

Fat Quality Impacts the Effect of a High-Fat Diet on the Fatty Acid Profile, Life History Traits and Gene Expression in Drosophila melanogaster

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Fly Strains and Husbandry
2.2. Synchronization of Eggs
2.3. Preparation of High-Fat Diets
2.4. Determination of the Fatty Acid Profile of Fruit Flies and Supplemented Oils and Fats
2.5. Determination of Food Intake
2.6. Determination of Body Weight and Body Composition
2.7. Lifespan Experiments and Determination of Gut Integrity
2.8. Induced Climbing Activity Assay
2.9. Determination of Spontaneous Locomotor Activity
2.10. Determination of Female Fecundity
2.11. Egg-Laying Preference Assay
2.12. Determination of the Development Rate
2.13. Determination of the Abdominal Width and Ovarian Area
2.14. Isolation and Sequencing of RNA
2.15. Statistical Analysis
3. Results
3.1. Increasing Concentrations of Dietary Butterfat Significantly Increased the Energy Intake without Affecting the Body Composition of Male and Female D. melanogaster
3.2. Female Fecundity Decreased in a Dose-Dependent Manner in Response to Butterfat Supplementation
3.3. Delayed Egg to Adult Development at High Dietary Butterfat Concentrations
3.4. Increasing Concentrations of Dietary Butterfat Did Not Affect the Spontaneous Locomotion Activity of Male and Female Fruit Flies
3.5. Dietary Butterfat Shortened the Lifespan of Both, Male and Female w1118 Flies in a Concentration-Dependent Manner
3.6. Different High-Fat Diet Qualities Led to Fat Source-Specific Shifts in the Fatty Acid Profile of D. melanogaster
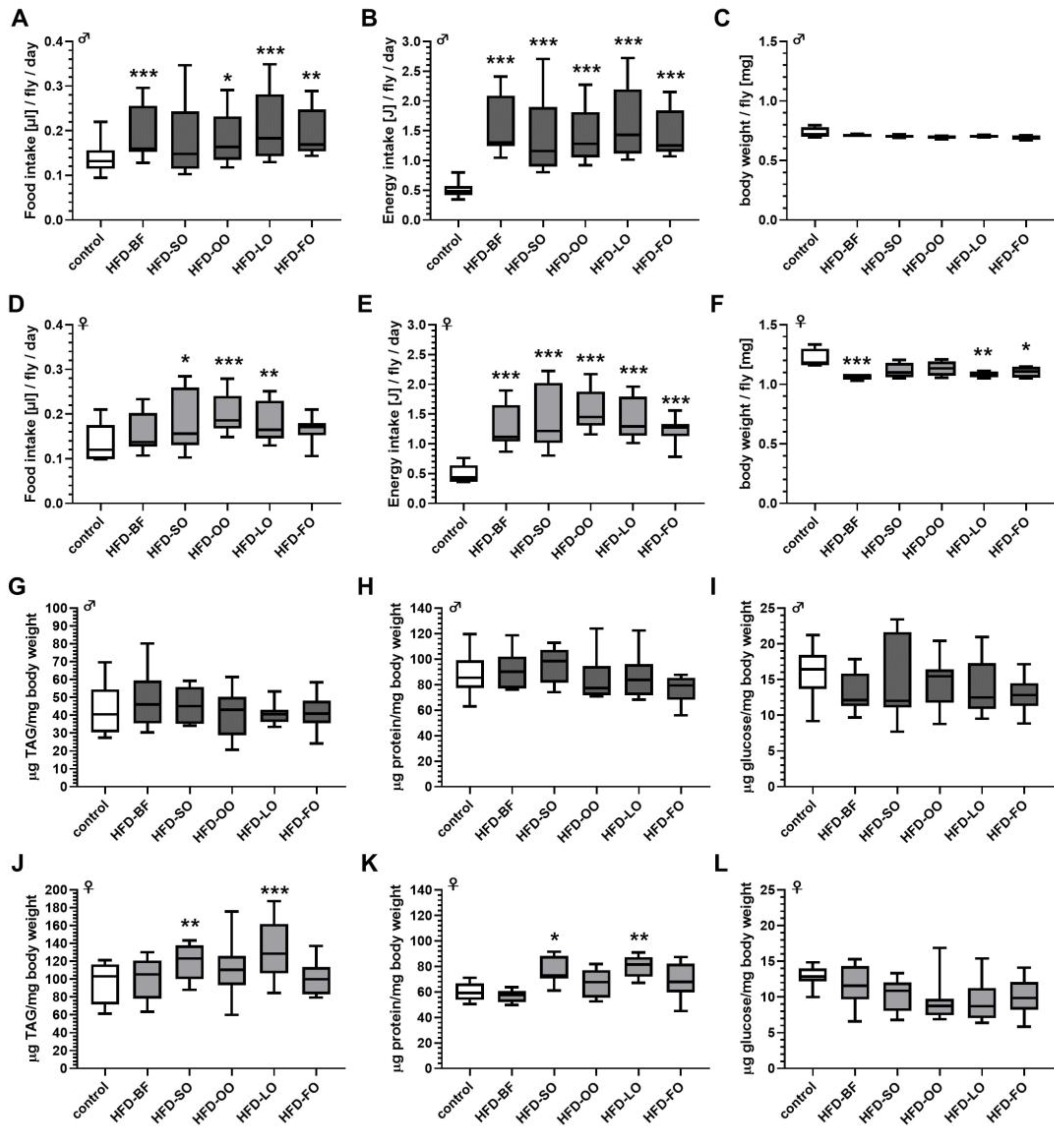
3.7. High-Fat Diet Feeding Resulted in an Increased Energy Intake Independent of the Fat Quality
3.8. Body Weight and Body Composition Were Altered by Ingestion of High-Fat Diets Comprising Different Fat Qualities Solely in Female Fruit Flies
3.9. High-Fat Diets Based on Animal Fats Had a Negative Impact on the Egg Laying Rate in D. melanogaster
