
The Genetic Structures and Molecular Mechanisms Underlying Ear Traits in Maize (Zea mays L.)

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. The Literature Review and QTL/QTN Data Collection
2.2. The Construction of a Consensus Map for Ear Traits
2.3. The Identification of QTN and QTL Clusters
2.4. The Construction of the Molecular Networks Regulating Ear Traits and Candidate Gene Identification of QTN and QTL Clusters
2.5. Abbreviations
3. Results and Discussion
3.1. The Genetic Basis of the Ear Traits and Summary of the Genetic Loci Identified by QTL Mapping
3.2. The Genetic Basis of the Ear Traits and Summary of the Genetic Loci Identified by GWAS
3.3. A summary of the Molecular Mechanisms Regulating Ear Traits Based on Information of the Known Genes, and the Instruction for Prediction of the Candidate Genes of QTL and QTN Clusters
3.3.1. The Auxin Pathway and Regulation of AM Activity and Lateral Primordia Initiation
3.3.2. Three-Amino-Acid Loop Extension (TALE) Homeodomain Proteins and the Regulation of AM Activity and EN
3.3.3. The CLAVATA (CLV)-WUSCHEL (WUS) Negative Feedback Loop and Regulation of IM Activity and KRN
3.3.4. Reactive Oxygen Species (ROS) Signaling and Regulation of IM Activity and KRN
3.3.5. The RAMOSA (RA) Pathway and Inhibition of Branch Initiation in Maize Ears
3.3.6. The Ethylene Pathway and Regulating IM Activity and SM Identity
3.3.7. The miR172 Pathway and SM Fate Regulation during Maize Ear Development
3.3.8. Other Genes Regulating IM Activity with an Undetermined Molecular Pathway
3.4. The Application Potential of the Mined Data for Important Loci Recognition and New Gene Discovery
4. Conclusions and Perspectives
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- FAO. Available online: https://www.Fao.Org/faostat (accessed on 24 March 2023).
- Duvick, D. Genetic progress in yield of United States maize (Zea mays L.). Maydica 2005, 50, 193. [Google Scholar]
- Rosegrant, M.; Ringler, C.; Sulser, T.B.; Ewing, M.; Palazzo, A.; Zhu, T.; Nelson, G.C.; Koo, J.; Robertson, R.; Msangi, S. Agriculture and Food Security under Global Change: Prospects for 2025/2050; International Food Policy Research Institute: Washington, DC, USA, 2009; p. 89. [Google Scholar]
- Cheng, P.-C.; Pareddy, D.R. Morphology and development of the tassel and ear. In The Maize Handbook; Springer: Berlin/Heidelberg, Germany, 1994; pp. 37–47. [Google Scholar]
- Bennett, T.; Leyser, O. Something on the side: Axillary meristems and plant development. Plant Mol. Biol. 2006, 60, 843–854. [Google Scholar] [CrossRef]
- Vollbrecht, E.; Schmidt, R.J. Development of the inflorescences. In Handbook of Maize: Its Biology; Springer: Berlin/Heidelberg, Germany, 2009; pp. 13–40. [Google Scholar]
- Ning, Q.; Jian, Y.; Du, Y.; Li, Y.; Shen, X.; Jia, H.; Zhao, R.; Zhan, J.; Yang, F.; Jackson, D. An ethylene biosynthesis enzyme controls quantitative variation in maize ear length and kernel yield. Nat. Commun. 2021, 12, 5832. [Google Scholar] [CrossRef]
- Pei, Y.; Deng, Y.; Zhang, H.; Zhang, Z.; Liu, J.; Chen, Z.; Cai, D.; Li, K.; Du, Y.; Zang, J. Ear apical degeneration1 regulates maize ear development by maintaining malate supply for apical inflorescence. Plant Cell 2022, 34, 2222–2241. [Google Scholar] [CrossRef]
- Matsuoka, Y.; Vigouroux, Y.; Goodman, M.M.; Sanchez, G.J.; Buckler, E.; Doebley, J. A single domestication for maize shown by multilocus microsatellite genotyping. Proc. Natl. Acad. Sci. USA 2002, 99, 6080–6084. [Google Scholar] [CrossRef]
- Doebley, J. The genetics of maize evolution. Annu. Rev. Genet. 2004, 38, 37–59. [Google Scholar] [CrossRef] [Green Version]
- Piperno, D.R.; Ranere, A.J.; Holst, I.; Iriarte, J.; Dickau, R. Starch grain and phytolith evidence for early ninth millennium BP maize from the Central Balsas River Valley, Mexico. Proc. Natl. Acad. Sci. USA 2009, 106, 5019–5024. [Google Scholar] [CrossRef]
- Iltis, H.H. From teosinte to maize: The catastrophic sexual transmutation. Science 1983, 222, 886–894. [Google Scholar] [CrossRef] [Green Version]
- Doebley, J.; Stec, A.; Hubbard, L. The evolution of apical dominance in maize. Nature 1997, 386, 485–488. [Google Scholar] [CrossRef]
- Wills, D.M.; Whipple, C.J.; Takuno, S.; Kursel, L.E.; Shannon, L.M.; Ross-Ibarra, J.; Doebley, J.F. From many, one: Genetic control of prolificacy during maize domestication. PLoS Genet. 2013, 9, e1003604. [Google Scholar] [CrossRef] [Green Version]
- Otegui, M. Prolificacy and grain yield components in modern Argentinian maize hybrids. Maydica 1995, 40, 371–376. [Google Scholar]
- Amelong, A.; Gambín, B.L.; Severini, A.D.; Borrás, L. Predicting maize kernel number using QTL information. Field Crops Res. 2015, 172, 119–131. [Google Scholar] [CrossRef]
- Brown, P.J.; Upadyayula, N.; Mahone, G.S.; Tian, F.; Bradbury, P.J.; Myles, S.; Holland, J.B.; Flint-Garcia, S.; McMullen, M.D.; Buckler, E.S. Distinct genetic architectures for male and female inflorescence traits of maize. PLoS Genet. 2011, 7, e1002383. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Zhong, W.; Yang, F.; Zhang, Z. Genetic and molecular mechanisms of quantitative trait loci controlling maize inflorescence architecture. Plant Cell Physiol. 2018, 59, 448–457. [Google Scholar] [CrossRef] [Green Version]
- Veit, B.; Schmidt, R.J.; Hake, S.; Yanofsky, M.F. Maize floral development: New genes and old mutants. Plant Cell 1993, 5, 1205. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Yang, X.; Li, G. Molecular insights into inflorescence meristem specification for yield potential in cereal crops. Int. J. Mol. Sci. 2021, 22, 3508. [Google Scholar] [CrossRef]
- Upadyayula, N.; Da Silva, H.; Bohn, M.O.; Rocheford, T. Genetic and QTL analysis of maize tassel and ear inflorescence architecture. Theor. Appl. Genet. 2006, 112, 592–606. [Google Scholar] [CrossRef]
- Wang, Y.; Bao, J.; Wei, X.; Wu, S.; Fang, C.; Li, Z.; Qi, Y.; Gao, Y.; Dong, Z.; Wan, X. Genetic Structure and Molecular Mechanisms Underlying the Formation of Tassel, Anther, and Pollen in the Male Inflorescence of Maize (Zea mays L.). Cells 2022, 11, 1753. [Google Scholar] [CrossRef]
- Tanaka, W.; Pautler, M.; Jackson, D.; Hirano, H.-Y. Grass meristems II: Inflorescence architecture, flower development and meristem fate. Plant Cell Physiol. 2013, 54, 313–324. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Yuan, Z. Molecular control of grass inflorescence development. Annu. Rev. Plant Biol. 2014, 65, 553–578. [Google Scholar] [CrossRef]
- Du, Y.; Wu, B.; Xing, Y.; Zhang, Z. Conservation and divergence: Regulatory networks underlying reproductive branching in rice and maize. J. Adv. Res. 2022, 41, 179–190. [Google Scholar] [CrossRef]
