Abstract
Cognitive deficits strongly affect the quality of life of patients with multiple sclerosis (MS). However, no cognitive MS biomarkers are currently available. Extracellular vesicles (EVs) contain markers of parental cells and are able to pass from the brain into blood, representing a source of disease biomarkers. The aim of this study was to investigate whether small non-coding microRNAs (miRNAs) targeting synaptic genes and packaged in plasma EVs may reflect cognitive deficits in MS patients. Total EVs were precipitated by Exoquick from the plasma of twenty-six cognitively preserved (CP) and twenty-three cognitively impaired (CI) MS patients belonging to two independent cohorts. Myeloid EVs were extracted by affinity capture from total EVs using Isolectin B4 (IB4). Fourteen miRNAs targeting synaptic genes were selected and measured by RT-PCR in both total and myeloid EVs. Myeloid EVs from CI patients expressed higher levels of miR-150-5p and lower levels of let-7b-5p compared to CP patients. Stratification for progressive MS (PMS) and relapsing-remitting MS (RRMS) and correlation with clinical parameters suggested that these alterations might be attributable to cognitive deficits rather than disease progression. This study identifies miR-150-5p and let-7b-5p packaged in blood myeloid EVs as possible biomarkers for cognitive deficits in MS.
1. Introduction
Although most work on multiple sclerosis (MS) has been addressing the role of immune cell activation on oligodendrocytes and myelin damage, current knowledge indicates that MS harbors several characteristic aspects of a neurodegenerative disorder, that is, damage to axons, synapses and nerve cell bodies [1,2]. Specifically, synaptic loss and axonal degeneration are early events in the disease [3,4] and could occur independently of demyelination.
Between 40% and 70% of patients suffer from MS-related cognitive impairment, which has been more strongly associated with the neurodegenerative aspects of the disease [5]. Cognitive impairment is present in all MS subtypes and is frequently seen early in the disease course [6,7], representing a predictor of future disease progression [8,9,10]. The cognitive domains most frequently impaired in MS are information-processing speed, working memory [8] and long-term memory [11], followed by dysfunction in executive functioning and attention [12,13]. Mood disturbances and personality abnormalities often occur in MS subjects too, and may also strongly affect patients’ quality of life [9].
Whereas functional deficits in the central nervous system (CNS) are routinely evaluated with the administration of the Expanded Disability Status Scale (EDSS) [14], assessment of cognitive functions by neuropsychological tests is time-consuming, and cognition cannot currently be monitored in standard clinical practice. From this point of view, a biomarker for cognitive deficit would be of great value. Many studies have tried to characterize the structural MRI correlates of cognitive impairment in patients with MS, finding an association between isolated measures of structural CNS damage and cognitive performance [15]. However, isolated MRI measurements, such as thalamic volume loss, are only moderately related to cognitive performance [16,17,18]. Thus, the development of more powerful/complementary cognitive MS biomarkers is still an unmet clinical need.
A full understanding of the molecular mechanisms underlying synaptic changes and altered connectivity in MS could be useful for identifying cognitive biomarkers. Recent evidence has linked synaptic alterations and cognitive deficits in MS to activation of microglia, the immune cells of the brain. Specifically, it was first demonstrated that microglia activation is necessary for hippocampal synaptic and cognitive alterations in experimental autoimmune encephalomyelitis (EAE) [19], an animal model for MS. Activation of NLRP3 inflammasome in microglia was then shown to play an important role in EAE cognitive deficits via the alteration of astrocyte phenotypes and the release of complement component 3 (C3) [20]. Consistently, higher complement deposits were detected in post-mortem hippocampi of MS patients with cognitive impairments, compared to cognitive preserved patients [21]. Other studies linked microgliosis to increased production of microglial extracellular vesicles (EVs) in MS patients and revealed that microglial EVs impair stability of excitatory synapses [22,23].
EVs recently entered the scene of neuroscience as important mediators of cell-to-cell communication, which transfer a variety of biomolecules, including genetic materials, between cells. EVs can pass from the brain into blood and circulate in body fluids, where they are emerging as promising markers for several neurological diseases, representing a brain “fluid biopsy” [24,25]. Among EV cargo which influence excitatory synapses, our recent data point to small non-coding microRNAs (miRNAs), which have validated synaptic targets [23]. By silencing synaptic proteins, these miRNAs, which are delivered to neurons by EVs produced from reactive microglia and astrocytes, impair synapse stability and function in the mouse brain, and may be responsible for defects in synaptic plasticity and cognition in MS patients. In this study, we tested the hypothesis that miRNAs targeting synaptic proteins in myeloid EVs could represent cognitive biomarkers in MS.
2. Materials and Methods
2.1. Patients Enrolled and Study Design
Patients with MS were included in the study with the following eligibility criteria: (i) diagnosis of MS according to the 2010 McDonald criteria [26], (ii) EDSS score ≤ 7, (iii) age ≥ 25 and ≤70 years, (iv) no steroid treatment for at least four weeks before the blood withdrawal and (v) ability to provide written informed consent. Details of demographic characteristics are shown in Table 1. A first Italian MS cohort from IRCCS Fondazione Ca’ Granda Ospedale Maggiore Policlinico, consisting of 21 patients, was initially enrolled. Patients were grouped as cognitively preserved (CP) and cognitively impaired (CI) according to their neuropsychological status assessed with the Rao Brief Repeatable Battery (BRB) [27] and were characterized for the neurofilament serum levels (sNf-L). Subsequently, a validation Amsterdam MS Center cohort from Amsterdam UMC, location VUmc, was enrolled, consisting of 28 patients. A test battery based on the MACFIMS [28] was administered to the second cohort, as outlined below.

Table 1.
Demographic and clinical features of the two cohorts of patients enrolled.
IRCCS Fondazione Ca’ Granda cohort. Eleven CP (all RRMS type) and 10 CI (six RRMS and four PMS) MS patients were recruited at the Neurology Unit of the Fondazione IRCCS Ca’ Granda Ospedale Maggiore Policlinico (Milan, Italy) (Supplementary Table S1). Cognitive functions were assessed by an experienced neuropsychologist with the use of the Italian version of the BRB [27]. It includes tests of: (1) attention and speed in information processing (paced auditory serial addition test (PASAT 3 and PASAT 2) and Symbol Digit Modality Test (SDMT)), (2) verbal memory (selective reminding test, long-term storage (SRT LTS), consistent long-term retrieval (SRT CLTR), and delayed (STR D)), and (3) spatial memory (spatial recall test immediate and delayed (SPART and SPART D)). Consistent with previous works [27], those patients who failed at least three tests were considered cognitively impaired, and those who failed less than three tests were considered cognitively preserved.
MS Center Amsterdam cohort. Fifteen CP (fourteen RRMS and one PMS) and thirteen CI (nine RRMS and four PMS) MS patients were recruited at Amsterdam UMC (Amsterdam, the Netherlands) (Supplementary Table S1). Cognitive functions were assessed by a test battery based on the MACFIMS [28], and consisted of the following five (sub-)domains: (1) verbal memory (Dutch version of the California Verbal Learning Test version 2), (2) visuospatial memory (Brief Visuospatial Memory Test–Revised), (3) processing speed (SDMT and Stroop Color–Word Test cards I and II), (4) executive function—verbal fluency (Controlled Oral Word Association Test) and (5) executive function—response inhibition (Stroop Color–Word Test interference score). Scores were corrected for age, education and sex when applicable, and transformed into five (sub-)domain-specific z-scores, as well as one composite score for overall cognitive functioning (i.e., average of all z-scores) based on a normative sample of healthy controls. Patients were classified as CI (i.e., ≥1.5 standard deviations (SDs) below the means of healthy controls on ≥20% of the neuropsychological test scores, corresponding to ≥3/11 test scores) or CP (i.e., remainder) [29]. Additional data included disease duration in years, disease type at the time of the visit and EDSS score.
2.2. Serum Nf-L Determination
Nf-L levels were determined in serum obtained from 7 mL of blood withdrawal after centrifugation for 10 min at 1500× g. sNf-L levels were measured by using an MSD (R-PLEX Human Neurofilament L Kit, according to the manufacturer’s instructions, Meso Scale Discovery, Rockville, MD, USA) in the Italian cohort. Analysis was performed using a QuickPlex SQ 120 instrument (MSD, Rockville, MD, USA) and DISCOVERY WORKBENCH®4.0 software. sNf-L levels of the Amsterdam MS Center cohort were measured using a single-molecule array (Simoa) assay to measure the Nf-L levels on an HD-X instrument (Quanterix, Billerica, MA, USA) following the manufacturer’s instructions.
2.3. Total EVs and Myeloid EV Isolation
For total and myeloid EVs’ isolation, 7–10 mL of venous blood was drawn by syringe into 0.5 mL of saline with EDTA, centrifuged for 15 min at 1500× g, aliquoted and stored at −80 °C pending use.
