
The Effect of Variable Light Source and Light Intensity on the Growth of Three Algal Species



Abstract
:1. Introduction
2. Materials and Methods
2.1. Microalgae Strains
2.2. Culture Conditions
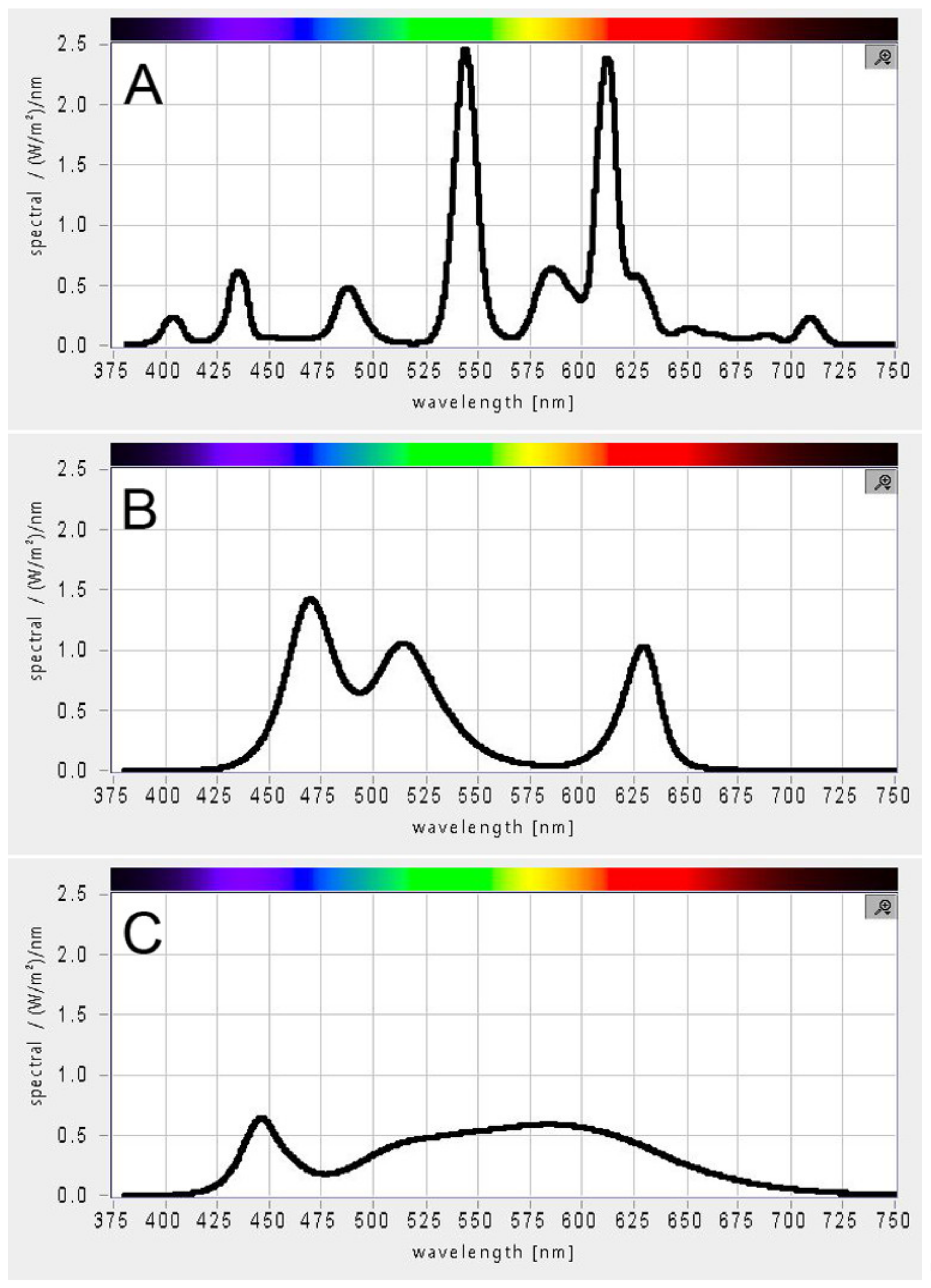
2.3. Light Spectrum and Intensity Measurement
2.4. Quantum Yield Measurement
2.5. Cell Volume and Number
2.6. Cell Doubling Time
2.7. Photosynthetic Pigment Contents
2.8. Statistical Analysis
3. Results
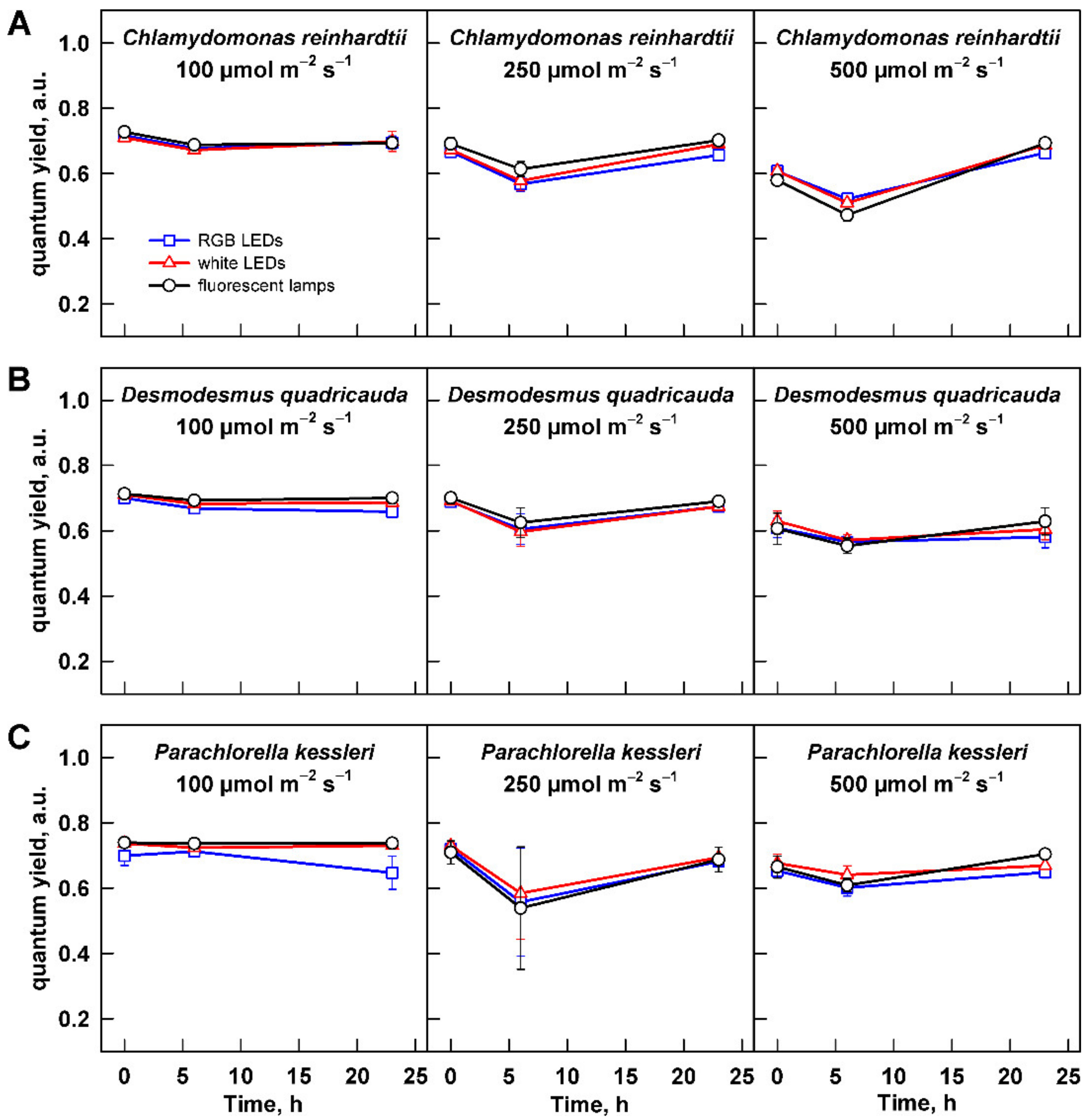
3.1. Maximum Potential Quantum Efficiency of Photosystem II
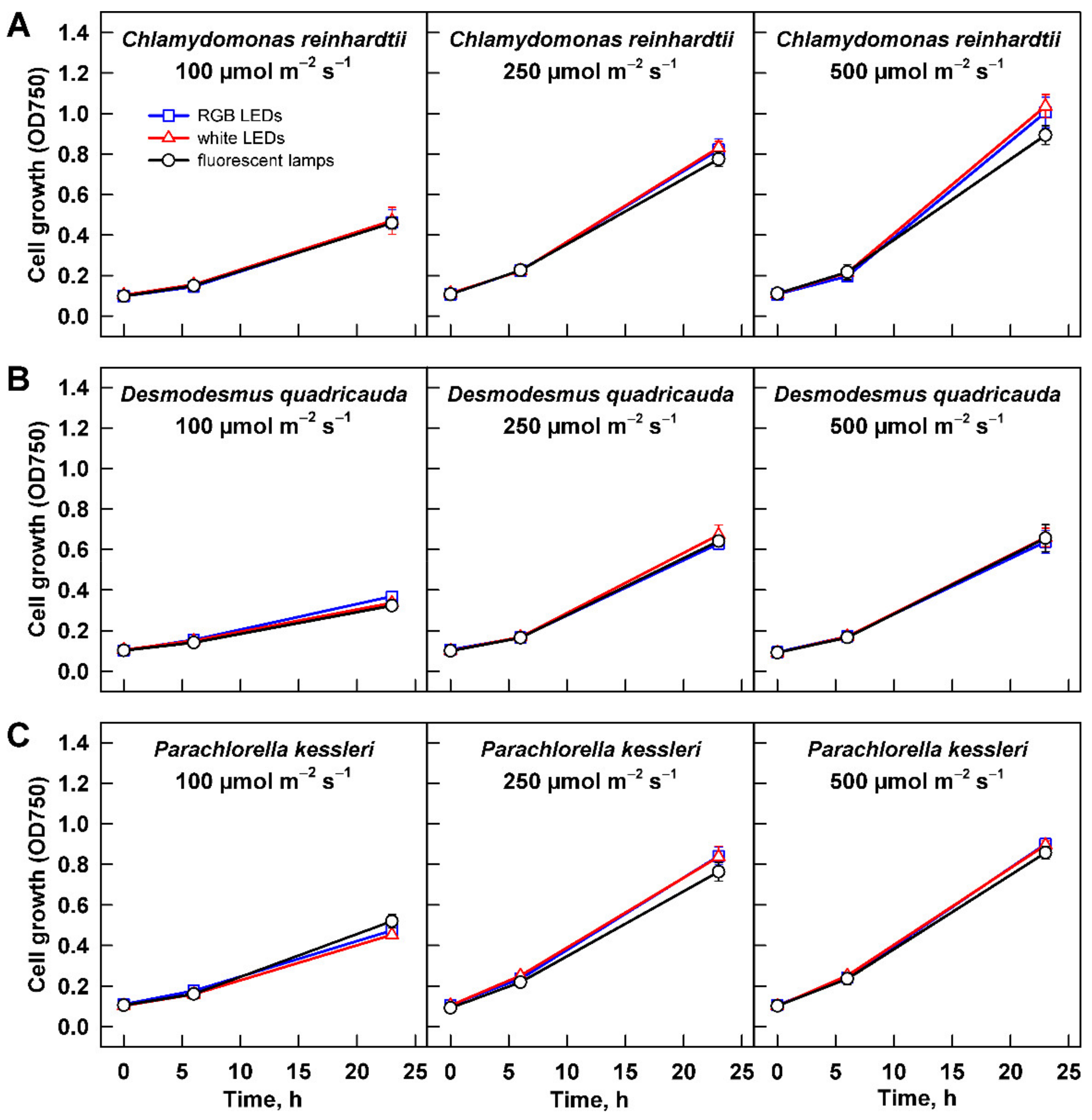
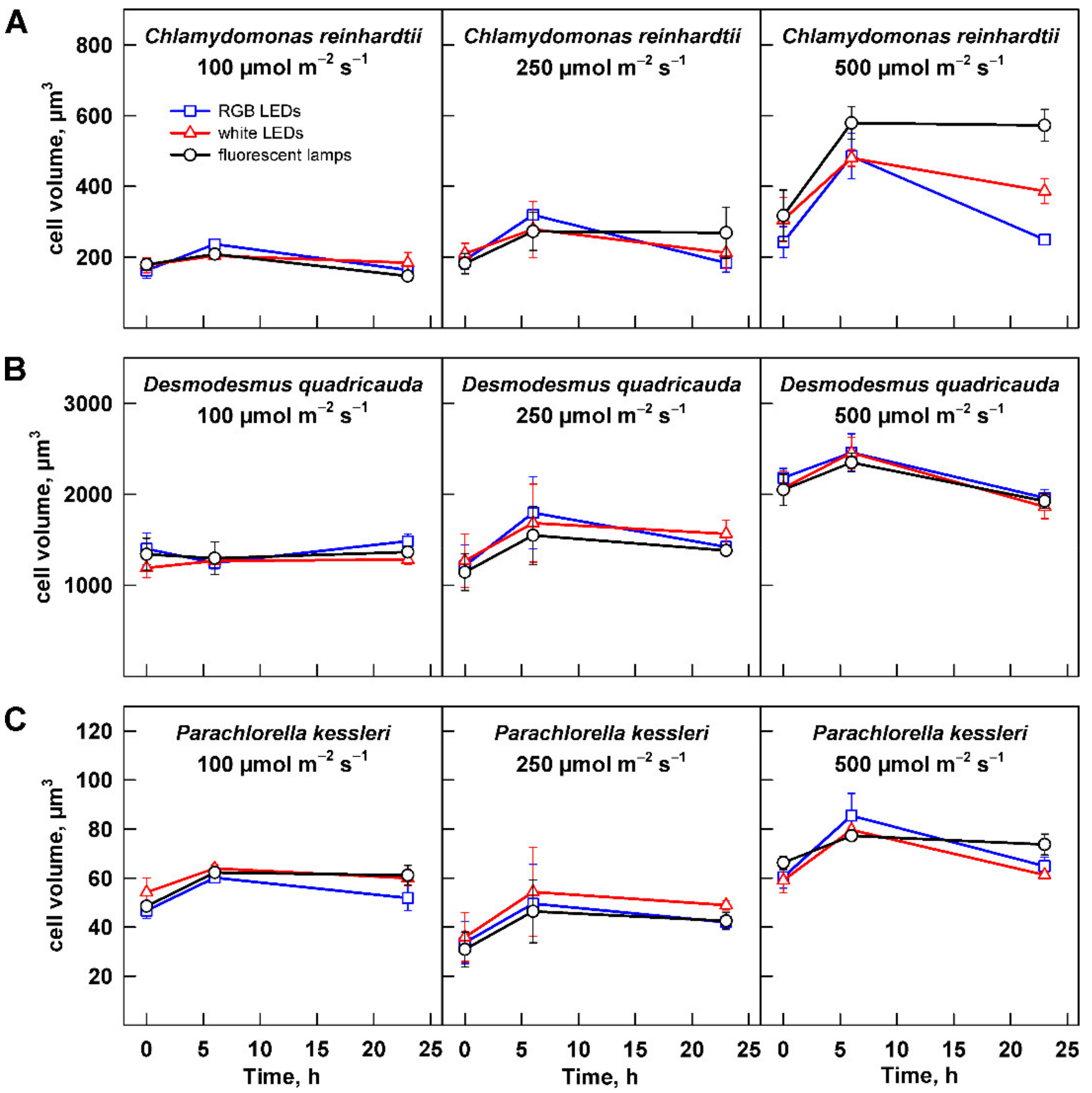
3.2. Effect of Light Intensity
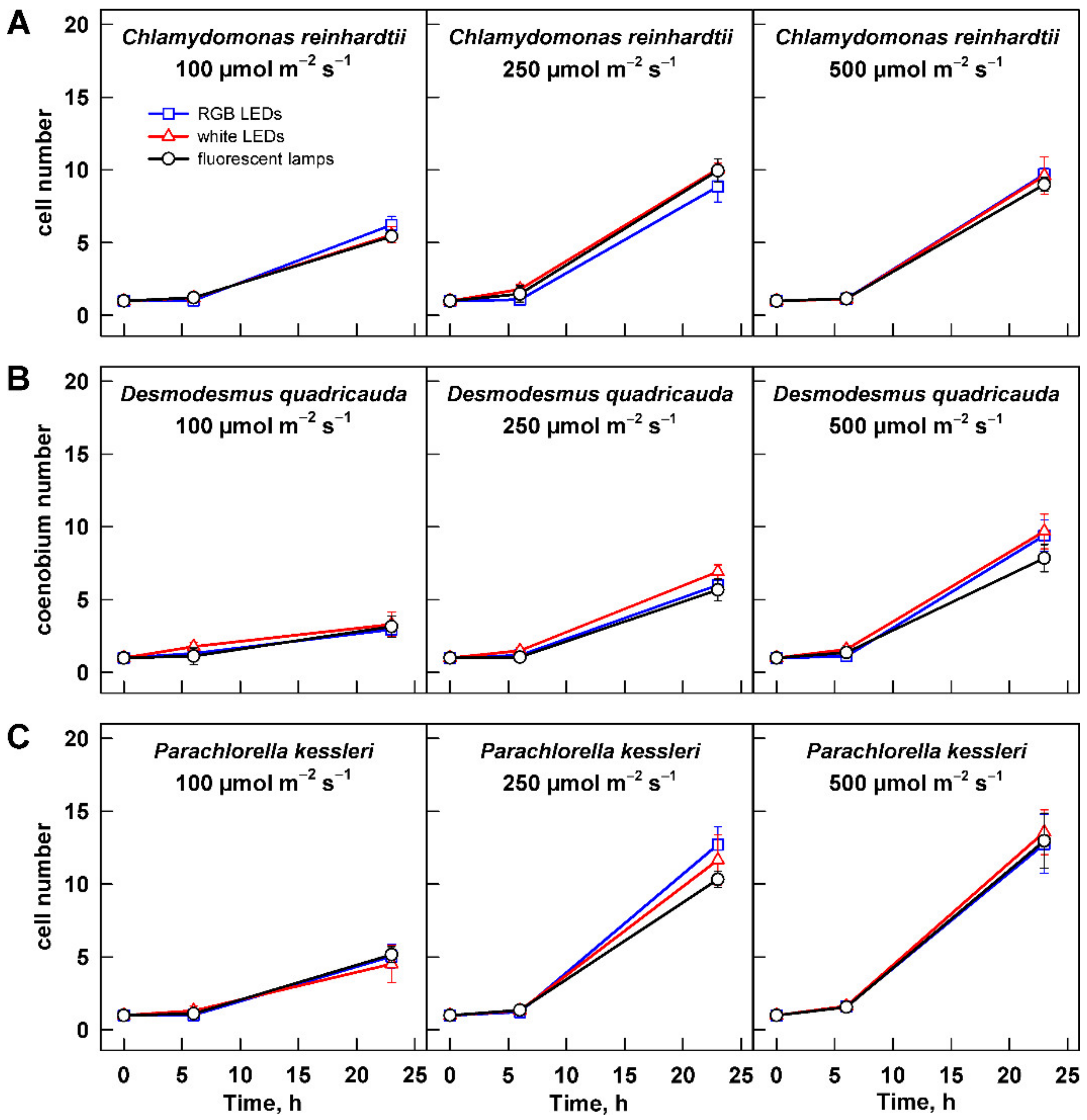
3.2.1. Cell Growth
3.2.2. Cell Size and Number
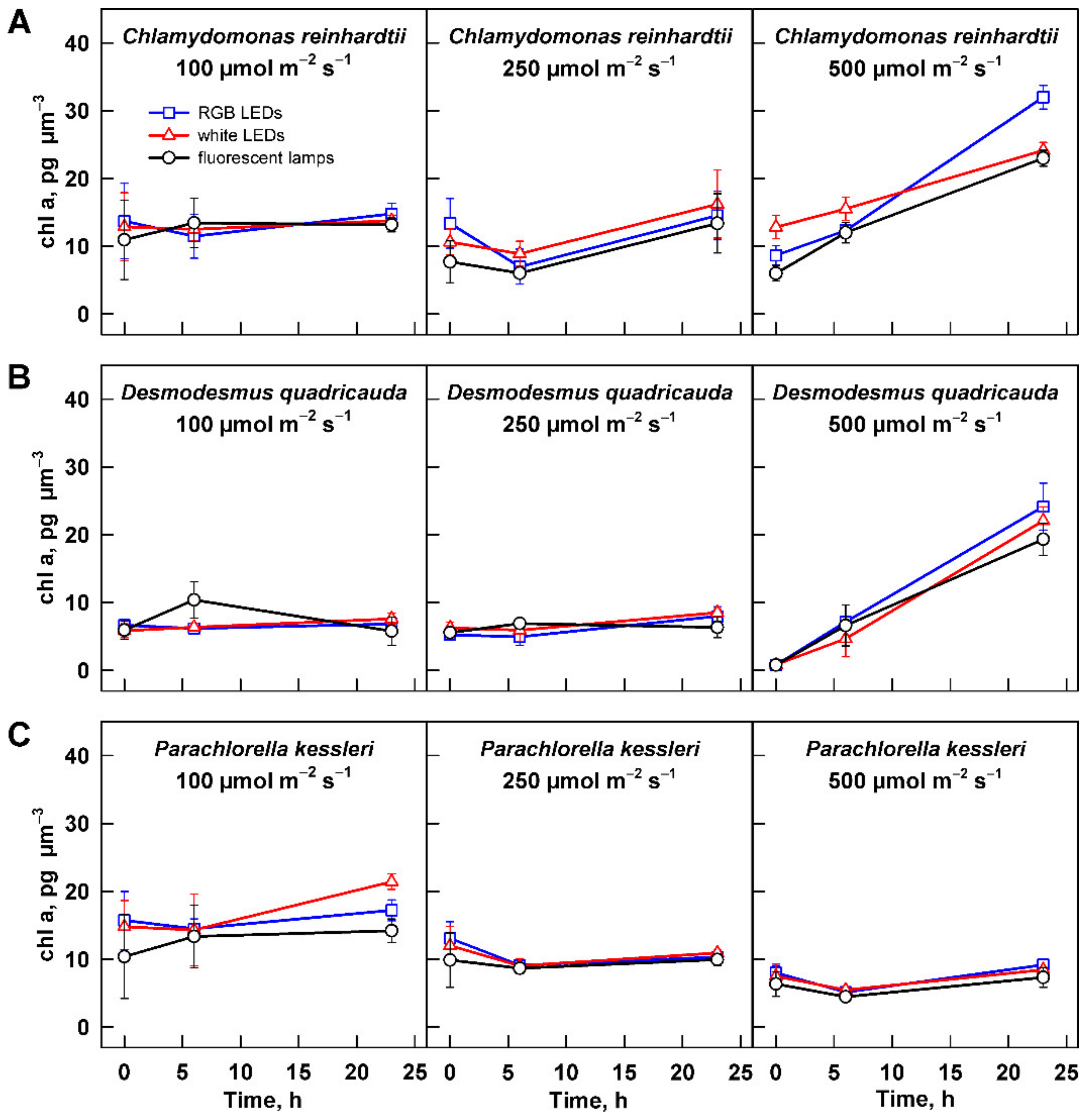
3.2.3. Photosynthetic Pigments
3.3. Effect of Light Quality
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mata, T.M.; Martins, A.A.; Caetano, N.S. Microalgae for biodiesel production and other applications: A review. Renew. Sustain. Energy Rev. 2010, 14, 217–232. [Google Scholar] [CrossRef] [Green Version]
- Spolaore, P.; Joannis-Cassan, C.; Duran, E.; Isambert, A. Commercial applications of microalgae. J. Biosci. Bioeng. 2006, 101, 87–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borowitzka, M.A. High-value products from microalgae—their development and commercialisation. J. Appl. Phycol. 2013, 25, 743–756. [Google Scholar] [CrossRef]
- Caporgno, M.P.; Mathys, A. Trends in microalgae incorporation into innovative food products with potential health benefits. Front. Nutr. 2018, 5, 58. [Google Scholar] [CrossRef] [PubMed]
- Chisti, Y. Biodiesel from microalgae beats bioethanol. Trends Biotechnol. 2008, 26, 126–131. [Google Scholar] [CrossRef]