- Eveland, A.L.; Goldshmidt, A.; Pautler, M.; Morohashi, K.; Liseron-Monfils, C.; Lewis, M.W.; Kumari, S.; Hiraga, S.; Yang, F.; Unger-Wallace, E.; et al. Regulatory modules controlling maize inflorescence architecture. Genome Res. 2014, 24, 431–443. [Google Scholar] [CrossRef] [Green Version]
- Hallauer, A.R.; Carena, M.J.; Miranda Filho, J.D. Quantitative Genetics in Maize Breeding; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2010; Volume 6. [Google Scholar]
- Veldboom, L.; Lee, M. Molecular-marker-facilitated studies of morphological traits in maize. II: Determination of QTLs for grain yield and yield components. Theor. Appl. Genet. 1994, 89, 451–458. [Google Scholar] [CrossRef]
- Edwards, M.; Helentjaris, T.; Wright, S.; Stuber, C. Molecular-marker-facilitated investigations of quantitative trait loci in maize. Theor. Appl. Genet. 1992, 83, 765–774. [Google Scholar] [CrossRef]
- Li, C.; Li, Y.; Sun, B.; Peng, B.; Liu, C.; Liu, Z.; Yang, Z.; Li, Q.; Tan, W.; Zhang, Y. Quantitative trait loci mapping for yield components and kernel-related traits in multiple connected RIL populations in maize. Euphytica 2013, 193, 303–316. [Google Scholar] [CrossRef]
- Fei, X.; Wang, Y.; Zheng, Y.; Shen, X.; Ding, J.; Lai, J.; Song, W.; Zhao, H. Identification of two new QTLs of maize (Zea mays L.) underlying kernel row number using the HNAU-NAM1 population. BMC Genom. 2022, 23, 593. [Google Scholar] [CrossRef]
- Lu, M.; Xie, C.-X.; Li, X.-H.; Hao, Z.-F.; Li, M.-S.; Weng, J.-F.; Zhang, D.-G.; Bai, L.; Zhang, S.-H. Mapping of quantitative trait loci for kernel row number in maize across seven environments. Mol. Breed. 2011, 28, 143–152. [Google Scholar] [CrossRef]
- Liu, L.; Du, Y.; Huo, D.; Wang, M.; Shen, X.; Yue, B.; Qiu, F.; Zheng, Y.; Yan, J.; Zhang, Z. Genetic architecture of maize kernel row number and whole genome prediction. Theor. Appl. Genet. 2015, 128, 2243–2254. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Ren, Z.; Luo, B.; Zhong, H.; Ma, P.; Zhang, H.; Hu, H.; Wang, Y.; Zhang, H.; Liu, D. Genetic architecture of maize yield traits dissected by QTL mapping and GWAS in maize. Crop J. 2022, 10, 436–446. [Google Scholar] [CrossRef]
- Liu, L.; Du, Y.; Shen, X.; Li, M.; Sun, W.; Huang, J.; Liu, Z.; Tao, Y.; Zheng, Y.; Yan, J. KRN4 controls quantitative variation in maize kernel row number. PLoS Genet. 2015, 11, e1005670. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Lin, Z.; Zhang, X.; Liu, H.; Zhou, L.; Zhong, S.; Li, Y.; Zhu, C.; Lin, Z. KRN1, a major quantitative trait locus for kernel row number in maize. New Phytol. 2019, 223, 1634–1646. [Google Scholar] [CrossRef]
- Jia, H.; Li, M.; Li, W.; Liu, L.; Jian, Y.; Yang, Z.; Shen, X.; Ning, Q.; Du, Y.; Zhao, R. A serine/threonine protein kinase encoding gene KERNEL NUMBER PER ROW6 regulates maize grain yield. Nat. Commun. 2020, 11, 988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.; Chen, L.; Zhang, X.; Yang, N.; Guo, J.; Wang, M.; Ji, S.; Zhao, X.; Yin, P.; Cai, L. Convergent selection of a WD40 protein that enhances grain yield in maize and rice. Science 2022, 375, eabg7985. [Google Scholar] [CrossRef]
- Lu, G.H.; Tang, J.H.; Yan, J.B.; Ma, X.Q.; Li, J.S.; Chen, S.J.; Ma, J.C.; Liu, Z.X.; Li-Zhu, E.; Zhang, Y.R. Quantitative trait loci mapping of maize yield and its components under different water treatments at flowering time. J. Integr. Plant Biol. 2006, 48, 1233–1243. [Google Scholar] [CrossRef]
- Yan, J.-B.; Tang, H.; Huang, Y.-Q.; Zheng, Y.-L.; Li, J.-S. Quantitative trait loci mapping and epistatic analysis for grain yield and yield components using molecular markers with an elite maize hybrid. Euphytica 2006, 149, 121–131. [Google Scholar] [CrossRef]
- Ma, X.; Tang, J.; Teng, W.; Yan, J.; Meng, Y.; Li, J. Epistatic interaction is an important genetic basis of grain yield and its components in maize. Mol. Breed. 2007, 20, 41–51. [Google Scholar] [CrossRef]
- Hao, Z.; Li, X.; Xie, C.; Li, M.; Zhang, D.; Bai, L.; Zhang, S. Two consensus quantitative trait loci clusters controlling anthesis–silking interval, ear setting and grain yield might be related with drought tolerance in maize. Ann. Appl. Biol. 2008, 153, 73–83. [Google Scholar] [CrossRef]
- Hongmei, Z.; Zuping, Z.; Xiaohong, L.; Zhong, L.; Chuan, H.; Daihui, L.; Yangchun, L.; Guoqin, Z.; Zhenbo, T.; Runzhi, L. QTL mapping for ear length and ear diameter under different nitrogen regimes in maize. Afr. J. Agric. Res. 2010, 5, 626–630. [Google Scholar]
- Liu, X.; Zheng, Z.; Tan, Z.; Li, Z.; He, C. Genetic analysis of two new quantitative trait loci for ear weight in maize inbred line Huangzao4. Genet. Mol. Res. 2010, 9, 2140–2147. [Google Scholar] [CrossRef]
- Liu, X.; He, S.; Zheng, Z.; Huang, Y.; Tan, Z.; Wu, X. QTL identification for row number per ear and grain number per row in maize. Maydica 2010, 55, 127–133. [Google Scholar]
- Tang, J.; Yan, J.; Ma, X.; Teng, W.; Wu, W.; Dai, J.; Dhillon, B.S.; Melchinger, A.E.; Li, J. Dissection of the genetic basis of heterosis in an elite maize hybrid by QTL mapping in an immortalized F2 population. Theor. Appl. Genet. 2010, 120, 333–340. [Google Scholar] [CrossRef]
- Guo, J.; Chen, Z.; Liu, Z.; Wang, B.; Song, W.; Li, W.; Chen, J.; Dai, J.; Lai, J. Identification of genetic factors affecting plant density response through QTL mapping of yield component traits in maize (Zea mays L.). Euphytica 2011, 182, 409–422. [Google Scholar] [CrossRef]
- Holland, J.B.; Coles, N.D. QTL controlling masculinization of ear tips in a maize (Zea mays L.) intraspecific cross. G3 Genes Genomes Genet. 2011, 1, 337–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, B.; Li, Y.; Wang, Y.; Liu, C.; Liu, Z.; Tan, W.; Zhang, Y.; Wang, D.; Shi, Y.; Sun, B. QTL analysis for yield components and kernel-related traits in maize across multi-environments. Theor. Appl. Genet. 2011, 122, 1305–1320. [Google Scholar] [CrossRef] [PubMed]
- Studer, A.J.; Doebley, J.F. Do large effect QTL fractionate? A case study at the maize domestication QTL teosinte branched1. Genetics 2011, 188, 673–681. [Google Scholar] [CrossRef] [Green Version]
- Choe, E.; Rocheford, T.R. Genetic and QTL analysis of pericarp thickness and ear architecture traits of Korean waxy corn germplasm. Euphytica 2012, 183, 243–260. [Google Scholar] [CrossRef]
- Nikolić, A.; Ignjatović-Micić, D.; Dodig, D.; Anđelković, V.; Lazić-Jančić, V. Identification of QTLs for yield and drought-related traits in maize: Assessment of their causal relationships. Biotechnol. Biotechnol. Equip. 2012, 26, 2952–2960. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Wang, X.; Wang, B.; Tian, Y.; Li, M.; Nie, Y.; Peng, Q.; Wang, Z. Fine mapping a major QTL for kernel number per row under different phosphorus regimes in maize (Zea mays L.). Theor. Appl. Genet. 2013, 126, 1545–1553. [Google Scholar] [CrossRef]
