, P.S.; Ibrahim, M.L.; Klement, J.D.; Sharman, S.K.; Paschall, A.V.; Yang, D.; Nayak-Kapoor, A.; Liu, K. SETD1B Activates iNOS Expression in Myeloid-Derived Suppressor Cells. Cancer Res. 2017, 77, 2834–2843. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Salazar, E.G.; Almeraya, E.V.; Lopez-Perez, T.V.; Jimenez-Salas, Z.; Patino, N.; Velazquez-Cruz, R. MicroRNA-1270 Inhibits Cell Proliferation, Migration, and Invasion via Targeting IRF8 in Osteoblast-like Cell Lines. Curr. Issues Mol. Biol. 2022, 44, 1182–1190. [Google Scholar] [CrossRef]
- Xiong, H.; Li, H.; Kong, H.J.; Chen, Y.; Zhao, J.; Xiong, S.; Huang, B.; Gu, H.; Mayer, L.; Ozato, K.; et al. Ubiquitin-dependent degradation of interferon regulatory factor-8 mediated by Cbl down-regulates interleukin-12 expression. J. Biol. Chem. 2005, 280, 23531–23539. [Google Scholar] [CrossRef] [PubMed]
- Unlu, S.; Kumar, A.; Waterman, W.R.; Tsukada, J.; Wang, K.Z.; Galson, D.L.; Auron, P.E. Phosphorylation of IRF8 in a pre-associated complex with Spi-1/PU.1 and non-phosphorylated Stat1 is critical for LPS induction of the IL1B gene. Mol. Immunol. 2007, 44, 3364–3379. [Google Scholar] [CrossRef]
- Gabrilovich, D.I.; Nagaraj, S. Myeloid-derived suppressor cells as regulators of the immune system. Nat. Rev. Immunol. 2009, 9, 162–174. [Google Scholar] [CrossRef]
- Patel, A.; Oba, T.; Kajihara, R.; Yokoi, T.; Abrams, S.I.; Ito, F. Multimodal Intralesional Therapy for Reshaping the Myeloid Compartment of Tumors Resistant to Anti-PD-L1 Therapy via IRF8 Expression. J. Immunol. 2021, 207, 1298–1309. [Google Scholar] [CrossRef]
- Krishnamoorthy, M.; Gerhardt, L.; Maleki Vareki, S. Immunosuppressive Effects of Myeloid-Derived Suppressor Cells in Cancer and Immunotherapy. Cells 2021, 10, 1170. [Google Scholar] [CrossRef]
- Ma, T.; Renz, B.W.; Ilmer, M.; Koch, D.; Yang, Y.; Werner, J.; Bazhin, A.V. Myeloid-Derived Suppressor Cells in Solid Tumors. Cells 2022, 11, 310. [Google Scholar] [CrossRef]
- Condamine, T.; Mastio, J.; Gabrilovich, D.I. Transcriptional regulation of myeloid-derived suppressor cells. J. Leukoc. Biol. 2015, 98, 913–922. [Google Scholar] [CrossRef]
- Youn, J.-I.; Nagaraj, S.; Collazo, M.; Gabrilovich, D.I. Subsets of Myeloid-Derived Suppressor Cells in Tumor-Bearing Mice. J. Immunol. 2008, 181, 5791–5802. [Google Scholar] [CrossRef] [PubMed]
- Netherby, C.S.; Messmer, M.N.; Burkard-Mandel, L.; Colligan, S.; Miller, A.; Cortes Gomez, E.; Wang, J.; Nemeth, M.J.; Abrams, S.I. The Granulocyte Progenitor Stage Is a Key Target of IRF8-Mediated Regulation of Myeloid-Derived Suppressor Cell Production. J. Immunol. 2017, 198, 4129–4139. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.D.; Lu, C.; Payne, D.; Paschall, A.V.; Klement, J.D.; Redd, P.S.; Ibrahim, M.L.; Yang, D.; Han, Q.; Liu, Z.; et al. Autocrine IL6-Mediated Activation of the STAT3-DNMT Axis Silences the TNFalpha-RIP1 Necroptosis Pathway to Sustain Survival and Accumulation of Myeloid-Derived Suppressor Cells. Cancer Res. 2020, 80, 3145–3156. [Google Scholar] [CrossRef] [PubMed]
