
Theoretical Concepts in Magnetobiology after 40 Years of Research


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
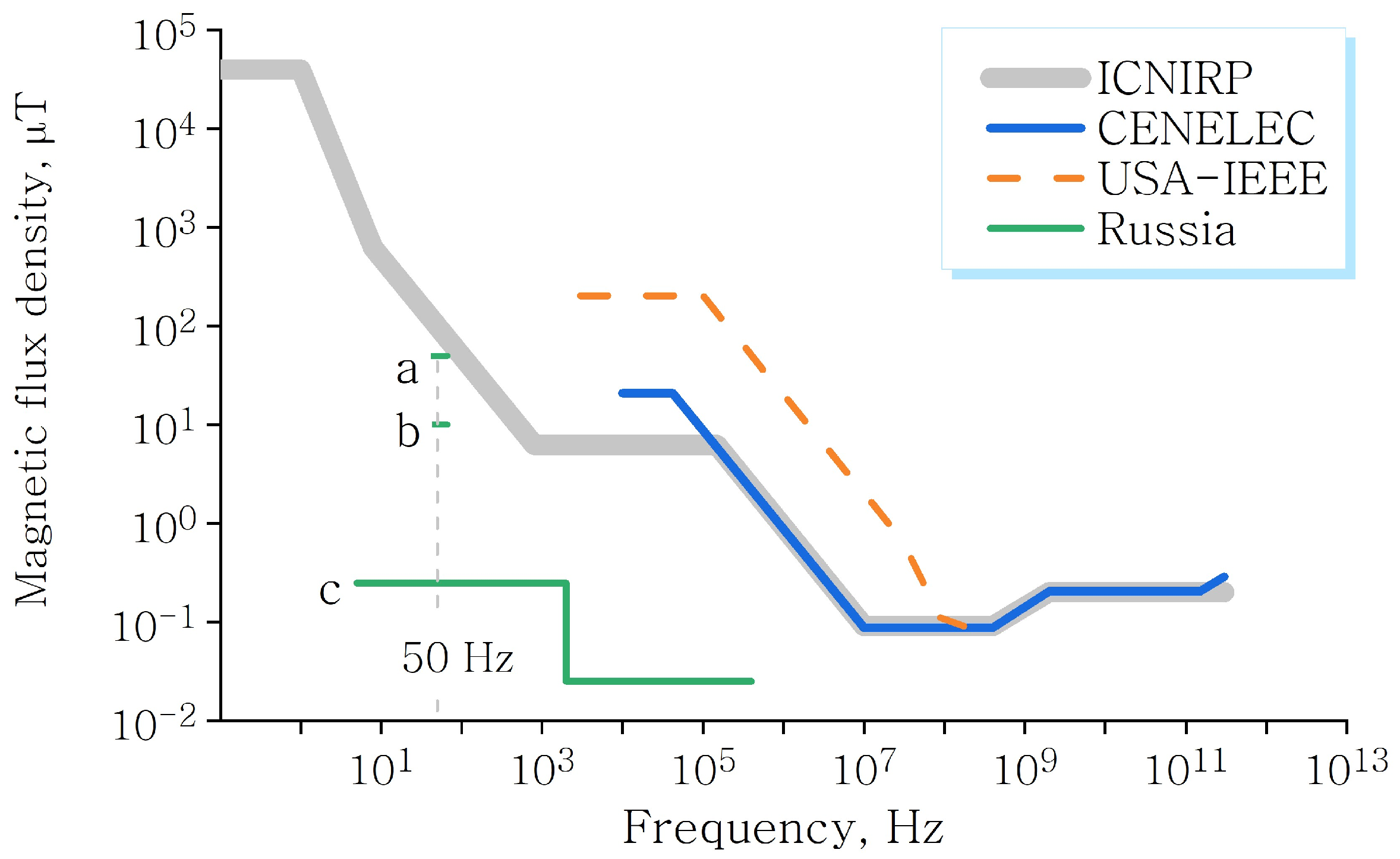
:1. Introduction
2. Phenomena Not Underlying Nonspecific MF Effects
3. Plausible Molecular Mechanisms of Nonspecific MF Effects
- S-T transitions in the spin state of a pair of radicals . Such pairs are in a spin-correlated state which is magnetically sensitive. They arise as intermediates in magnetochemical reactions involving free radicals, for example:and in many enzyme–substrate reactions [39]. The MF, while modulating the S-T conversion, thereby changes the yield of free radicals. The energy of S-T conversion—that of the flip of spins—in the geoMF is orders of magnitude less than the energy of chemical activation; MF, here, is essentially a controlling factor only.