3.10. An Olive Oil Based High-Fat Diet Was Preferred for Oviposition
3.11. High-Fat Diets Slightly Delayed the Development of D. melanogaster without Affecting the Developmental Rate
3.12. The Fat Quality of High-Fat Diets Affected the Degree of Lifespan Reduction in Fruit Flies
3.13. High-Fat Diets Affected the Climbing Activity of Fruit Flies in a Fat Quality-Dependent Manner
3.14. The Spontaneous Locomotor Activities of Male and Female Fruit Flies Were Differently Affected by Short-Term Exposure to Plant Oil-Based High-Fat Diets
3.15. Fat Quality Had a Large Influence on the Transcriptional Response of D. melanogaster to a High-Fat Diet
3.16. The Small Overlap of Genes Differentially Expressed in Response to All of the Three High-Fat Diet Qualities Were Associated with the Functional Terms Immune/Stress Response and Fatty Acid Metabolism
3.17. Feeding an HFD Containing 12% Butterfat Had a Negative Impact on Gut Integrity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- NCD-RisC. Trends in adult body-mass index in 200 countries from 1975 to 2014: A pooled analysis of 1698 population-based measurement studies with 19·2 million participants. Lancet 2016, 387, 1377–1396. [Google Scholar] [CrossRef] [PubMed]
- WHO. Noncommunicable Diseases Country Profiles 2018. Available online: https://apps.who.int/iris/handle/10665/274512 (accessed on 10 November 2022).
- Kopp, W. How western diet and lifestyle drive the pandemic of obesity and civilization diseases. Diabetes Metab. Syndr. Obes. Targets Ther. 2019, 12, 2221–2236. [Google Scholar] [CrossRef] [PubMed]
- Meijaard, E.; Abrams, J.F.; Slavin, J.L.; Sheil, D. Dietary fats, human nutrition and the environment: Balance and sustainability. Front. Nutr. 2022, 9, 878644. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Liu, H. Nutritional indices for assessing fatty acids: A mini-review. Int. J. Mol. Sci. 2020, 21, 5695. [Google Scholar] [CrossRef]
- WHO. Obesity and Overweight. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 10 November 2022).