- Cai, L.; Li, K.; Yang, X.; Li, J. Identification of large-effect QTL for kernel row number has potential for maize yield improvement. Mol. Breed. 2014, 34, 1087–1096. [Google Scholar] [CrossRef]
- Chen, Z.; Wang, B.; Dong, X.; Liu, H.; Ren, L.; Chen, J.; Hauck, A.; Song, W.; Lai, J. An ultra-high density bin-map for rapid QTL mapping for tassel and ear architecture in a large F2 maize population. BMC Genom. 2014, 15, 433. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Jia, H.; Liu, L.; Zhang, C.; Liu, Z.; Zhang, Z. Quantitative trait loci mapping for kernel row number using chromosome segment substitution lines in maize. Genet. Mol. Res. 2014, 13, 1707–1716. [Google Scholar] [CrossRef]
- Park, K.J.; Sa, K.J.; Kim, B.W.; Koh, H.-J.; Lee, J.K. Genetic mapping and QTL analysis for yield and agronomic traits with an F2: 3 population derived from a waxy corn × sweet corn cross. Genes Genom. 2014, 36, 179–189. [Google Scholar] [CrossRef]
- Tian, B.; Wang, J.; Wang, G. Confirmation of a major QTL on chromosome 10 for maize kernel row number in different environments. Plant Breed. 2014, 133, 184–188. [Google Scholar] [CrossRef]
- Yu, Y.; Li, G.; Yang, Z.; Hu, J.; Zheng, J.; Qi, X. Identification of a major quantitative trait locus for ear size induced by space flight in sweet corn. Genet. Mol. Res. 2014, 13, 3069–3078. [Google Scholar] [CrossRef]
- Mendes-Moreira, P.; Alves, M.L.; Satovic, Z.; Dos Santos, J.P.; Santos, J.N.; Souza, J.C.; Pego, S.E.; Hallauer, A.R.; Vaz Patto, M.C. Genetic architecture of ear fasciation in maize (Zea mays) under QTL scrutiny. PLoS ONE 2015, 10, e0124543. [Google Scholar] [CrossRef]
- Sa, K.J.; Park, J.Y.; Woo, S.Y.; Ramekar, R.V.; Jang, C.-S.; Lee, J.K. Mapping of QTL traits in corn using a RIL population derived from a cross of dent corn × waxy corn. Genes Genom. 2015, 37, 1–14. [Google Scholar] [CrossRef]
- Yang, C.; Liu, J.; Rong, T. Detection of quantitative trait loci for ear row number in F2 populations of maize. Genet. Mol. Res. 2015, 14, 14229–14238. [Google Scholar] [CrossRef]
- Yang, C.; Tang, D.; Zhang, L.; Liu, J.; Rong, T. Identification of QTL for ear row number and two-ranked versus many-ranked ear in maize across four environments. Euphytica 2015, 206, 33–47. [Google Scholar] [CrossRef]
- Zhou, G.; Zhu, Q.; Yang, G.; Huang, J.; Cheng, S.; Yue, B.; Zhang, Z. qEL7.2 is a pleiotropic QTL for kernel number per row, ear length and ear weight in maize (Zea mays L.). Euphytica 2015, 203, 429–436. [Google Scholar] [CrossRef]
- Calderon, C.I.; Yandell, B.S.; Doebley, J.F. Fine mapping of a QTL associated with kernel row number on chromosome 1 of maize. PLoS ONE 2016, 11, e0150276. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Li, C.; Li, Y.; Song, Y.; Zhang, D.; Wang, T.; Li, Y.; Shi, Y. Quantitative trait loci mapping of yield and related traits using a high-density genetic map of maize. Mol. Breed. 2016, 36, 134. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, L.; Liu, S.; Li, Z.; Huang, R.; Li, Y.; Cheng, H.; Li, X.; Zhou, B.; Wu, S. The genetic basis of natural variation in kernel size and related traits using a four-way cross population in maize. PLoS ONE 2016, 11, e0153428. [Google Scholar] [CrossRef] [Green Version]
- Ding, J.; Ma, J.; Chen, J.; Ai, T.; Li, Z.; Tian, Z.; Wu, S.; Chen, W.; Wu, J. QTL mapping for ear tip-barrenness in maize. Span. J. Agric. Res. 2016, 14, e07SC01. [Google Scholar] [CrossRef] [Green Version]
- Huo, D.; Ning, Q.; Shen, X.; Liu, L.; Zhang, Z. QTL mapping of kernel number-related traits and validation of one major QTL for ear length in maize. PLoS ONE 2016, 11, e0155506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Zhou, Q.; Dong, L.; Wang, H.; Liu, F.; Weng, J.; Li, X.; Xie, C. Genetic architecture of the maize kernel row number revealed by combining QTL mapping using a high-density genetic map and bulked segregant RNA sequencing. BMC Genom. 2016, 17, 915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, X.; Lu, X.; Zhang, Z.; Xu, M.; Mao, K.; Li, W.; Wei, F.; Sun, P.; Tang, J. Genetic analysis of heterosis for maize grain yield and its components in a set of SSSL testcross populations. Euphytica 2016, 210, 181–193. [Google Scholar] [CrossRef]
- Yang, C.; Zhang, L.; Jia, A.; Rong, T. Identification of QTL for maize grain yield and kernel-related traits. J. Genet. 2016, 95, 239–247. [Google Scholar] [CrossRef]
- Hu, X.; Wang, H.; Li, K.; Wu, Y.; Liu, Z.; Huang, C. Genome-wide proteomic profiling reveals the role of dominance protein expression in heterosis in immature maize ears. Sci. Rep. 2017, 7, 16130. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Yang, Q.; Gao, L.; Zhang, M.; Ni, Z.; Zhang, Y. Identification of heterosis-associated stable QTLs for ear-weight-related traits in an elite maize hybrid Zhengdan 958 by design III. Front. Plant Sci. 2017, 8, 561. [Google Scholar] [CrossRef] [Green Version]
- Pan, L.; Yin, Z.; Huang, Y.; Chen, J.; Zhu, L.; Zhao, Y.; Guo, J. QTL for maize grain yield identified by QTL mapping in six environments and consensus loci for grain weight detected by meta-analysis. Plant Breed. 2017, 136, 820–833. [Google Scholar] [CrossRef]
- Su, C.; Wang, W.; Gong, S.; Zuo, J.; Li, S.; Xu, S. High density linkage map construction and mapping of yield trait QTLs in maize (Zea mays) using the genotyping-by-sequencing (GBS) technology. Front. Plant Sci. 2017, 8, 706. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Zhou, Z.; Yong, H.; Zhang, X.; Hao, Z.; Zhang, F.; Li, M.; Zhang, D.; Li, X.; Wang, Z. Analysis of the genetic architecture of maize ear and grain morphological traits by combined linkage and association mapping. Theor. Appl. Genet. 2017, 130, 1011–1029. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.; Zhang, R.; Xing, J.; Duan, M.; Wang, Y.; Su, A.; Wang, F.; Xu, L.; Tian, H.; Wang, J. QTL mapping of three ear traits using a doubled haploid population of maize. Plant Breed. 2018, 137, 706–713. [Google Scholar] [CrossRef]
- Zhan, J.; Wang, F.; Xing, W.; Liu, J.; Fan, Z.; Tao, Y. Fine mapping and candidate gene prediction of a major QTL for kernel number per ear in maize. Mol. Breed. 2018, 38, 1–9. [Google Scholar] [CrossRef]
- Zhou, B.; Zhou, Z.; Ding, J.; Zhang, X.; Mu, C.; Wu, Y.; Gao, J.; Song, Y.; Wang, S.; Ma, J. Combining three mapping strategies to reveal quantitative trait loci and candidate genes for maize ear length. Plant Genome 2018, 11, 170107. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.-K.; Sa, K.J.; Park, D.H.; Lim, S.E.; Ryu, S.-H.; Park, J.Y.; Park, K.J.; Rhee, H.-I.; Lee, M.; Lee, J.K. Construction of genetic linkage map and identification of QTLs related to agronomic traits in DH population of maize (Zea mays L.) using SSR markers. Genes Genom. 2019, 41, 667–678. [Google Scholar] [CrossRef]