Aliquots of 500 μL of plasma were incubated with 5 μL of Thrombin (Thrombin Plasma Prep 611 U/mL, System biosciences, SBI, Palo Alto, CA, USA) for 5 min in order to eliminate the large amount of fibrin present in plasma. After centrifugation for 5 min at 3000× g at RT, total EVs were harvested from the supernatants by precipitation with 126 µL of Exoquick exosome precipitation solution (EXOQ; System biosciences, SBI, Palo Alto, CA, USA) per 500 μL of supernatants and, after incubation for 30 min at 4 °C, centrifuged for 30 min at RT. Each pellet containing EVs was resuspended in 500 μL of phosphate buffer. To enrich in myeloid EVs, 100 µL aliquots of the total EV suspensions were incubated with 40 μL of streptavidin magnetic beads (Exo-Flow streptavidin 9.1 μm beads, System biosciences, SBI, Palo Alto, CA, USA), preincubated with 10 μL of biotinylated Isolectin B4 (IB4; 1 mg/mL; Sigma Aldrich, St. Louis, MO, USA) at 4 °C with continuous mixing overnight. Then, IB4-positive EVs were isolated using a magnetic plate and treated for subsequent RNA extraction and Western blot analysis. Biotinylated IB4 pretreated with the IB4 interacting molecule Melibiose (6-O-a-d-galactopyranosyl-d-glucose, 1M) [22] was used as a negative control for Western blot analysis. For size distribution analysis, myeloid EVs were resuspended in 300 μL of Exosome Elution Buffer (Exo-Flow, System biosciences, SBI, Palo Alto, CA, USA) and incubated on a rotating rank for 40 min at RT to detach IB4-positive EVs from bead–antibody complexes. Final suspensions containing EVs were stored at −80 °C for further analysis.
2.4. Analysis of EV Size and Concentration
The size distribution and concentration of total and myeloid EVs were measured by the Tunable Resistive Pulse Sensing (TRPS) technique, using the qNano instrument (Izon, Christchurch, New Zealand). Then, 0.1 µm of filtered PBS supplemented with 0.3% Wetting Solution (from Izon EV reagent kit) was used for both pre-treating the pore and suspending EVs in order to prevent EVs binding to the pore or spontaneous EV aggregation. Total and myeloid-enriched EV samples were analyzed using a NP200 nanopore (Izon, Christchurch, New Zealand). CPC200 calibration particles (carboxylated polystyrene particles, supplied by Izon and diluted following the manufacturer’s instructions) were used as standards, and they were measured immediately before or after the experimental samples under identical conditions. Data acquisition and analysis were performed using Izon Control Suite software (version V3.2, Izon, Christchurch, New Zealand).
2.5. Western Blotting
The protein concentration of total EVs and IB4-positive EVs isolated with Exo-Flow 2.0 streptavidin beads (System biosciences, SBI, Palo Alto, CA, USA) was quantified by the Micro BCA Protein Assay Kit (Thermo Fisher Scientific, Waltham, MA, USA) to load equal amounts of total EVs and IB4-positive EVs. The negative control (EVs captured by beads coated with melibiose-treated IB4) was immune-captured from an equal volume of total EVs compared to IB4-positive EVs. EVs were dissolved in PBS with LDS sample buffer (Life Technologies, Waltham, MA, USA) and separated using 12% SDS polyacrylamide gels. Then, samples were electro-transferred onto nitrocellulose membranes and blot was incubated overnight at 4 °C with specific primary antibodies: anti-AnnexinA2 (ANXA2) (diluted 1:500; Abcam, Cambridge, MA, USA), anti-TMEM119 (diluted 1:1000; BioLegend, San Diego, CA, USA) and anti-CD171 (diluted 1:1000; Thermo Fisher Scientific, Waltham, MA, USA). After washing, membranes were incubated with horseradish peroxidase-conjugated secondary antibodies (Pierce, Thermo Fisher Scientific, Waltham, MA, USA) for 1 h and immuno-positive bands were detected by enhanced chemiluminescence (ECL, Euroclone, Milan, Italy, or FEMTO, Thermo Fisher, Waltham, MA, USA) according to the manufacturer’s instructions. Densitometric analysis of immunoreactive bands was performed using ImageJ software (www.imagej.nih.gov/ij/, accessed on 1 July 2021).
2.6. Selection of miRNAs with Synaptic Targets
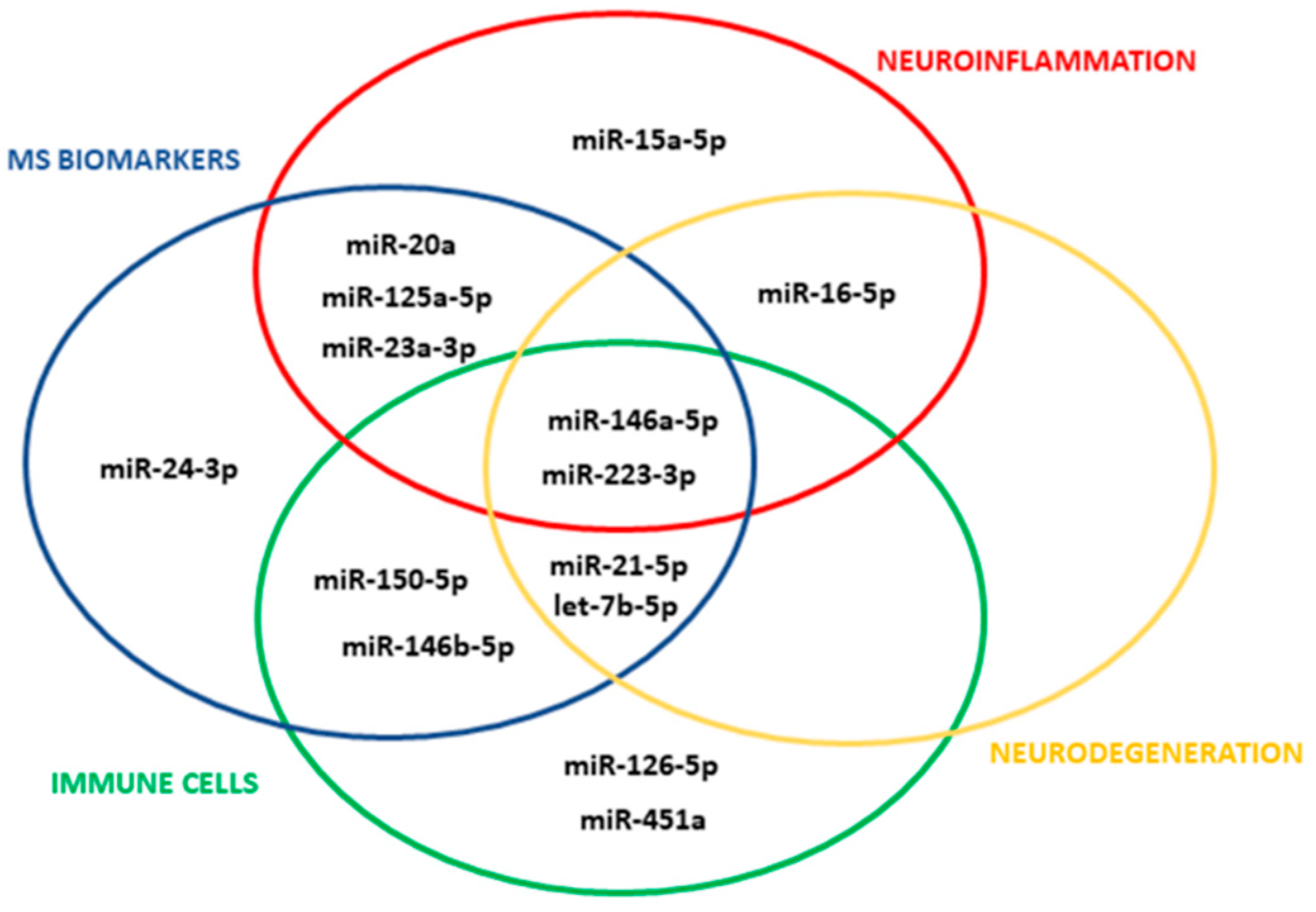
Only a fraction of miRNAs are sorted from cells into EVs [23,30] and are detectable in EVs circulating in human blood [31,32]. In addition, the limited volume of plasma available from biobanks (about 500 µL) prevents multiple analyses on the same patient. Therefore, to increase the chance to identify vesicular miRNAs reflecting/predicting cognitive symptoms in MS patients, we restricted the analysis to a small group of miRNAs with validated synaptic targets in humans. We focused on miRNAs highly expressed in human circulating EVs (top 10 miRNAs enriched in plasma EVs) [32], and/or consistently detected in plasma EVs from healthy subjects/Alzheimer’s disease patients (n = 58) [31] (Supplementary Table S2). According to the microRNA database, miRTarBase (http://miRTarBase.mbc.nctu.edu.tw/, accessed on 1 July 2021), and Panther classification (http://pantherdb.org/news/news20201218.jsp, accessed on 1 July 2021), the vast majority of these miRNAs have validated synaptic targets, as listed in Supplementary Table S2. To narrow their number, the miRNAs were further selected using the following criteria: enrichment in microglia/peripheral immune cells and EVs thereof [33,34,35,36,37,38,39,40,41,42,43], release under inflammatory or neurodegenerative stimuli [23], previous identification as candidate biomarkers for MS pathology [44,45,46,47,48,49,50,51,52,53,54,55] and dysregulation in neurodegenerative diseases [31,56,57,58]. The miRNAs that passed the selection criteria were the following: miR-146a-5p, miR-223-3p, miR-16-5p, miR-23a-3p, miR-20a, miR-21-5p, miR-125a-5p, miR-146b-5p, miR-150-5p, miR-24-3p, miR-15a-5p, let-7b-5p, miR-451a and miR-126-5p (Table 2). Experimentally validated synaptic targets with strong evidence in humans (by report assays, qPCR and Western blot analysis) of candidate miRNAs are listed in Supplementary Table S3. In addition, cel-miR-39, a C. elegans miRNA, not expressed in human cells, was selected as an exogenous control for RNA extraction and the retro-transcription process.
Table 2.
Characteristics of selected miRNAs targeting synaptic genes.
