

FUT2 Facilitates Autophagy and Suppresses Apoptosis via p53 and JNK Signaling in Lung Adenocarcinoma Cells

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents, Plasmids, and Antibodies
2.2. Bioinformatics Analysis
2.3. Cell Culture and Transfection
2.4. RNA Extraction and Quantitative RT-PCR (RT-qPCR)
2.5. Western Blot Analysis
2.6. Autophagy Activity Detection
2.7. Transcriptome Analysis
2.8. Apoptosis Detection
2.9. Mitochondrial Membrane Potential (MMP) Detection
2.10. Immunofluorescence
2.11. Co-Immunoprecipitation (Co-IP)
2.12. In Vivo Experiment
2.13. Statistical Analysis
3. Results
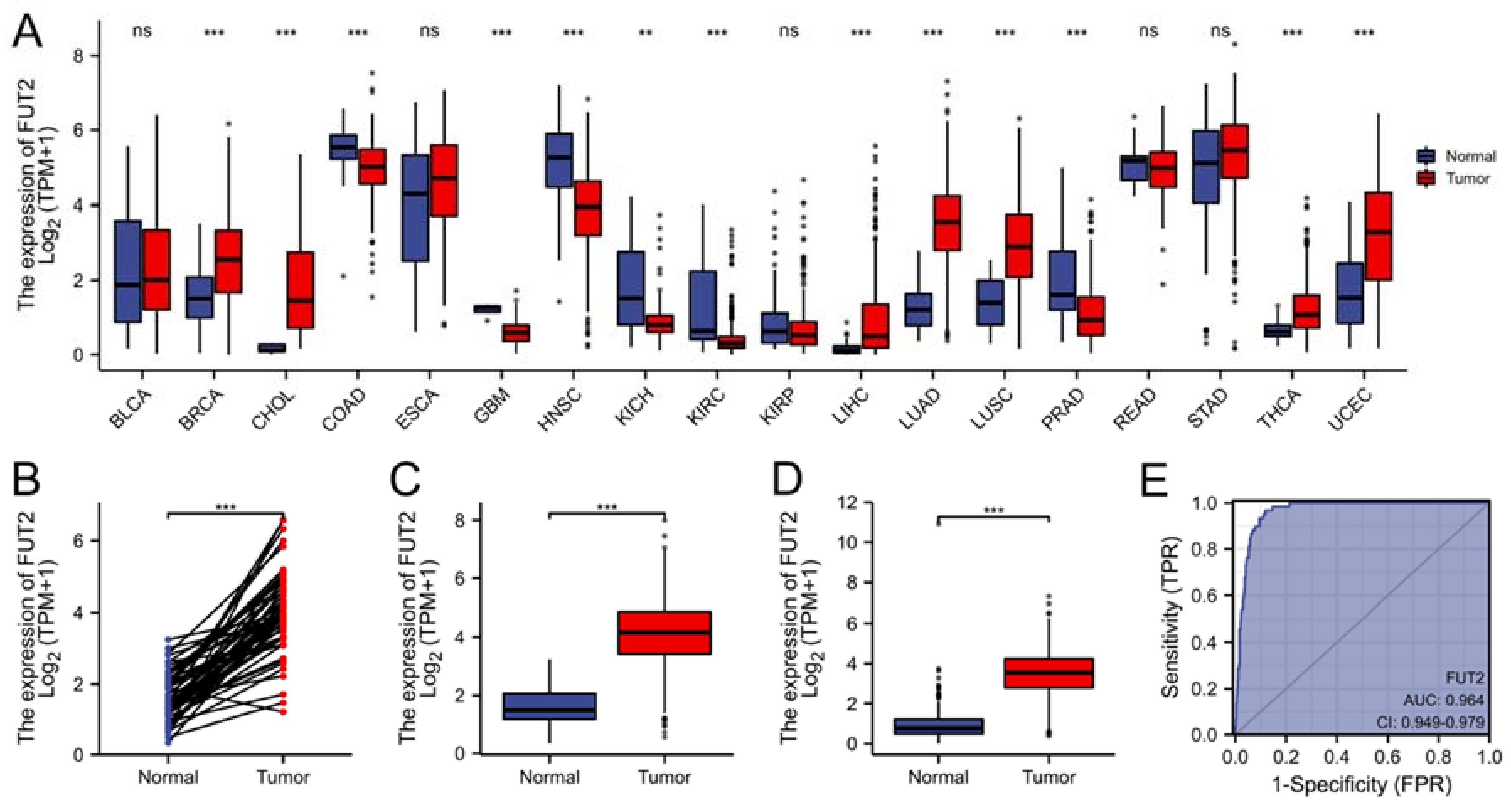
3.1. Upregulated mRNA Expression of FUT2 in Patients with Lung Adenocarcinoma
3.2. Relationships between the mRNA Levels of FUT2 and Clinical Pathological Characteristics of Lung Adenocarcinoma Patients
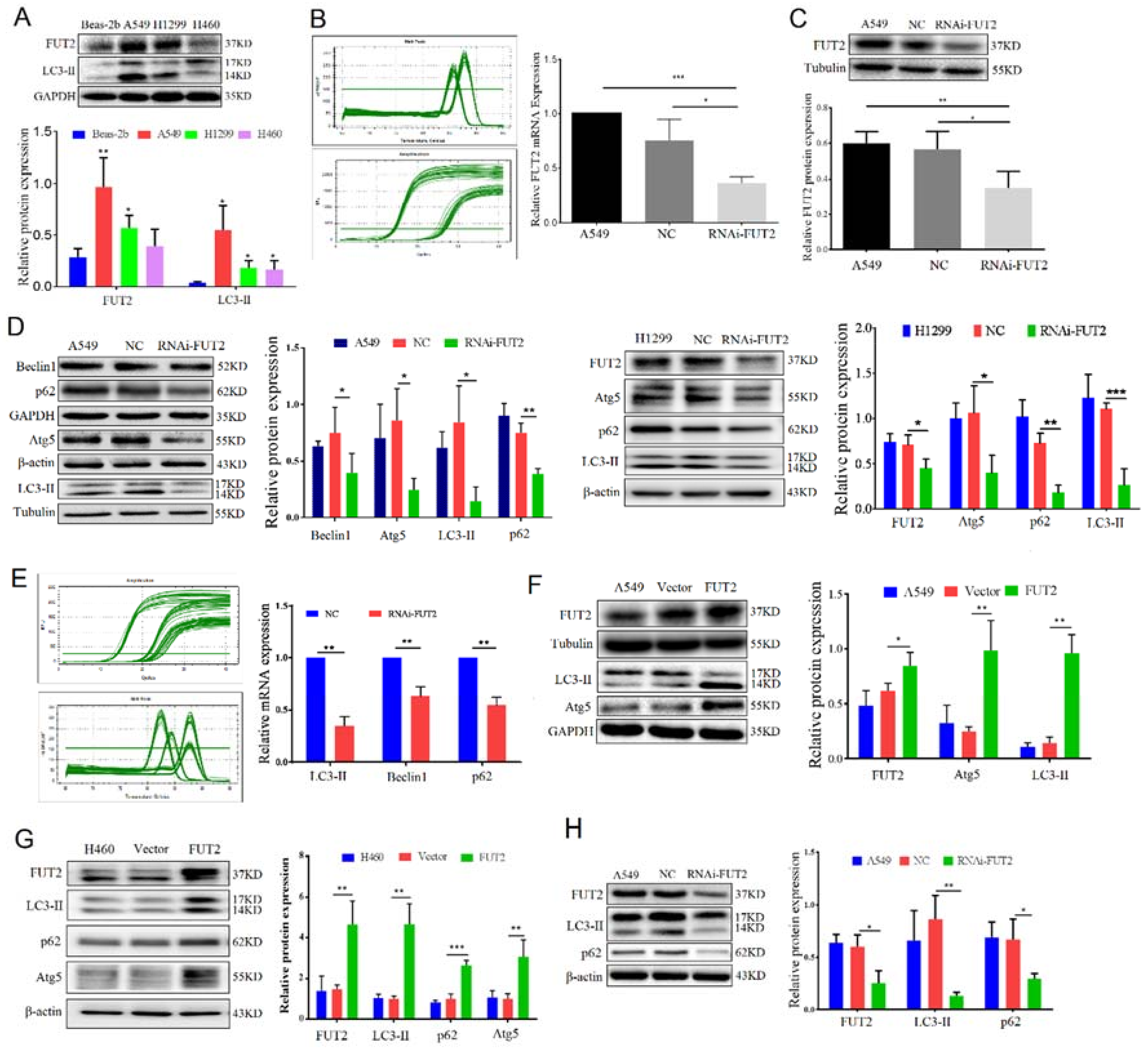
3.3. FUT2 and LC3-II Were Highly Expressed in Lung Adenocarcinoma Cell Lines
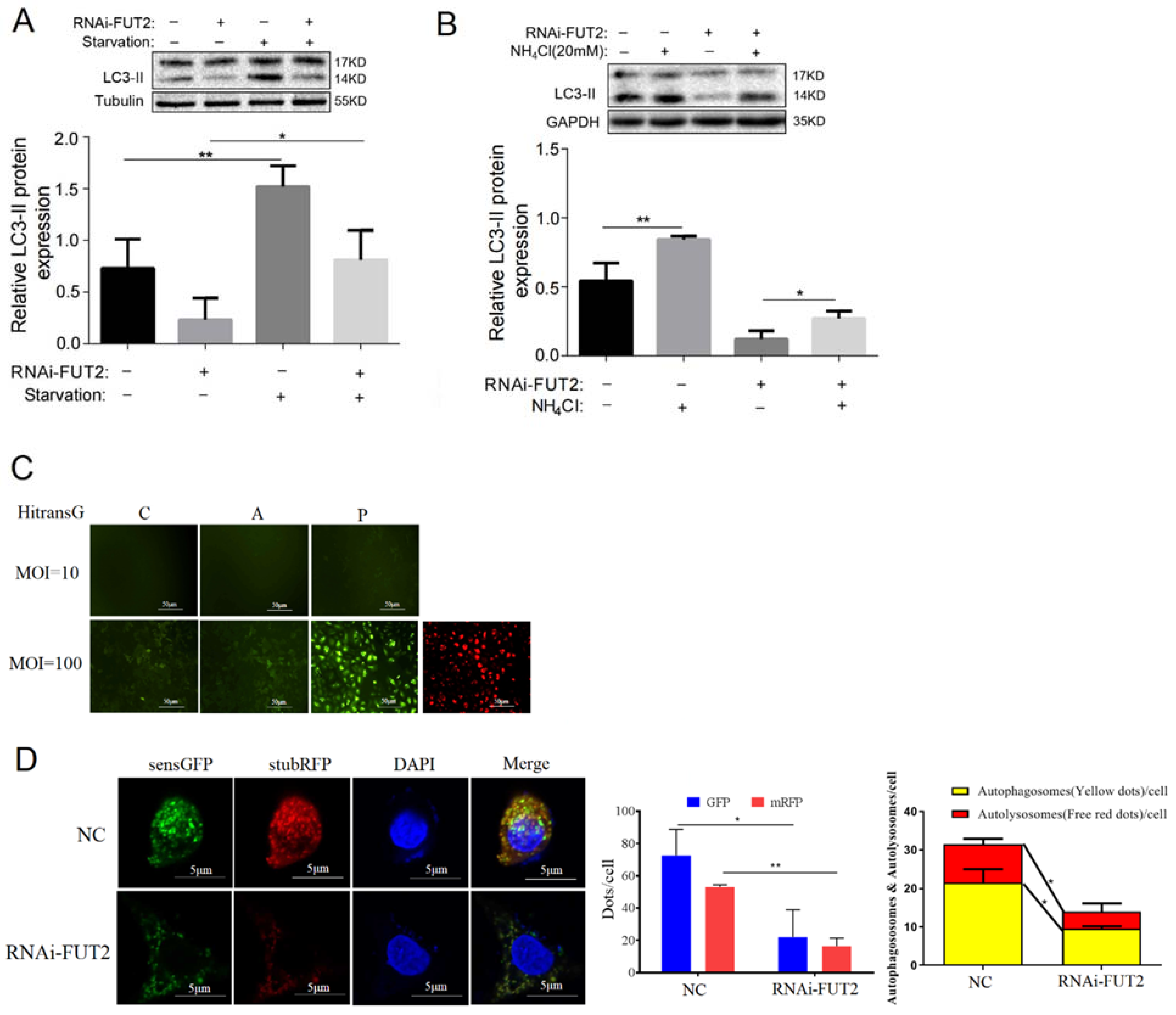
3.4. FUT2 Promoted the Autophagy of Lung Adenocarcinoma Cells
3.5. FUT2 Affected the Early Stage of Autophagy in Lung Adenocarcinoma Cells
3.6. FUT2 Promotes Autophagy by AMPK/ULK1/PI3K III Pathways in LUAD Cells
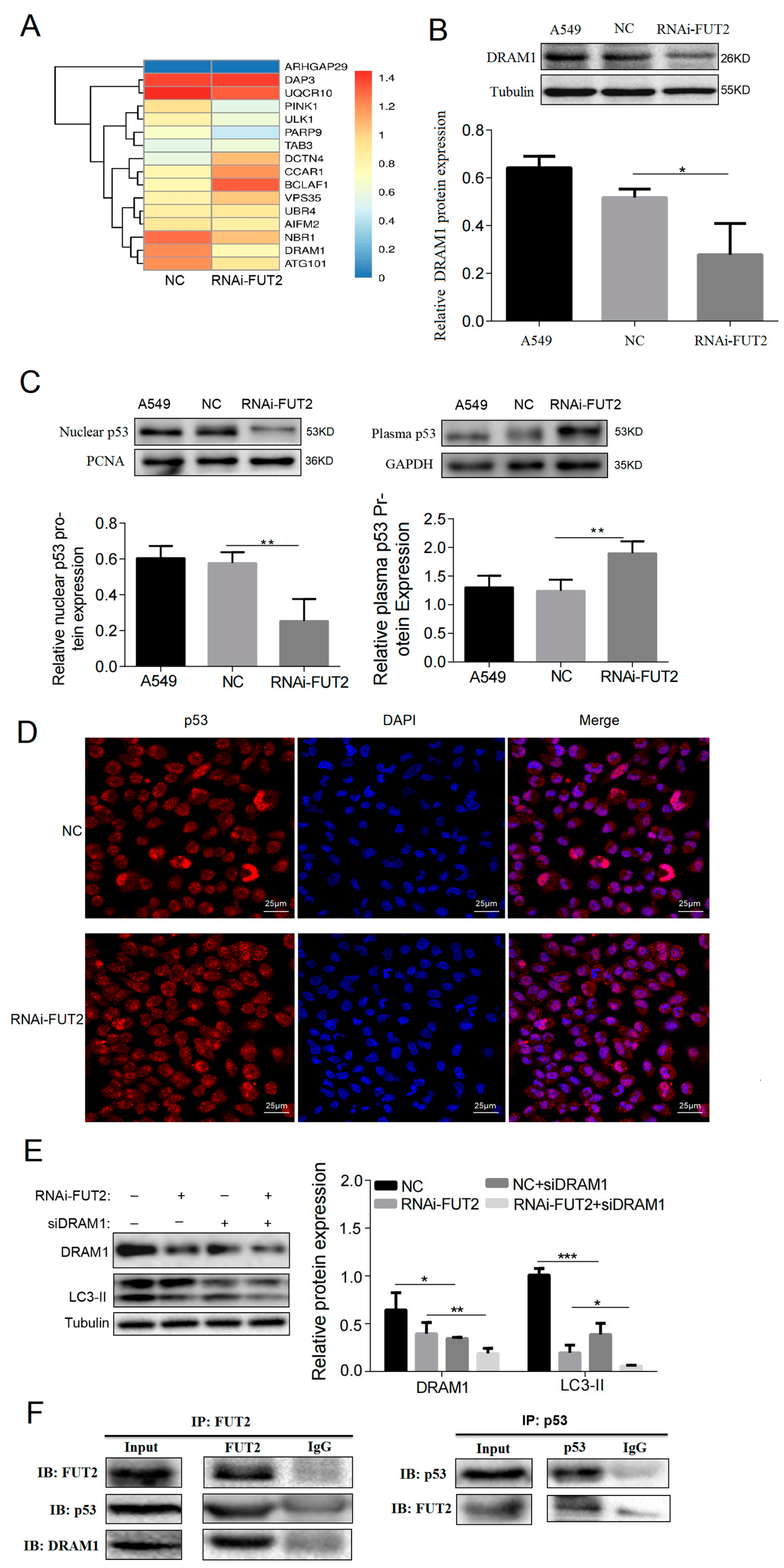
3.7. FUT2 Stimulates Autophagy through p53/DRAM1 Signaling in LUAD Cells
3.8. FUT2 Inhibited the Mitochondrial-Mediated Apoptosis in LUAD Cells
3.9. FUT2 Modulates Apoptosis via JNK Signaling in LUAD Cells
3.10. The Regulation of Autophagy and Apoptosis by FUT2 and Their Antagonism in LUAD Cells
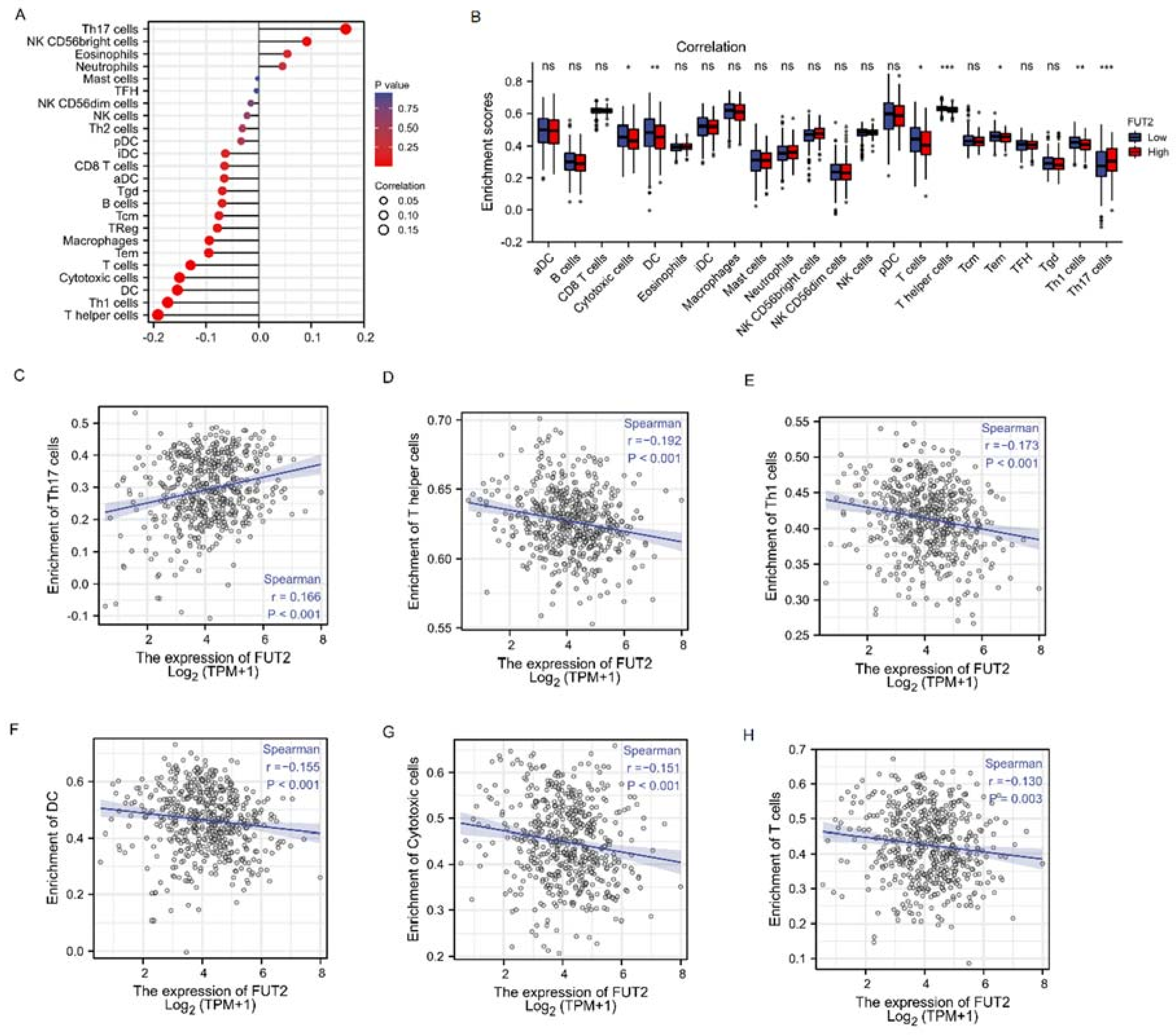
3.11. The Correlation between FUT2 Expression and Immune Infiltration