- Nie, N.; Ding, X.; Chen, L.; Wu, X.; An, Y.; Li, C.; Song, Y.; Zhang, D.; Liu, Z.; Wang, T. Characterization and fine mapping of qkrnw4, a major QTL controlling kernel row number in maize. Theor. Appl. Genet. 2019, 132, 3321–3331. [Google Scholar] [CrossRef]
- Yi, Q.; Liu, Y.; Hou, X.; Zhang, X.; Li, H.; Zhang, J.; Liu, H.; Hu, Y.; Yu, G.; Li, Y. Genetic dissection of yield-related traits and mid-parent heterosis for those traits in maize (Zea mays L.). BMC Plant Biol. 2019, 19, 392. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Su, C. Mapping quantitative trait loci for yield-related traits and predicting candidate genes for grain weight in maize. Sci. Rep. 2019, 9, 16112. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Liu, P.; Zhang, X.; Zhang, Y.; Ma, L.; Liu, M.; Guan, Z.; Zhang, Y.; Li, P.; Zou, C. Genome-wide association studies and QTL mapping uncover the genetic architecture of ear tip-barrenness in maize. Physiol. Plant. 2020, 170, 27–39. [Google Scholar] [CrossRef]
- Yi, Q.; Liu, Y.; Hou, X.; Zhang, X.; Zhang, J.; Liu, H.; Hu, Y.; Yu, G.; Li, Y.; Wang, Y. Quantitative trait loci mapping for yield-related traits under low and high planting densities in maize (Zea mays). Plant Breed. 2020, 139, 227–240. [Google Scholar] [CrossRef]
- Zhang, X.; Guan, Z.; Li, Z.; Liu, P.; Ma, L.; Zhang, Y.; Pan, L.; He, S.; Zhang, Y.; Li, P. A combination of linkage mapping and GWAS brings new elements on the genetic basis of yield-related traits in maize across multiple environments. Theor. Appl. Genet. 2020, 133, 2881–2895. [Google Scholar] [CrossRef]
- Liu, X.; Hu, X.; Li, K.; Liu, Z.; Wu, Y.; Feng, G.; Huang, C.; Wang, H. Identifying quantitative trait loci for the general combining ability of yield-relevant traits in maize. Breed. Sci. 2021, 71, 217–228. [Google Scholar] [CrossRef]
- Mei, X.; Dong, E.; Liang, Q.; Bai, Y.; Nan, J.; Yang, Y.; Cai, Y. Identification of QTL for fasciated ear related traits in maize. Crop Sci. 2021, 61, 1184–1193. [Google Scholar] [CrossRef]
- Yang, L.; Li, T.; Liu, B.; Li, R.; Yu, R.; Zhang, X.; Li, Q.; Xu, S.; Xue, J. Genetic analysis of ear-related traits under different pollination treatments in maize (Zea mays). Plant Breed. 2021, 140, 211–222. [Google Scholar] [CrossRef]
- Xiao, Y.; Liu, H.; Wu, L.; Warburton, M.; Yan, J. Genome-wide association studies in maize: Praise and stargaze. Mol. Plant 2017, 10, 359–374. [Google Scholar] [CrossRef] [Green Version]
- Yang, N.; Lu, Y.; Yang, X.; Huang, J.; Zhou, Y.; Ali, F.; Wen, W.; Liu, J.; Li, J.; Yan, J. Genome wide association studies using a new nonparametric model reveal the genetic architecture of 17 agronomic traits in an enlarged maize association panel. PLoS Genet. 2014, 10, e1004573. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Y.; Tong, H.; Yang, X.; Xu, S.; Pan, Q.; Qiao, F.; Raihan, M.S.; Luo, Y.; Liu, H.; Zhang, X. Genome-wide dissection of the maize ear genetic architecture using multiple populations. New Phytol. 2016, 210, 1095–1106. [Google Scholar] [CrossRef] [Green Version]
- Park, J.Y.; Ramekar, R.V.; Sa, K.J.; Lee, J.K. Genetic diversity, population structure, and association mapping of biomass traits in maize with simple sequence repeat markers. Genes Genom. 2015, 37, 725–735. [Google Scholar] [CrossRef]
- Sa, K.; Park, J.; Choi, S.; Kim, B.; Park, K.; Lee, J. Genetic diversity, population structure, and association mapping of agronomic traits in waxy and normal maize inbred lines. Genet. Mol. Res. 2015, 14, 7502–7518. [Google Scholar] [CrossRef]
- Xue, S.; Bradbury, P.J.; Casstevens, T.; Holland, J.B. Genetic architecture of domestication-related traits in maize. Genetics 2016, 204, 99–113. [Google Scholar] [CrossRef] [Green Version]
- Bouchet, S.; Bertin, P.; Presterl, T.; Jamin, P.; Coubriche, D.; Gouesnard, B.; Laborde, J.; Charcosset, A. Association mapping for phenology and plant architecture in maize shows higher power for developmental traits compared with growth influenced traits. Heredity 2017, 118, 249–259. [Google Scholar] [CrossRef] [Green Version]
- Lai, X.; Schnable, J.C.; Liao, Z.; Xu, J.; Zhang, G.; Li, C.; Hu, E.; Rong, T.; Xu, Y.; Lu, Y. Genome-wide characterization of non-reference transposable element insertion polymorphisms reveals genetic diversity in tropical and temperate maize. BMC Genom. 2017, 18, 702. [Google Scholar] [CrossRef]
- Xu, Y.; Xu, C.; Xu, S. Prediction and association mapping of agronomic traits in maize using multiple omic data. Heredity 2017, 119, 174–184. [Google Scholar] [CrossRef] [Green Version]
- An, Y.; Chen, L.; Li, Y.-X.; Li, C.; Shi, Y.; Song, Y.; Zhang, D.; Li, Y.; Wang, T. Candidate loci for the kernel row number in maize revealed by a combination of transcriptome analysis and regional association mapping. BMC Plant Biol. 2019, 19, 201. [Google Scholar] [CrossRef]
- Chen, Q.; Yang, C.J.; York, A.M.; Xue, W.; Daskalska, L.L.; DeValk, C.A.; Krueger, K.W.; Lawton, S.B.; Spiegelberg, B.G.; Schnell, J.M. TeoNAM: A nested association mapping population for domestication and agronomic trait analysis in maize. Genetics 2019, 213, 1065–1078. [Google Scholar] [CrossRef] [Green Version]
- An, Y.; Chen, L.; Li, Y.-X.; Li, C.; Shi, Y.; Zhang, D.; Li, Y.; Wang, T. Genome-wide association studies and whole-genome prediction reveal the genetic architecture of KRN in maize. BMC Plant Biol. 2020, 20, 490. [Google Scholar] [CrossRef]
- Chen, H.; Hao, Z.; Zhao, Y.; Yang, R. A fast-linear mixed model for genome-wide haplotype association analysis: Application to agronomic traits in maize. BMC Genom. 2020, 21, 151. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Qu, J.; Tian, X.; Lao, Y.; Wei, N.; Wang, Y.; Hao, Y.; Zhang, X.; Xue, J.; Xu, S. Identification of ear morphology genes in maize (Zea mays L.) using selective sweeps and association mapping. Front. Genet. 2020, 11, 747. [Google Scholar] [CrossRef]
- Liu, H.-J.; Wang, X.; Xiao, Y.; Luo, J.; Qiao, F.; Yang, W.; Zhang, R.; Meng, Y.; Sun, J.; Yan, S. CUBIC: An atlas of genetic architecture promises directed maize improvement. Genome Biol. 2020, 21, 20. [Google Scholar] [CrossRef]
- Rice, B.R.; Fernandes, S.B.; Lipka, A.E. Multi-trait genome-wide association studies reveal loci associated with maize inflorescence and leaf architecture. Plant Cell Physiol. 2020, 61, 1427–1437. [Google Scholar] [CrossRef]
- Yang, L.; Li, T.; Tian, X.; Yang, B.; Lao, Y.; Wang, Y.; Zhang, X.; Xue, J.; Xu, S. Genome-wide association study (GWAS) reveals genetic basis of ear-related traits in maize. Euphytica 2020, 216, 172. [Google Scholar] [CrossRef]
- Liu, N.; Du, Y.; Warburton, M.L.; Xiao, Y.; Yan, J. Phenotypic plasticity contributes to maize adaptation and heterosis. Mol. Biol. Evol. 2021, 38, 1262–1275. [Google Scholar] [CrossRef]