- Precession (in the vector model) of single moments—for example, the spin magnetic moments of electrons. The precession becomes uneven in a variable MF or slows down in a weakened MF [40].
- Interference of angular quantum states or a change in the phase of angular states of rotating molecular groups and electron clouds.
4. Why Are Nonspecific Effects Difficult to Reproduce?
5. Can the Sensitivity of Organisms to Tens of nT Be Explained?
6. Magnetobiological Phenomena in Social Life
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AC | alternating current |
| CENELEC | Comité Européen de Normalisation Électrotechnique |
| DC | direct current |
| EMF | electromagnetic field |
| ICNIRP | the International Commission on Non-Ionizing Radiation Protection |
| IEEE | Institute of Electrical and Electronics Engineers |
| MF | magnetic field |
| S-T | singlet-triplet |
References
- Gilbert, W. De Magnete; Dover Publication Inc.: New York, NY, USA, 1958. [Google Scholar]
- Von Middendorff, A.T. Die Isepiptesen Russlands; Kaiserlichen Akademie der Wissenschaften: St. Petersburg, Russia, 1855. [Google Scholar]
- Schott, H. Zur Geschichte der Elektrotherapie und ihrer Beziehung zum Heilmagnetismus. In Naturheilverfahren und Unkonventionelle Medizinische Richtunge; und Andere, V.F., Ed.; Springer: Berlin/Heidelberg, Germany, 1996. [Google Scholar]
- Kholodov, Y.A. Magnetism in Biology; Nauka: Moscow, Russia, 1970. [Google Scholar]
- Webb, S.J.; Dodds, D.E. Microwave inhibition of bacterial growth. Nature 1968, 218, 374–375. [Google Scholar] [CrossRef]
- Devyatkov, N.D. Influence of millimeter-band electromagnetic radiation on biological objects. Sov. Phys. Uspekhi 1974, 16, 568–569. [Google Scholar] [CrossRef]
- Presman, A.S. Electromagnetic Fields and Life; Plenum: New York, NY, USA, 1970. [Google Scholar]
- Anikin, V.M. Magnetobiology and magnetotherapy: “Karmilov” period. Heteromagn. Microelectron. 2016, 21, 131–140. (In Russian) [Google Scholar]
- Moulder, J.E. The Electric and Magnetic Fields Research and Public Information Dissemination (EMF-RAPID) Program. Radiat. Res. 2000, 153, 613–616. [Google Scholar] [CrossRef]
- IARC. IARC Monographs on the Evaluation of Carcinogenic Risks to Humans, Volume 80. Non-Ionizing Radiation, Part 1: Static and Extremely Low-Frequency (ELF) Electric and Magnetic Fields; IARC Press: Lyon, France, 2002. [Google Scholar]
- Huss, A.; Peters, S.; Vermeulen, R. Occupational exposure to extremely low-frequency magnetic fields and the risk of ALS: A systematic review and meta-analysis. Bioelectromagnetics 2018, 39, 156–163. [Google Scholar] [CrossRef]
- Miller, A.B.; Morgan, L.L.; Udasin, I.; Davis, D.L. Cancer epidemiology update, following the 2011 IARC evaluation of radiofrequency electromagnetic fields (Monograph 102). Environ. Res. 2018, 167, 673–683. [Google Scholar] [CrossRef]
- Li, D.K.; Chen, H.; Ferber, J.R.; Odouli, R.; Quesenberry, C. Exposure to magnetic field non-ionizing radiation and the risk of miscarriage: A prospective cohort study. Sci. Rep. 2017, 7, 17541. [Google Scholar] [CrossRef] [PubMed]
- Belyaev, I.; Dean, A.; Eger, H.; Hubmann, G.; Jandrisovits, R.; Kern, M.; Kundi, M.; Moshammer, H.; Lercher, P.; Müller, K.; et al. EUROPAEM EMF Guideline 2016 for the prevention, diagnosis and treatment of EMF-related health problems and illnesses. Rev. Environ. Health 2016, 31, 363–397. [Google Scholar] [CrossRef] [PubMed]
- Environmental Health Criteria 238. Extremely Low Frequency Fields; World Health Organization: Geneva, Switzerland, 2007.
