
Follistatin Is a Novel Chemoattractant for Migration and Invasion of Placental Trophoblasts of Mice


Abstract

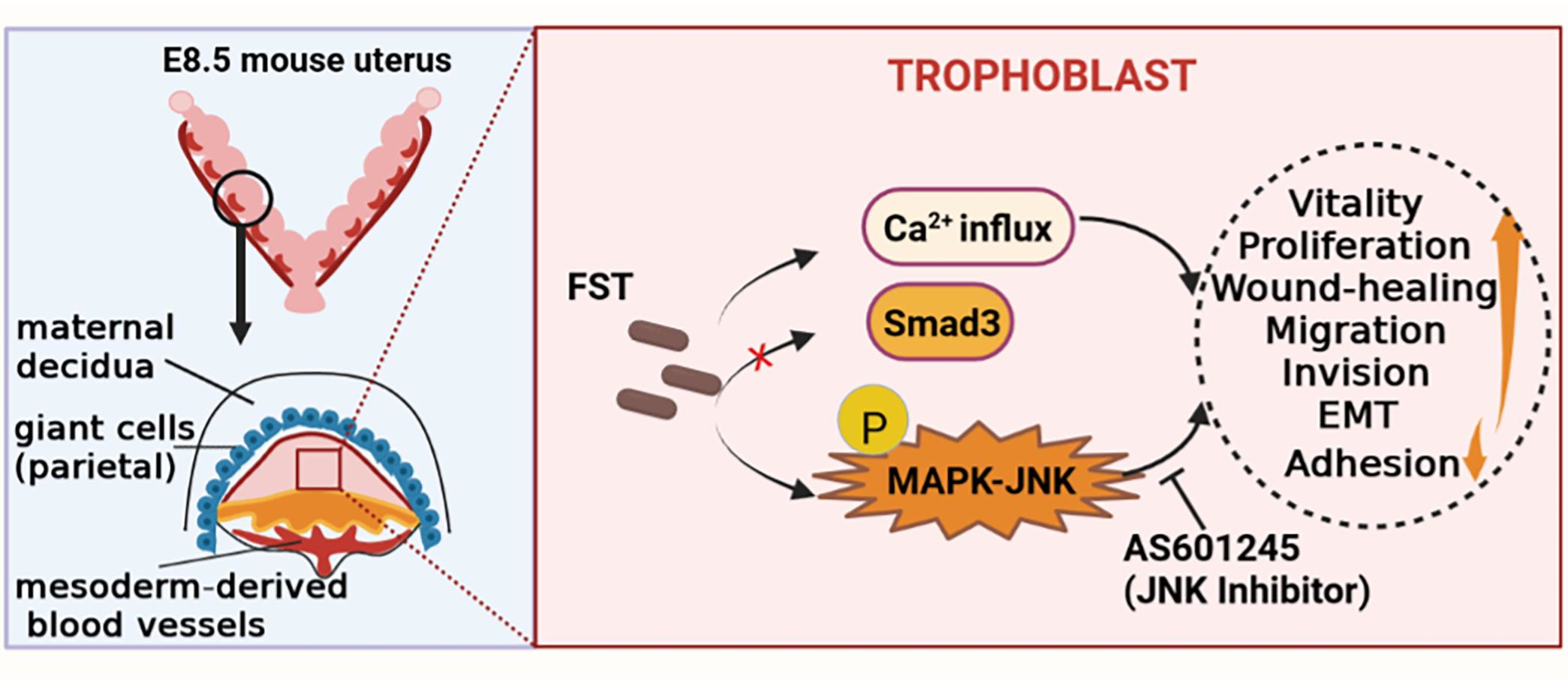{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Cell Culture
2.3. Pretreatment of Primary Cultured Trophoblasts of Mouse with AS601245
2.4. CCK-8 Assay
2.5. Real-Time Cell Analysis
2.6. Giemsa Staining
2.7. Wound Healing Assay
2.8. Microfluidic Cell Migration Assay
2.8.1. Microfluidic Chip Preparation
2.8.2. Cell Loading
2.8.3. Gradient Generation
2.8.4. Image Capture
2.8.5. Image Analysis
2.9. Matrigel-Coated Transwell Invasion Assay
2.10. Immunofluorescence Assay
2.11. Calcium Flux Assay
2.12. Western Blotting
2.13. Statistical Analysis
3. Results
3.1. FST Increased Viability and Proliferation of Primary Cultured Trophoblasts of Mouse
3.2. FST Promoted Wound Healing of Primary Cultured Trophoblasts of Mouse
3.3. FST Induced Migration and Invasion of Primary Cultured Trophoblasts of Mice
3.4. FST Suppressed Adhesion of Primary Cultured Trophoblasts of Mouse
3.5. FST Increased Expression of Migration and Polarity-Related Protein of Trophoblasts
3.6. FST Increased Intracellular Calcium Flux and p-JNK Protein Level of Primary Cultured Trophoblasts of Mice
3.7. JNK Inhibitor AS601245 Suppressed FST-induced Migration and Invasion of Primary Cultured Trophoblasts of Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Maltepe, E.; Fisher, S.J. Placenta: The forgotten organ. Annu. Rev. Cell Dev. Biol. 2015, 31, 523–552. [Google Scholar] [CrossRef]
- Gerri, C.; McCarthy, A.; Alanis-Lobato, G.; Demtschenko, A.; Bruneau, A.; Loubersac, S.; Fogarty, N.M.E.; Hampshire, D.; Elder, K.; Snell, P.; et al. Initiation of a conserved trophectoderm program in human, cow and mouse embryos. Nature 2020, 587, 443–447. [Google Scholar] [CrossRef]
- Mohan, A.; Asselin, J.; Sargent, I.L.; Groome, N.P.; Muttukrishna, S. Effect of cytokines and growth factors on the secretion of inhibin A, activin A and follistatin by term placental villous trophoblasts in culture. Eur. J. Endocrinol. 2001, 145, 505–511. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, Y.; Nakamura, T.; Nakamura, R.; Irahara, M.; Aono, T.; Sugino, H. Identification of activins and follistatin proteins in human follicular fluid and placenta. J. Clin. Endocrinol. Metab. 1995, 80, 915–921. [Google Scholar] [CrossRef] [PubMed]
- Hardy, J.T.; Buhimschi, I.A.; McCarthy, M.E.; Zhao, G.; Laky, C.A.; Shook, L.L.; Buhimschi, C.S. Imbalance of Amniotic Fluid Activin-A and Follistatin in Intraamniotic Infection, Inflammation, and Preterm Birth. J. Clin. Endocrinol. Metab. 2016, 101, 2785–2793. [Google Scholar] [CrossRef]
- Bloise, E.; Ciarmela, P.; Dela Cruz, C.; Luisi, S.; Petraglia, F.; Reis, F.M. Activin A in Mammalian Physiology. Physiol. Rev. 2019, 99, 739–780. [Google Scholar] [CrossRef] [PubMed]