- Ma, J.; Cao, Y. Genetic dissection of grain yield of maize and yield-related traits through association mapping and genomic prediction. Front. Plant Sci. 2021, 12, 690059. [Google Scholar] [CrossRef]
- Zeng, T.; Meng, Z.; Yue, R.; Lu, S.; Li, W.; Li, W.; Meng, H.; Sun, Q. Genome wide association analysis for yield related traits in maize. BMC Plant Biol. 2022, 22, 449. [Google Scholar] [CrossRef]
- Galli, M.; Liu, Q.; Moss, B.L.; Malcomber, S.; Li, W.; Gaines, C.; Federici, S.; Roshkovan, J.; Meeley, R.; Nemhauser, J.L. Auxin signaling modules regulate maize inflorescence architecture. Proc. Natl. Acad. Sci. USA 2015, 112, 13372–13377. [Google Scholar] [CrossRef]
- Gallavotti, A.; Barazesh, S.; Malcomber, S.; Hall, D.; Jackson, D.; Schmidt, R.J.; McSteen, P. Sparse inflorescence1 encodes a monocot-specific YUCCA-like gene required for vegetative and reproductive development in maize. Proc. Natl. Acad. Sci. USA 2008, 105, 15196–15201. [Google Scholar] [CrossRef]
- Phillips, K.A.; Skirpan, A.L.; Liu, X.; Christensen, A.; Slewinski, T.L.; Hudson, C.; Barazesh, S.; Cohen, J.D.; Malcomber, S.; McSteen, P. Vanishing tassel2 encodes a grass-specific tryptophan aminotransferase required for vegetative and reproductive development in maize. Plant Cell 2011, 23, 550–566. [Google Scholar] [CrossRef] [Green Version]
- Friml, J.; Yang, X.; Michniewicz, M.; Weijers, D.; Quint, A.; Tietz, O.; Benjamins, R.; Ouwerkerk, P.B.; Ljung, K.; Sandberg, G. A PINOID-dependent binary switch in apical-basal PIN polar targeting directs auxin efflux. Science 2004, 306, 862–865. [Google Scholar] [CrossRef] [Green Version]
- McSteen, P.; Malcomber, S.; Skirpan, A.; Lunde, C.; Wu, X.; Kellogg, E.; Hake, S. Barren inflorescence2 encodes a co-ortholog of the PINOID serine/threonine kinase and is required for organogenesis during inflorescence and vegetative development in maize. Plant Physiol. 2007, 144, 1000–1011. [Google Scholar] [CrossRef] [Green Version]
- Skirpan, A.; Culler, A.H.; Gallavotti, A.; Jackson, D.; Cohen, J.D.; McSteen, P. BARREN INFLORESCENCE2 interaction with ZmPIN1a suggests a role in auxin transport during maize inflorescence development. Plant Cell Physiol. 2009, 50, 652–657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carraro, N.; Forestan, C.; Canova, S.; Traas, J.; Varotto, S. ZmPIN1a and ZmPIN1b encode two novel putative candidates for polar auxin transport and plant architecture determination of maize. Plant Physiol. 2006, 142, 254–264. [Google Scholar] [CrossRef] [Green Version]
- Gallavotti, A.; Yang, Y.; Schmidt, R.J.; Jackson, D. The relationship between auxin transport and maize branching. Plant Physiol. 2008, 147, 1913–1923. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Zhang, X.; Zhao, Y.; Li, Y.; Zhang, G.; Peng, Z.; Zhang, J. Enhancing auxin accumulation in maize root tips improves root growth and dwarfs plant height. Plant Biotechnol. J. 2018, 16, 86–99. [Google Scholar] [CrossRef] [Green Version]
- Salehin, M.; Bagchi, R.; Estelle, M. SCFTIR1/AFB-based auxin perception: Mechanism and role in plant growth and development. Plant Cell 2015, 27, 9–19. [Google Scholar] [CrossRef] [Green Version]
- Gallavotti, A.; Zhao, Q.; Kyozuka, J.; Meeley, R.B.; Ritter, M.K.; Doebley, J.F.; Pe, M.E.; Schmidt, R.J. The role of barren stalk1 in the architecture of maize. Nature 2004, 432, 630–635. [Google Scholar] [CrossRef]
- Skirpan, A.; Wu, X.; McSteen, P. Genetic and physical interaction suggest that BARREN STALK1 is a target of BARREN INFLORESCENCE2 in maize inflorescence development. Plant J. 2008, 55, 787–797. [Google Scholar] [CrossRef]
- Gallavotti, A.; Malcomber, S.; Gaines, C.; Stanfield, S.; Whipple, C.; Kellogg, E.; Schmidt, R.J. BARREN STALK FASTIGIATE1 is an AT-hook protein required for the formation of maize ears. Plant Cell 2011, 23, 1756–1771. [Google Scholar] [CrossRef] [Green Version]
- Yao, H.; Skirpan, A.; Wardell, B.; Matthes, M.S.; Best, N.B.; McCubbin, T.; Durbak, A.; Smith, T.; Malcomber, S.; McSteen, P. The barren stalk2 gene is required for axillary meristem development in maize. Mol. Plant 2019, 12, 374–389. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Zhao, R.; Du, Y.; Shen, X.; Ning, Q.; Li, Y.; Liu, D.; Xiong, Q.; Zhang, Z. The Coordinated KNR6–AGAP–ARF1 Complex Modulates Vegetative and Reproductive Traits by Participating in Vesicle Trafficking in Maize. Cells 2021, 10, 2601. [Google Scholar] [CrossRef]
- Zhuang, X.; Jiang, J.; Li, J.; Ma, Q.; Xu, Y.; Xue, Y.; Xu, Z.; Chong, K. Over-expression of OsAGAP, an ARF-GAP, interferes with auxin influx, vesicle trafficking and root development. Plant J. 2006, 48, 581–591. [Google Scholar] [CrossRef] [PubMed]
- Serikawa, K.A.; Martinez-Laborda, A.; Zambryski, P. Three knotted1-like homeobox genes in Arabidopsis. Plant Mol. Biol. 1996, 32, 673–683. [Google Scholar] [CrossRef] [PubMed]
- Venglat, S.; Dumonceaux, T.; Rozwadowski, K.; Parnell, L.; Babic, V.; Keller, W.; Martienssen, R.; Selvaraj, G.; Datla, R. The homeobox gene BREVIPEDICELLUS is a key regulator of inflorescence architecture in Arabidopsis. Proc. Natl. Acad. Sci. USA 2002, 99, 4730–4735. [Google Scholar] [CrossRef]
- Ragni, L.; Belles-Boix, E.; Günl, M.; Pautot, V. Interaction of KNAT6 and KNAT2 with BREVIPEDICELLUS and PENNYWISE in Arabidopsis inflorescences. Plant Cell 2008, 20, 888–900. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Pi, L.; Huang, H.; Xu, L. ATH1 and KNAT2 proteins act together in regulation of plant inflorescence architecture. J. Exp. Bot. 2012, 63, 1423–1433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kerstetter, R.A.; Laudencia-Chingcuanco, D.; Smith, L.G.; Hake, S. Loss-of-function mutations in the maize homeobox gene, knotted1, are defective in shoot meristem maintenance. Development 1997, 124, 3045–3054. [Google Scholar] [CrossRef] [PubMed]
- Vollbrecht, E.; Reiser, L.; Hake, S. Shoot meristem size is dependent on inbred background and presence of the maize homeobox gene, knotted1. Development 2000, 127, 3161–3172. [Google Scholar] [CrossRef]
- Bolduc, N.; Yilmaz, A.; Mejia-Guerra, M.K.; Morohashi, K.; O’Connor, D.; Grotewold, E.; Hake, S. Unraveling the KNOTTED1 regulatory network in maize meristems. Genes Dev. 2012, 26, 1685–1690. [Google Scholar] [CrossRef] [Green Version]
- Bolduc, N.; Hake, S. The maize transcription factor KNOTTED1 directly regulates the gibberellin catabolism gene ga2ox1. Plant Cell 2009, 21, 1647–1658. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Wen, Y.; Cui, M.; Qi, X.; Deng, R.; Gao, J.; Cheng, Z. Histological, physiological and transcriptomic analysis reveal gibberellin-induced axillary meristem formation in garlic (Allium sativum). Plants 2020, 9, 970. [Google Scholar] [CrossRef]