- International Commission on Non-Ionizing Radiation Protection. Guidelines for limiting exposure to time-varying, electric, magnetic and electromagnetic fields up to 300 GHz. Health Phys. 1998, 74, 494–521. [Google Scholar]
- Wiltschko, W.; Wiltschko, R. Magnetic compass of european robins. Science 1972, 176, 62–64. [Google Scholar] [CrossRef]
- Röösli, M. (Ed.) Epidemiology of Electromagnetic Fields; CRC Press: Boca Raton, FL, USA, 2014. [Google Scholar]
- Kudryashov, Y.B.; Rubin, A.B. Radiation Biophysics: Ultra-Low Frequency Electromagnetic Radiation; Fizmatlit: Moscow, Russia, 2014. [Google Scholar]
- Markov, M.S. (Ed.) Electromagnetic Fields in Biology and Medicine; CRC Press: Boca Raton, FL, USA, 2015. [Google Scholar]
- Greenebaum, B.; Barnes, F. (Eds.) Biological and Medical Aspects of Electromagnetic Fields, 4th ed.; CRC Press: Boca Raton, FL, USA, 2019. [Google Scholar]
- Binhi, V.N.; Prato, F.S. Biological effects of the hypomagnetic field: An analytical review of experiments and theories. PLoS ONE 2017, 12, e0179340. [Google Scholar] [CrossRef]
- Binhi, V.N. Nonspecific magnetic biological effects: A model assuming the spin-orbit coupling. J. Chem. Phys. 2019, 151, 204101. [Google Scholar] [CrossRef] [PubMed]
- Hore, P.J.; Mouritsen, H. The radical-pair mechanism of magnetoreception. Annu. Rev. Biophys. 2016, 45, 299–344. [Google Scholar] [CrossRef]
- Wan, G.; Hayden, A.N.; Iiams, S.E.; Merlin, C. Cryptochrome 1 mediates light-dependent inclination magnetosensing in monarch butterflies. Nat. Commun. 2021, 12, 771. [Google Scholar] [CrossRef] [PubMed]
- Kishkinev, D.; Chernetsov, N.; Heyers, D.; Mouritsen, H. Migratory reed warblers need intact trigeminal nerves to correct for a 1000 km eastward displacement. PLoS ONE 2013, 8, e65847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhiman, S.K.; Galland, P. Effects of weak static magnetic fields on the gene expression of seedlings of Arabidopsis thaliana. J. Plant Physiol. 2018, 231, 9–18. [Google Scholar] [CrossRef]
- Kholodov, Y.A. Bypassing the Sensory Organs? New in Life, Science, and Technology—Biology, No. 11; Znanie: Moscow, Russia, 1991. [Google Scholar]
- Novitsky, Y.I.; Novitskaya, G.V. The Effect of a Constant Magnetic Field on Plants; Nauka: Moscow, Russia, 2016. (In Russian) [Google Scholar]
- Breus, T.K.; Binhi, V.N.; Petrukovich, A.A. Magnetic factor of the solar terrestrial relations and its impact on the human body: Physical problems and prospects for research. Physics–Uspekhi 2016, 59, 502–510. [Google Scholar] [CrossRef]
- Fu, J.P.; Mo, W.C.; Liu, Y.; He, R.Q. Decline of cell viability and mitochondrial activity in mouse skeletal muscle cell in a hypomagnetic field. Bioelectromagnetics 2016, 37, 212–222. [Google Scholar] [CrossRef]
- Paponov, I.A.; Fliegmann, J.; Narayana, R.; Maffei, M.E. Differential root and shoot magnetoresponses in Arabidopsis thaliana. Sci. Rep. 2021, 11, 9195. [Google Scholar] [CrossRef]
- Binhi, V.N. Magnetobiology: Underlying Physical Problems; Academic Press: San Diego, CA, USA, 2002. [Google Scholar]
- Johnsen, S.; Lohmann, K.J. The physics and neurobiology of magnetoreception. Nat. Rev. Neurosci. 2005, 6, 703–712. [Google Scholar] [CrossRef]