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics, 2022. CA Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Fehlmann, T.; Kahraman, M.; Ludwig, N.; Backes, C.; Galata, V.; Keller, V.; Geffers, L.; Mercaldo, N.; Hornung, D.; Weis, T.; et al. Evaluating the use of circulating microRNA profiles for lung cancer detection in symptomatic patients. JAMA Oncol. 2020, 6, 714–723. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zhu, J.; Lubman, D.M.; Gao, C. Aberrant glycosylation and cancer biomarker discovery: A promising and thorny journey. Clin. Chem. Lab. Med. 2019, 57, 407–416. [Google Scholar] [CrossRef]
- Munkley, J.; Elliott, D.J. Hallmarks of glycosylation in cancer. Oncotarget 2016, 7, 35478–35489. [Google Scholar] [CrossRef] [Green Version]
- Jia, L.; Zhang, J.; Ma, T.; Guo, Y.; Yu, Y.; Cui, J. The function of fucosylation in progression of lung cancer. Front. Oncol. 2018, 8, 565. [Google Scholar] [CrossRef]
- Blanas, A.; Sahasrabudhe, N.M.; Rodríguez, E.; van Kooyk, Y.; van Vliet, S.J. Fucosylated antigens in cancer: An alliance toward tumor progression, metastasis, and resistance to chemotherapy. Front. Oncol. 2018, 8, 39. [Google Scholar] [CrossRef]
- Honma, R.; Kinoshita, I.; Miyoshi, E.; Tomaru, U.; Matsuno, Y.; Shimizu, Y.; Takeuchi, S.; Kobayashi, Y.; Kaga, K.; Taniguchi, N.; et al. Expression of fucosyltransferase 8 is associated with an unfavorable clinical outcome in non-small cell lung cancers. Oncology 2015, 88, 298–308. [Google Scholar] [CrossRef]