2.7. Total RNA Isolation and Quantitative Real-Time PCR Analysis
RNA was extracted from total and IB4-positive EVs using the Total Exosome RNA Isolation Kit (Invitrogen, Waltham, MA, USA) following the manufacturer’s protocol. The reverse transcription (RT) reaction was performed using the MicroRNA Reverse Transcription Kit (Applied Biosystem, Waltham, MA, USA). Given the low RNA amount in our samples, cDNA was pre-amplified using TaqMan®Pre-Amp Master Mix (Applied Biosystem, Waltham, MA, USA). Each well of a 96-well plate was loaded with 5 μL of pre-amplified cDNA (diluted 1:5) and 15 μL of PCR Reaction Mix, composed of TaqMan® Universal PCR Master Mix, TaqMan® miRNA Assay and RNase- free water (Applied Biosystem, Waltham, MA, USA). The reaction and amplification protocol was performed according to the manufacturers’ instructions. Finally, the quantitative RT-PCR was run on a QuantStudio™ 12K Flex Real-Time PCR System (Thermo Fischer Scientific, Waltham, MA, USA). Raw data underwent quality control analysis, by applying a Crt (relative threshold cycle) cutoff < 28 and the amplification score > 1 (measure of good amplification quality). Candidate miRNAs were detectable in total EVs from most MS patients, except for miR-146a-5p and miR-23a-3p, whose expression levels did not reach good quality control parameters and were excluded from the analysis. In most IB4-positive EVs, miR-15a-5p and miR-125a-5p were undetectable. As an endogenous control reference, miR-16-5p and miR-21-5p were used for the Italian population, while miR-16-5p and miR-451a for the Amsterdam MS Center one, identified through the algorithm GeNorm. The relative quantification (RQ) was calculated with the 2−ΔΔCt method [59]. Patients’ classification in terms of cognitive performance was revealed upon completion of the PCR analysis.
2.8. Statistical Analysis
Principal component analysis (PCA) for both cohorts was performed using ClustVis (https://biit.cs.ut.ee/clustvis/, accessed on 16 April 2022). Unit variance row scaling was applied for PCA. Statistical analysis was performed using Prism GraphPad 9.0 software (San Diego, CA, USA). Data distribution was tested for normality with the Kolmogorov–Smirnov and Shapiro–Wilk tests. All data are shown as mean ± SEM. Differences between two groups were analyzed with the Mann−Whitney test. Multiple comparisons were performed by one-way ANOVA followed by Tukey’s test, or the Kruskal–Wallis test followed by Dunn’s Multiple Comparison test as post hoc tests. For EV miRNA enrichment analysis, the Wilcoxon matched-pairs signed-rank test was used. Differences were considered significant when p < 0.05 and p < 0.01, and they are indicated by one or two asterisks, respectively. The Spearman test for correlation between miR-150-5p/let-7b-5p and biochemical, demographic, clinical variables and neuropsychological test scores was also used.
3. Results
We selected 14 miRNAs with validated synaptic targets in humans (Figure 1; for selection criteria see Section 2) and probed these miRNAs for both total and myeloid EVs isolated from plasma of CP and CI MS patients belonging to 2 independent cohorts. Total EVs were isolated from human plasma with the Exoquick commercial kit, and myeloid EVs were extracted from total EVs by immune-affinity capture using the myeloid marker IB4.
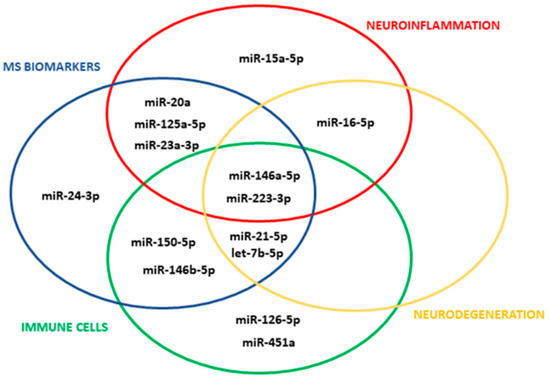
Figure 1.
miRNA selection. The Venn diagram shows the 14 circulating miRNAs with validated synaptic targets which meet the selection criteria.
3.1. Isolation of Total and Myeloid EVs from Blood Plasma
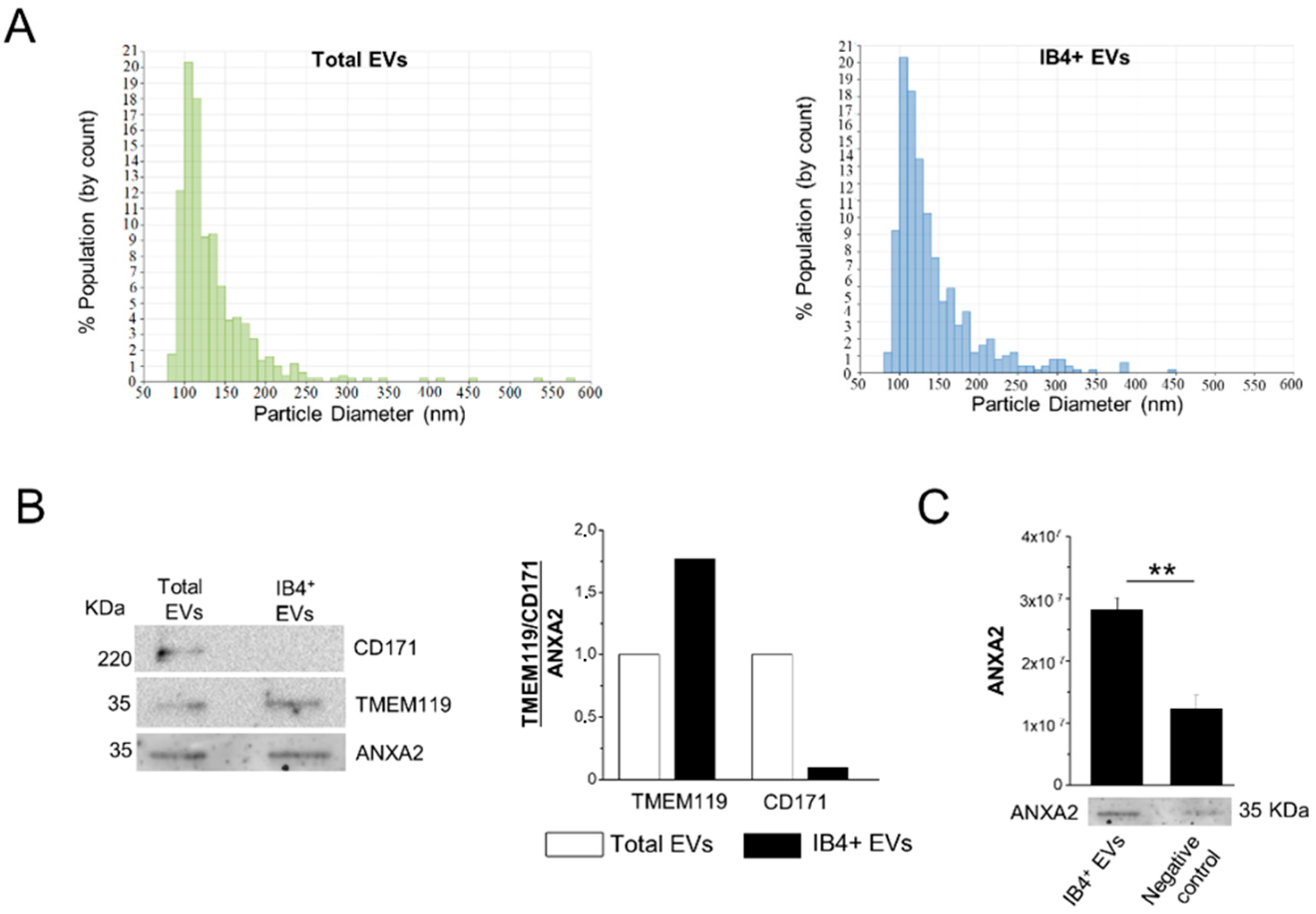
According to MISEV 2018 guidelines [60], we previously characterized, via multiple approaches, i.e., electron microscopy, flow cytometry, Nanoparticle Tracking Analysis (NTA) and Western blotting, human plasma EVs isolated by Exoquick, and a subpopulation affinity-captured from total plasma EVs, finding pure populations of mainly oval-shaped particles, approximately 150 nm in diameter [31]. The presence of miRNAs packaged inside plasma EVs was also previously assessed using an RNase protection assay [31]. In this study, total EVs precipitated with Exoquick from frozen MS plasma samples were further analyzed by TRPS (number of MS patients = 4). The analysis showed that an average of 3.49 × 1011 ± 2.62 × 1011 EVs can be extracted from 1 mL of plasma in the Exoquick pellet, and confirmed that EVs were relatively small in size, ranging from 84 to 661 nm (mean diameter = 140 nm, mode of the diameter = 110 nm; Figure 2A, left). TRPS analysis showed that myeloid EVs, isolated from total EVs by affinity capture with IB4, were similar in size to total EVs (mean diameter = 144.83 nm, mode of the diameter = 111.83 nm; Figure 2A, right), and were about 1% of the total EVs.
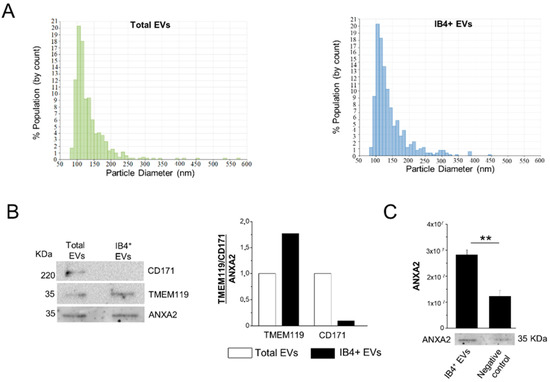
Figure 2.