- Nelson, N.; Kanno, Y.; Hong, C.; Contursi, C.; Fujita, T.; Fowlkes, B.J.; O’Connell, E.; Hu-Li, J.; Paul, W.E.; Jankovic, D.; et al. Expression of IFN regulatory factor family proteins in lymphocytes. Induction of Stat-1 and IFN consensus sequence binding protein expression by T cell activation. J. Immunol. 1996, 156, 3711–3720. [Google Scholar] [PubMed]
- Zhan, F.; Tian, E.; Bumm, K.; Smith, R.; Barlogie, B.; Shaughnessy, J., Jr. Gene expression profiling of human plasma cell differentiation and classification of multiple myeloma based on similarities to distinct stages of late-stage B-cell development. Blood J. Am. Soc. Hematol. 2003, 101, 1128–1140. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H.; Melchers, M.; Wang, H.; Torrey, T.A.; Slota, R.; Qi, C.-F.; Kim, J.Y.; Lugar, P.; Kong, H.J.; Farrington, L.; et al. Regulation of the germinal center gene program by interferon (IFN) regulatory factor 8/IFN consensus sequence-binding protein. J. Exp. Med. 2005, 203, 63–72. [Google Scholar] [CrossRef]
- Qi, C.-F.; Li, Z.; Raffeld, M.; Wang, H.; Kovalchuk, A.L.; Morse, H.C. Differential expression of IRF8 in subsets of macrophages and dendritic cells and effects of IRF8 deficiency on splenic B cell and macrophage compartments. Immunol. Res. 2009, 45, 62–74. [Google Scholar] [CrossRef]
- Feng, J.; Wang, H.; Shin, D.M.; Masiuk, M.; Qi, C.F.; Morse, H.C., 3rd. IFN regulatory factor 8 restricts the size of the marginal zone and follicular B cell pools. J. Immunol. 2011, 186, 1458–1466. [Google Scholar] [CrossRef]
- Tang, W.Y.; Zhang, Y.H.; Zhang, Y.S.; Liao, Y.; Luo, J.S.; Liu, J.H.; Peng, C.J.; Tang, Y.L.; Huang, D.P.; Sun, X.; et al. Abnormal thymic B cell activation and impaired T cell differentiation in pristane-induced lupus mice. Immunol. Lett. 2021, 231, 49–60. [Google Scholar] [CrossRef]
- Carotta, S.; Willis, S.N.; Hasbold, J.; Inouye, M.; Pang, S.H.M.; Emslie, D.; Light, A.; Chopin, M.; Shi, W.; Wang, H.; et al. The transcription factors IRF8 and PU.1 negatively regulate plasma cell differentiation. J. Exp. Med. 2014, 211, 2169–2181. [Google Scholar] [CrossRef]
- Choi, J.K.; Yu, C.R.; Bing, S.J.; Jittayasothorn, Y.; Mattapallil, M.J.; Kang, M.; Park, S.B.; Lee, H.S.; Dong, L.; Shi, G.; et al. IL-27-producing B-1a cells suppress neuroinflammation and CNS autoimmune diseases. Proc. Natl. Acad. Sci. USA 2021, 118, e2109548118. [Google Scholar] [CrossRef] [PubMed]
- Bouamar, H.; Abbas, S.; Lin, A.-P.; Wang, L.; Jiang, D.; Holder, K.N.; Kinney, M.C.; Hunicke-Smith, S.; Aguiar, R.C.T. A capture-sequencing strategy identifies IRF8, EBF1, and APRIL as novel IGH fusion partners in B-cell lymphoma. Blood 2013, 122, 726–733. [Google Scholar] [CrossRef] [PubMed]
- Pang, S.H.M.; Minnich, M.; Gangatirkar, P.; Zheng, Z.; Ebert, A.; Song, G.; Dickins, R.A.; Corcoran, L.M.; Mullighan, C.G.; Busslinger, M.; et al. PU.1 cooperates with IRF4 and IRF8 to suppress pre-B-cell leukemia. Leukemia 2016, 30, 1375–1387. [Google Scholar] [CrossRef]
- McQuaid, D.C.; Katz, S.G.; Xu, M.L. IRF8 as a Novel Marker to Differentiate Between CD30-Positive Large Cell Lymphomas. Am. J. Clin. Pathol. 2022, 158, 173–176. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.X.; Lee, C.H.; Qi, C.F.; Wang, H.; Naghashfar, Z.; Abbasi, S.; Morse, H.C. IFN Regulatory Factor 8 Regulates MDM2 in Germinal Center B Cells. J. Immunol. 2009, 183, 3188–3194. [Google Scholar] [CrossRef]