- Blank, M.; Soo, L.; Lin, H.; Henderson, A.S.; Goodman, R. Changes in transcription in HL-60 cells following exposure to alternating currents from electric fields. Bioelectrochem. Bioenerg. 1992, 28, 301–309. [Google Scholar] [CrossRef]
- Binhi, V.N. Principles of Electromagnetic Biophysics; Fizmatlit: Moscow, Russia, 2011. (In Russian) [Google Scholar]
- Binhi, V.N.; Chernavsky, D.S. Stochastic resonance of magnetosomes fixed in the cytoskeleton. Biophysics 2005, 50, 599–603. [Google Scholar]
- Kirschvink, J.L.; Winklhofer, M.; Walker, M.M. Biophysics of magnetic orientation: Strengthening the interface between theory and experimental design. J. R. Soc. Interface 2010, 7, S171–S179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afanasyeva, M.S.; Taraban, M.B.; Purtov, P.A.; Leshina, T.V.; Grissom, C.B. Magnetic spin effects in enzymatic reactions: Radical oxidation of NADH by horseradish peroxidase. J. Am. Chem. Soc. 2006, 128, 8651–8658. [Google Scholar] [CrossRef]
- Binhi, V.N.; Prato, F.S. A physical mechanism of magnetoreception: Extension and analysis. Bioelectromagnetics 2017, 38, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Li, Z.; Polyakova, T.; Dejneka, A.; Zablotskii, V.; Zhang, X. Effect of static magnetic field on DNA synthesis: The interplay between DNA chirality and magnetic field left–right asymmetry. FASEB BioAdv. 2020, 2, 254–263. [Google Scholar] [CrossRef]
- Bingi, V.N. Mechanism of magnetosensitive binding of ions by certain proteins. Biofizika 1997, 42, 338–342. [Google Scholar]
- Lambert, C.J. Basic concepts of quantum interference and electron transport in single-molecule electronics. Chem. Soc. Rev. 2015, 44, 875–888. [Google Scholar] [CrossRef]
- Binhi, V.N. Nuclear spins in the primary mechanisms of biological action of magnetic fields. Biophysics 1995, 40, 671–685. [Google Scholar]
- Buchachenko, A.L. Magnetic field-dependent molecular and chemical processes in biochemistry, genetics and medicine. Russ. Chem. Rev. 2014, 83, 1–12. [Google Scholar] [CrossRef]
- Barnes, F.S.; Greenebaum, B. The effects of weak magnetic fields on radical pairs. Bioelectromagnetics 2015, 36, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Binhi, V.N.; Savin, A.V. Molecular gyroscopes and biological effects of weak extremely low-frequency magnetic fields. Phys. Rev. E 2002, 65, 051912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tikhonov, V.I.; Volkov, A.A. Separation of water into its ortho and para isomers. Science 2002, 296, 2363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryzhkina, I.S.; Kiseleva, Y.V.; Murtazina, L.I.; Konovalov, A.I. Effect of ultralow concentrations and electromagnetic fields. Dokl. Phys. Chem. 2012, 446, 153–157. [Google Scholar] [CrossRef]
- Shatalov, V.M.; Filippov, A.E.; Noga, I.V. Bubbles induced fluctuations of some properties of aqueous solutions. Biophysics 2012, 57, 421–427. [Google Scholar] [CrossRef]
- Pershin, S.M. Effect of quantum differences of ortho and para H2O spin-isomers on water properties: Biophysical aspect. Biophysics 2013, 58, 723–730. [Google Scholar] [CrossRef]
