
Crosstalk between Bone and Muscles during Physical Activity

 ,
,  ,
, 
 and
and
Abstract
:
1. Physical Activity and Wellness
2. Progenitor Cells
3. Bone and Muscle Cytokines
4. Extracellular Vesicles Modulation during Physical Activity
5. Interconnected Bone and Muscle Molecular Pathways
6. In Vitro and In Vivo Model to Evaluate the Effects of Physical Activity on the Bone–Muscle Crosstalk
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Karsenty, G.; Mera, P. Molecular bases of the crosstalk between bone and muscle. Bone 2018, 115, 43–49. [Google Scholar]
- Mera, P.; Laue, K.; Ferron, M.; Confavreux, C.; Wei, J.; Galán-Díez, M.; Lacampagne, A.; Mitchell, S.J.; Mattison, J.A.; Chen, Y. Osteocalcin signaling in myofibers is necessary and sufficient for optimum adaptation to exercise. Cell Metab. 2016, 23, 1078–1092. [Google Scholar] [PubMed]
- Whitham, M.; Febbraio, M.A. The ever-expanding myokinome: Discovery challenges and therapeutic implications. Nat. Rev. Drug Discov. 2016, 15, 719–729. [Google Scholar] [PubMed]
- Mahalakshmi, B.; Maurya, N.; Lee, S.-D.; Bharath Kumar, V. Possible neuroprotective mechanisms of physical exercise in neurodegeneration. Int. J. Mol. Sci. 2020, 21, 5895. [Google Scholar] [PubMed]
- Valenti, M.T.; Dalle Carbonare, L.; Dorelli, G.; Mottes, M. Effects of physical exercise on the prevention of stem cells senescence. Stem Cell Rev. Rep. 2020, 16, 33–40. [Google Scholar] [PubMed]
- Kirk, B.; Zanker, J.; Duque, G. Osteosarcopenia: Epidemiology, diagnosis, and treatment—Facts and numbers. J. Cachexia Sarcopenia Muscle 2020, 11, 609–618. [Google Scholar] [PubMed]
- Brunet, A.; Goodell, M.A.; Rando, T.A. Ageing and rejuvenation of tissue stem cells and their niches. Nat. Rev. Mol. Cell Biol. 2023, 24, 45–62. [Google Scholar]
- Durstine, J.L.; Painter, P.; Franklin, B.A.; Morgan, D.; Pitetti, K.H.; Roberts, S.O. Physical activity for the chronically ill and disabled. Sports Med. 2000, 30, 207–219. [Google Scholar]
- White, L.J.; Dressendorfer, R.H. Exercise and multiple sclerosis. Sports Med. 2004, 34, 1077–1100. [Google Scholar]
- Singh, M.A.F. Exercise comes of age: Rationale and recommendations for a geriatric exercise prescription. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2002, 57, M262–M282. [Google Scholar]
- Booth, F.W.; Roberts, C.K.; Laye, M.J. Lack of exercise is a major cause of chronic diseases. Compr. Physiol. 2012, 2, 1143–1211. [Google Scholar] [PubMed]
- Pedersen, B.K.; Saltin, B. Exercise as medicine–evidence for prescribing exercise as therapy in 26 different chronic diseases. Scand. J. Med. Sci. Sports 2015, 25 (Suppl. S3), 1–72. [Google Scholar] [PubMed]
- Tieland, M.; Trouwborst, I.; Clark, B.C. Skeletal muscle performance and ageing. J. Cachexia Sarcopenia Muscle 2018, 9, 3–19. [Google Scholar]
- Cariati, I.; Bonanni, R.; Onorato, F.; Mastrogregori, A.; Rossi, D.; Iundusi, R.; Gasbarra, E.; Tancredi, V.; Tarantino, U. Role of physical activity in bone–muscle crosstalk: Biological aspects and clinical implications. J. Funct. Morphol. Kinesiol. 2021, 6, 55. [Google Scholar] [PubMed]
- Mannino, G.; Russo, C.; Maugeri, G.; Musumeci, G.; Vicario, N.; Tibullo, D.; Giuffrida, R.; Parenti, R.; Lo Furno, D. Adult stem cell niches for tissue homeostasis. J. Cell. Physiol. 2022, 237, 239–257. [Google Scholar] [PubMed]
- Relaix, F.; Rocancourt, D.; Mansouri, A.; Buckingham, M. A Pax3/Pax7-dependent population of skeletal muscle progenitor cells. Nature 2005, 435, 948–953. [Google Scholar] [PubMed]