- Gillingham, L.G.; Harris-Janz, S.; Jones, P.J. Dietary monounsaturated fatty acids are protective against metabolic syndrome and cardiovascular disease risk factors. Lipids 2011, 46, 209–228. [Google Scholar] [CrossRef]
- Beulen, Y.; Martínez-González, M.; van de Rest, O.; Salas-Salvadó, J.; Sorlí, J.; Gómez-Gracia, E.; Fiol, M.; Estruch, R.; Santos-Lozano, J.; Schröder, H.; et al. Quality of dietary fat intake and body weight and obesity in a mediterranean population: Secondary analyses within the predimed trial. Nutrients 2018, 10, 2011. [Google Scholar] [CrossRef]
- Neuenschwander, M.; Barbaresko, J.; Pischke, C.R.; Iser, N.; Beckhaus, J.; Schwingshackl, L.; Schlesinger, S.; Basu, S. Intake of dietary fats and fatty acids and the incidence of type 2 diabetes: A systematic review and dose-response meta-analysis of prospective observational studies. PLoS Med. 2020, 17, e1003347. [Google Scholar] [CrossRef]
- Hooper, L. Dietary fat intake and prevention of cardiovascular disease: Systematic review. BMJ 2001, 322, 757–763. [Google Scholar] [CrossRef]
- Gáliková, M.; Klepsatel, P. Obesity and aging in the Drosophila model. Int. J. Mol. Sci. 2018, 19, 1896. [Google Scholar] [CrossRef]
- Eickelberg, V.; Lüersen, K.; Staats, S.; Rimbach, G. Phenotyping of Drosophila melanogaster—A nutritional perspective. Biomolecules 2022, 12, 221. [Google Scholar] [CrossRef]
- Bier, E. Drosophila, the golden bug, emerges as a tool for human genetics. Nat. Rev. Genet. 2005, 6, 9–23. [Google Scholar] [CrossRef] [PubMed]
- Lachaise, D.; Cariou, M.L.; David, J.R.; Lemeunier, F.; Tsacas, L.; Ashburner, M. Historical biogeography of the Drosophila melanogaster species subgroup. J. Evol. Biol. 1988, 22, 159–225. [Google Scholar]
- Yang, D. Carnivory in the larvae of Drosophila melanogaster and other Drosophila species. Sci. Rep. 2018, 8, 15484. [Google Scholar] [CrossRef] [PubMed]
- Miguel-Aliaga, I.; Jasper, H.; Lemaitre, B. Anatomy and physiology of the digestive tract of Drosophila melanogaster. Genetics 2018, 210, 357–396. [Google Scholar] [CrossRef] [PubMed]
- Horne, I.; Haritos, V.S.; Oakeshott, J.G. Comparative and functional genomics of lipases in holometabolous insects. Insect Biochem. Mol. Biol. 2009, 39, 547–567. [Google Scholar] [CrossRef]
- Lüersen, K.; Röder, T.; Rimbach, G. Drosophila melanogaster in nutrition research—The importance of standardizing experimental diets. Genes Nutr. 2019, 14, 3. [Google Scholar] [CrossRef]
- Wen, D.-T.; Zheng, L.; Yang, F.; Li, H.-Z.; Hou, W.-Q. Endurance exercise prevents high-fat-diet induced heart and mobility premature aging and dsir2 expression decline in aging Drosophila. Oncotarget 2018, 9, 7298–7311. [Google Scholar] [CrossRef]
- Liao, S.; Amcoff, M.; Nässel, D.R. Impact of high-fat diet on lifespan, metabolism, fecundity and behavioral senescence in Drosophila. Insect Biochem. Mol. Biol. 2020, 133, 103495. [Google Scholar] [CrossRef]