- Zhang, Q.Q.; Wang, J.G.; Wang, L.Y.; Wang, J.F.; Wang, Q.; Yu, P.; Bai, M.Y.; Fan, M. Gibberellin repression of axillary bud formation in Arabidopsis by modulation of DELLA-SPL9 complex activity. J. Integr. Plant Biol. 2020, 62, 421–432. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.-H.; Lin, H.; Joo, S.; Goodenough, U. Early sexual origins of homeoprotein heterodimerization and evolution of the plant KNOX/BELL family. Cell 2008, 133, 829–840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Byrne, M.E.; Groover, A.T.; Fontana, J.R.; Martienssen, R.A. Phyllotactic pattern and stem cell fate are determined by the Arabidopsis homeobox gene BELLRINGER. Development 2003, 130, 3941–3950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, H.M.; Hake, S. The interaction of two homeobox genes, BREVIPEDICELLUS and PENNYWISE, regulates internode patterning in the Arabidopsis inflorescence. Plant Cell 2003, 15, 1717–1727. [Google Scholar] [CrossRef] [Green Version]
- Tsuda, K.; Abraham-Juarez, M.-J.; Maeno, A.; Dong, Z.; Aromdee, D.; Meeley, R.; Shiroishi, T.; Nonomura, K.-I.; Hake, S. KNOTTED1 cofactors, BLH12 and BLH14, regulate internode patterning and vein anastomosis in maize. Plant Cell 2017, 29, 1105–1118. [Google Scholar] [CrossRef] [Green Version]
- Schoof, H.; Lenhard, M.; Haecker, A.; Mayer, K.F.; Jürgens, G.; Laux, T. The stem cell population of Arabidopsis shoot meristems is maintained by a regulatory loop between the CLAVATA and WUSCHEL genes. Cell 2000, 100, 635–644. [Google Scholar] [CrossRef] [Green Version]
- Fletcher, J.C. The CLV-WUS stem cell signaling pathway: A roadmap to crop yield optimization. Plants 2018, 7, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, S.E.; Williams, R.W.; Meyerowitz, E.M. The CLAVATA1 gene encodes a putative receptor kinase that controls shoot and floral meristem size in Arabidopsis. Cell 1997, 89, 575–585. [Google Scholar] [CrossRef] [Green Version]
- Fletcher, J.C.; Brand, U.; Running, M.P.; Simon, R.; Meyerowitz, E.M. Signaling of cell fate decisions by CLAVATA3 in Arabidopsis shoot meristems. Science 1999, 283, 1911–1914. [Google Scholar] [CrossRef]
- Jeong, S.; Trotochaud, A.E.; Clark, S.E. The Arabidopsis CLAVATA2 gene encodes a receptor-like protein required for the stability of the CLAVATA1 receptor-like kinase. Plant Cell 1999, 11, 1925–1933. [Google Scholar] [CrossRef] [Green Version]
- Trotochaud, A.E.; Jeong, S.; Clark, S.E. CLAVATA3, a multimeric ligand for the CLAVATA1 receptor-kinase. Science 2000, 289, 613–617. [Google Scholar] [CrossRef] [PubMed]
- Brand, U.; Grunewald, M.; Hobe, M.; Simon, R. Regulation of CLV3 expression by two homeobox genes in Arabidopsis. Plant Physiol. 2002, 129, 565–575. [Google Scholar] [CrossRef] [Green Version]
- Brand, U.; Fletcher, J.C.; Hobe, M.; Meyerowitz, E.M.; Simon, R. Dependence of stem cell fate in Arabidopsis on a feedback loop regulated by CLV3 activity. Science 2000, 289, 617–619. [Google Scholar] [CrossRef] [PubMed]
- Bommert, P.; Lunde, C.; Nardmann, J.; Vollbrecht, E.; Running, M.; Jackson, D.; Hake, S.; Werr, W. Thick tassel dwarf1 encodes a putative maize ortholog of the Arabidopsis CLAVATA1 leucine-rich repeat receptor-like kinase. Development 2005, 132, 1235–1245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taguchi-Shiobara, F.; Yuan, Z.; Hake, S.; Jackson, D. The fasciated ear2 gene encodes a leucine-rich repeat receptor-like protein that regulates shoot meristem proliferation in maize. Genes Dev. 2001, 15, 2755–2766. [Google Scholar] [CrossRef] [Green Version]
- Je, B.I.; Gruel, J.; Lee, Y.K.; Bommert, P.; Arevalo, E.D.; Eveland, A.L.; Wu, Q.; Goldshmidt, A.; Meeley, R.; Bartlett, M. Signaling from maize organ primordia via FASCIATED EAR3 regulates stem cell proliferation and yield traits. Nat. Genet. 2016, 48, 785–791. [Google Scholar] [CrossRef] [Green Version]
- Bommert, P.; Nagasawa, N.S.; Jackson, D. Quantitative variation in maize kernel row number is controlled by the FASCIATED EAR2 locus. Nat. Genet. 2013, 45, 334–337. [Google Scholar] [CrossRef]
- Bommert, P.; Je, B.I.; Goldshmidt, A.; Jackson, D. The maize Gα gene COMPACT PLANT2 functions in CLAVATA signalling to control shoot meristem size. Nature 2013, 502, 555–558. [Google Scholar] [CrossRef]
- Je, B.; Xu, F.; Wu, Q.; Liu, L.; Meeley, R.; Gallagher, J.; Corcilius, L.; Payne, R.; Bartlett, M.; Jackson, D. The CLAVATA receptor FASCIATED EAR2 responds to distinct CLE peptides by signaling through two downstream effectors. Elife 2018, 7, e35673. [Google Scholar] [CrossRef]
- Urano, D.; Jackson, D.; Jones, A.M. AG protein alpha null mutation confers prolificacy potential in maize. J. Exp. Bot. 2015, 66, 4511–4515. [Google Scholar] [CrossRef] [Green Version]
- Nardmann, J.; Werr, W. The shoot stem cell niche in angiosperms: Expression patterns of WUS orthologues in rice and maize imply major modifications in the course of mono-and dicot evolution. Mol. Biol. Evol. 2006, 23, 2492–2504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Li, W.; Gaines, C.; Buck, A.; Galli, M.; Gallavotti, A. Structural variation at the maize WUSCHEL1 locus alters stem cell organization in inflorescences. Nat. Commun. 2021, 12, 2378. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Tian, C.; Zhang, C.; Shi, B.; Cao, X.; Zhang, T.-Q.; Zhao, Z.; Wang, J.-W.; Jiao, Y. Cytokinin signaling activates WUSCHEL expression during axillary meristem initiation. Plant Cell 2017, 29, 1373–1387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greenboim-Wainberg, Y.; Maymon, I.; Borochov, R.; Alvarez, J.; Olszewski, N.; Ori, N.; Eshed, Y.; Weiss, D. Cross talk between gibberellin and cytokinin: The Arabidopsis GA response inhibitor SPINDLY plays a positive role in cytokinin signaling. Plant Cell 2005, 17, 92–102. [Google Scholar] [CrossRef] [Green Version]
- Pautler, M.; Eveland, A.L.; LaRue, T.; Yang, F.; Weeks, R.; Lunde, C.; Je, B.I.; Meeley, R.; Komatsu, M.; Vollbrecht, E. FASCIATED EAR4 encodes a bZIP transcription factor that regulates shoot meristem size in maize. Plant Cell 2015, 27, 104–120. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Bui, H.T.; Pautler, M.; Llaca, V.; Johnston, R.; Lee, B.-H.; Kolbe, A.; Sakai, H.; Jackson, D. A maize glutaredoxin gene, abphyl2, regulates shoot meristem size and phyllotaxy. Plant Cell 2015, 27, 121–131. [Google Scholar] [CrossRef] [Green Version]
- Yang, R.; Xu, F.; Wang, Y.; Zhong, W.; Dong, L.; Shi, Y.; Tang, T.; Sheng, H.; Jackson, D.; Yang, F. Glutaredoxins regulate maize inflorescence meristem development via redox control of TGA transcriptional activity. Nat. Plants 2021, 7, 1589–1601. [Google Scholar] [CrossRef]
- Zeng, J.; Dong, Z.; Wu, H.; Tian, Z.; Zhao, Z. Redox regulation of plant stem cell fate. EMBO J. 2017, 36, 2844–2855. [Google Scholar] [CrossRef]