- Muttukrishna, S.; Tannetta, D.; Groome, N.; Sargent, I. Activin and follistatin in female reproduction. Mol. Cell Endocrinol. 2004, 225, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Prakash, A.; Laird, S.; Tuckerman, E.; Li, T.C.; Ledger, W.L. Inhibin A and activin A may be used to predict pregnancy outcome in women with recurrent miscarriage. Fertil. Steril. 2005, 83, 1758–1763. [Google Scholar] [CrossRef] [PubMed]
- Prakash, A.; Li, T.C.; Tuckerman, E.; Laird, S.; Wells, M.; Ledger, W.L. A study of luteal phase expression of inhibin, activin, and follistatin subunits in the endometrium of women with recurrent miscarriage. Fertil. Steril. 2006, 86, 1723–1730. [Google Scholar] [CrossRef] [PubMed]
- D’Antona, D.; Reis, F.M.; Benedetto, C.; Evans, L.W.; Groome, N.P.; de Kretser, D.M.; Wallace, E.M.; Petraglia, F. Increased maternal serum activin A but not follistatin levels in pregnant women with hypertensive disorders. J. Endocrinol. 2000, 165, 157–162. [Google Scholar] [CrossRef]
- Prakash, A.; Tuckerman, E.; Laird, S.; Ola, B.; Li, T.C.; Ledger, W.L. A preliminary study comparing the endometrial expression of inhibin, activin and follistatin in women with a history of implantation failure after IVF treatment and a control group. BJOG 2008, 115, 532–537. [Google Scholar] [CrossRef] [PubMed]
- Fullerton, P.T.J.; Monsivais, D.; Kommagani, R.; Matzuk, M.M. Follistatin is critical for mouse uterine receptivity and decidualization. Proc. Natl. Acad. Sci. USA 2017, 114, E4772–E4781. [Google Scholar] [CrossRef]
- Matzuk, M.M.; Lu, N.; Vogel, H.; Sellheyer, K.; Roop, D.R.; Bradley, A. Multiple defects and perinatal death in mice deficient in follistatin. Nature 1995, 374, 360–363. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Kumar, T.R.; Woodruff, T.; Hadsell, L.A.; DeMayo, F.J.; Matzuk, M.M. Overexpression of mouse follistatin causes reproductive defects in transgenic mice. Mol. Endocrinol. 1998, 12, 96–106. [Google Scholar] [CrossRef] [PubMed]
- Lamouille, S.; Xu, J.; Derynck, R. Molecular mechanisms of epithelial-mesenchymal transition. Nat. Rev. Mol. Cell Biol. 2014, 15, 178–196. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Klausen, C.; Cheng, J.C.; Zhu, H.; Leung, P.C. Activin A, B, and AB increase human trophoblast cell invasion by up-regulating N-cadherin. J. Clin. Endocrinol. Metab. 2014, 99, E2216–E2225. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Moustakas, A.; Heldin, P. TGFβ and matrix-regulated epithelial to mesenchymal transition. Biochim. Biophys. Acta 2014, 1840, 2621–2634. [Google Scholar] [CrossRef]
- Liu, H.; Wang, L.; Wang, Y.; Zhu, Q.; Aldo, P.; Ding, J.; Mor, G.; Liao, A. Establishment and characterization of a new human first trimester Trophoblast cell line, AL07. Placenta 2020, 100, 122–132. [Google Scholar] [CrossRef]
- Kojima, J.; Fukuda, A.; Taira, H.; Kawasaki, T.; Ito, H.; Kuji, N.; Isaka, K.; Umezawa, A.; Akutsu, H. Efficient production of trophoblast lineage cells from human induced pluripotent stem cells. Lab. Investig. 2017, 97, 1188–1200. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Xu, L.; Jin, H.; Zhong, Y.; Di, J.; Lin, Q.D. CXCL12 enhances exogenous CD4+CD25+ T cell migration and prevents embryo loss in non-obese diabetic mice. Fertil. Steril. 2009, 91, 2687–2696. [Google Scholar] [CrossRef] [PubMed]