- Zhong, W.; Xu, X.; Zhu, Z.; Du, Q.; Du, H.; Yang, L.; Ling, Y.; Xiong, H.; Li, Q. Increased expression of IRF8 in tumor cells inhibits the generation of Th17 cells and predicts unfavorable survival of diffuse large B cell lymphoma patients. Oncotarget 2017, 8, 49757–49772. [Google Scholar] [CrossRef]
- Ouyang, X.; Zhang, R.; Yang, J.; Li, Q.; Qin, L.; Zhu, C.; Liu, J.; Ning, H.; Shin, M.S.; Gupta, M.; et al. Transcription factor IRF8 directs a silencing programme for TH17 cell differentiation. Nat. Commun. 2011, 2, 314. [Google Scholar] [CrossRef]
- Newman, D.M.; Leung, P.S.; Putoczki, T.L.; Nutt, S.L.; Cretney, E. Th17 cell differentiation proceeds independently of IRF8. Immunol. Cell Biol. 2016, 94, 796–801. [Google Scholar] [CrossRef]
- Veldhoen, M.; Hocking, R.J.; Atkins, C.J.; Locksley, R.M.; Stockinger, B. TGFβ in the context of an inflammatory cytokine milieu supports de novo differentiation of IL-17-producing T cells. Immunity 2006, 24, 179–189. [Google Scholar] [CrossRef]
- Ivanov, I.I.; McKenzie, B.S.; Zhou, L.; Tadokoro, C.E.; Lepelley, A.; Lafaille, J.J.; Cua, D.J.; Littman, D.R. The orphan nuclear receptor RORγt directs the differentiation program of proinflammatory IL-17+ T helper cells. Cell 2006, 126, 1121–1133. [Google Scholar] [CrossRef]
- Lehtonen, A.; Lund, R.; Lahesmaa, R.; Julkunen, I.; Sareneva, T.; Matikainen, S. IFN-α and IL-12 activate IFN regulatory factor 1 (IRF-1), IRF-4, and IRF-8 gene expression in human NK and T cells. Cytokine 2003, 24, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Wang, I.-M.; Contursi, C.; Masumi, A.; Ma, X.; Trinchieri, G.; Ozato, K. An IFN-γ-inducible transcription factor, IFN consensus sequence binding protein (ICSBP), stimulates IL-12 p40 expression in macrophages. J. Immunol. 2000, 165, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Purwar, R.; Schlapbach, C.; Xiao, S.; Kang, H.S.; Elyaman, W.; Jiang, X.; Jetten, A.M.; Khoury, S.J.; Fuhlbrigge, R.C.; Kuchroo, V.K. Robust tumor immunity to melanoma mediated by interleukin-9–producing T cells. Nat. Med. 2012, 18, 1248–1253. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Hong, S.; Li, H.; Park, J.; Hong, B.; Wang, L.; Zheng, Y.; Liu, Z.; Xu, J.; He, J. Th9 cells promote antitumor immune responses in vivo. J. Clin. Investig. 2012, 122, 4160–4171. [Google Scholar] [CrossRef]
- Végran, F.; Berger, H.; Boidot, R.; Mignot, G.; Bruchard, M.; Dosset, M.; Chalmin, F.; Rébé, C.; Dérangère, V.; Ryffel, B. The transcription factor IRF1 dictates the IL-21-dependent anticancer functions of TH 9 cells. Nat. Immunol. 2014, 15, 758–766. [Google Scholar] [CrossRef]
- Miyagawa, F.; Zhang, H.; Terunuma, A.; Ozato, K.; Tagaya, Y.; Katz, S.I. Interferon regulatory factor 8 integrates T-cell receptor and cytokine-signaling pathways and drives effector differentiation of CD8 T cells. Proc. Natl. Acad. Sci. USA 2012, 109, 12123–12128. [Google Scholar] [CrossRef]
- Li, D.; Zhang, Y.; Qiu, Q.; Wang, J.; Zhao, X.; Jiao, B.; Zhang, X.; Yu, S.; Xu, P.; Dan, Y.; et al. IRF8 Impacts Self-Renewal of Hematopoietic Stem Cells by Regulating TLR9 Signaling Pathway of Innate Immune Cells. Adv. Sci. 2021, 8, e2101031. [Google Scholar] [CrossRef]