- Wosczyna, M.N.; Rando, T.A. A muscle stem cell support group: Coordinated cellular responses in muscle regeneration. Dev. Cell 2018, 46, 135–143. [Google Scholar]
- Yin, H.; Price, F.; Rudnicki, M.A. Satellite cells and the muscle stem cell niche. Physiol. Rev. 2013, 93, 23–67. [Google Scholar]
- Salhotra, A.; Shah, H.N.; Levi, B.; Longaker, M.T. Mechanisms of bone development and repair. Nat. Rev. Mol. Cell Biol. 2020, 21, 696–711. [Google Scholar]
- Kim, P.; Park, J.; Lee, D.-J.; Mizuno, S.; Shinohara, M.; Hong, C.P.; Jeong, Y.; Yun, R.; Park, H.; Park, S. Mast4 determines the cell fate of MSCs for bone and cartilage development. Nat. Commun. 2022, 13, 3960. [Google Scholar]
- Carapeto, P.V.; Aguayo-Mazzucato, C. Effects of exercise on cellular and tissue aging. Aging 2021, 13, 14522. [Google Scholar] [CrossRef]
- Kimmel, J.C.; Hwang, A.B.; Scaramozza, A.; Marshall, W.F.; Brack, A.S. Aging induces aberrant state transition kinetics in murine muscle stem cells. Development 2020, 147, dev183855. [Google Scholar] [CrossRef] [PubMed]
- Lombardi, G.; Ziemann, E.; Banfi, G. Physical activity and bone health: What is the role of immune system? A narrative review of the third way. Front. Endocrinol. 2019, 10, 60. [Google Scholar]
- Marędziak, M.; Śmieszek, A.; Chrząstek, K.; Basinska, K.; Marycz, K. Physical activity increases the total number of bone-marrow-derived mesenchymal stem cells, enhances their osteogenic potential, and inhibits their adipogenic properties. Stem Cells Int. 2015, 2015, 379093. [Google Scholar] [CrossRef]
- Hsu, W.-B.; Lin, S.-J.; Hung, J.-S.; Chen, M.-H.; Lin, C.-Y.; Hsu, W.-H.; Hsu, W.-W.R. Effect of resistance training on satellite cells in old mice–a transcriptome study: Implications for sarcopenia. Bone Jt. Res. 2022, 11, 121–133. [Google Scholar] [CrossRef] [PubMed]
- Pugh, J.K.; Faulkner, S.H.; Turner, M.C.; Nimmo, M.A. Satellite cell response to concurrent resistance exercise and high-intensity interval training in sedentary, overweight/obese, middle-aged individuals. Eur. J. Appl. Physiol. 2018, 118, 225–238. [Google Scholar] [CrossRef] [PubMed]
- Caldow, M.K.; Thomas, E.E.; Dale, M.J.; Tomkinson, G.R.; Buckley, J.D.; Cameron-Smith, D. Early myogenic responses to acute exercise before and after resistance training in young men. Physiol. Rep. 2015, 3, e12511. [Google Scholar] [CrossRef]
- Brett, J.O.; Arjona, M.; Ikeda, M.; Quarta, M.; de Morree, A.; Egner, I.M.; Perandini, L.A.; Ishak, H.D.; Goshayeshi, A.; Benjamin, D.I. Exercise rejuvenates quiescent skeletal muscle stem cells in old mice through restoration of Cyclin D1. Nat. Metab. 2020, 2, 307–317. [Google Scholar] [CrossRef]
- Dalle Carbonare, L.; Dorelli, G.; Li Vigni, V.; Minoia, A.; Bertacco, J.; Cheri, S.; Deiana, M.; Innamorati, G.; Cominacini, M.; Tarperi, C. Physical activity modulates miRNAs levels and enhances MYOD expression in myoblasts. Stem Cell Rev. Rep. 2022, 18, 1865–1874. [Google Scholar] [CrossRef]
- Valenti, M.T.; Deiana, M.; Cheri, S.; Dotta, M.; Zamboni, F.; Gabbiani, D.; Schena, F.; Dalle Carbonare, L.; Mottes, M. Physical exercise modulates miR-21-5p, miR-129-5p, miR-378-5p, and miR-188-5p expression in progenitor cells promoting osteogenesis. Cells 2019, 8, 742. [Google Scholar] [CrossRef]
- Hirschfeld, H.; Kinsella, R.; Duque, G. Osteosarcopenia: Where bone, muscle, and fat collide. Osteoporos. Int. 2017, 28, 2781–2790. [Google Scholar] [CrossRef] [PubMed]