- Birse, R.T.; Choi, J.; Reardon, K.; Rodriguez, J.; Graham, S.; Diop, S.; Ocorr, K.; Bodmer, R.; Oldham, S. High-fat-diet-induced obesity and heart dysfunction are regulated by the tor pathway in Drosophila. Cell Metab. 2010, 12, 533–544. [Google Scholar] [CrossRef]
- Rivera, O.; McHan, L.; Konadu, B.; Patel, S.; Sint Jago, S.; Talbert, M.E. A high-fat diet impacts memory and gene expression of the head in mated female Drosophila melanogaster. J. Comp. Physiol. B 2019, 189, 179–198. [Google Scholar] [CrossRef]
- Villanueva, J.E.; Livelo, C.; Trujillo, A.S.; Chandran, S.; Woodworth, B.; Andrade, L.; Le, H.D.; Manor, U.; Panda, S.; Melkani, G.C. Time-restricted feeding restores muscle function in Drosophila models of obesity and circadian-rhythm disruption. Nat. Commun. 2019, 10, 2700. [Google Scholar] [CrossRef] [PubMed]
- Schultzhaus, J.N.; Bennett, C.J.; Iftikhar, H.; Yew, J.Y.; Mallett, J.; Carney, G.E. High fat diet alters Drosophila melanogaster sexual behavior and traits: Decreased attractiveness and changes in pheromone profiles. Sci. Rep. 2018, 8, 5387. [Google Scholar] [CrossRef] [PubMed]
- Heinrichsen, E.T.; Haddad, G.G.; Missirlis, F. Role of high-fat diet in stress response of Drosophila. PLoS ONE 2012, 7, e42587. [Google Scholar] [CrossRef] [PubMed]
- Diop, S.B.; Bisharat-Kernizan, J.; Birse, R.T.; Oldham, S.; Ocorr, K.; Bodmer, R. Pgc-1/spargel counteracts high-fat-diet-induced obesity and cardiac lipotoxicity downstream of tor and brummer atgl lipase. Cell Rep. 2015, 10, 1572–1584. [Google Scholar] [CrossRef] [PubMed]
- Kayashima, Y.; Murata, S.; Sato, M.; Matsuura, K.; Asanuma, T.; Chimoto, J.; Ishii, T.; Mochizuki, K.; Kumazawa, S.; Nakayama, T.; et al. Tea polyphenols ameliorate fat storage induced by high-fat diet in Drosophila melanogaster. Biochem. Biophys. Rep. 2015, 4, 417–424. [Google Scholar] [CrossRef] [PubMed]
- Woodcock, K.J.; Kierdorf, K.; Pouchelon, C.A.; Vivancos, V.; Dionne, M.S.; Geissmann, F. Macrophage-derived upd3 cytokine causes impaired glucose homeostasis and reduced lifespan in Drosophila fed a lipid-rich diet. Immunity 2015, 42, 133–144. [Google Scholar] [CrossRef]
- Jung, J.; Kim, D.-I.; Han, G.-Y.; Kwon, H. The effects of high fat diet-induced stress on olfactory sensitivity, behaviors, and transcriptional profiling in Drosophila melanogaster. Int. J. Mol. Sci. 2018, 19, 2855. [Google Scholar] [CrossRef]
- Lindmark Månsson, H. Fatty acids in bovine milk fat. Food Nutr. Res. 2008, 52, 1821. [Google Scholar] [CrossRef]
- Diop, S.; Birse, R.; Bodmer, R. High fat diet feeding and high throughput triacylglyceride assay in Drosophila melanogaster. J. Vis. Exp. 2017, 127, 56029. [Google Scholar] [CrossRef]
- Staats, S.; Wagner, A.E.; Lüersen, K.; Künstner, A.; Meyer, T.; Kahns, A.K.; Derer, S.; Graspeuntner, S.; Rupp, J.; Busch, H.; et al. Dietary ursolic acid improves health span and life span in male Drosophila melanogaster. BioFactors 2018, 45, 169–186. [Google Scholar] [CrossRef]