- Lu, H.; Lin, S.Y.; Weng, R.R.; Juan, Y.H.; Chen, Y.W.; Hou, H.H.; Hung, Z.; Oswita, G.A.; Huang, Y.-J.; Guu, S.-H. Fucosyltransferase 4 shapes oncogenic glycoproteome to drive metastasis of lung adenocarcinoma. EBioMedicine 2020, 57, 102846. [Google Scholar] [CrossRef]
- Lai, T.Y.; Chen, I.J.; Lin, R.J.; Liao, G.S.; Yeo, H.L.; Ho, C.L.; Wu, J.; Chang, N.; Lee, A.C.; Yu, A.L. Fucosyltransferase 1 and 2 play pivotal roles in breast cancer cells. Cell Death Discov. 2019, 5, 74. [Google Scholar] [CrossRef]
- Montesino, B.; Steenackers, A.; Lozano, J.M.; Young, G.D.; Hu, N.; Sackstein, R.; Chandler, K.B. Identification of α1,2-fucosylated signaling and adhesion molecules in head and neck squamous cell carcinoma. Glycobiology 2022, 32, 441–455. [Google Scholar] [CrossRef]
- Zhou, W.; Ma, H.; Deng, G.; Tang, L.; Lu, J.; Chen, X. Clinical significance and biological function of fucosyltransferase 2 in lung adenocarcinoma. Oncotarget 2017, 8, 97246–97259. [Google Scholar] [CrossRef] [Green Version]
- Deng, G.; Chen, L.; Zhang, Y.; Fan, S.; Li, W.; Lu, J.; Chen, X. Fucosyltransferase 2 induced epithelial-mesenchymal transition via TGF-β/Smad signaling pathway in lung adenocarcinaoma. Exp. Cell Res. 2018, 370, 613–622. [Google Scholar] [CrossRef]
- White, E. The role for autophagy in cancer. J. Clin. Investig. 2015, 125, 42–46. [Google Scholar] [CrossRef] [Green Version]
- Mulcahy Levy, J.M.; Thorburn, A. Autophagy in cancer: Moving from understanding mechanism to improving therapy responses in patients. Cell Death Differ. 2020, 27, 843–857. [Google Scholar] [CrossRef]
- Das, C.K.; Banerjee, I.; Mandal, M. Pro-survival autophagy: An emerging candidate of tumor progression through maintaining hallmarks of cancer. Semin. Cancer Biol. 2020, 66, 59–74. [Google Scholar] [CrossRef]