- Hachicha, R.; Elleuch, F.; Ben Hlima, H.; Dubessay, P.; de Baynast, H.; Delattre, C.; Pierre, G.; Hachicha, R.; Abdelkafi, S.; Michaud, P. Biomolecules from microalgae and cyanobacteria: Applications and market survey. Appl. Sci. 2022, 12, 1924. [Google Scholar] [CrossRef]
- Metsoviti, M.N.; Papapolymerou, G.; Karapanagiotidis, I.T.; Katsoulas, N. Effect of light intensity and quality on growth rate and composition of Chlorella vulgaris. Plants 2019, 9, 31. [Google Scholar] [CrossRef] [Green Version]
- Patel, A.K.; Choi, Y.Y.; Sim, S.J. Emerging prospects of mixotrophic microalgae: Way forward to sustainable bioprocess for environmental remediation and cost-effective biofuels. Bioresour. Technol. 2020, 300, 122741. [Google Scholar] [CrossRef]
- Zachleder, V.; Kselíková, V.; Ivanov, I.N.; Bialevich, V.; Vítová, M.; Ota, S.; Takeshita, T.; Kawano, S.; Bišová, K. Supra-optimal temperature: An efficient approach for overaccumulation of starch in the green alga Parachlorella kessleri. Cells 2021, 10, 1806. [Google Scholar] [CrossRef]
- Narala, R.R.; Garg, S.; Sharma, K.K.; Thomas-Hall, S.R.; Deme, M.; Li, Y.; Schenk, P.M. Comparison of microalgae cultivation in photobioreactor, open raceway pond, and a two-stage hybrid system. Front. Energy Res. 2016, 4, 29. [Google Scholar] [CrossRef]
- Valdovinos-García, E.M.; Petriz-Prieto, M.A.; Olán-Acosta, M.d.l.Á.; Barajas-Fernández, J.; Guzmán-López, A.; Bravo-Sánchez, M.G. Production of microalgal biomass in photobioreactors as feedstock for bioenergy and other uses: A techno-economic study of harvesting stage. Appl. Sci. 2021, 11, 4386. [Google Scholar] [CrossRef]