Characterization of total and IB4-positive EVs. (A) Representative size distribution of total and IB4-positive (IB4+) EVs detected by TRPS. (B) Representative Western blot and relative quantification of microglial TMEM119 marker and neuronal CD171 marker in total EVs and IB4+ EVs. CD171 and TMEM119 expression are normalized to ANXA2 staining. (C) Expression and quantification of EV marker ANXA2 in IB4+ EVs and negative control (IB4-coated beads pretreated with melibiose) (n = 3). Data are shown as mean ± SEM. ** p < 0.01, one-way ANOVA, Tukey’s multiple comparisons test.
3.2. Assessment of Myeloid EV Enrichment
The specificity of the affinity method to extract myeloid EVs from the total EV population was assessed by Western blotting on total EVs and IB4-positive EVs isolated from a plasma pool of three subjects. Staining for the EV marker ANXA2, the microglia marker TMEM119 and the neural marker CD171 showed immunoreactivity of total EVs for both microglial and neuronal markers (Figure 2B). As expected, densitometric analysis revealed that IB4-positive EVs were enriched in TMEM119 (by 1.75-fold) compared to total EVs, while they were almost negative (depleted to 0.09-fold) for the neuronal marker CD171 (Figure 2B). Less EVs were isolated upon pretreatment of IB4-coated beads with melibiose [22], which inhibits IB4 binding to myeloid cell-specific glucose residues, as evidenced by decreased ANXA2 immunoreactivity (Figure 2C).
3.3. Expression of Selected miRNAs in Total and Myeloid EVs
We initially investigated a population consisting of 11 CP MS patients and 10 CI MS patients assessed by a short version of BRB, a series of tests previously used for cognitive evaluation in MS [61].
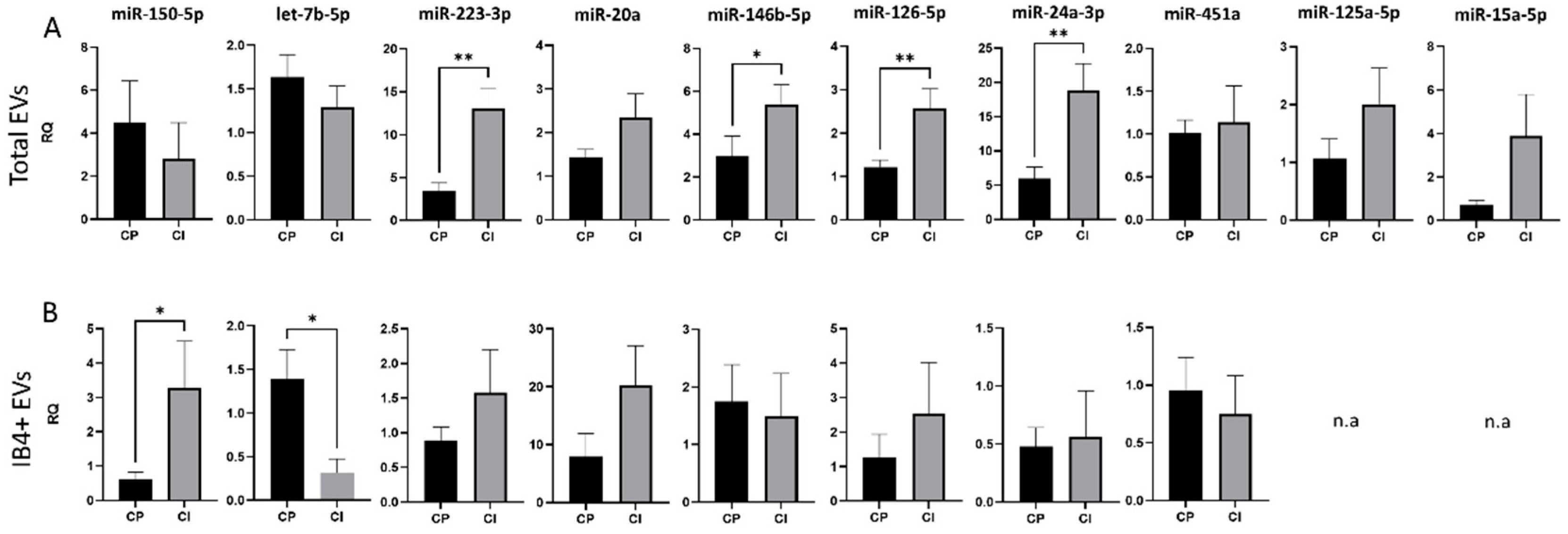
Results of the study cohort characteristics are provided in Table 1. We found an upregulation of miR-24-3p, miR-126-5p, miR-223-3p and miR-146b-5p in total EVs from CI compared to CP patients (RQ ± SEM: 18.82 ± 3.93 vs. 5.93 ± 1.70, p = 0.003; 2.70 ± 0.44 vs. 1.22 ± 0.16, p = 0.005; 13.01 ± 2.45 vs. 3.50 ± 0.89, p = 0.001; 5.41 ± 0.89 vs. 2.99 ± 0.92, p = 0.04, respectively, Figure 3A), while in IB4-positive EVs, miR-150-5p was upregulated and let-7b-5p was downregulated in patients with cognitive disabilities (RQ ± SEM: 3.29 ± 0.11 vs. 0.60 ± 0.2, p = 0.03; 0.323 ± 0.15 vs. 1.39 ± 0.33, p = 0.03, respectively, Figure 3B).
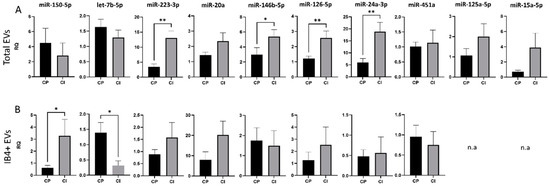
Figure 3.
miRNA cargo in total and IB4-positive (IB4+) EVs in the Italian cohort of MS patients. (A) RQ of selected miRNA levels in total plasma EVs from CP and CI patients. (B) RQ of selected miRNA levels in IB4+ EVs from CP and CI patients. Data were normalized to miR-16-5p and miR-21-5p, expressed as 2–ΔΔCt. Mann–Whitney test, * p < 0.05, ** p < 0.001.
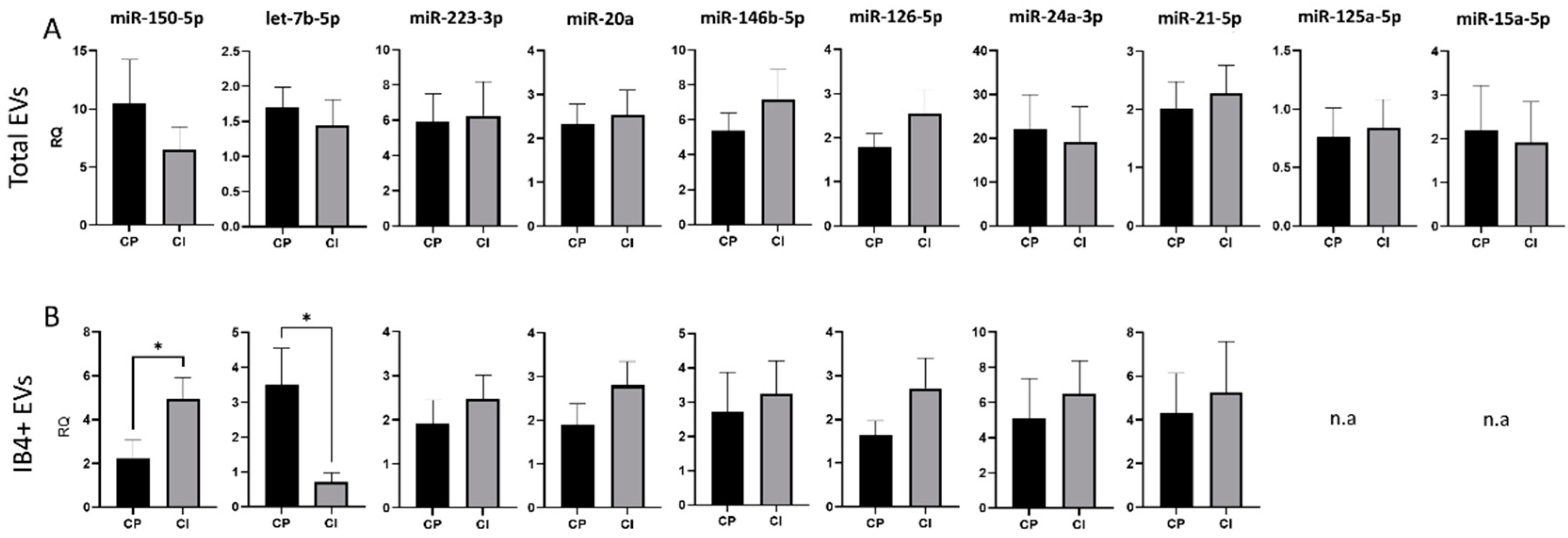
To validate these data, the expression of the 14 miRNAs was analyzed in a second cohort consisting of 15 CP MS patients and 13 CI MS patients, who underwent a larger battery of neuropsychological tests (7 tests) to deeply measure the most affected cognitive domains in MS. No significant changes in the content of any miRNAs with validated synaptic targets were detected between CI and CP patients in total EVs (Figure 4A). Conversely, analysis of the miRNA cargo confirmed the upregulation of miR-150-5p, a miRNA involved in the development of myeloid lineages [62] and in MS inflammation, and the downregulation of let-7b-5p, a miRNA controlling immune cell responses [63], in IB4-positive EVs from CI patients compared to CP patients (RQ ± SEM: 4.93 ± 0.99 vs. 2.22 ± 0.84, p = 0.03; 0.71 ± 0.27 vs. 3.52 ± 1.03, p < 0.05, Figure 4B). To exclude the possibility that miR-150-5p and let7b were randomly selected as statistically different, we performed PCA for myeloid EVs in both cohorts. PCA showed that miRNA measurements in myeloid EVs could distinguish patients based on their cognitive status (Supplementary Figure S1).