- Linford, N.J.; Bilgir, C.; Ro, J.; Pletcher, S.D. Measurement of lifespan in Drosophila melanogaster. J. Vis. Exp. 2013, 71, 50068. [Google Scholar]
- Wu, Q.; Yu, G.; Park, S.J.; Gao, Y.; Ja, W.W.; Yang, M. Excreta quantification (ex-q) for longitudinal measurements of food intake in Drosophila. iScience 2020, 23, 100776. [Google Scholar] [CrossRef] [PubMed]
- Rera, M.; Clark, R.I.; Walker, D.W. Intestinal barrier dysfunction links metabolic and inflammatory markers of aging to death in Drosophila. PNAS 2012, 109, 21528–21533. [Google Scholar] [CrossRef]
- Madabattula, S.; Strautman, J.; Bysice, A.; O’Sullivan, J.; Androschuk, A.; Rosenfelt, C.; Doucet, K. Quantitative analysis of climbing defects in a Drosophila model of neurodegenerative disorders. J. Vis. Exp. 2015, 100, e52741. [Google Scholar] [CrossRef]
- Chiu, J.C.; Low, K.H.; Pike, D.H.; Yildirim, E.; Edery, I. Assaying locomotor activity to study circadian rhythms and sleep parameters in Drosophila. J. Vis. Exp. 2010, 43, 2157. [Google Scholar] [CrossRef] [PubMed]
- Barnes, A.I.; Wigby, S.; Boone, J.M.; Partridge, L.; Chapman, T. Feeding, fecundity and lifespan in female Drosophila melanogaster. Proc. Biol. Sci. 2008, 275, 1675–1683. [Google Scholar] [CrossRef]
- Karageorgi, M.; Bräcker, L.B.; Lebreton, S.; Minervino, C.; Cavey, M.; Siju, K.P.; Grunwald Kadow, I.C.; Gompel, N.; Prud’homme, B. Evolution of multiple sensory systems drives novel egg-laying behavior in the fruit pest Drosophila suzukii. Curr. Biol. 2017, 27, 847–853. [Google Scholar] [CrossRef] [PubMed]
- Gompel, N.; Schröder, E.A. Fly Imaging. Available online: http://gompel.org/methods (accessed on 1 November 2021).
- Weil, T.T.; Parton, R.M.; Davis, I. Preparing individual Drosophila egg chambers for live imaging. J. Vis. Exp. 2012, 60, 3679. [Google Scholar] [CrossRef]
- Kaufman, J.D.; Seidler, Y.; Bailey, H.R.; Whitacre, L.; Bargo, F.; Lüersen, K.; Rimbach, G.; Pighetti, G.M.; Ipharraguerre, I.R.; Ríus, A.G. A postbiotic from Aspergillus oryzae attenuates the impact of heat stress in ectothermic and endothermic organisms. Sci. Rep. 2021, 11, 6407. [Google Scholar] [CrossRef]
- Pinheiro, J.C.; Bates, D.M. Mixed-Effects Models in S and S-Plus, 1st ed.; Springer: New York, NY, USA, 2000. [Google Scholar]
- Nakagawa, S.; Schielzeth, H.; O’Hara, R.B. A general and simple method for obtaining r2 from generalized linear mixed-effects models. Methods Ecol. Evol. 2013, 4, 133–142. [Google Scholar] [CrossRef]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [PubMed]
- Scarpati, M.; Qi, Y.; Govind, S.; Singh, S. A combined computational strategy of sequence and structural analysis predicts the existence of a functional eicosanoid pathway in Drosophila melanogaster. PLoS One 2019, 14, e0211897. [Google Scholar] [CrossRef]
- Gutierrez, E.; Wiggins, D.; Fielding, B.; Gould, A.P. Specialized hepatocyte-like cells regulate drosophila lipid metabolism. Nature 2007, 445, 275–280. [Google Scholar] [CrossRef]