- Ruan, H.B.; Ma, Y.; Torres, S.; Zhang, B.; Feriod, C.; Heck, R.M.; Qian, K.; Fu, M.; Li, X.; Nathanson, M.H.; et al. Calcium-dependent O-GlcNAc signaling drives liver autophagy in adaptation to starvation. Gene Dev. 2017, 31, 1655–1665. [Google Scholar] [CrossRef] [Green Version]
- Pyo, K.E.; Kim, C.R.; Lee, M.; Kim, J.; Kim, K.; Baek, S.H. ULK1 O-GlcNAcylation is crucial for activating VPS34 via ATG14L during autophagy initiation. Cell Rep. 2018, 25, 2878–2890. [Google Scholar] [CrossRef] [Green Version]
- Matarrese, P.; Garofalo, T.; Manganelli, V.; Gambardella, L.; Marconi, M.; Grasso, M.; Tinari, A.; Misasi, R.; Malorn, W.; Sorice, M.; et al. Evidence for the involvement of GD3 ganglioside in autophagosome formation and maturation. Autophagy 2014, 10, 750–765. [Google Scholar] [CrossRef]
- Fahie, K.; Zachara, N.E. Molecular Functions of glycoconjugates in autophagy. J. Mol. Biol. 2016, 428, 3305–3324. [Google Scholar] [CrossRef]
- Joshi, S.; Ryan, K.M. Autophagy chews Fap to promote apoptosis. Nat. Cell Biol. 2014, 16, 23–25. [Google Scholar] [CrossRef] [PubMed]
- Mariño, G.; Niso-Santano, M.; Baehrecke, E.H.; Kroemer, G. Self-consumption: The interplay of autophagy and apoptosis. Nat. Rev. Mol. Cell Biol. 2014, 15, 81–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vivian, J.; Rao, A.A.; Nothaft, F.A.; Ketchum, C.; Armstrong, J.; Novak, A.; Pfeil, J.; Narkizian, J.; Deran, A.D.; Musselman-Brown, A.; et al. Toil enables reproducible, open source, big biomedical data analyses. Nat. Biotechnol. 