- Mace, E.M.; Bigley, V.; Gunesch, J.T.; Chinn, I.K.; Angelo, L.S.; Care, M.A.; Maisuria, S.; Keller, M.D.; Togi, S.; Watkin, L.B. Biallelic mutations in IRF8 impair human NK cell maturation and function. J. Clin. Investig. 2017, 127, 306–320. [Google Scholar] [CrossRef]
- Bigley, V.; Maisuria, S.; Cytlak, U.; Jardine, L.; Care, M.A.; Green, K.; Gunawan, M.; Milne, P.; Dickinson, R.; Wiscombe, S. Biallelic interferon regulatory factor 8 mutation: A complex immunodeficiency syndrome with dendritic cell deficiency, monocytopenia, and immune dysregulation. J. Allergy Clin. Immunol. 2018, 141, 2234–2248. [Google Scholar] [CrossRef]
- Adams, N.M.; Lau, C.M.; Fan, X.; Rapp, M.; Geary, C.D.; Weizman, O.E.; Diaz-Salazar, C.; Sun, J.C. Transcription Factor IRF8 Orchestrates the Adaptive Natural Killer Cell Response. Immunity 2018, 48, 1172–1182. [Google Scholar] [CrossRef]
- Rajabi, H.; Tagde, A.; Alam, M.; Bouillez, A.; Pitroda, S.; Suzuki, Y.; Kufe, D. DNA methylation by DNMT1 and DNMT3b methyltransferases is driven by the MUC1-C oncoprotein in human carcinoma cells. Oncogene 2016, 35, 6439–6445. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Xiong, X.; Shao, Q.; Xiang, T.; Li, L.; Yin, X.; Li, X.; Tao, Q.; Ren, G. The tumor suppressor interferon regulatory factor 8 inhibits beta-catenin signaling in breast cancers, but is frequently silenced by promoter methylation. Oncotarget 2017, 8, 48875–48888. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, M.; Toyota, M.; Suzuki, H.; Nojima, M.; Yamamoto, E.; Kamimae, S.; Watanabe, Y.; Kai, M.; Akashi, H.; Maruyama, R. DNA methylation of interferon regulatory factors in gastric cancer and noncancerous gastric mucosae. Cancer Sci. 2010, 101, 1708–1716. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Geng, H.; Ng, K.; Yu, J.; Van Hasselt, A.; Cao, Y.; Zeng, Y.; Wong, A.; Wang, X.; Ying, J. Epigenetic disruption of interferon-γ response through silencing the tumor suppressor interferon regulatory factor 8 in nasopharyngeal, esophageal and multiple other carcinomas. Oncogene 2008, 27, 5267–5276. [Google Scholar] [CrossRef]
- Ye, L.; Xiang, T.; Zhu, J.; Li, D.; Shao, Q.; Peng, W.; Tang, J.; Li, L.; Ren, G. Interferon consensus sequence-binding protein 8, a tumor suppressor, suppresses tumor growth and invasion of non-small cell lung cancer by interacting with the wnt/β-catenin pathway. Cell. Physiol. Biochem. 2018, 51, 961–978. [Google Scholar] [CrossRef]
- Wu, H.; You, L.; Li, Y.; Zhao, Z.; Shi, G.; Chen, Z.; Wang, Z.; Li, X.; Du, S.; Ye, W.; et al. Loss of a Negative Feedback Loop between IRF8 and AR Promotes Prostate Cancer Growth and Enzalutamide Resistance. Cancer Res. 2020, 80, 2927–2939. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhang, L.; Li, L.; Wang, Z.; Ying, J.; Fan, Y.; Xu, B.; Wang, L.; Liu, Q.; Chen, G.; et al. Interferon regulatory factor 8 functions as a tumor suppressor in renal cell carcinoma and its promoter methylation is associated with patient poor prognosis. Cancer Lett. 2014, 354, 227–234. [Google Scholar] [CrossRef]