- Stobdan, T.; Sahoo, D.; Azad, P.; Hartley, I.; Heinrichsen, E.; Zhou, D.; Haddad, G.G.; Skoulakis, E.M.C. High fat diet induces sex-specific differential gene expression in Drosophila melanogaster. PLoS ONE 2019, 14, e0213474. [Google Scholar] [CrossRef]
- Azuma, M.; Fat Le, T.; Yoshimoto, Y.; Hiraki, N.; Yamanaka, M.; Omura, F.; Inoue, Y.H. Rna-seq analysis of diet-driven obesity and anti-obesity effects of quercetin glucoside or epigallocatechin gallate in Drosophila adults. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 857–876. [Google Scholar]
- Warwick, Z.S.; Schiffman, S.S. Role of dietary fat in calorie intake and weight gain. Neurosci. Biobehav. Rev. 1992, 16, 585–596. [Google Scholar] [CrossRef]
- Han, S.; Schroeder, E.A.; Silva-Garcia, C.G.; Hebestreit, K.; Mair, W.B.; Brunet, A. Mono-unsaturated fatty acids link h3k4me3 modifiers to C. elegans lifespan. Nature 2017, 544, 185–190. [Google Scholar] [CrossRef]
- O’Rourke, E.J.; Kuballa, P.; Xavier, R.; Ruvkun, G. Omega-6 polyunsaturated fatty acids extend life span through the activation of autophagy. Genes Dev. 2013, 27, 429–440. [Google Scholar] [CrossRef]
- Gargano, J.W.; Martin, I.; Bhandari, P.; Grotewiel, M.S. Rapid iterative negative geotaxis (ring): A new method for assessing age-related locomotor decline in Drosophila. Exp. Gerontol. 2005, 40, 386–395. [Google Scholar] [CrossRef]
- Musselman, L.P.; Kuhnlein, R.P. Drosophila as a model to study obesity and metabolic disease. J. Exp. Biol. 2018, 221, jeb163881. [Google Scholar] [CrossRef]
- Hohos, N.M.; Cho, K.J.; Swindle, D.C.; Skaznik-Wikiel, M.E. High-fat diet exposure, regardless of induction of obesity, is associated with altered expression of genes critical to normal ovulatory function. Mol. Cell. Endocrinol. 2018, 470, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Raubenheimer, D.; Machovsky-Capuska, G.E.; Chapman, C.A.; Rothman, J.M. Geometry of nutrition in field studies: An illustration using wild primates. Oecologia 2015, 177, 223–234. [Google Scholar] [CrossRef] [PubMed]
- Jang, T.; Lee, K.P. Comparing the impacts of macronutrients on life-history traits in larval and adult Drosophila melanogaster: The use of nutritional geometry and chemically defined diets. J. Exp. Biol. 2018, 221, jeb181115. [Google Scholar] [CrossRef] [PubMed]
- Fougeron, A.S.; Farine, J.P.; Flaven-Pouchon, J.; Everaerts, C.; Ferveur, J.F. Fatty-acid preference changes during development in Drosophila melanogaster. PLoS ONE 2011, 6, e26899. [Google Scholar] [CrossRef] [PubMed]
- Heinrichsen, E.T.; Zhang, H.; Robinson, J.E.; Ngo, J.; Diop, S.; Bodmer, R.; Joiner, W.J.; Metallo, C.M.; Haddad, G.G. Metabolic and transcriptional response to a high-fat diet in Drosophila melanogaster. Mol. Metab. 2014, 3, 42–54. [Google Scholar] [CrossRef] [PubMed]
- Hemphill, W.; Rivera, O.; Talbert, M. RNA-sequencing of Drosophila melanogaster head tissue on high-sugar and high-fat diets. G3 (Bethesda) 2018, 8, 279–290. [Google Scholar] [CrossRef] [PubMed]