2017, 35, 314–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hänzelmann, S.; Castelo, R.; Guinney, J. GSVA: Gene set variation analysis for microarray and RNA-seq data. BMC Bioinform. 2013, 14, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paquette, M.; El-Houjeiri, L.; Pause, A. mTOR Pathways in Cancer and Autophagy. Cancers 2018, 10, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Kundu, M.; Viollet, B.; Guan, K.L. AMPK and mTOR regulate autophagy through direct phosphorylation of Ulk1. Nat. Cell Biol. 2011, 13, 132–141. [Google Scholar] [CrossRef] [Green Version]
- Crighton, D.; Wilkinson, S.; O’Prey, J.; Syed, N.; Smith, P.; Harrison, P.R.; Gasco, M.; Garrone, O.; Crook, T.; Ryan, K.M. DRAM, a p53-induced modulator of autophagy, is critical for apoptosis. Cell 2006, 126, 121–134. [Google Scholar] [CrossRef] [Green Version]
- Bindea, G.; Mlecnik, B.; Tosolini, M.; Kirilovsky, A.; Waldner, M.; Obenauf, A.C.; Angell, H.; Fredriksen, T.; Lafontaine, L.; Berger, A.; et al. Spatiotemporal dynamics of intratumoral immune cells reveal the immune landscape in human cancer. Immunity 2013, 39, 782–795. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; He, S.; Ma, B. Autophagy and autophagy-related proteins in cancer. Mol. Cancer 2020, 19, 12. [Google Scholar] [CrossRef]
- Onorati, A.V.; Dyczynski, M.; Ojha, R.; Amaravadi, R.K. Targeting autophagy in cancer. Cancer 2018, 124, 3307–3318. [Google Scholar] [CrossRef]
- Russell, R.C.; Tian, Y.; Yuan, H.; Park, H.W.; Chang, Y.Y.; Kim, J.; Kim, H.; Neufeld, T.P.; Dillin, A.; Guan, K. ULK1 induces autophagy by phosphorylating Beclin-1 and activating VPS34 lipid kinase. Nat. Cell Biol. 2013, 15, 741–750. [Google Scholar] [CrossRef] [Green Version]