- Chaubal, R.; Anderson, J.R.; Trimnell, M.R.; Fox, T.W.; Albertsen, M.C.; Bedinger, P. The transformation of anthers in the msca1 mutant of maize. Planta 2003, 216, 778–788. [Google Scholar] [CrossRef]
- An, X.; Ma, B.; Duan, M.; Dong, Z.; Liu, R.; Yuan, D.; Hou, Q.; Wu, S.; Zhang, D.; Liu, D. Molecular regulation of ZmMs7 required for maize male fertility and development of a dominant male-sterility system in multiple species. Proc. Natl. Acad. Sci. USA 2020, 117, 23499–23509. [Google Scholar] [CrossRef]
- Wan, X.; Wu, S.; Li, X. Breeding with dominant genic male-sterility genes to boost crop grain yield in the post-heterosis utilization era. Mol. Plant 2021, 14, 531–534. [Google Scholar] [CrossRef] [PubMed]
- Igamberdiev, A.U.; Eprintsev, A.T. Organic acids: The pools of fixed carbon involved in redox regulation and energy balance in higher plants. Front. Plant Sci. 2016, 7, 1042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Igamberdiev, A.U.; Bykova, N.V. Role of organic acids in the integration of cellular redox metabolism and mediation of redox signalling in photosynthetic tissues of higher plants. Free Radic. Biol. Med. 2018, 122, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Luo, L.; Xu, J.; Xin, P.; Guo, H.; Wu, J.; Bai, L.; Wang, G.; Chu, J.; Zuo, J. Malate transported from chloroplast to mitochondrion triggers production of ROS and PCD in Arabidopsis thaliana. Cell Res. 2018, 28, 448–461. [Google Scholar] [CrossRef] [Green Version]
- Selinski, J.; Scheibe, R. Malate valves: Old shuttles with new perspectives. Plant Biol. 2019, 21, 21–30. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Galli, M.; Liu, X.; Federici, S.; Buck, A.; Cody, J.; Labra, M.; Gallavotti, A. NEEDLE1 encodes a mitochondria localized ATP-dependent metalloprotease required for thermotolerant maize growth. Proc. Natl. Acad. Sci. USA 2019, 116, 19736–19742. [Google Scholar] [CrossRef] [Green Version]
- Vollbrecht, E.; Springer, P.S.; Goh, L.; Buckler IV, E.S.; Martienssen, R. Architecture of floral branch systems in maize and related grasses. Nature 2005, 436, 1119–1126. [Google Scholar] [CrossRef]
- Bortiri, E.; Chuck, G.; Vollbrecht, E.; Rocheford, T.; Martienssen, R.; Hake, S. Ramosa2 encodes a LATERAL ORGAN BOUNDARY domain protein that determines the fate of stem cells in branch meristems of maize. Plant Cell 2006, 18, 574–585. [Google Scholar] [CrossRef] [Green Version]
- Satoh-Nagasawa, N.; Nagasawa, N.; Malcomber, S.; Sakai, H.; Jackson, D. A trehalose metabolic enzyme controls inflorescence architecture in maize. Nature 2006, 441, 227–230. [Google Scholar] [CrossRef]
- Gallavotti, A.; Long, J.A.; Stanfield, S.; Yang, X.; Jackson, D.; Vollbrecht, E.; Schmidt, R.J. The control of axillary meristem fate in the maize ramosa pathway. Development 2010, 137, 2849–2856. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Sun, W.; Singh, R.; Zheng, Y.; Cao, Z.; Li, M.; Lunde, C.; Hake, S.; Zhang, Z. GRF-interacting factor1 regulates shoot architecture and meristem determinacy in maize. Plant Cell 2018, 30, 360–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Zheng, Y.; Cui, D.; Du, Y.; Zhang, D.; Sun, W.; Du, H.; Zhang, Z. GIF1 controls ear inflorescence architecture and floral development by regulating key genes in hormone biosynthesis and meristem determinacy in maize. BMC Plant Biol. 2022, 22, 127. [Google Scholar] [CrossRef] [PubMed]
- Whipple, C.J.; Kebrom, T.H.; Weber, A.L.; Yang, F.; Hall, D.; Meeley, R.; Schmidt, R.; Doebley, J.; Brutnell, T.P.; Jackson, D.P. Grassy tillers1 promotes apical dominance in maize and responds to shade signals in the grasses. Proc. Natl. Acad. Sci. USA 2011, 108, E506–E512. [Google Scholar] [CrossRef] [PubMed]
- Klein, H.; Gallagher, J.; Demesa-Arevalo, E.; Abraham-Juárez, M.J.; Heeney, M.; Feil, R.; Lunn, J.E.; Xiao, Y.; Chuck, G.; Whipple, C. Recruitment of an ancient branching program to suppress carpel development in maize flowers. Proc. Natl. Acad. Sci. USA 2022, 119, e2115871119. [Google Scholar] [CrossRef] [PubMed]
- Kebrom, T.H.; Burson, B.L.; Finlayson, S.A. Phytochrome B represses Teosinte Branched1 expression and induces sorghum axillary bud outgrowth in response to light signals. Plant Physiol. 2006, 140, 1109–1117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chuck, G.; Whipple, C.; Jackson, D.; Hake, S. The maize SBP-box transcription factor encoded by tasselsheath4 regulates bract development and the establishment of meristem boundaries. Development 2010, 137, 1243–1250. [Google Scholar] [CrossRef] [Green Version]
- Chuck, G.S.; Brown, P.J.; Meeley, R.; Hake, S. Maize SBP-box transcription factors unbranched2 and unbranched3 affect yield traits by regulating the rate of lateral primordia initiation. Proc. Natl. Acad. Sci. USA 2014, 111, 18775–18780. [Google Scholar] [CrossRef]
- Wang, H.; Nussbaum-Wagler, T.; Li, B.; Zhao, Q.; Vigouroux, Y.; Faller, M.; Bomblies, K.; Lukens, L.; Doebley, J.F. The origin of the naked grains of maize. Nature 2005, 436, 714–719. [Google Scholar] [CrossRef] [Green Version]
- Chuck, G.; Cigan, A.M.; Saeteurn, K.; Hake, S. The heterochronic maize mutant Corngrass1 results from overexpression of a tandem microRNA. Nat. Genet. 2007, 39, 544–549. [Google Scholar] [CrossRef]
- Du, Y.; Liu, L.; Peng, Y.; Li, M.; Li, Y.; Liu, D.; Li, X.; Zhang, Z. UNBRANCHED3 expression and inflorescence development is mediated by UNBRANCHED2 and the distal enhancer, KRN4, in maize. PLoS Genet. 2020, 16, e1008764. [Google Scholar] [CrossRef]
- Chuck, G.; Muszynski, M.; Kellogg, E.; Hake, S.; Schmidt, R.J. The control of spikelet meristem identity by the branched silkless1 gene in maize. Science 2002, 298, 1238–1241. [Google Scholar] [CrossRef] [PubMed]
- Chuck, G.; Meeley, R.B.; Hake, S. The control of maize spikelet meristem fate by theAPETALA2-like gene indeterminate spikelet1. Genes Dev. 1998, 12, 1145–1154. [Google Scholar] [CrossRef]
- Chuck, G.; Meeley, R.; Hake, S. Floral meristem initiation and meristem cell fate are regulated by the maize AP2 genes ids1 and sid1. Development 2008, 135, 3013–3019. [Google Scholar] [CrossRef] [Green Version]
- Du, Y.; Lunde, C.; Li, Y.; Jackson, D.; Hake, S.; Zhang, Z. Gene duplication at the Fascicled ear1 locus controls the fate of inflorescence meristem cells in maize. Proc. Natl. Acad. Sci. USA 2021, 118, e2019218118. [Google Scholar] [CrossRef]
- Clark, R.M.; Wagler, T.N.; Quijada, P.; Doebley, J. A distant upstream enhancer at the maize domestication gene tb1 has pleiotropic effects on plant and inflorescent architecture. Nat. Genet. 2006, 38, 594–597. [Google Scholar] [CrossRef] [PubMed]