- Hamidi, H.; Lilja, J.; Ivaska, J. Using xCELLigence RTCA Instrument to Measure Cell Adhesion. Bio Protoc. 2017, 7, e2646. [Google Scholar] [CrossRef]
- Yang, K.; Yang, X.; Gao, C.; Hua, C.; Hong, C.; Zhu, L. A Novel Microfluidic Device for the Neutrophil Functional Phenotype Analysis: Effects of Glucose and Its Derivatives AGEs. Micromachines 2021, 12, 944. [Google Scholar] [CrossRef] [PubMed]
- Schulz, L.C.; Widmaier, E.P. The effect of leptin on mouse trophoblast cell invasion. Biol. Reprod. 2004, 71, 1963–1967. [Google Scholar] [CrossRef] [PubMed]
- Hemberger, M.; Hughes, M.; Cross, J.C. Trophoblast stem cells differentiate in vitro into invasive trophoblast giant cells. Dev. Biol. 2004, 271, 362–371. [Google Scholar] [CrossRef] [PubMed]
- Perez-Garcia, V.; Lea, G.; Lopez-Jimenez, P.; Okkenhaug, H.; Burton, G.J.; Moffett, A.; Turco, M.Y.; Hemberger, M. BAP1/ASXL complex modulation regulates epithelial-mesenchymal transition during trophoblast differentiation and invasion. Elife 2021, 10, e63254. [Google Scholar] [CrossRef]
- Gupta, S.; Maitra, A. EMT: Matter of Life or Death? Cell 2016, 164, 840–842. [Google Scholar] [CrossRef]
- Yu, M.; Bardia, A.; Wittner, B.S.; Stott, S.L.; Smas, M.E.; Ting, D.T.; Isakoff, S.J.; Ciciliano, J.C.; Wells, M.N.; Shah, A.M.; et al. Circulating breast tumor cells exhibit dynamic changes in epithelial and mesenchymal composition. Science 2013, 339, 580–584. [Google Scholar] [CrossRef]
- Eastham, A.M.; Spencer, H.; Soncin, F.; Ritson, S.; Merry, C.L.; Stern, P.L.; Ward, C.M. Epithelial-mesenchymal transition events during human embryonic stem cell differentiation. Cancer Res. 2007, 67, 11254–11262. [Google Scholar] [CrossRef]
- Chen, T.; You, Y.; Jiang, H.; Wang, Z.Z. Epithelial-mesenchymal transition (EMT): A biological process in the development, stem cell differentiation, and tumorigenesis. J. Cell Physiol. 2017, 232, 3261–3272. [Google Scholar] [CrossRef]
- Tsukita, S.; Yonemura, S. Cortical actin organization: Lessons from ERM (ezrin/radixin/moesin) proteins. J. Biol. Chem. 1999, 274, 34507–34510. [Google Scholar] [CrossRef]
- Song, X.; Wang, W.; Wang, H.; Yuan, X.; Yang, F.; Zhao, L.; Mullen, M.; Du, S.; Zohbi, N.; Muthusamy, S.; et al. Acetylation of ezrin regulates membrane-cytoskeleton interaction underlying CCL18-elicited cell migration. J. Mol. Cell Biol. 2020, 12, 424–437. [Google Scholar] [CrossRef] [PubMed]
- Xie, D.; Liu, Z.; Wu, J.; Feng, W.; Yang, K.; Deng, J.; Tian, G.; Santos, S.; Cui, X.; Lin, F. The effects of activin A on the migration of human breast cancer cells and neutrophils and their migratory interaction. Exp. Cell Res. 2017, 357, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Ramathal, C.Y.; Bagchi, I.C.; Taylor, R.N.; Bagchi, M.K. Endometrial decidualization: Of mice and men. Semin. Reprod. Med. 2010, 28, 17–26. [Google Scholar] [CrossRef]