- Paul, S.; Bera, S.; Dasgupta, R.; Mondal, S.; Roy, S. Review on the recent structural advances in open and closed systems for carbon capture through algae. Energy Nexus 2021, 4, 100032. [Google Scholar] [CrossRef]
- Singh, S.P.; Singh, P. Effect of temperature and light on the growth of algae species: A review. Renew. Sustain. Energy Rev. 2015, 50, 431–444. [Google Scholar] [CrossRef]
- Difusa, A.; Talukdar, J.; Kalita, M.C.; Mohanty, K.; Goud, V.V. Effect of light intensity and pH condition on the growth, biomass and lipid content of microalgae Scenedesmus species. Biofuels 2015, 6, 37–44. [Google Scholar] [CrossRef]
- Lee, E.; Jalalizadeh, M.; Zhang, Q. Growth kinetic models for microalgae cultivation: A review. Algal Res. 2015, 12, 497–512. [Google Scholar] [CrossRef]
- Xu, Y.; Ibrahim, I.M.; Harvey, P.J. The influence of photoperiod and light intensity on the growth and photosynthesis of Dunaliella salina (Chlorophyta) CCAP 19/30. Plant Physiol. Biochem. 2016, 106, 305–315. [Google Scholar] [CrossRef] [Green Version]
- Atta, M.; Idris, A.; Bukhari, A.; Wahidin, S. Intensity of blue LED light: A potential stimulus for biomass and lipid content in fresh water microalgae Chlorella vulgaris. Bioresour. Technol. 2013, 148, 373–378. [Google Scholar] [CrossRef]
- Vélez-Landa, L.; Hernández-De León, H.R.; Pérez-Luna, Y.D.C.; Velázquez-Trujillo, S.; Moreira-Acosta, J.; Berrones-Hernández, R.; Sánchez-Roque, Y. Influence of light intensity and photoperiod on the photoautotrophic growth and lipid content of the microalgae Verrucodesmus verrucosus in a photobioreactor. Sustainability 2021, 13, 6606. [Google Scholar] [CrossRef]
- Zhou, P.; Guo, H.; Fang, Z.; He, Y.; Weerasinghe, R. Effect of light quality on the cultivation of Chlorella pyrenoidosa. E3S Web Conf. 2020, 143, 02033. [Google Scholar] [CrossRef]