- Fawley, J.; Gourlay, D.M. Intestinal alkaline phosphatase: A summary of its role in clinical disease. J. Surg. Res. 2016, 202, 225–234. [Google Scholar] [CrossRef]
- Obata, F.; Kuranaga, E.; Tomioka, K.; Ming, M.; Takeishi, A.; Chen, C.H.; Soga, T.; Miura, M. Necrosis-driven systemic immune response alters sam metabolism through the foxo-gnmt axis. Cell Rep. 2014, 7, 821–833. [Google Scholar] [CrossRef]
- Georgescu, B.; Boaru, A.M.; Muntean, L.; Sima, N.; Struti, D.I.; Papuc, T.A.; Georgescu, C. Modulating the fatty acid profiles of Hermetia illucens larvae fats by dietary enrichment with different oilseeds: A sustainable way for future use in feed and food. Insects 2022, 13, 801. [Google Scholar] [CrossRef]
- Shen, L.R.; Lai, C.Q.; Feng, X.; Parnell, L.D.; Wan, J.B.; Wang, J.D.; Li, D.; Ordovas, J.M.; Kang, J.X. Drosophila lacks c20 and c22 pufas. J. Lipid Res. 2010, 51, 2985–2992. [Google Scholar] [CrossRef]
- Carvalho, M.; Sampaio, J.L.; Palm, W.; Brankatschk, M.; Eaton, S.; Shevchenko, A. Effects of diet and development on the Drosophila lipidome. Mol. Syst. Biol. 2012, 8, 600. [Google Scholar] [CrossRef] [PubMed]
- Raj, R.R.; Lofquist, S.; Lee, M.J. Remodeling of adipose tissues by fatty acids: Mechanistic update on browning and thermogenesis by n-3 polyunsaturated fatty acids. Pharm. Res. 2022; ahead of print. [Google Scholar]
- Qiu, J.; Ma, Y.; Qiu, J. Regulation of intestinal immunity by dietary fatty acids. Mucosal Immunol. 2022, 15, 846–856. [Google Scholar] [CrossRef] [PubMed]
- Obniski, R.; Sieber, M.; Spradling, A.C. Dietary lipids modulate notch signaling and influence adult intestinal development and metabolism in Drosophila. Dev. Cell 2018, 47, 98–111.e5. [Google Scholar] [CrossRef] [PubMed]
- Von Frieling, J.; Faisal, M.N.; Sporn, F.; Pfefferkorn, R.; Nolte, S.S.; Sommer, F.; Rosenstiel, P.; Roeder, T. A high-fat diet induces a microbiota-dependent increase in stem cell activity in the Drosophila intestine. PLoS Genet. 2020, 16, e1008789. [Google Scholar] [CrossRef] [PubMed]
- Broderick, N.A.; Buchon, N.; Lemaitre, B.; McFall-Ngai, M.J. Microbiota-induced changes in Drosophila melanogaster host gene expression and gut morphology. mBio 2014, 5, e01117-14. [Google Scholar] [CrossRef] [PubMed]
- Burcelin, R. Gut microbiota and immune crosstalk in metabolic disease. Mol. Metab. 2016, 5, 771–781. [Google Scholar] [CrossRef]
- Zaidman-Rémy, A.; Poidevin, M.; Hervé, M.; Welchman, D.P.; Paredes, J.C.; Fahlander, C.; Steiner, H.; Mengin-Lecreulx, D.; Lemaitre, B.; Bereswill, S. Drosophila immunity: Analysis of pgrp-sb1 expression, enzymatic activity and function. PLoS ONE 2011, 6, e17231. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Males | Females | |||||
|---|---|---|---|---|---|---|
| Median | Mean | Maximum | Median | Mean | Maximum | |
| Control | 55.0 ± 2.1 d | 54.0 ± 2.0 d | 85.3 ± 4.6 d | 62.7 ± 3.4 d | 60.7 ± 4.3 d | 97.0 ± 3.1 d |
| HFD-3 | −5.5% | −4.9% | −2.3% | −5.9% | −5.5% | −6.5% |
| HFD-6 | −5.5% | −5.6% | −4.7% | −8.5% | −10.4% | −8.9% |