- Perez-Garcia, V.; Fineberg, E.; Wilson, R.; Murray, A.; Mazzeo, C.I.; Tudor, C.; Sienerth, A.; White, J.K.; Tuck, E.; Ryder, E.J.; et al. Placentation defects are highly prevalent in embryonic lethal mouse mutants. Nature 2018, 555, 463–468. [Google Scholar] [CrossRef] [PubMed]
- O’Connell, A.R.; McNatty, K.P.; Hurst, P.R.; Spencer, T.E.; Bazers, F.W.; Reader, K.L.; Johnstone, P.D.; Davis, G.H.; Juengel, J.L. Activin A and follistatin during the oestrous cycle and early pregnancy in ewes. J. Endocrinol. 2016, 228, 193–203. [Google Scholar] [CrossRef][Green Version]
- Bearfield, C.; Jauniaux, E.; Groome, N.; Sargent, I.L.; Muttukrishna, S. The secretion and effect of inhibin A, activin A and follistatin on first-trimester trophoblasts in vitro. Eur. J. Endocrinol. 2005, 152, 909–916. [Google Scholar] [CrossRef]
- Garcés, M.F.; Vallejo, S.A.; Sanchez, E.; Palomino-Palomino, M.A.; Leal, L.G.; Ángel-Muller, E.; Díaz-Cruz, L.A.; Ruíz-Parra, A.I.; González-Clavijo, A.M.; Castaño, J.P.; et al. Longitudinal analysis of maternal serum Follistatin concentration in normal pregnancy and preeclampsia. Clin. Endocrinol. 2015, 83, 229–235. [Google Scholar] [CrossRef]
- Kimura, F.; Sidis, Y.; Bonomi, L.; Xia, Y.; Schneyer, A. The follistatin-288 isoform alone is sufficient for survival but not for normal fertility in mice. Endocrinology 2010, 151, 1310–1319. [Google Scholar] [CrossRef][Green Version]
- Fahmy-Garcia, S.; Farrell, E.; Witte-Bouma, J.; Robbesom-van den Berge, I.; Suarez, M.; Mumcuoglu, D.; Walles, H.; Kluijtmans, S.G.J.M.; van de Eerden, B.C.J.; van Osch, G.J.V.M.; et al. Follistatin Effects in Migration, Vascularization, and Osteogenesis in vitro and Bone Repair in vivo. Front. Bioeng. Biotechnol. 2019, 7, 38. [Google Scholar] [CrossRef]
- Arai, K.Y.; Nishiyama, T. Developmental changes in extracellular matrix messenger RNAs in the mouse placenta during the second half of pregnancy: Possible factors involved in the regulation of placental extracellular matrix expression. Biol. Reprod. 2007, 77, 923–933. [Google Scholar] [CrossRef]
- Natale, D.R.; Hemberger, M.; Hughes, M.; Cross, J.C. Activin promotes differentiation of cultured mouse trophoblast stem cells towards a labyrinth cell fate. Dev. Biol. 2009, 335, 120–131. [Google Scholar] [CrossRef]
- Thiery, J.P.; Acloque, H.; Huang, R.Y.; Nieto, M.A. Epithelial-mesenchymal transitions in development and disease. Cell 2009, 139, 871–890. [Google Scholar] [CrossRef] [PubMed]
- Kalluri, R.; Weinberg, R.A. The basics of epithelial-mesenchymal transition. J. Clin. Investig. 2009, 119, 1420–1428. [Google Scholar] [CrossRef]
- De Robertis, M.; Greco, M.R.; Cardone, R.A.; Mazza, T.; Marzano, F.; Mehterov, N.; Kazakova, M.; Belev, N.; Tullo, A.; Pesole, G.; et al. Upregulation of YKL-40 Promotes Metastatic Phenotype and Correlates with Poor Prognosis and Therapy Response in Patients with Colorectal Cancer. Cells 2022, 11, 3568. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, M.; Christofori, G. EMT, the cytoskeleton, and cancer cell invasion. Cancer Metastasis Rev. 2009, 28, 15–33. [Google Scholar] [CrossRef] [PubMed]