- Darko, E.; Heydarizadeh, P.; Schoefs, B.; Sabzalian, M.R. Photosynthesis under artificial light: The shift in primary and secondary metabolism. Philos. Trans. R. Soc. B 2014, 369, 20130243. [Google Scholar] [CrossRef]
- Bula, R.J.; Morrow, R.C.; Tibbitts, T.; Barta, D.; Ignatius, R.; Martin, T. Light-emitting diodes as a radiation source for plants. HortScience 1991, 26, 203–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulze, P.S.; Barreira, L.A.; Pereira, H.G.; Perales, J.A.; Varela, J.C. Light emitting diodes (LEDs) applied to microalgal production. Trends Biotechnol. 2014, 32, 422–430. [Google Scholar] [CrossRef] [PubMed]
- Glemser, M.; Heining, M.; Schmidt, J.; Becker, A.; Garbe, D.; Buchholz, R.; Bruck, T. Application of light-emitting diodes (LEDs) in cultivation of phototrophic microalgae: Current state and perspectives. Appl. Microbiol. Biotechnol. 2016, 100, 1077–1088. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Jin, P.; Cheng, J.J. A comprehensive comparable study of the physiological properties of four microalgal species under different light wavelength conditions. Planta 2018, 248, 489–498. [Google Scholar] [CrossRef] [PubMed]
- Shu, C.H.; Tsai, C.C.; Liao, W.H.; Chen, K.Y.; Huang, H.C. Effects of light quality on the accumulation of oil in a mixed culture of Chlorella sp. and Saccharomyces cerevisiae. J. Chem. Technol. Biotechnol. 2012, 87, 601–607. [Google Scholar] [CrossRef]
- Oldenhof, H.; Zachleder, V.; van den Ende, H. Blue- and red-light regulation of the cell cycle in Chlamydomonas reinhardtii (Chlorophyta). Eur. J. Phycol. 2006, 41, 313–320. [Google Scholar] [CrossRef] [Green Version]