- Bunkin, N.F.; Shkirin, A.V.; Suyazov, N.V.; Babenko, V.A.; Sychev, A.A.; Penkov, N.V.; Belosludtsev, K.N.; Gudkov, S.V. Formation and dynamics of ion-stabilized gas nanobubble phase in the bulk of aqueous NaCl solutions. J. Phys. Chem. B 2016, 120, 1291–1303. [Google Scholar] [CrossRef]
- Novikov, V.V.; Yablokova, E.V.; Fesenko, E.E. The role of water in the effect of weak combined magnetic fields on production of reactive oxygen species (ROS) by neutrophils. Appl. Sci. 2020, 10, 3326. [Google Scholar] [CrossRef]
- Sarimov, R.M.; Binhi, V.N. Low-frequency magnetic fields in cars and office premises and the geomagnetic field variations. Bioelectromagnetics 2020, 41, 360–368. [Google Scholar] [CrossRef]
- Portelli, L.A.; Schomay, T.E.; Barnes, F.S. Inhomogeneous background magnetic field in biological incubators is a potential confounder for experimental variability and reproducibility. Bioelectromagnetics 2013, 34, 337–348. [Google Scholar] [CrossRef]
- Makinistian, L.; Muehsam, D.J.; Bersani, F.; Belyaev, I. Some recommendations for experimental work in magnetobiology, revisited. Bioelectromagnetics 2018, 39, 556–564. [Google Scholar] [CrossRef] [PubMed]
- Prato, F.S.; Desjardins-Holmes, D.; Keenliside, L.D.; DeMoor, J.M.; Robertson, J.A.; Thomas, A.W. Magnetoreception in laboratory mice: Sensitivity to extremely low frequency fields exceeds 33 nT at 30 Hz. J. R. Soc. Interface 2013, 10, 20121046. [Google Scholar] [CrossRef] [Green Version]
- Engels, S.; Schneider, N.L.; Lefeldt, N.; Hein, C.M.; Zapka, M.; Michalik, A.; Elbers, D.; Kittel, A.; Hore, P.J.; Mouritsen, H. Anthropogenic electromagnetic noise disrupts magnetic compass orientation in a migratory bird. Nature 2014, 509, 353–356. [Google Scholar] [CrossRef]
- Valberg, P.A. Designing EMF experiments: What is required to characterize “exposure”? Bioelectromagnetics 1995, 16, 396–401. [Google Scholar] [CrossRef]
- Buchachenko, A. Why magnetic and electromagnetic effects in biology are irreproducible and contradictory? Bioelectromagnetics 2016, 37, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Portelli, L. Overcoming the irreproducibility barrier: Considerations to improve the quality of experimental practice when investigating the effects of low-level electric and magnetic fields on in vitro biological systems. In Bioengineering and Biophysical Aspects of Electromagnetic Fields, 4th ed.; Greenebaum, B., Barnes, F., Eds.; CRC Press: Boca Raton, FL, USA, 2019; pp. 435–462. [Google Scholar]
- Zenchenko, T.A.; Breus, T.K. The possible effect of space weather factors on various physiological systems of the human organism. Atmosphere 2021, 12, 346. [Google Scholar] [CrossRef]
- Krylov, V.V. Biological effects related to geomagnetic activity and possible mechanisms. Bioelectromagnetics 2017, 38, 497–510. [Google Scholar] [CrossRef]
- Tsurutani, B.T.; Gonzalez, W.D.; Kamide, Y.; Arballo, J.K. (Eds.) Magnetic Storms; American Geophysical Union: Washington, DC, USA, 1997. [Google Scholar]
- Pishchalnikov, R.Y.; Gurfinkel, Y.I.; Sarimov, R.M.; Vasin, A.L.; Sasonko, M.L.; Matveeva, T.A.; Binhi, V.N.; Baranov, M.V. Cardiovascular response as a marker of environmental stress caused by variations in geomagnetic field and local weather. Biomed. Signal Process. Control 2019, 51, 401–410. [Google Scholar] [CrossRef]