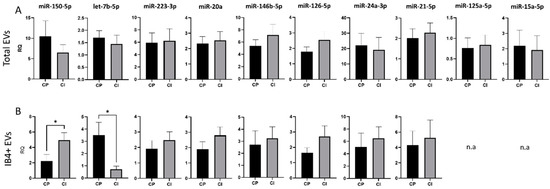
Figure 4.
miRNA cargo in total and IB4-positive (IB4+) EVs in the Amsterdam MS Center cohort of MS patients. (A) RQ of selected miRNA levels in total plasma EVs from CP and CI patients. (B) RQ of selected miRNA levels in IB4+ EVs from CP and CI patients. Data were normalized to miR-16-5p and miR-451a, expressed as 2–ΔΔCt. Mann–Whitney test, * p < 0.05.
As expected for the two miRNAs highly expressed in myeloid cells [33,34,35,36,39], analysis of let-7b-5p and miR-150-5p expression in IB4-positive EVs compared to total EVs revealed an enrichment for both miRNAs in the IB4-positive fraction, albeit the results reached the significant threshold only for miR-150-5p (p < 0.0001; Supplementary Figure S2). This evidence confirmed the enrichment of myeloid EVs in the IB4-positive fraction. Moreover, stratification of let-7b-5p and miR-150-5p levels according to disease type showed no significant differences between PMS and RRMS in both of the cohorts considered (Supplementary Figure S3), suggesting that the miRNA dysregulation might be attributable to cognitive deficits rather than MS progression.
3.4. Correlations between miRNA Levels and Clinical Data
Finally, we performed correlation analysis between miR-150-5p and let-7b-5p expression levels in IB4-positive EVs and clinical features relevant to MS course (age at blood withdrawal, age at disease onset (AAO), gender, EDSS, disease duration), both in the total group and stratified for cognitive status. Correlations between miRNA levels and the axonal damage biomarker Nf-L in the serum were also carried out. miR-150-5p/let-7b-5p expression in myeloid EVs was not correlated with the clinical data, either when patients were grouped or stratified according to their cognitive status (Supplementary Table S4). Conversely, a moderate positive correlation between miR-150-5p levels in IB4-positive EVs and sNf-L was observed both in the overall Italian MS population (r = 0.46, p = 0.04) and in the Amsterdam MS Center one (r = 0.47, p = 0.04).
4. Discussion
By measuring a small set of miRNAs, which may cause synaptic dysfunction, in plasma EVs from two independent small cohorts of CP and CI MS patients, we found a specific signature of cognitive dysfunction, consisting of high levels of miR-150-5p and low levels of let-7b-5p in IB4-positive EVs, derived from microglia and other phagocytes. No significant changes of these miRNAs were detected between CP and CI MS patients in total plasma EVs, which originate from several cell types, indicating that myeloid EVs are a better source for miRNAs reflecting synaptic alterations in MS compared to total plasma EVs and, likely, circulating miRNAs. Although the battery of cognitive tests used was different, both batteries in the two patient cohorts assessed the cognitive domains most frequently impaired in MS.
Overall, our findings suggest that miRNA dysregulation might be attributable to cognitive deficits rather than MS progression. In fact, no differences in the miRNA levels were observed in IB4-positive EVs from PMS compared to RRMS patients, and no direct correlation was found between miRNAs levels in CI patients and sNf-L serum concentration, a proposed predictor of poor clinical outcome in MS [64], whose role in tracking cognitive deficit in MS is still controversial [65,66,67,68]. However, higher miR150-5p levels directly correlated with sNf-L levels in the overall Italian cohort as well as in the Amsterdam MS Center cohort, implying a role of the miRNA in neurodegenerative process. sNf-L were evaluated in the two populations with two different platforms, whose results have already been demonstrated to be highly correlated [69].
A major limitation of our study is the small sample size. Thus, data should be interpreted with caution. This holds true, especially for PMS patients, whose limited number does not allow us to accurately investigate miRNA expression changes between PMS and RRMS. Another apparent limitation is the heterogeneity of the two cohorts, which differ in terms of disease duration and EDSS. However, we would like to point out that the ability of miR150-5p and let-7b-5p to capture cognitive performance in such heterogeneous MS populations is in favor of their value as cognitive biomarkers in all MS forms.
miR-150-5p is a miRNA enriched in both brain-resident microglia [23,33] and peripheral myeloid cells [34,36], which targets many neuronal genes controlling synaptic stability and functions. Among miR-150-5p targets, ephrin type B receptor2 (EPHB2), Gβ5, a member of the signal-transducing G protein β, and Wnt7b regulate dendrite formation, dendritic spines’ maturation and/or stimulate the formation of excitatory synapses [70], while another target, PAK3, mediates cytoskeletal reorganization in dendritic spines during synaptic plasticity [71]. Importantly, defects in PAK3 as well as in Patched domain-containing protein 1 (PTCHD1), another validated miR150-5p target, are associated with intellectual disability in humans [72,73], and PTCHD1 deficiency causes synaptic and cognitive dysfunctions in mice [74]. This suggests that EVs mediate the transfer of miR-150-5p to neurons, and consequent PAK3/PTCHD1 downregulation may be implicated in cognitive impairments of MS patients. Immunohistochemistry of brain tissue from CP and CI patients will be necessary to test this intriguing hypothesis in future studies.
Let-7b-5p targets several relevant synaptic genes, involved in synaptic transmission and cognition, including synaptotagmin1, a fundamental regulator of calcium-dependent exocytosis, Fragile X-Related Protein2 (FXR2), a RNA-binding protein involved in dendritic maturation [75], and neuroplastin (NPTN), a synaptic-enriched adhesion molecule that acts as a key regulator of synaptic plasticity and function [76]. However, let-7b-5p also silences the expression of multiple inflammatory genes in myeloid cells, including type 1 interferon (IFNB1), interleukin 8 (CXCL8) and signal transducer and activator of transcription 2 (STAT2), thereby strongly influencing the immune cell function. Members of the let-7 family were previously shown to be downregulated in microglia exposed to neurodegenerative stimulus [23], and their reduction might be instrumental in bringing about maladaptive myeloid cell activation in neurodegeneration. According to this hypothesis, here, we showed a significant decrease in the expression of let-7b-5p in myeloid EVs from CI compared to CP MS patients, which may reflect the transcriptional and functional signature of dysfunctional microglia in neurodegenerative conditions, termed disease-associated microglia (DAM) [77] or neurodegenerative microglia [78]. Our data are in perfect agreement with a recent study showing that let-7b-5p levels in the cerebrospinal fluid (CSF) directly correlated with a better cognitive performance of RRMS patients, while they were reduced in progressive MS [49]. The latter finding, along with the present study, suggests that let-7b-5p content in myeloid but not total blood EVs might reflect the miRNA levels in the CSF and help to define the activation state of myeloid cells in the course of the disease.
To conclude, our data suggested that let-7b-5p and miR-150-5p profiling in myeloid EVs may represent a possible useful diagnostic tool to measure cognitive dysfunction in MS, which provides complementary information to MRI indication (e.g., thalamic volume loss). Further research is needed, however, to validate the expression changes of the synaptic and/or myeloid targets controlled by miR-150-5p and let-7b-5p, and to see how well the miRNAs perform as biomarkers of cognitive impairment, also in relation to other proposed biomarkers for MS cognitive impairment, such as CSF Tau [79].
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/cells11091551/s1, Table S1: Gender distribution among CI and CP RRMS and PMS patients in the two cohorts; Table S2: miRNAs present in plasma EVs and their validated key synaptic targets; Table S3: Selected miRNAs and their validated key synaptic targets; Table S4: Correlation analysis among clinical variables and miR150-5p and let7b-5p levels; Figure S1: Principal component analysis based on the selected miRNAs in IB4-positive EVs for the Italian and Amsterdam cohorts; Figure S2: miR-150-5p and let-7b-5p content in IB4-positive EVs compared to total EVs from the same patients; Figure S3: Relative quantification of miR-150-5p and let-7-5p in total EVs and IB4-positive EVs from RRMS vs. PMS patients.
Author Contributions
Conceptualization, C.V. (Claudia Verderio) and C.F.; formal analysis, F.S., C.V. (Caterina Visconte), M.S., M.T.G., M.G. and M.H.; investigation, H.E.H., T.C., M.D.R., A.P., E.R., M.v.D. and B.A.d.J.; writing—original draft preparation, C.V. (Caterina Visconte) and C.F.; writing—review and editing, F.S., H.E.H., C.E.T., D.G. and B.A.d.J.; supervision, E.S., D.G., C.E.T. and C.V. (Claudia Verderio); funding acquisition, C.V. (Caterina Visconte), M.G. and F.S. All authors have read and agreed to the published version of the manuscript.