- Koc, C.; Anderson, G.A.; Kommareddy, A. Use of red and blue light-emitting diodes (LED) and fluorescent lamps to grow microalgae in a photobioreactor. Isr. J. Aquac.-Bamidgeh 2013, 65, 797–804. [Google Scholar] [CrossRef]
- Sueoka, N. Mitotic replication of deoxyribonucleic acid in Chlamydomonas reinhardtii. Proc. Natl. Acad. Sci. USA 1960, 46, 83–91. [Google Scholar] [CrossRef] [Green Version]
- Zachleder, V.; Ivanov, I.; Vítová, M.; Bišová, K. Cell cycle arrest by supraoptimal temperature in the alga Chlamydomonas reinhardtii. Cells 2019, 8, 1237–1257. [Google Scholar] [CrossRef] [Green Version]
- Zachleder, V.; Šetlík, I. Effect of irradiance on the course of RNA synthesis in the cell cycle of Scenedesmus quadricauda. Biol. Plant. 1982, 24, 341–353. [Google Scholar] [CrossRef]
- Korzyńska, A.; Zychowicz, M. A method of estimation of the cell doubling time on basis of the cell culture monitoring data. Biocybern. Biomed. Eng. 2008, 28, 75–82. [Google Scholar]
- Řezanka, T.; Kaineder, K.; Mezricky, D.; Řezanka, M.; Bišová, K.; Zachleder, V.; Vítová, M. The effect of lanthanides on photosynthesis, growth, and chlorophyll profile of the green alga Desmodesmus quadricauda. Photosynth. Res. 2016, 130, 335–346. [Google Scholar] [CrossRef] [PubMed]
- MacKinney, G. Absorption of light by chlorophyll solutions. J. Biol. Chem. 1941, 140, 315–322. [Google Scholar] [CrossRef]
- Lichtenthaler, D.K.; Wellburn, A.R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 603, 591–592. [Google Scholar] [CrossRef] [Green Version]
- Lukavský, J.; Simmer, J.; Kubín, Š. Methods for algal growth evaluation. In Algal Essays and Monitoring Eutrophication; Marvan, P., Přibil, S., Lhotský, O., Eds.; E. Schweitzerbartsche Verlags-buchhandlung: Stuttgart, Germany, 1979; pp. 77–85. [Google Scholar]