- Zilionis, R.; Engblom, C.; Pfirschke, C.; Savova, V.; Zemmour, D.; Saatcioglu, H.D.; Krishnan, I.; Maroni, G.; Meyerovitz, C.V.; Kerwin, C.M.; et al. Single-Cell Transcriptomics of Human and Mouse Lung Cancers Reveals Conserved Myeloid Populations across Individuals and Species. Immunity 2019, 50, 1317–1334.e10. [Google Scholar] [CrossRef]
- Mehner, C.; Miller, E.; Nassar, A.; Bamlet, W.R.; Radisky, E.S.; Radisky, D.C. Tumor cell expression of MMP3 as a prognostic factor for poor survival in pancreatic, pulmonary, and mammary carcinoma. Genes Cancer 2015, 6, 480. [Google Scholar] [CrossRef]
- Mendes, O.; Kim, H.-T.; Stoica, G. Expression of MMP2, MMP9 and MMP3 in breast cancer brain metastasis in a rat model. Clin. Exp. Metastasis 2005, 22, 237–246. [Google Scholar] [CrossRef]
- Shoshan, E.; Braeuer, R.R.; Kamiya, T.; Mobley, A.K.; Huang, L.; Vasquez, M.E.; Velazquez-Torres, G.; Chakravarti, N.; Ivan, C.; Prieto, V. NFAT1 directly regulates IL8 and MMP3 to promote melanoma tumor growth and metastasis. Cancer Res. 2016, 76, 3145–3155. [Google Scholar] [CrossRef] [PubMed]
- Banik, D.; Netherby, C.S.; Bogner, P.N.; Abrams, S.I. MMP3-mediated tumor progression is controlled transcriptionally by a novel IRF8-MMP3 interaction. Oncotarget 2015, 6, 15164–15179. [Google Scholar] [CrossRef] [PubMed]
- Sung, J.Y.; Park, S.Y.; Kim, J.H.; Kang, H.G.; Yoon, J.H.; Na, Y.S.; Kim, Y.N.; Park, B.K. Interferon consensus sequence-binding protein (ICSBP) promotes epithelial-to-mesenchymal transition (EMT)-like phenomena, cell-motility, and invasion via TGF-β signaling in U2OS cells. Cell Death Dis. 2014, 5, e1224. [Google Scholar] [CrossRef] [PubMed]
- Egwuagu, C.E.; Li, W.; Yu, C.R.; Che Mei Lin, M.; Chan, C.C.; Nakamura, T.; Chepelinsky, A.B. Interferon-gamma induces regression of epithelial cell carcinoma: Critical roles of IRF-1 and ICSBP transcription factors. Oncogene 2006, 25, 3670–3679. [Google Scholar] [CrossRef]
- Villa-Morales, M.; Fernandez-Piqueras, J. Targeting the Fas/FasL signaling pathway in cancer therapy. Expert Opin. Ther. Targets 2012, 16, 85–101. [Google Scholar] [CrossRef]
- Mollinedo, F.; Gajate, C. Fas/CD95 death receptor and lipid rafts: New targets for apoptosis-directed cancer therapy. Drug Resist. Updates 2006, 9, 51–73. [Google Scholar] [CrossRef]
- Merting, A.D.; Poschel, D.B.; Lu, C.; Klement, J.D.; Yang, D.; Li, H.; Shi, H.; Chapdelaine, E.; Montgomery, M.; Redman, M.T.; et al. Restoring FAS Expression via Lipid-Encapsulated FAS DNA Nanoparticle Delivery Is Sufficient to Suppress Colon Tumor Growth In Vivo. Cancers 2022, 14, 361. [Google Scholar] [CrossRef]
- Xiao, W.; Ibrahim, M.L.; Redd, P.S.; Klement, J.D.; Lu, C.; Yang, D.; Savage, N.M.; Liu, K. Loss of Fas expression and function is coupled with colon cancer resistance to immune checkpoint inhibitor immunotherapy. Mol. Cancer Res. 2019, 17, 420–430. [Google Scholar] [CrossRef]
- Yang, D.; Wang, S.; Brooks, C.; Dong, Z.; Schoenlein, P.V.; Kumar, V.; Ouyang, X.; Xiong, H.; Lahat, G.; Hayes-Jordan, A.; et al. IFN regulatory factor 8 sensitizes soft tissue sarcoma cells to death receptor-initiated apoptosis via repression of FLICE-like protein expression. Cancer Res. 2009, 69, 1080–1088. [Google Scholar] [CrossRef]