- Park, J.M.; Jung, C.H.; Seo, M.; Otto, N.M.; Grunwald, D.; Kim, K.H.; Moriarity, B.; Kim, Y.; Starker, C.; Nho, R.S.; et al. The ULK1 complex mediates mTORC1 signaling to the autophagy initiation machinery via binding and phosphorylating ATG14. Autophagy 2016, 12, 547–564. [Google Scholar] [CrossRef] [Green Version]
- Su, Z.; Yang, Z.; Xu, Y.; Chen, Y.; Yu, Q. Apoptosis, autophagy, necroptosis, and cancer metastasis. Mol. Cancer 2015, 14, 48. [Google Scholar] [CrossRef] [Green Version]
- D’Arcy, M.S. Cell death: A review of the major forms of apoptosis, necrosis and autophagy. Cell Biol. Int. 2019, 43, 582–592. [Google Scholar] [CrossRef]
- Ling, Q.; Xu, X.; Wei, X.; Wang, W.; Zhou, B.; Wang, B.; Zheng, S. Oxymatrine induces human pancreatic cancer PANC-1 cells apoptosis via regulating expression of Bcl-2 and IAP families, and releasing of cytochrome c. J. Exp. Clin. Cancer Res. 2011, 30, 66. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.C.; Bratton, S.B. Regulation of the intrinsic apoptosis pathway by reactive oxygen species. Antioxid. Redox Signal. 2013, 19, 546–558. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Qi, F.; Meng, X.; Zhu, C.; Gao, Y. Mst1 regulates colorectal cancer stress response via inhibiting Bnip3-related mitophagy by activation of JNK/p53 pathway. Cell Biol. Toxicol. 2018, 34, 263–277. [Google Scholar] [CrossRef]
- Fang, H.; Yang, K.; Tang, P.; Zhao, N.; Ma, R.; Luo, X.; Liu, Q. Glycosylation end products mediate damage and apoptosis of periodontal ligament stem cells induced by the JNK-mitochondrial pathway. Aging 2020, 12, 12850–12868. [Google Scholar] [CrossRef]