- McCree, K.J. The action spectrum, absorptance and quantum yield of photosynthesis in crop plants. J. Agric. Meteorol. 1971, 9, 191–216. [Google Scholar] [CrossRef]
- Zachleder, V.; Bišová, K.; Vítová, M. The cell cycle of microalgae. In The Physiology of Microalgae; Borowitzka, M.A., Beardall, J., Raven, J.A., Eds.; Springer: Dordrecht, The Netherlands, 2016; Volume 6, pp. 3–46. [Google Scholar]
- Li, S.F.; Fanesi, A.; Martin, T.; Lopes, F. Biomass production and physiology of Chlorella vulgaris during the early stages of immobilized state are affected by light intensity and inoculum cell density. Algal Res. 2021, 59, 452–461. [Google Scholar] [CrossRef]
- He, Q.; Yang, H.; Wu, L.; Hu, C. Effect of light intensity on physiological changes, carbon allocation and neutral lipid accumulation in oleaginous microalgae. Bioresour. Technol. 2015, 191, 219–228. [Google Scholar] [CrossRef]
- Beale, S.I.; Appleman, D. Chlorophyll synthesis in Chlorella. J. Plant Physiol. 1971, 47, 230–235. [Google Scholar] [CrossRef] [Green Version]
- Da Silva Ferreira, V.; Sant’Anna, C. Impact of culture conditions on the chlorophyll content of microalgae for biotechnological applications. World J. Microbiol. Biotechnol. 2017, 33, 20. [Google Scholar] [CrossRef]
- Bonente, G.; Pippa, S.; Castellano, S.; Bassi, R.; Ballottari, M. Acclimation of Chlamydomonas reinhardtii to different growth irradiances. J. Biol. Chem. 2012, 287, 5833–5847. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, V.S.; Pinto, R.F.; Sant’Anna, C. Low light intensity and nitrogen starvation modulate the chlorophyll content of Scenedesmus dimorphus. J. Appl. Microbiol. 