Funding
The work has been supported by FOE (Fondo Ordinario Enti di ricerca) 2022 and FISM—Fondazione Italiana Sclerosi Multipla—cod. 2018/R/22 to Claudia Verderio and 2016/B/2 to M.G., and financed or co-financed with the ‘5 per mille’ public funding. F.S. was the recipient of a fellowship from Regione Lombardia “NeOn”, POR-FESR 2014–2020, ID 239047. M.S. is supported by MoH, grant GR-2019-12369100.
Institutional Review Board Statement
The study was approved by the Institutional Review Board (IRB) of the Fondazione IRCCS Ca’ Granda Ospedale Maggiore Policlinico (Milan, Italy) Parere 857-2021 CE, and the IRB of VU University medical center (VUmc) of Amsterdam, The Netherlands.
Informed Consent Statement
Informed consent was obtained from all patients involved in the study.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Acknowledgments
We thank Elga de Vries (MS Center Amsterdam, The Netherlands) for helpful discussion and suggestions.
Conflicts of Interest
C.E.T. has a collaboration contract with ADx Neurosciences, Quanterix and Eli Lilly, and performed contract research or received grants from AC-Immune, Axon Neurosciences, Biogen, Brainstorm Therapeutics, Celgene, EIP Pharma, Eisai, PeopleBio, Roche, Toyama and Vivoryon. She serves on editorial boards of Medidact Neurologie/Springer, Alzheimer Research and Therapy, Neurology: Neuroimmunology & Neuroinflammation, and is the editor of a Neuromethods book, Springer.
References
- Draheim, T.; Liessem, A.; Scheld, M.; Wilms, F.; Weißflog, M.; Denecke, B.; Kensler, T.W.; Zendedel, A.; Beyer, C.; Kipp, M.; et al. Activation of the astrocytic Nrf2/ARE system ameliorates the formation of demyelinating lesions in a multiple sclerosis animal model. Glia 2016, 64, 2219–2230. [Google Scholar] [CrossRef] [PubMed]
- Kipp, M.; Nyamoya, S.; Hochstrasser, T.; Amor, S. Multiple sclerosis animal models: A clinical and histopathological perspective. Brain Pathol. 2017, 27, 123–137. [Google Scholar] [CrossRef] [PubMed]
- Geurts, J.J.; Barkhof, F. Grey matter pathology in multiple sclerosis. Lancet Neurol. 2008, 7, 841–851. [Google Scholar] [CrossRef]
- Fong, J.S.; Rae-Grant, A.; Huang, D.R. Neurodegeneration and neuroprotective agents in multiple sclerosis. Recent Pat. CNS Drug Discov. 2008, 3, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Benedict, R.H.B.; Amato, M.P.; DeLuca, J.; Geurts, J.J.G. Cognitive impairment in multiple sclerosis: Clinical management, MRI, and therapeutic avenues. Lancet Neurol. 2020, 19, 860–871. [Google Scholar] [CrossRef]
- Glanz, B.I.; Holland, C.M.; Gauthier, S.A.; Amunwa, E.L.; Liptak, Z.; Houtchens, M.K.; Sperling, R.A.; Khoury, S.J.; Guttmann, C.R.G.; Weiner, H.L. Cognitive dysfunction in patients with clinically isolated syndromes or newly diagnosed multiple sclerosis. Mult. Scler. 2007, 13, 1004–1010. [Google Scholar] [CrossRef]
- Nocentini, U.; Pasqualetti, P.; Bonavita, S.; Buccafusca, M.; De Caro, M.F.; Farina, D.; Girlanda, P.; Le Pira, F.; Lugaresi, A.; Quattrone, A.; et al. Cognitive dysfunction in patients with relapsing-remitting multiple sclerosis. Mult. Scler. 2006, 12, 77–87. [Google Scholar] [CrossRef]
- Zipoli, V.; Goretti, B.; Hakiki, B.; Siracusa, G.; Sorbi, S.; Portaccio, E.; Amato, M.P. Cognitive impairment predicts conversion to multiple sclerosis in clinically isolated syndromes. Mult. Scler. 2010, 16, 62–67. [Google Scholar] [CrossRef]
- Reuter, F.; Zaaraoui, W.; Crespy, L.; Faivre, A.; Rico, A.; Malikova, I.; Confort-Gouny, S.; Cozzone, P.J.; Ranjeva, J.P.; Pelletier, J.; et al. Cognitive impairment at the onset of multiple sclerosis: Relationship to lesion location. Mult. Scler. J. 2011, 17, 755–758. [Google Scholar] [CrossRef]
- Pitteri, M.; Romualdi, C.; Magliozzi, R.; Monaco, S.; Calabrese, M. Cognitive impairment predicts disability progression and cortical thinning in MS: An 8-year study. Mult. Scler. 2017, 23, 848–854. [Google Scholar] [CrossRef]
- Lazeron, R.H.C.; Rombouts, S.A.R.B.; Scheltens, P.; Polman, C.H.; Barkhof, F. An fMRI study of planning-related brain activity in patients with moderately advanced multiple sclerosis. Mult. Scler. 2004, 10, 549–555. [Google Scholar] [CrossRef]
- McCarthy, M.L.; MacKenzie, E.J.; Durbin, D.R.; Aitken, M.E.; Jaffe, K.M.; Paidas, C.N.; Slomine, B.S.; Dorsch, A.M.; Berk, R.A.; Christensen, J.R.; et al. The Pediatric Quality of Life Inventory: An evaluation of its reliability and validity for children with traumatic brain injury. Arch. Phys. Med. Rehabil. 2005, 86, 1901–1909. [Google Scholar] [CrossRef]
- Langdon, D.W. Cognition in multiple sclerosis. Curr. Opin. Neurol. 2011, 24, 244–249. [Google Scholar] [CrossRef]
- Kurtzke, J.F. Rating neurologic impairment in multiple sclerosis: An expanded disability status scale (EDSS). Neurology 1983, 33, 1444–1452. [Google Scholar] [CrossRef] [Green Version]
- Preziosa, P.; Rocca, M.A.; Pagani, E.; Stromillo, M.L.; Enzinger, C.; Gallo, A.; Hulst, H.E.; Atzori, M.; Pareto, D.; Riccitelli, G.C.; et al. Structural MRI correlates of cognitive impairment in patients with multiple sclerosis. Hum. Brain Mapp. 2016, 37, 1627–1644. [Google Scholar] [CrossRef]
- Houtchens, M.K.; Benedict, R.H.B.; Killiany, R.; Sharma, J.; Jaisani, Z.; Singh, B.; Weinstock-Guttman, B.; Guttmann, C.R.G.; Bakshi, R. Thalamic atrophy and cognition in multiple sclerosis. Neurology 2007, 69, 1213–1223. [Google Scholar] [CrossRef]
- Schoonheim, M.M.; Popescu, V.; Lopes, F.C.R.; Wiebenga, O.T.; Vrenken, H.; Douw, L.; Polman, C.H.; Geurts, J.J.G.; Barkhof, F. Subcortical atrophy and cognition. Neurology 2012, 79, 1754–1761. [Google Scholar] [CrossRef]
- Bergsland, N.; Pelizzari, L.; Laganá, M.M.; Di Tella, S.; Rossetto, F.; Nemni, R.; Clerici, M.; Baglio, F. Automated Assessment of the Substantia Nigra Pars Compacta in Parkinson’s Disease: A Diffusion Tensor Imaging Study. J. Pers. Med. 2021, 11, 1235. [Google Scholar] [CrossRef]
- Di Filippo, M.; De Iure, A.; Giampà, C.; Chiasserini, D.; Tozzi, A.; Orvietani, P.L.; Ghiglieri, V.; Tantucci, M.; Durante, V.; Quiroga-Varela, A.; et al. Persistent activation of microglia and NADPH drive hippocampal dysfunction in experimental multiple sclerosis. Sci. Rep. 2016, 6, 20926. [Google Scholar] [CrossRef]
- Hou, B.; Zhang, Y.; Liang, P.; He, Y.; Peng, B.; Liu, W.; Han, S.; Yin, J.; He, X. Inhibition of the NLRP3-inflammasome prevents cognitive deficits in experimental autoimmune encephalomyelitis mice via the alteration of astrocyte phenotype. Cell Death Dis. 2020, 11, 377. [Google Scholar] [CrossRef]
- Ramaglia, V.; Dubey, M.; Malpede, M.A.; Petersen, N.; de Vries, S.I.; Ahmed, S.M.; Lee, D.S.; Schenk, G.J.; Gold, S.M.; Huitinga, I.; et al. Complement-associated loss of CA2 inhibitory synapses in the demyelinated hippocampus impairs memory. Acta Neuropathol. 2021, 142, 643–667. [Google Scholar] [CrossRef]
- Verderio, C.; Muzio, L.; Turola, E.; Bergami, A.; Novellino, L.; Ruffini, F.; Riganti, L.; Corradini, I.; Francolini, M.; Garzetti, L.; et al. Myeloid microvesicles are a marker and therapeutic target for neuroinflammation. Ann. Neurol. 2012, 72, 610–624. [Google Scholar] [CrossRef] [Green Version]
- Prada, I.; Gabrielli, M.; Turola, E.; Iorio, A.; D’Arrigo, G.; Parolisi, R.; De Luca, M.; Pacifici, M.; Bastoni, M.; Lombardi, M.; et al. Glia-to-neuron transfer of miRNAs via extracellular vesicles: A new mechanism underlying inflammation-induced synaptic alterations. Acta Neuropathol. 2018, 135, 529–550. [Google Scholar] [CrossRef] [Green Version]