- Tsuruta, F.; Sunayama, J.; Mori, Y.; Hattori, S.; Shimizu, S.; Tsujimoto, Y.; Yoshioka, K.; Masuyama, N.; Gotoh, Y. JNK promotes Bax translocation to mitochondria through phosphorylation of 14-3-3 proteins. EMBO J. 2004, 23, 1889–1899. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Li, X.; Li, Y.; Li, Y.; Wang, Y.; Zhu, L.; Chen, P.; Tian, Z.; Qiu, Y.; Feng, R.; et al. DNA damage-regulated autophagy modulator 1 (DRAM1) mediates autophagy and apoptosis of intestinal epithelial cells in inflammatory bowel disease. Dig. Dis. Sci. 2021, 66, 3375–3390. [Google Scholar] [CrossRef]
- Su, M.; Mei, Y.; Sinha, S. Role of the Crosstalk between Autophagy and Apoptosis in Cancer. J. Oncol. 2013, 2013, 102735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasof, G.M.; Goyal, L.; White, E. Btf, a novel death-promoting transcriptional repressor that interacts with Bcl-2-related proteins. Mol. Cell. Biol. 1994, 19, 4390–4404. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| FUT2 | 5′-GTGGTGTTTGCTGGCGATGG-3′ | 5′-AAAGATTTTGAGGAAAGGGGAGTCG-3′ |
| Beclin1 | 5′-AACCAGATGCGTTATGCCC-3′ | 5′-ATTGATTGTGCCAAACTGTCC-3′ |
| LC3 | 5′-AACATGAGCGAGTTGGTCAAG-3′ | 5′-GCTCGTAGATGTCCGCGAT-3′ |
| P62 | 5′-AGCGTCAGGAAGGTGCCATT-3′ | 5′-CCTTTCTCAAGCCCCATGTTG-3′ |
| GAPDH | 5′-GAACATCATCCCTGCCTCTACT-3′ | 5′-CCTGCTTCACCACCTTCTTG-3′ |
| Characteristic | Low Expression of FUT2 | High Expression of FUT2 | p |
|---|---|---|---|
| n | 267 | 268 | |
| T stage, n (%) | 0.001 | ||
| T1 | 101 (19%) | 74 (13.9%) | |
| T2 | 143 (26.9%) | 146 (27.4%) | |
| T3 | 13 (2.4%) | 36 (6.8%) | |
| T4 | 8 (1.5%) | 11 (2.1%) | |
| N stage, n (%) | 0.522 | ||