2016, 120, 661–670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guidi, L.; Tattini, M.; Landi, M. How does chloroplast protect chlorophyll against excessive light. In Chlorophyll; Jacob-Lopes, L.Q.Z.E., Queiroz, M.I., Eds.; IntechOpen: London, UK, 2017; Volume 21. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Incident Light Intensity (µmol m−2 s−1) | |||
|---|---|---|---|
| Organism | 100 | 250 | 500 |
| Fluorescent Lamps | |||
| Chlamydomonas reinhardtii | 4.68 ± 0.36 | 7.18 ± 0.28 | 8.12 ± 0.83 |
| Desmodesmus quadricauda | 3.19 ± 0.28 | 6.4 ± 0.1 | 7.24 ± 0.22 |
| Parachlorella kessleri | 4.96 ± 0.29 | 8.26 ± 0.46 | 8.35 ± 0.17 |
| RGB LED | |||
| Chlamydomonas reinhardtii | 4.79 ± 0.86 | 7.69 ± 0.69 | 9.29 ± 0.37 |
| Desmodesmus quadricauda | 3.63 ± 0.28 | 5.94 ± 0.39 | 6.94 ± 0.52 |
| Parachlorella kessleri | 4.3 ± 0.23 | 7.96 ± 0.65 | 8.52 ± 0.28 |
| White LED | |||
| Chlamydomonas reinhardtii | 4.52 ± 0.57 | 7.37 ± 0.41 | 9.51 ± 0.92 |
| Desmodesmus quadricauda | 3.21 ± 0.29 | 6.65 ± 0.28 | 6.68 ± 0.52 |
| Parachlorella kessleri | 4.4 ± 0.18 | 8.1 ± 0.8 | 8.83 ± 0.17 |
| Incident Light Intensity (µmol m−2 s−1) | |||
|---|---|---|---|
| Organism | 100 | 250 | 500 |
| Fluorescent lamps | |||
| Chlamydomonas reinhardtii | 208 ± 12 | 323 ± 21 | 579 ± 46 |
| Desmodesmus quadricauda | 1297 ± 180 | 1964 ± 169 | 2349 ± 1032 |
| Parachlorella kessleri | 62 ± 1 | 46 ± 4 | 77 ± 2 |
| RGB LED | |||
| Chlamydomonas reinhardtii | 236 ± 11 | 355 ± 14 | 485 ± 64 |
| Desmodesmus quadricauda | 1244 ± 67 | 2218 ± 132 | 2459 ± 204 |
| Parachlorella kessleri | 60 ± 2 | 50 ± 2 | 85 ± 9 |
| White LED | |||
| Chlamydomonas reinhardtii | 215 ± 11 | 393 ± 32 | 613 ± 24 |
| Desmodesmus quadricauda | 1267 ± 32 | 2209 ± 141 | 2456 ± 172 |