- Ookawa, T.; Hobo, T.; Yano, M.; Murata, K.; Ando, T.; Miura, H.; Asano, K.; Ochiai, Y.; Ikeda, M.; Nishitani, R. New approach for rice improvement using a pleiotropic QTL gene for lodging resistance and yield. Nat. Commun. 2010, 1, 132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, A.; Hao, C.; Wang, Z.; Geng, S.; Jia, M.; Wang, F.; Han, X.; Kong, X.; Yin, L.; Tao, S. Wheat breeding history reveals synergistic selection of pleiotropic genomic sites for plant architecture and grain yield. Mol. Plant 2022, 15, 504–519. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Zhou, Y.; Zhang, X.; Zhang, J.; Zhou, Z.; Weng, J.; Li, X.; Wang, Z. Natural variation in the THICK TASSEL DWARF1 (TD1) gene in the regulation of maize (Zea mays L.) ear-related traits. Breed. Sci. 2019, 69, 323–331. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; An, X.; Li, Z.; Yan, T.; Zhu, T.; Xie, K.; Liu, S.; Hou, Q.; Zhao, L.; Wu, S. CRISPR/Cas9-based discovery of maize transcription factors regulating male sterility and their functional conservation in plants. Plant Biotechnol. J. 2021, 19, 1769–1784. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, S.; Jiang, Y.; Yan, T.; Fang, C.; Hou, Q.; Wu, S.; Xie, K.; An, X.; Wan, X. Use of CRISPR/Cas9-Based Gene Editing to Simultaneously Mutate Multiple Homologous Genes Required for Pollen Development and Male Fertility in Maize. Cells 2022, 11, 439. [Google Scholar] [CrossRef]
- Wei, X.; Pu, A.; Liu, Q.; Hou, Q.; Zhang, Y.; An, X.; Long, Y.; Jiang, Y.; Dong, Z.; Wu, S. The bibliometric landscape of gene editing innovation and regulation in the worldwide. Cells 2022, 11, 2682. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Gallagher, J.; Arevalo, E.D.; Chen, R.; Skopelitis, T.; Wu, Q.; Bartlett, M.; Jackson, D. Enhancing grain-yield-related traits by CRISPR–Cas9 promoter editing of maize CLE genes. Nat. Plants 2021, 7, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Hoisington, D.; Melchinger, A. From theory to practice: Marker-assisted selection in maize. In Molecular Marker Systems in Plant Breeding and Crop Improvement; Springer: Berlin/Heidelberg, Germany, 2004; pp. 335–352. [Google Scholar]
- Desta, Z.A.; Ortiz, R. Genomic selection: Genome-wide prediction in plant improvement. Trends Plant Sci. 2014, 19, 592–601. [Google Scholar] [CrossRef] [PubMed]
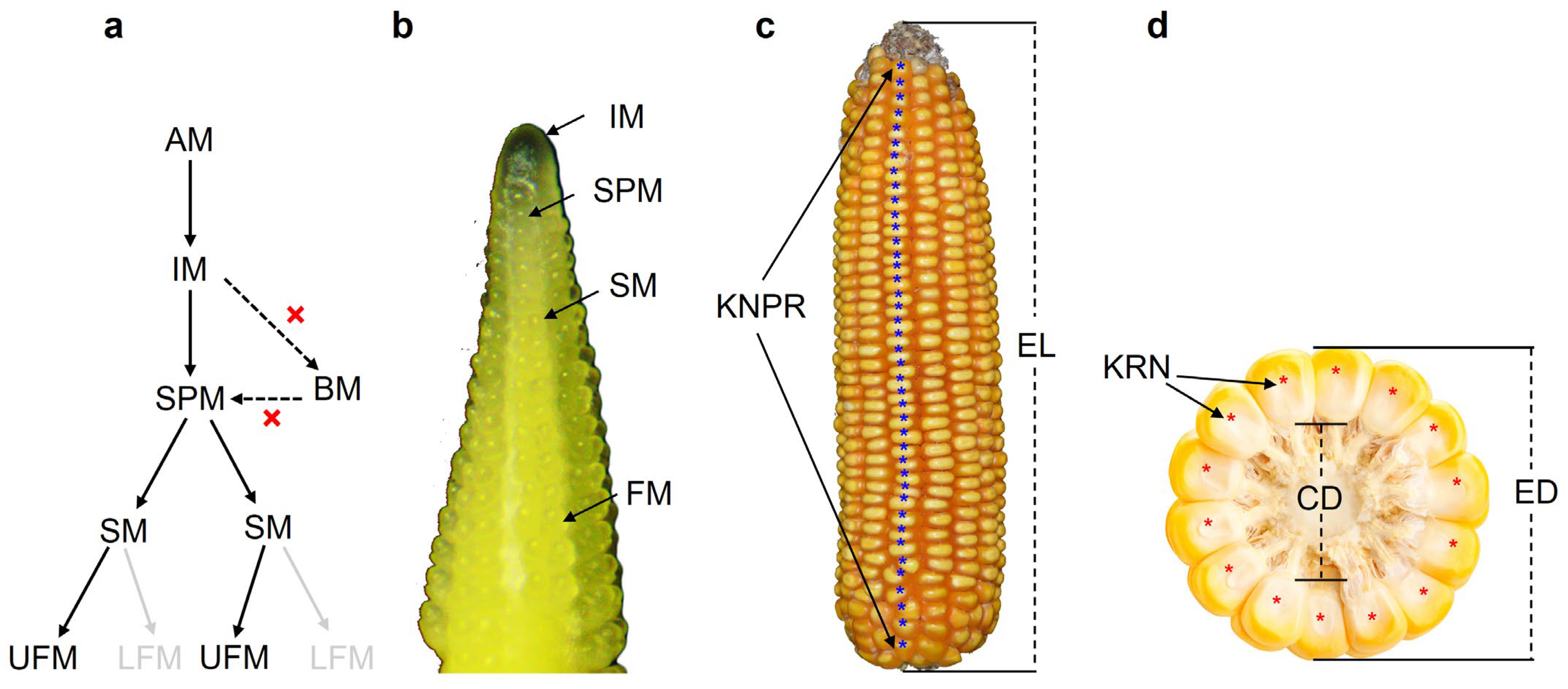


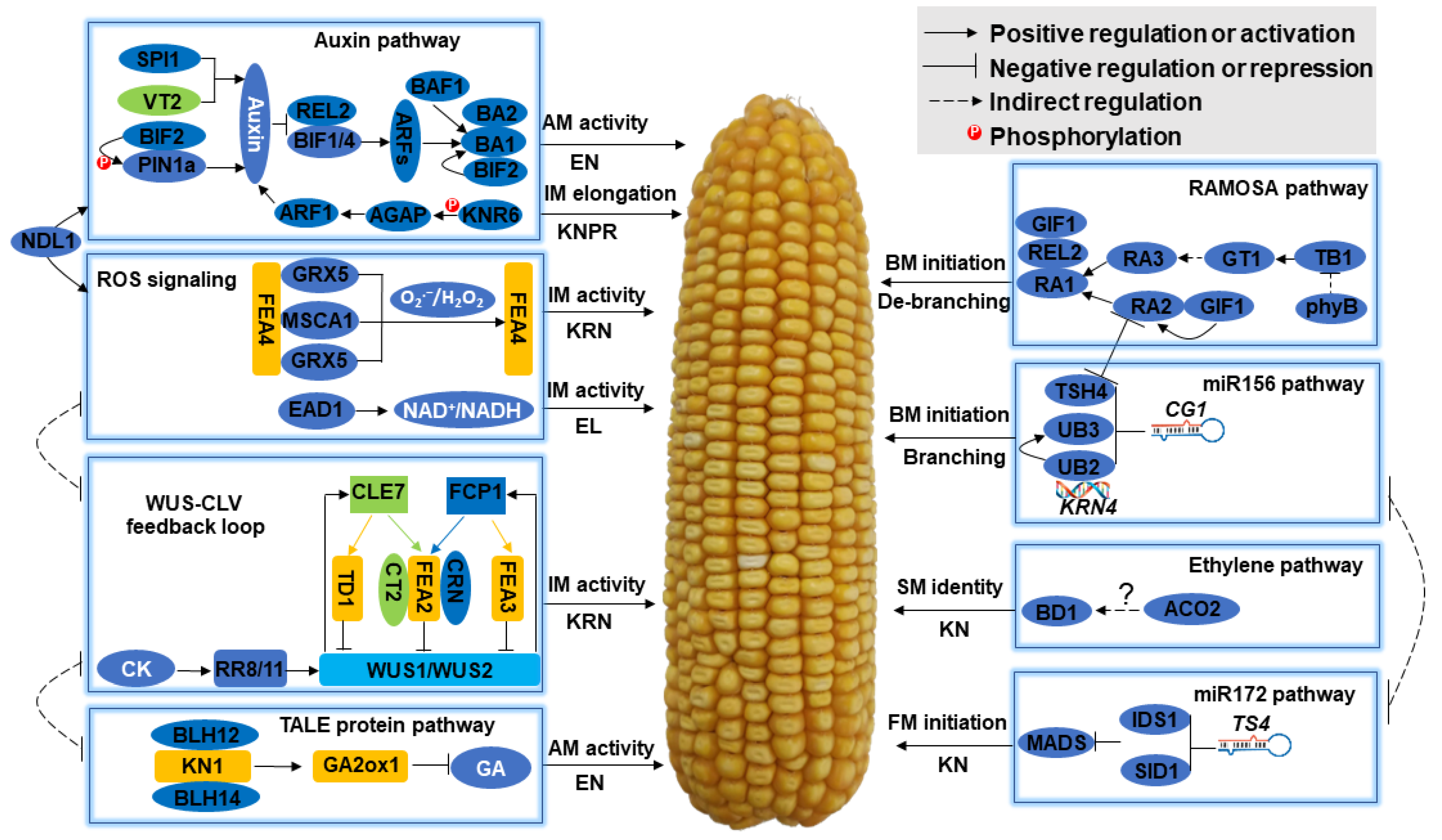

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Chromosome | QTL Cluster | QTN Cluster | Known Gene | ||
|---|---|---|---|---|---|---|
| Start (Mb) | End (Mb) | Start (Mb) | End (Mb) | |||
| 1 | 1 | 293.44 | 304.60 | 297.63 | 298.13 | |
| 2 | 1 | 293.44 | 304.60 | 299.78 | 300.29 | IDS1/KRN1 |
| 3 | 2 | 5.47 | 5.85 | 4.84 | 5.83 | |
| 4 | 2 | 19.86 | 26.25 | 21.48 | 22.02 | |
| 5 | 2 | 19.86 | 26.25 | 23.49 | 24.19 | |
| 6 | 3 | 14.89 | 26.40 | 17.27 | 18.08 | |
| 7 | 3 | 226.09 | 228.09 | 225.85 | 226.54 | |
| 8 | 4 | 195.32 | 239.24 | 204.74 | 205.67 | RKN4, UB3 |
| 9 | 4 | 195.32 | 239.24 | 205.76 | 206.52 | RKN4, UB3 |
| 10 | 5 | 20.41 | 23.71 | 22.80 | 23.30 | |
| 11 | 36.01 | 51.50 | 35.58 | 36.47 | ||
| 12 | 5 | 36.01 | 51.50 | 41.07 | 41.62 | |
| 13 | 5 | 36.01 | 51.50 | 44.21 | 44.71 | |
| 14 | 5 | 60.99 | 67.09 | 65.79 | 66.29 | TD1 |
| 15 | 5 | 141.98 | 184.11 | 146.17 | 146.67 | |
| 16 | 7 | 11.68 | 12.63 | 12.00 | 12.62 | |
| 17 | 7 | 21.96 | 45.32 | 41.96 | 42.46 | SID1 |
| 18 | 8 | 112.72 | 122.82 | 113.29 | 113.98 | |
| 19 | 9 | 27.76 | 29.76 | 28.29 | 29.03 | |
| No. | Chromosome | Pleiotropic Region | Data Resource | Ear Traits | Gene | |
|---|---|---|---|---|---|---|
| Start (Mb) | End (Mb) | |||||
| 1 | 1 | 0.00 | 0.61 | QTN cluster | CD, KRN | |
| 2 | 1 | 1.96 | 2.11 | QTN cluster | KRN, ED, CD | |
| 3 | 1 | 2.50 | 2.68 | QTN cluster | CD, KRN | |
| 4 | 1 | 3.23 | 3.72 | QTN cluster | CD, ED | |
| 5 | 1 | 26.40 | 26.90 | QTN cluster | EL, KRN | |
| 6 | 1 | 28.45 | 28.48 | QTN cluster | EL, CD, KRN | |
| 7 | 1 | 34.89 | 35.55 | QTN cluster/QTL cluster | CD, EL | |
| 8 | 1 | 35.99 | 36.49 | QTN cluster/QTL cluster | EL, ED | |
| 9 | 1 | 42.54 | 43.54 | QTL cluster | EL, KNPR | |
| 10 | 1 | 44.18 | 44.90 | QTL cluster | KWPE, ED | |
| 11 | 1 | 220.28 | 221.93 | QTL cluster | KRN, CD | |
| 12 | 1 | 285.34 | 292.19 | QTL cluster | KRN, KWPE | |
| 13 | 2 | 23.49 | 23.81 | QTN cluster | CD, KRN | |
| 14 | 2 | 65.94 | 66.23 | QTL cluster | KRN, KNPR | BA2 |
| 15 | 2 | 153.51 | 154.97 | QTL cluster | EL, KRN | |
| 16 | 2 | 200.01 | 200.07 | QTL cluster | KRN, ED | |
| 17 | 3 | 2.42 | 3.42 | QTN cluster/QTL cluster | CD, KRN | |
| 18 | 3 | 3.19 | 3.28 | QTL cluster | CD, KRN, EL | |
| 19 | 3 | 9.57 | 10.06 | QTL cluster | EL, KRN | |
| 20 | 3 | 10.44 | 11.10 | QTL cluster | KRN, EL | |
| 21 | 3 | 24.90 | 25.40 | QTN cluster/QTL cluster | KRN, CD | |
| 22 | 3 | 154.92 | 155.42 | QTN cluster/QTL cluster | KRN, ED | |
| 23 | 4 | 0.33 | 0.83 | QTN cluster | CD, KRN | |
| 24 | 4 | 191.57 | 191.82 | QTN cluster/QTL cluster | KRN, ED, EW | |
| 25 | 4 | 215.58 | 216.26 | QTN cluster/QTL cluster | KRN, EL | |
| 26 | 4 | 244.05 | 244.61 | QTN cluster/QTL cluster | KRN, ED | |
| 27 | 4 | 244.74 | 245.54 | QTL cluster | KRN, ED | |
| 28 | 5 | 126.98 | 127.53 | QTN cluster/QTL cluster | EL, KRN | |
| 29 | 5 | 137.90 | 138.57 | QTL cluster | KNPR, EL | |
| 30 | 5 | 145.16 | 145.22 | QTL cluster | CW, KRN, KWPE | |
| 31 | 5 | 164.69 | 166.10 | QTL cluster | KRN, EL | |
| 32 | 5 | 196.69 | 197.81 | QTL cluster | EL, KRN | |
| 33 | 5 | 213.77 | 214.96 | QTL cluster | KRN, EL | |
| 34 | 5 | 216.13 | 216.72 | QTN cluster/QTL cluster | EL, CD | |
| 35 | 8 | 121.24 | 121.27 | QTN cluster/QTL cluster | KRN, CD, EL | |
| 36 | 8 | 167.11 | 167.61 | QTN cluster/QTL cluster | KRN, ED | |
| 37 | 9 | 10.90 | 12.21 | QTL cluster | ED, KRN | |
| 38 | 9 | 13.09 | 14.78 | QTL cluster | ED, KRN | |
| 39 | 10 | 94.76 | 98.77 | QTL cluster | EW, KNPR | ACO2 |
| 40 | 10 | 148.31 | 148.42 | QTN cluster/QTL cluster | KRN, EL | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, Z.; Wang, Y.; Bao, J.; Li, Y.; Yin, Z.; Long, Y.; Wan, X. The Genetic Structures and Molecular Mechanisms Underlying Ear Traits in Maize (Zea mays L.). Cells 2023, 12, 1900. https://doi.org/10.3390/cells12141900
Dong Z, Wang Y, Bao J, Li Y, Yin Z, Long Y, Wan X. The Genetic Structures and Molecular Mechanisms Underlying Ear Traits in Maize (Zea mays L.). Cells. 2023; 12(14):1900. https://doi.org/10.3390/cells12141900
Chicago/Turabian StyleDong, Zhenying, Yanbo Wang, Jianxi Bao, Ya’nan Li, Zechao Yin, Yan Long, and Xiangyuan Wan. 2023. "The Genetic Structures and Molecular Mechanisms Underlying Ear Traits in Maize (Zea mays L.)" Cells 12, no. 14: 1900. https://doi.org/10.3390/cells12141900
APA StyleDong, Z., Wang, Y., Bao, J., Li, Y., Yin, Z., Long, Y., & Wan, X. (2023). The Genetic Structures and Molecular Mechanisms Underlying Ear Traits in Maize (Zea mays L.). Cells, 12(14), 1900. https://doi.org/10.3390/cells12141900