| N0 | 178 (34.3%) | 170 (32.8%) | |
| N1 | 44 (8.5%) | 51 (9.8%) | |
| N2 | 36 (6.9%) | 38 (7.3%) | |
| N3 | 0 (0%) | 2 (0.4%) | |
| M stage, n (%) | 0.936 | ||
| M0 | 177 (45.9%) | 184 (47.7%) | |
| M1 | 13 (3.4%) | 12 (3.1%) | |
| Pathologic stage, n (%) | 0.159 | ||
| Stage I | 156 (29.6%) | 138 (26.2%) | |
| Stage II | 51 (9.7%) | 72 (13.7%) | |
| Stage III | 39 (7.4%) | 45 (8.5%) | |
| Stage IV | 14 (2.7%) | 12 (2.3%) | |
| Residual tumor, n (%) | 0.839 | ||
| R0 | 179 (48.1%) | 176 (47.3%) | |
| R1 | 5 (1.3%) | 8 (2.2%) | |
| R2 | 2 (0.5%) | 2 (0.5%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Yao, E.; Liu, Y.; Zhang, Y.; Ding, M.; Liu, J.; Chen, X.; Fan, S. FUT2 Facilitates Autophagy and Suppresses Apoptosis via p53 and JNK Signaling in Lung Adenocarcinoma Cells. Cells 2022, 11, 4031. https://doi.org/10.3390/cells11244031
Zhang Y, Yao E, Liu Y, Zhang Y, Ding M, Liu J, Chen X, Fan S. FUT2 Facilitates Autophagy and Suppresses Apoptosis via p53 and JNK Signaling in Lung Adenocarcinoma Cells. Cells. 2022; 11(24):4031. https://doi.org/10.3390/cells11244031
Chicago/Turabian StyleZhang, Yuqi, Enze Yao, Yijing Liu, Yining Zhang, Mengyang Ding, Jingyu Liu, Xiaoming Chen, and Sairong Fan. 2022. "FUT2 Facilitates Autophagy and Suppresses Apoptosis via p53 and JNK Signaling in Lung Adenocarcinoma Cells" Cells 11, no. 24: 4031. https://doi.org/10.3390/cells11244031
APA StyleZhang, Y., Yao, E., Liu, Y., Zhang, Y., Ding, M., Liu, J., Chen, X., & Fan, S. (2022). FUT2 Facilitates Autophagy and Suppresses Apoptosis via p53 and JNK Signaling in Lung Adenocarcinoma Cells. Cells, 11(24), 4031. https://doi.org/10.3390/cells11244031