| Parachlorella kessleri | 64 ± 1 | 54 ± 5 | 80 ± 1 |
| Incident Light Intensity (µmol m−2 s−1) | |||
|---|---|---|---|
| Organism | 100 | 250 | 500 |
| Fluorescent lamps | |||
| Chlamydomonas reinhardtii | 9.44 ± 0.16 | 6.98 ± 0.25 | 7.28 ± 0.18 |
| Desmodesmus quadricauda | 12.68 ± 0.92 | 9.43 ± 0.83 | 7.85 ± 0.49 |
| Parachlorella kessleri | 9.89 ± 0.73 | 6.84 ± 0.16 | 6.33 ± 0.41 |
| RGB LED | |||
| Chlamydomonas reinhardtii | 8.82 ± 0.5 | 7.4 ± 0.37 | 7.03 ± 0.13 |
| Desmodesmus quadricauda | 12.36 ± 0.47 | 8.57 ± 0.12 | 7.19 ± 0.34 |
| Parachlorella kessleri | 9.27 ± 0.29 | 6.31 ± 0.26 | 6.39 ± 0.44 |
| White LED | |||
| Chlamydomonas reinhardtii | 9.43 ± 0.56 | 6.91 ± 0.11 | 7.17 ± 0.49 |
| Desmodesmus quadricauda | 12.61 ± 0.55 | 8.28 ± 0.28 | 7.12 ± 0.44 |
| Parachlorella kessleri | 9.11 ± 0.41 | 6.62 ± 0.47 | 6.16 ± 0.25 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bialevich, V.; Zachleder, V.; Bišová, K. The Effect of Variable Light Source and Light Intensity on the Growth of Three Algal Species. Cells 2022, 11, 1293. https://doi.org/10.3390/cells11081293
Bialevich V, Zachleder V, Bišová K. The Effect of Variable Light Source and Light Intensity on the Growth of Three Algal Species. Cells. 2022; 11(8):1293. https://doi.org/10.3390/cells11081293
Chicago/Turabian StyleBialevich, Vitali, Vilém Zachleder, and Kateřina Bišová. 2022. "The Effect of Variable Light Source and Light Intensity on the Growth of Three Algal Species" Cells 11, no. 8: 1293. https://doi.org/10.3390/cells11081293
APA StyleBialevich, V., Zachleder, V., & Bišová, K. (2022). The Effect of Variable Light Source and Light Intensity on the Growth of Three Algal Species. Cells, 11(8), 1293. https://doi.org/10.3390/cells11081293