- Yang, D.; Thangaraju, M.; Browning, D.D.; Dong, Z.; Korchin, B.; Lev, D.C.; Ganapathy, V.; Liu, K. IFN Regulatory Factor 8 Mediates Apoptosis in Nonhemopoietic Tumor Cells via Regulation of Fas Expression. J. Immunol. 2007, 179, 4775–4782. [Google Scholar] [CrossRef]
- Seelan, R.S.; Qian, C.; Yokomizo, A.; Bostwick, D.G.; Smith, D.I.; Liu, W. Human acid ceramidase is overexpressed but not mutated in prostate cancer. Genes Chromosomes Cancer 2000, 29, 137–146. [Google Scholar] [CrossRef]
- Saad, A.F.; Meacham, W.D.; Bai, A.; Anelli, V.; Anelli, V.; Mahdy, A.E.; Turner, L.S.; Cheng, J.; Bielawska, A.; Bielawski, J. The functional effects of acid ceramidase over-expression in prostate cancer progression and resistance to chemotherapy. Cancer Biol. Ther. 2007, 6, 1451–1456. [Google Scholar] [CrossRef]
- Lucki, N.C.; Sewer, M.B. Genistein stimulates MCF-7 breast cancer cell growth by inducing acid ceramidase (ASAH1) gene expression. J. Biol. Chem. 2011, 286, 19399–19409. [Google Scholar] [CrossRef]
- Mattei, F.; Schiavoni, G.; De Ninno, A.; Lucarini, V.; Sestili, P.; Sistigu, A.; Fragale, A.; Sanchez, M.; Spada, M.; Gerardino, A. A multidisciplinary study using in vivo tumor models and microfluidic cell-on-chip approach to explore the cross-talk between cancer and immune cells. J. Immunotoxicol. 2014, 11, 337–346. [Google Scholar] [CrossRef]
- Klement, J.D.; Poschel, D.B.; Lu, C.; Merting, A.D.; Yang, D.; Redd, P.S.; Liu, K. Osteopontin Blockade Immunotherapy Increases Cytotoxic T Lymphocyte Lytic Activity and Suppresses Colon Tumor Progression. Cancers 2021, 13, 1006. [Google Scholar] [CrossRef]
- Klement, J.D.; Paschall, A.V.; Redd, P.S.; Ibrahim, M.L.; Lu, C.; Yang, D.; Celis, E.; Abrams, S.I.; Ozato, K.; Liu, K. An osteopontin/CD44 immune checkpoint controls CD8+ T cell activation and tumor immune evasion. J. Clin. Investig. 2018, 128, 5549–5560. [Google Scholar] [CrossRef]
- Lu, C.; Liu, Z.; Klement, J.D.; Yang, D.; Merting, A.D.; Poschel, D.; Albers, T.; Waller, J.L.; Shi, H.; Liu, K. WDR5-H3K4me3 epigenetic axis regulates OPN expression to compensate PD-L1 function to promote pancreatic cancer immune escape. J. Immunother. Cancer 2021, 9, e002624. [Google Scholar] [CrossRef]
- Lee, J.L.; Wang, M.J.; Sudhir, P.R.; Chen, G.D.; Chi, C.W.; Chen, J.Y. Osteopontin promotes integrin activation through outside-in and inside-out mechanisms: OPN-CD44V interaction enhances survival in gastrointestinal cancer cells. Cancer Res. 2007, 67, 2089–2097. [Google Scholar] [CrossRef]
- Nallasamy, P.; Nimmakayala, R.K.; Karmakar, S.; Leon, F.; Seshacharyulu, P.; Lakshmanan, I.; Rachagani, S.; Mallya, K.; Zhang, C.; Ly, Q.P.; et al. Pancreatic Tumor Microenvironment Factor Promotes Cancer Stemness via SPP1-CD44 Axis. Gastroenterology 2021, 161, 1998–2013.e7. [Google Scholar] [CrossRef]
- Qi, J.; Sun, H.; Zhang, Y.; Wang, Z.; Xun, Z.; Li, Z.; Ding, X.; Bao, R.; Hong, L.; Jia, W.; et al. Single-cell and spatial analysis reveal interaction of FAP(+) fibroblasts and SPP1(+) macrophages in colorectal cancer. Nat. Commun. 2022, 13, 1742. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, Q.; Xing, B.; Luo, N.; Gao, R.; Yu, K.; Hu, X.; Bu, Z.; Peng, J.; Ren, X.; et al. Immune phenotypic linkage between colorectal cancer and liver metastasis. Cancer Cell 2022, 40, 424–437.e5. [Google Scholar] [CrossRef]