- Koonin, E.V. The Logic of Chance: The Nature and Origin of Biological Evolution; FT Press: Upper Saddle River, NJ, USA, 2012. [Google Scholar]
- Binhi, V.N.; Prato, F.S. Rotations of macromolecules affect nonspecific biological responses to magnetic fields. Sci. Rep. 2018, 8, 13495. [Google Scholar] [CrossRef] [PubMed]
- Binhi, V.N. Random effects in magnetobiology and a way to summarize them. Bioelectromagnetics 2021, 42, 501–515. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Jarocha, L.E.; Zollitsch, T.; Konowalczyk, M.; Henbest, K.B.; Richert, S.; Golesworthy, M.J.; Schmidt, J.; Dejean, V.; Sowood, D.J.C.; et al. Magnetic sensitivity of cryptochrome 4 from a migratory songbird. Nature 2021, 594, 535–540. [Google Scholar] [CrossRef] [PubMed]
- Hoff, A.J.; Rademaker, H.; Van Grondelle, R.; Duysens, L.N.M. On the magnetic field dependence of the yield of the triplet state in reaction centers of photosynthetic bacteria. Biochim. Biophys. Acta-Bioenerg. 1977, 460, 547–554. [Google Scholar] [CrossRef]
- Schulten, K.; Swenberg, C.; Weller, A. A biomagnetic sensory mechanism based on magnetic field modulated coherent electron spin motion. Z. Phys. Chem. 1978, 111, 1–5. [Google Scholar] [CrossRef]
- Richards, O.W.; Davies, R.G. Imms’ General Textbook of Entomology: Structure, Physiology and Development, 10th ed.; Chapman and Hall: London, UK, 1977. [Google Scholar]
- Simons, A.M. Many wrongs: The advantage of group navigation. Trends Ecol. Evol. 2004, 19, 453–455. [Google Scholar] [CrossRef] [PubMed]
- Binhi, V.N. A limit in the dynamic increase in the accuracy of group migration. BioSystems 2018, 166, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Kattnig, D.R.; Solov’yov, I.; Hore, P.J. Electron spin relaxation in cryptochrome-based magnetoreception. Phys. Chem. Chem. Phys. 2016, 18, 12443–12456. [Google Scholar] [CrossRef] [Green Version]
- Gauger, E.M.; Rieper, E.; Morton, J.J.L.; Benjamin, S.C.; Vedral, V. Sustained quantum coherence and entanglement in the avian compass. Phys. Rev. Lett. 2011, 106, 040503. [Google Scholar] [CrossRef] [Green Version]
- Yan, M.M.; Zhang, L.; Cheng, Y.X.; Sappington, T.W.; Pan, W.D.; Jiang, X.F. Effect of a near-zero magnetic field on development and flight of oriental armyworm (Mythimna separata). J. Integr. Agric. 2020, 19, 2–11. [Google Scholar] [CrossRef]
- Wan, G.J.; Jiang, S.L.; Zhang, M.; Zhao, J.Y.; Zhang, Y.C.; Pan, W.D.; Sword, G.A.; Chen, F.J. Geomagnetic field absence reduces adult body weight of amigratory insect by disrupting feeding behavior and appetiteregulation. Insect Sci. 2021, 28, 251–260. [Google Scholar] [CrossRef]
- Xue, X.; Ali, Y.F.; Luo, W.; Liu, C.; Zhou, G.; Liu, N.A. Biological effects of space hypomagnetic environment on circadian rhythm. Front. Physiol. 2021, 12, 643943. [Google Scholar] [CrossRef]