- Vella, L.J.; Hill, A.F.; Cheng, L. Focus on extracellular vesicles: Exosomes and their role in protein trafficking and biomarker potential in Alzheimer’s and Parkinson’s disease. Int. J. Mol. Sci. 2016, 17, 173. [Google Scholar] [CrossRef]
- Selmaj, I.; Cichalewska, M.; Namiecinska, M.; Galazka, G.; Horzelski, W.; Selmaj, K.W.; Mycko, M.P. Global exosome transcriptome profiling reveals biomarkers for multiple sclerosis. Ann. Neurol. 2017, 81, 703–717. [Google Scholar] [CrossRef]
- Polman, C.H.; Reingold, S.C.; Banwell, B.; Clanet, M.; Cohen, J.A.; Filippi, M.; Fujihara, K.; Havrdova, E.; Hutchinson, M.; Kappos, L.; et al. Diagnostic criteria for multiple sclerosis: 2010 Revisions to the McDonald criteria. Ann. Neurol. 2011, 69, 292–302. [Google Scholar] [CrossRef] [Green Version]
- Amato, M.P.; Portaccio, E.; Goretti, B.; Zipoli, V.; Ricchiuti, L.; De Caro, M.F.; Patti, F.; Vecchio, R.; Sorbi, S.; Trojano, M. The Rao’s Brief Repeatable Battery and Stroop Test: Normative values with age, education and gender corrections in an Italian population. Mult. Scler. 2006, 12, 787–793. [Google Scholar] [CrossRef]
- Benedict, R.H.; Cookfair, D.; Gavett, R.; Gunther, M.; Munschauer, F.; Garg, N.; Weinstock-Guttman, B. Validity of the minimal assessment of cognitive function in multiple sclerosis (MACFIMS). J. Int. Neuropsychol. Soc. 2006, 12, 549–558. [Google Scholar] [CrossRef]
- Nauta, I.M.; Bertens, D.; van Dam, M.; Huiskamp, M.; Driessen, S.; Geurts, J.J.; Uitdehaag, B.M.; Fasotti, L.; Hulst, H.E.; de Jong, B.A.; et al. Performance validity in outpatients with multiple sclerosis and cognitive complaints. Mult. Scler. 2021, 28, 642–653. [Google Scholar] [CrossRef]
- Jeppesen, D.K.; Fenix, A.M.; Franklin, J.L.; Higginbotham, J.N.; Zhang, Q.; Zimmerman, L.J.; Liebler, D.C.; Ping, J.; Liu, Q.; Evans, R.; et al. Reassessment of Exosome Composition. Cell 2019, 177, 428–445. [Google Scholar] [CrossRef] [Green Version]
- Serpente, M.; Fenoglio, C.; D’Anca, M.; Arcaro, M.; Sorrentino, F.; Visconte, C.; Arighi, A.; Fumagalli, G.G.; Porretti, L.; Cattaneo, A.; et al. MiRNA Profiling in Plasma Neural-Derived Small Extracellular Vesicles from Patients with Alzheimer’s Disease. Cells 2020, 9, 1443. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Cheng, L.; Shao, Q.; Chen, Z.; Lv, X.; Li, J.; He, L.; Sun, Y.; Ji, Q.; Lu, P.; et al. Characterization of serum small extracellular vesicles and their small RNA contents across humans, rats, and mice. Sci. Rep. 2020, 10, 4197. [Google Scholar] [CrossRef] [PubMed]
- Jovičić, A.; Roshan, R.; Moisoi, N.; Pradervand, S.; Moser, R.; Pillai, B.; Luthi-Carter, R. Comprehensive expression analyses of neural cell-type-specific miRNAs identify new determinants of the specification and maintenance of neuronal phenotypes. Ann. Intern. Med. 2013, 158, 5127–5137. [Google Scholar] [CrossRef] [Green Version]
- Ma, Z.; Shen, Y.; Zeng, Q.; Liu, J.; Yang, L.; Fu, R.; Hu, G. MiR-150-5p regulates EGR2 to promote the development of chronic rhinosinusitis via the DC-Th axis. Int. Immunopharmacol. 2018, 54, 188–197. [Google Scholar] [CrossRef]
- Rong, J.; Xu, L.; Hu, Y.; Liu, F.; Yu, Y.; Guo, H.; Ni, X.; Huang, Y.; Zhao, L.; Wang, Z. Inhibition of let-7b-5p contributes to an anti-tumorigenic macrophage phenotype through the SOCS1/STAT pathway in prostate cancer. Cancer Cell Int 2020, 20, 470. [Google Scholar] [CrossRef]
- Selimoglu-Buet, D.; Rivière, J.; Ghamlouch, H.; Bencheikh, L.; Lacout, C.; Morabito, M.; Diop, M.; Meurice, G.; Breckler, M.; Chauveau, A.; et al. A miR-150/TET3 pathway regulates the generation of mouse and human non-classical monocyte subset. Nat. Commun. 2018, 9, 5455. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Zhou, Y.; Wang, J.; Yan, Y.; Peng, L.; Qiu, W. Dysregulated microRNA involvement in multiple sclerosis by induction of T helper 17 cell differentiation. Front. Immunol. 2018, 9, 1256. [Google Scholar] [CrossRef]
- Gaudet, A.D.; Fonken, L.K.; Watkins, L.R.; Nelson, R.J.; Popovich, P.G. MicroRNAs: Roles in Regulating Neuroinflammation. Neuroscientist 2018, 24, 221–245. [Google Scholar] [CrossRef] [Green Version]
- Marques-Rocha, J.L.; Garcia-Lacarte, M.; Samblas, M.; Bressan, J.; Martínez, J.A.; Milagro, F.I. Regulatory roles of miR-155 and let-7b on the expression of inflammation-related genes in THP-1 cells: Effects of fatty acids. J. Physiol. Biochem. 2018, 74, 579–589. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, M.; Zhong, M.; Suo, Q.; Lv, K. Expression profiles of miRNAs in polarized macrophages. Int. J. Mol. Med. 2013, 31, 797–802. [Google Scholar] [CrossRef] [Green Version]
- Hulsmans, M.; van Dooren, E.; Mathieu, C.; Holvoet, P. Decrease of miR-146b-5p in monocytes during obesity is associated with loss of the anti-inflammatory but not insulin signaling action of adiponectin. PLoS ONE 2012, 7, e32794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coleman, L.G.; Zou, J.; Crews, F.T. Microglial-derived miRNA let-7 and HMGB1 contribute to ethanol-induced neurotoxicity via TLR7. J. Neuroinflamm. 2017, 14, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Youn, Y.J.; Shrestha, S.; Lee, Y.B.; Kim, J.K.; Lee, J.H.; Hur, K.; Mali, N.M.; Nam, S.W.; Kim, S.H.; Lee, S.; et al. Neutrophil-derived trail is a proinflammatory subtype of neutrophil-derived extracellular vesicles. Theranostics 2021, 11, 2770–2787. [Google Scholar] [CrossRef] [PubMed]
- Amoruso, A.; Blonda, M.; Gironi, M.; Grasso, R.; Di Francescantonio, V.; Scaroni, F.; Furlan, R.; Verderio, C.; Avolio, C. Immune and central nervous system-related miRNAs expression profiling in monocytes of multiple sclerosis patients. Sci. Rep. 2020, 10, 6125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vistbakka, J.; Sumelahti, M.L.; Lehtimäki, T.; Elovaara, I.; Hagman, S. Evaluation of serum miR-191-5p, miR-24-3p, miR-128-3p, and miR-376c-3 in multiple sclerosis patients. Acta Neurol. Scand. 2018, 138, 130–136. [Google Scholar] [CrossRef] [PubMed]
- Lescher, J.; Paap, F.; Schultz, V.; Redenbach, L.; Scheidt, U.; Rosewich, H.; Nessler, S.; Fuchs, E.; Gärtner, J.; Brück, W.; et al. MicroRNA regulation in experimental autoimmune encephalomyelitis in mice and marmosets resembles regulation in human multiple sclerosis lesions. J. Neuroimmunol. 2012, 246, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Angelou, C.C.; Wells, A.C.; Vijayaraghavan, J.; Dougan, C.E.; Lawlor, R.; Iverson, E.; Lazarevic, V.; Kimura, M.Y.; Peyton, S.R.; Minter, L.M.; et al. Differentiation of Pathogenic Th17 Cells Is Negatively Regulated by Let-7 MicroRNAs in a Mouse Model of Multiple Sclerosis. Front. Immunol. 2020, 10, 3125. [Google Scholar] [CrossRef] [Green Version]
- Vistbakka, J.; Elovaara, I.; Lehtimäki, T.; Hagman, S. Circulating microRNAs as biomarkers in progressive multiple sclerosis. Mult. Scler. 2017, 23, 403–412. [Google Scholar] [CrossRef]
- Mandolesi, G.; Rizzo, F.R.; Balletta, S.; Stampanoni Bassi, M.; Gilio, L.; Guadalupi, L.; Nencini, M.; Moscatelli, A.; Ryan, C.P.; Licursi, V.; et al. The microRNA let-7b-5p Is Negatively Associated with Inflammation and Disease Severity in Multiple Sclerosis. Cells 2021, 10, 330. [Google Scholar] [CrossRef]
- Liguori, M.; Nuzziello, N.; Licciulli, F.; Consiglio, A.; Simone, M.; Viterbo, R.G.; Creanza, T.M.; Ancona, N.; Tortorella, C.; Margari, L.; et al. Combined microRNA and mRNA expression analysis in pediatric multiple sclerosis: An integrated approach to uncover novel pathogenic mechanisms of the disease. Hum. Mol. Genet. 2018, 27, 66–79. [Google Scholar] [CrossRef]