| HFD-9 | −10.3% | −8.6% | −4.7% | −16.0% | −14.3% | −13.1% |
| HFD-12 | −10.9% | −12.3% | −13.3% | −22.9% | −19.2% | −10.7% |
| Males | Females | |||||
|---|---|---|---|---|---|---|
| Median | Mean | Maximum | Median | Mean | Maximum | |
| Control | 52.0 | 50.0 | 75.5 | 56.0 | 55.5 | 82.5 |
| HFD-BF | −29% | −24% | −9% | −63% | −56% | −27% |
| HFD-SO | −38% | −34% | −20% | −82% | −77% | −48% |
| HFD-OO | −44% | −36% | −17% | −84% | −82% | −54% |
| HFD-LO | −39% | −32% | −25% | −74% | −72% | −56% |
| HFD-FO | −29% | −26% | −17% | −71% | −69% | −39% |
| HFD-BF vs. Control (FDR ≤ 0.1) | |||
| Functional cluster/ Orphan categories | Number of Genes | p-Value | Benjamini |
| Upregulated (39) | |||
| Innate immunity/stress response | 17 (+11 *) | 1.5 × 10−24 | 4.6 × 10−24 |
| FA metabolism/FA elongation | 2 | n.d. | n.d. |
| Downregulated (0) | |||
| (no cluster identified) | - | - | - |
| HFD-FO vs. Control (FDR ≤ 0.1) | |||
| Functional Cluster/ Orphan categories | Number of Genes | p-Value | Benjamini |
| Upregulated (23) | |||
| Innate immunity/stress response | 5 (+1 *) | 3.7 × 10−7 | 7.3 × 10−7 |
| Monooxygenase activity | 3 | 6.6 × 10−3 | 9.5 × 10−2 |
| FA metabolism/FA elongation | 0 (+5 *) | n.d. | n.d. |
| Alkaline phosphatase activity | 1 (+2 *) | n.d. | n.d. |
| Downregulated (43) | |||
| Proteolysis | 11 | 3.0 × 10−8 | 8.0 × 10−7 |
| Lipase | 3 (+1 *) | 4.6 × 10−3 | 2.5 × 10−2 |
| Odorant binding | 3 | 6.8 × 10−3 | 3.2 × 10−2 |
| HFD-SO vs. Control (FDR ≤ 0.1) | |||
| Functional Cluster/ Orphan categories | Number of Genes | p-Value | Benjamini |
| Upregulated (11) | |||
| FA metabolism/FA elongation | 1 (+3 *) | n.d. | n.d. |
| Downregulated (4) | |||
| (no cluster identified) | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eickelberg, V.; Rimbach, G.; Seidler, Y.; Hasler, M.; Staats, S.; Lüersen, K. Fat Quality Impacts the Effect of a High-Fat Diet on the Fatty Acid Profile, Life History Traits and Gene Expression in Drosophila melanogaster. Cells 2022, 11, 4043. https://doi.org/10.3390/cells11244043
Eickelberg V, Rimbach G, Seidler Y, Hasler M, Staats S, Lüersen K. Fat Quality Impacts the Effect of a High-Fat Diet on the Fatty Acid Profile, Life History Traits and Gene Expression in Drosophila melanogaster. Cells. 2022; 11(24):4043. https://doi.org/10.3390/cells11244043
Chicago/Turabian StyleEickelberg, Virginia, Gerald Rimbach, Yvonne Seidler, Mario Hasler, Stefanie Staats, and Kai Lüersen. 2022. "Fat Quality Impacts the Effect of a High-Fat Diet on the Fatty Acid Profile, Life History Traits and Gene Expression in Drosophila melanogaster" Cells 11, no. 24: 4043. https://doi.org/10.3390/cells11244043
APA StyleEickelberg, V., Rimbach, G., Seidler, Y., Hasler, M., Staats, S., & Lüersen, K. (2022). Fat Quality Impacts the Effect of a High-Fat Diet on the Fatty Acid Profile, Life History Traits and Gene Expression in Drosophila melanogaster. Cells, 11(24), 4043. https://doi.org/10.3390/cells11244043