- Zhang, B.; Wang, L.; Zhan, A.; Wang, M.; Tian, L.; Guo, W.; Pan, Y. Long-term exposure to a hypomagnetic field attenuates adult hippocampal neurogenesis and cognition. Nat. Commun. 2021, 12, 1174. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Feng, S.; Yu, Y.; Zhang, Y.; Wei, S. Near-null magnetic field suppresses fruit growth in Arabidopsis. Bioelectromagnetics 2021, 42, 593–602. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Xue, Y.; Yang, J.; Shang, P.; Yuan, X. Biological effects of hypomagnetic field: Ground-based data for space exploration. Bioelectromagnetics 2021, 42, 516–531. [Google Scholar] [CrossRef]
- Binhi, V.N. Primary physical mechanism of the biological effects of weak magnetic fields. Biophysics 2016, 61, 170–176. [Google Scholar] [CrossRef]
- Syfers, R. EMF and childhood leukemia. EPRI J. 2006, Spring, 24–31. [Google Scholar]
- Karimi, A.; Moghaddam, F.G.; Valipour, M. Insights in the biology of extremely low-frequency magnetic fields exposure on human health. Mol. Biol. Rep. 2020, 47, 5621–5633. [Google Scholar] [CrossRef]
- Shine, M.B.; Guruprasad, K.N.; Anand, A. Enhancement of germination, growth, and photosynthesis in soybean by pre-treatment of seeds with magnetic field. Bioelectromagnetics 2011, 32, 474–484. [Google Scholar] [CrossRef] [PubMed]
- Purygin, P.P.; Vassilieva, T.I.; Purygin, V.A.; Sovetkin, D.A.; Tsaplev, D.A. Influence of Pre-Sowing Treatment of Flax Seeds on the Growth and Biochemical Parameters of Seedlings; Natural Science Series; Samara State University Bulletin: Samara, Russia, 2015; pp. 166–173. (In Russian) [Google Scholar]
- Michalak, I.; Lewandowska, S.; Niemczyk, K.; Detyna, J.; Bujak, H.; Arik, P.; Bartniczak, A. Germination of soybean seeds exposed to the static/alternating magnetic field and algal extract. Eng. Life Sci. 2019, 19, 986–999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammad, M.; Albaqami, M.; Pooam, M.; Kernevez, E.; Witczak, J.; Ritz, T.; Martino, C.; Ahmad, M. Cryptochrome mediated magnetic sensitivity in Arabidopsis occurs independently of light-induced electron transfer to the flavin. Photochem. Photobiol. Sci. 2020, 19, 341–352. [Google Scholar] [CrossRef] [PubMed]
- Finegold, L.; Flamm, B.L. Magnet therapy: Extraordinary claims, but no proved benefits. Br. Med. J. 2006, 332, 4. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Yu, Y.; Zhang, Y.; Li, Y.; Wei, S. Gibberellins are involved in effect of near-null magnetic field on Arabidopsis flowering. Bioelectromagnetics 2017, 38, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Agliassa, C.; Maffei, M.E. Reduction of geomagnetic field (GMF) to near null magnetic field (NNMF) affects some Arabidopsis thaliana clock genes amplitude in a light independent manner. J. Plant Physiol. 2019, 232, 23–26. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Binhi, V.N.; Rubin, A.B. Theoretical Concepts in Magnetobiology after 40 Years of Research. Cells 2022, 11, 274. https://doi.org/10.3390/cells11020274
Binhi VN, Rubin AB. Theoretical Concepts in Magnetobiology after 40 Years of Research. Cells. 2022; 11(2):274. https://doi.org/10.3390/cells11020274
Chicago/Turabian StyleBinhi, Vladimir N., and Andrei B. Rubin. 2022. "Theoretical Concepts in Magnetobiology after 40 Years of Research" Cells 11, no. 2: 274. https://doi.org/10.3390/cells11020274