- Theveneau, E.; Mayor, R. Cadherins in collective cell migration of mesenchymal cells. Curr. Opin. Cell Biol. 2012, 24, 677–684. [Google Scholar] [CrossRef]
- Wang, R.; Wang, X.; Yin, L.; Yin, L.; Chu, G.C.-Y.; Hu, P.; Ou, Y.; Zhang, Y.; Lewis, M.S.; Pandol, S.J. Breast Cancer MCF-7 Cells Acquire Heterogeneity during Successive Co-Culture with Hematopoietic and Bone Marrow-Derived Mesenchymal Stem/Stromal Cells. Cells 2022, 11, 3553. [Google Scholar] [CrossRef]
- Nisticò, P.; Bissell, M.J.; Radisky, D.C. Epithelial-mesenchymal transition: General principles and pathological relevance with special emphasis on the role of matrix metalloproteinases. Cold Spring Harb. Perspect. Biol. 2012, 4, a011908. [Google Scholar] [CrossRef]
- Siddiqui, T.A.; Lively, S.; Vincent, C.; Schlichter, L.C. Regulation of podosome formation, microglial migration and invasion by Ca(2+)-signaling molecules expressed in podosomes. J. Neuroinflamm. 2012, 9, 250. [Google Scholar] [CrossRef]
- Li, Y.; Klausen, C.; Zhu, H.; Leung, P.C. Activin A Increases Human Trophoblast Invasion by Inducing SNAIL-Mediated MMP2 Up-Regulation Through ALK4. J. Clin. Endocrinol. Metab. 2015, 100, E1415–E1427. [Google Scholar] [CrossRef]
- de Guise, C.; Lacerte, A.; Rafiei, S.; Reynaud, R.; Roy, M.; Brue, T.; Lebrun, J.J. Activin inhibits the human Pit-1 gene promoter through the p38 kinase pathway in a Smad-independent manner. Endocrinology 2006, 147, 4351–4362. [Google Scholar] [CrossRef]
- Jiang, L.; Qi, Y.; Kong, X.; Wang, R.; Qi, J.; Lin, F.; Cui, X.; Liu, Z. Activin A as a Novel Chemokine Induces Migration of L929 Fibroblasts by ERK Signaling in Microfluidic Devices. Front. Cell Dev. Biol. 2021, 9, 660316. [Google Scholar] [CrossRef] [PubMed]
- Bao, Y.L.; Tsuchida, K.; Liu, B.; Kurisaki, A.; Matsuzaki, T.; Sugino, H. Synergistic activity of activin A and basic fibroblast growth factor on tyrosine hydroxylase expression through Smad3 and ERK1/ERK2 MAPK signaling pathways. J. Endocrinol. 2005, 184, 493–504. [Google Scholar] [CrossRef] [PubMed]
- Bildik, G.; Akin, N.; Esmaeilian, Y.; Hela, F.; Yildiz, C.S.; Iltumur, E.; İncir, S.; Karahuseyinoglu, S.; Yakin, K.; Oktem, O. Terminal differentiation of human granulosa cells as luteinization is reversed by activin-A through silencing of Jnk pathway. Cell Death Discov. 2020, 6, 93. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Zheng, Y.; Zhang, H.; Liu, Y.; Sun, H.; Zhang, P. Galectin-1 induces metastasis and epithelial-mesenchymal transition (EMT) in human ovarian cancer cells via activation of the MAPK JNK/p38 signalling pathway. Am. J. Transl. Res. 2019, 11, 3862–3878. [Google Scholar]
- Xu, M.; Wang, S.; Wang, Y.; Wu, H.; Frank, J.A.; Zhang, Z.; Luo, J. Role of p38γ MAPK in regulation of EMT and cancer stem cells. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 1864, 3605–3617. [Google Scholar] [CrossRef]