- Baulina, N.; Kulakova, O.; Kiselev, I.; Osmak, G.; Popova, E.; Boyko, A.; Favorova, O. Immune-related miRNA expression patterns in peripheral blood mononuclear cells differ in multiple sclerosis relapse and remission. J. Neuroimmunol. 2018, 317, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Pan, W.; Qian, L. Identification of the miRNA–mRNA regulatory network in multiple sclerosis. Neurol. Res. 2017, 39, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Bergman, P.; Piket, E.; Khademi, M.; James, T.; Brundin, L.; Olsson, T.; Piehl, F.; Jagodic, M. Circulating miR-150 in CSF is a novel candidate biomarker for multiple sclerosis. Neurol. Neuroimmunol. NeuroInflammation 2016, 3, e219. [Google Scholar] [CrossRef] [Green Version]
- Martinez, B.; Peplow, P. MicroRNAs as disease progression biomarkers and therapeutic targets in experimental autoimmune encephalomyelitis model of multiple sclerosis. Neural Regen. Res. 2020, 15, 1831–1837. [Google Scholar] [CrossRef] [PubMed]
- Angerstein, C.; Hecker, M.; Paap, B.K.; Koczan, D.; Thamilarasan, M.; Thiesen, H.J.; Zettl, U.K. Integration of MicroRNA databases to study MicroRNAs associated with multiple sclerosis. Mol. Neurobiol. 2012, 45, 520–535. [Google Scholar] [CrossRef]
- Juźwik, C.A.; Drake, S.S.; Zhang, Y.; Paradis-Isler, N.; Sylvester, A.; Amar-Zifkin, A.; Douglas, C.; Morquette, B.; Moore, C.S.; Fournier, A.E. microRNA dysregulation in neurodegenerative diseases: A systematic review. Prog. Neurobiol. 2019, 182, 101664. [Google Scholar] [CrossRef]
- Herrera-Espejo, S.; Santos-Zorrozua, B.; Álvarez-González, P.; Lopez-Lopez, E.; Garcia-Orad, Á. A Systematic Review of MicroRNA Expression as Biomarker of Late-Onset Alzheimer’s Disease. Mol. Neurobiol. 2019, 56, 8376–8391. [Google Scholar] [CrossRef]
- Lehmann, S.M.; Krüger, C.; Park, B.; Derkow, K.; Rosenberger, K.; Baumgart, J.; Trimbuch, T.; Eom, G.; Hinz, M.; Kaul, D.; et al. An unconventional role for miRNA: Let-7 activates Toll-like receptor 7 and causes neurodegeneration. Nat. Neurosci. 2012, 15, 827–835. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [Green Version]
- Portaccio, E.; Goretti, B.; Zipoli, V.; Siracusa, G.; Sorbi, S.; Amato, M.P. A short version of Rao’s brief repeatable battery as a screening tool for cognitive impairment in multiple sclerosis. Clin. Neuropsychol. 2009, 23, 268–275. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Jiang, X.; Chen, J. The role of miR-150 in normal and malignant hematopoiesis. Oncogene 2014, 33, 3887–3893. [Google Scholar] [CrossRef]
- Wells, A.C.; Daniels, K.A.; Angelou, C.C.; Fagerberg, E.; Burnside, A.S.; Markstein, M.; Alfandari, D.; Welsh, R.M.; Pobezinskaya, E.L.; Pobezinsky, L.A. Modulation of let-7 miRNAs controls the differentiation of effector CD8 T cells. Elife 2017, 6, e26398. [Google Scholar] [CrossRef]
- Thebault, S.; Abdoli, M.; Fereshtehnejad, S.M.; Tessier, D.; Tabard-Cossa, V.; Freedman, M.S. Serum neurofilament light chain predicts long term clinical outcomes in multiple sclerosis. Sci. Rep. 2020, 10, 10381. [Google Scholar] [CrossRef] [PubMed]
- Aktas, O.; Renner, A.; Huss, A.; Filser, M.; Baetge, S.; Stute, N.; Gasis, M.; Lepka, K.; Goebels, N.; Senel, M.; et al. Serum neurofilament light chain. Neurol. Neuroimmunol. Neuroinflamm. 2020, 7, e885. [Google Scholar] [CrossRef]
- Gaetani, L.; Salvadori, N.; Lisetti, V.; Eusebi, P.; Mancini, A.; Gentili, L.; Borrelli, A.; Portaccio, E.; Sarchielli, P.; Blennow, K.; et al. Cerebrospinal fluid neurofilament light chain tracks cognitive impairment in multiple sclerosis. J. Neurol. 2019, 266, 2157–2163. [Google Scholar] [CrossRef]
- Friedova, L.; Motyl, J.; Srpova, B.; Oechtering, J.; Barro, C.; Vodehnalova, K.; Andelova, M.; Noskova, L.; Fialová, L.; Havrdova, E.K.; et al. The weak association between neurofilament levels at multiple sclerosis onset and cognitive performance after 9 years. Mult. Scler. Relat. Disord. 2020, 46, 102534. [Google Scholar] [CrossRef]
- Mattioli, F.; Bellomi, F.; Stampatori, C.; Mariotto, S.; Ferrari, S.; Monaco, S.; Mancinelli, C.; Capra, R. Longitudinal serum neurofilament light chain (sNfL) concentration relates to cognitive function in multiple sclerosis patients. J. Neurol. 2020, 267, 2245–2251. [Google Scholar] [CrossRef]
- Kuhle, J.; Barro, C.; Andreasson, U.; Derfuss, T.; Lindberg, R.; Sandelius, Å.; Liman, V.; Norgren, N.; Blennow, K.; Zetterberg, H. Comparison of three analytical platforms for quantification of the neurofilament light chain in blood samples: ELISA, electrochemiluminescence immunoassay and Simoa. Clin. Chem. Lab. Med. 2016, 54, 1655–1661. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.H.; Pandey, M.; Seigneur, E.M.; Panicker, L.M.; Koo, L.; Schwartz, O.M.; Chen, W.; Chen, C.K.; Simonds, W.F. Knockout of G protein β5 impairs brain development and causes multiple neurologic abnormalities in mice. J. Neurochem. 2011, 119, 544–554. [Google Scholar] [CrossRef] [Green Version]
- Thévenot, E.; Moreau, A.W.; Rousseau, V.; Combeau, G.; Domenichini, F.; Jacquet, C.; Goupille, O.; Amar, M.; Kreis, P.; Fossier, P.; et al. p21-activated kinase 3 (PAK3) protein regulates synaptic transmission through its interaction with the Nck2/Grb4 protein adaptor. J. Biol. Chem. 2011, 286, 40044–40059. [Google Scholar] [CrossRef] [Green Version]
- Qian, Y.; Wu, B.; Lu, Y.; Zhou, W.; Wang, S.; Wang, H. Novel PAK3 gene missense variant associated with two Chinese siblings with intellectual disability: A case report. BMC Med. Genet. 2020, 21, 31. [Google Scholar] [CrossRef] [Green Version]
- Chaudhry, A.; Noor, A.; Degagne, B.; Baker, K.; Bok, L.A.; Brady, A.F.; Chitayat, D.; Chung, B.H.; Cytrynbaum, C.; Dyment, D.; et al. Phenotypic spectrum associated with PTCHD1 deletions and truncating mutations includes intellectual disability and autism spectrum disorder. Clin. Genet. 2015, 88, 224–233. [Google Scholar] [CrossRef] [PubMed]
- Ung, D.C.; Iacono, G.; Méziane, H.; Blanchard, E.; Papon, M.A.; Selten, M.; van Rhijn, J.R.; Montjean, R.; Rucci, J.; Martin, S.; et al. Ptchd1 deficiency induces excitatory synaptic and cognitive dysfunctions in mouse. Mol. Psychiatry 2018, 23, 1356–1367. [Google Scholar] [CrossRef] [Green Version]
- Guo, W.; Polich, E.D.; Su, J.; Gao, Y.; Christopher, D.M.; Allan, A.M.; Wang, M.; Wang, F.; Wang, G.; Zhao, X. Fragile X Proteins FMRP and FXR2P Control Synaptic GluA1 Expression and Neuronal Maturation via Distinct Mechanisms. Cell Rep. 2015, 11, 1651–1666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beesley, P.W.; Herrera-Molina, R.; Smalla, K.H.; Seidenbecher, C. The Neuroplastin adhesion molecules: Key regulators of neuronal plasticity and synaptic function. J. Neurochem. 2014, 131, 268–283. [Google Scholar] [CrossRef] [PubMed]
- Keren-Shaul, H.; Spinrad, A.; Weiner, A.; Matcovitch-Natan, O.; Dvir-Szternfeld, R.; Ulland, T.K.; David, E.; Baruch, K.; Lara-Astaiso, D.; Toth, B.; et al. A Unique Microglia Type Associated with Restricting Development of Alzheimer’s Disease. Cell 2017, 169, 1276–1290.e17. [Google Scholar] [CrossRef]
- Krasemann, S.; Madore, C.; Cialic, R.; Baufeld, C.; Calcagno, N.; El Fatimy, R.; Beckers, L.; O’Loughlin, E.; Xu, Y.; Fanek, Z.; et al. The TREM2-APOE Pathway Drives the Transcriptional Phenotype of Dysfunctional Microglia in Neurodegenerative Diseases. Immunity 2017, 47, 566–581.e9. [Google Scholar] [CrossRef] [Green Version]
- Virgilio, E.; Vecchio, D.; Crespi, I.; Puricelli, C.; Barbero, P.; Galli, G.; Cantello, R.; Dianzani, U.; Comi, C. Cerebrospinal fluid biomarkers and cognitive functions at multiple sclerosis diagnosis. J. Neurol. 2022, 1, 1–9. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).