- Rud, A.K.; Boye, K.; Oijordsbakken, M.; Lund-Iversen, M.; Halvorsen, A.R.; Solberg, S.K.; Berge, G.; Helland, A.; Brustugun, O.T.; Maelandsmo, G.M. Osteopontin is a prognostic biomarker in non-small cell lung cancer. BMC Cancer 2013, 13, 540. [Google Scholar] [CrossRef]
- Rittling, S.R.; Chambers, A.F. Role of osteopontin in tumour progression. Br. J. Cancer 2004, 90, 1877–1881. [Google Scholar] [CrossRef]
- Sreekanthreddy, P.; Srinivasan, H.; Kumar, D.M.; Nijaguna, M.B.; Sridevi, S.; Vrinda, M.; Arivazhagan, A.; Balasubramaniam, A.; Hegde, A.S.; Chandramouli, B.A.; et al. Identification of potential serum biomarkers of glioblastoma: Serum osteopontin levels correlate with poor prognosis. Cancer Epidemiol. Biomark. Prev. 2010, 19, 1409–1422. [Google Scholar] [CrossRef]
- Chakraborty, G.; Jain, S.; Kundu, G.C. Osteopontin promotes vascular endothelial growth factor-dependent breast tumor growth and angiogenesis via autocrine and paracrine mechanisms. Cancer Res. 2008, 68, 152–161. [Google Scholar] [CrossRef]
- Sangaletti, S.; Tripodo, C.; Sandri, S.; Torselli, I.; Vitali, C.; Ratti, C.; Botti, L.; Burocchi, A.; Porcasi, R.; Tomirotti, A.; et al. Osteopontin shapes immunosuppression in the metastatic niche. Cancer Res. 2014, 74, 4706–4719. [Google Scholar] [CrossRef]
- Moorman, H.R.; Poschel, D.; Klement, J.D.; Lu, C.; Redd, P.S.; Liu, K. Osteopontin: A Key Regulator of Tumor Progression and Immunomodulation. Cancers 2020, 12, 3379. [Google Scholar] [CrossRef]
- Coppola, D.; Szabo, M.; Boulware, D.; Muraca, P.; Alsarraj, M.; Chambers, A.F.; Yeatman, T.J. Correlation of osteopontin protein expression and pathological stage across a wide variety of tumor histologies. Clin. Cancer Res. 2004, 10, 184–190. [Google Scholar] [CrossRef]
- Wei, J.; Marisetty, A.; Schrand, B.; Gabrusiewicz, K.; Hashimoto, Y.; Ott, M.; Grami, Z.; Kong, L.Y.; Ling, X.; Caruso, H.; et al. Osteopontin mediates glioblastoma-associated macrophage infiltration and is a potential therapeutic target. J. Clin. Investig. 2019, 129, 137–149. [Google Scholar] [CrossRef]
- Agrawal, D.; Chen, T.; Irby, R.; Quackenbush, J.; Chambers, A.F.; Szabo, M.; Cantor, A.; Coppola, D.; Yeatman, T.J. Osteopontin identified as lead marker of colon cancer progression, using pooled sample expression profiling. J. Natl. Cancer Inst. 2002, 94, 513–521. [Google Scholar] [CrossRef]
- Paschall, A.V.; Yang, D.; Lu, C.; Choi, J.H.; Li, X.; Liu, F.; Figueroa, M.; Oberlies, N.H.; Pearce, C.; Bollag, W.B.; et al. H3K9 Trimethylation Silences Fas Expression to Confer Colon Carcinoma Immune Escape and 5-Fluorouracil Chemoresistance. J. Immunol. 2015, 195, 1868–1882. [Google Scholar] [CrossRef]
- Grzelak, L.; Roesch, F.; Vaysse, A.; Biton, A.; Porrot, F.; Commère, P.-H.; Planchais, C.; Mouquet, H.; Vignuzzi, M.; Bruel, T.; et al. IRF8 regulates efficacy of therapeutic anti-CD20 monoclonal antibodies. bioRxiv 2022. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).