- Chaly, Y.; Blair, H.C.; Smith, S.M.; Bushnell, D.S.; Marinov, A.D.; Campfield, B.T.; Hirsch, R. Follistatin-like protein 1 regulates chondrocyte proliferation and chondrogenic differentiation of mesenchymal stem cells. Ann. Rheum. Dis. 2015, 74, 1467–1473. [Google Scholar] [CrossRef]
- Ni, S.; Li, C.; Xu, N.; Liu, X.; Wang, W.; Chen, W.; Wang, Y.; van Wijnen, A.J. Follistatin-like protein 1 induction of matrix metalloproteinase 1, 3 and 13 gene expression in rheumatoid arthritis synoviocytes requires MAPK, JAK/STAT3 and NF-κB pathways. J. Cell Physiol. 2018, 234, 454–463. [Google Scholar] [CrossRef]
- Liu, X.; Li, H.; Rajurkar, M.; Li, Q.; Cotton, J.L.; Ou, J.; Zhu, L.J.; Goel, H.L.; Mercurio, A.M.; Park, J.S.; et al. Tead and AP1 Coordinate Transcription and Motility. Cell Rep. 2016, 14, 1169–1180. [Google Scholar] [CrossRef]
- Jie, Z.; Xie, Z.; Zhao, X.; Sun, X.; Yu, H.; Pan, X.; Shen, S.; Qin, A.; Fang, X.; Fan, S. Glabridin inhibits osteosarcoma migration and invasion via blocking the p38- and JNK-mediated CREB-AP1 complexes formation. J. Cell Physiol. 2019, 234, 4167–4178. [Google Scholar] [CrossRef]
- Zheng, Q.; Yang, Y.; Cui, X.; Zhang, D.; Liu, S.; Yan, Q. AP1 mediates uPA/uPAR induced FUT4 expression and trophoblast invasion. J. Cell Biochem. 2018, 119, 6442–6451. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhao, J.; Luan, X.; Li, S.; Zhai, J.; Liu, J.; Du, Y. SPARCL1 impedes trophoblast migration and invasion by down-regulating ERK phosphorylation and AP-1 production and altering EMT-related molecule expression. Placenta 2020, 89, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Mori, Y.; Higuchi, M.; Hirabayashi, Y.; Fukuda, M.; Gotoh, Y. JNK phosphorylates synaptotagmin-4 and enhances Ca2+-evoked release. EMBO J. 2008, 27, 76–87. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.N.; Xia, Y.Z.; Zhang, C.; Zhang, H.; Luo, J.G.; Yang, L.; Kong, L.Y. Vielanin K enhances doxorubicin-induced apoptosis via activation of IRE1α-TRAF2-JNK pathway and increases mitochondrial Ca2 + influx in MCF-7 and MCF-7/MDR cells. Phytomedicine 2020, 78, 153329. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Qi, Y.; Yang, K.; Zhu, L.; Cui, X.; Liu, Z. Follistatin Is a Novel Chemoattractant for Migration and Invasion of Placental Trophoblasts of Mice. Cells 2022, 11, 3816. https://doi.org/10.3390/cells11233816
Li J, Qi Y, Yang K, Zhu L, Cui X, Liu Z. Follistatin Is a Novel Chemoattractant for Migration and Invasion of Placental Trophoblasts of Mice. Cells. 2022; 11(23):3816. https://doi.org/10.3390/cells11233816
Chicago/Turabian StyleLi, Jing, Yan Qi, Ke Yang, Linjing Zhu, Xueling Cui, and Zhonghui Liu. 2022. "Follistatin Is a Novel Chemoattractant for Migration and Invasion of Placental Trophoblasts of Mice" Cells 11, no. 23: 3816. https://doi.org/10.3390/cells11233816
APA StyleLi, J., Qi, Y., Yang, K., Zhu, L., Cui, X., & Liu, Z. (2022). Follistatin Is a Novel Chemoattractant for Migration and Invasion of Placental Trophoblasts of Mice. Cells, 11(23), 3816. https://doi.org/10.3390/cells11233816