- Kawao, N.; Kaji, H. Interactions between muscle tissues and bone metabolism. J. Cell. Biochem. 2015, 116, 687–695. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.W.; White, J.D.; Walker, E.C.; Martin, T.J.; Sims, N.A. Myokines (muscle-derived cytokines and chemokines) including ciliary neurotrophic factor (CNTF) inhibit osteoblast differentiation. Bone 2014, 64, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Di Raimondo, D.; Miceli, G.; Musiari, G.; Tuttolomondo, A.; Pinto, A. New insights about the putative role of myokines in the context of cardiac rehabilitation and secondary cardiovascular prevention. Ann. Transl. Med. 2017, 5, 300. [Google Scholar] [CrossRef] [PubMed]
- Colaianni, G.; Cuscito, C.; Mongelli, T.; Oranger, A.; Mori, G.; Brunetti, G.; Colucci, S.; Cinti, S.; Grano, M. Irisin enhances osteoblast differentiation in vitro. Int. J. Endocrinol. 2014, 2014, 902186. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.; Wu, H.; Lu, J.; Zhang, R.; Shen, X.; Gu, Y.; Shi, C.; Zhang, Y.; Yuan, W. Irisin reduces bone fracture by facilitating osteogenesis and antagonizing TGF-β/Smad signaling in a growing mouse model of osteogenesis imperfecta. J. Orthop. Transl. 2023, 38, 175–189. [Google Scholar] [CrossRef]
- Colaianni, G.; Cuscito, C.; Mongelli, T.; Pignataro, P.; Buccoliero, C.; Liu, P.; Lu, P.; Sartini, L.; Di Comite, M.; Mori, G. The myokine irisin increases cortical bone mass. Proc. Natl. Acad. Sci. USA 2015, 112, 12157–12162. [Google Scholar] [CrossRef]
- Kim, J.A.; Roh, E.; Hong, S.-h.; Lee, Y.-B.; Kim, N.H.; Yoo, H.J.; Seo, J.A.; Kim, N.H.; Kim, S.G.; Baik, S.H. Association of serum sclerostin levels with low skeletal muscle mass: The Korean Sarcopenic Obesity Study (KSOS). Bone 2019, 128, 115053. [Google Scholar] [CrossRef]
- Śliwicka, E.; Cisoń, T.; Pilaczyńska-Szcześniak, Ł.; Ziemba, A.; Straburzyńska-Lupa, A. Effects of marathon race on selected myokines and sclerostin in middle-aged male amateur runners. Sci. Rep. 2021, 11, 1–9. [Google Scholar] [CrossRef]
- Kirk, B.; Feehan, J.; Lombardi, G.; Duque, G. Muscle, bone, and fat crosstalk: The biological role of myokines, osteokines, and adipokines. Curr. Osteoporos. Rep. 2020, 18, 388–400. [Google Scholar] [CrossRef]
- Zhi, X.; Chen, Q.; Song, S.; Gu, Z.; Wei, W.; Chen, H.; Chen, X.; Weng, W.; Zhou, Q.; Cui, J. Myostatin promotes osteoclastogenesis by regulating ccdc50 gene expression and RANKL-induced NF-κB and MAPK pathways. Front. Pharmacol. 2020, 11, 565163. [Google Scholar] [CrossRef]
- Pedersen, B.K.; Steensberg, A.; Schjerling, P. Muscle-derived interleukin-6: Possible biological effects. J. Physiol. 2001, 536, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Collins, C.A.; Olsen, I.; Zammit, P.S.; Heslop, L.; Petrie, A.; Partridge, T.A.; Morgan, J.E. Stem cell function, self-renewal, and behavioral heterogeneity of cells from the adult muscle satellite cell niche. Cell 2005, 122, 289–301. [Google Scholar] [CrossRef] [PubMed]
- Lara-Castillo, N.; Johnson, M.L. Bone-muscle mutual interactions. Curr. Osteoporos. Rep. 2020, 18, 408–421. [Google Scholar] [CrossRef]
- Whitham, M.; Parker, B.L.; Friedrichsen, M.; Hingst, J.R.; Hjorth, M.; Hughes, W.E.; Egan, C.L.; Cron, L.; Watt, K.I.; Kuchel, R.P. Extracellular vesicles provide a means for tissue crosstalk during exercise. Cell Metab. 2018, 27, 237–251.e4. [Google Scholar] [CrossRef]
- Li, S.R.; Man, Q.W.; Gao, X.; Lin, H.; Wang, J.; Su, F.C.; Wang, H.Q.; Bu, L.L.; Liu, B.; Chen, G. Tissue-derived extracellular vesicles in cancers and non-cancer diseases: Present and future. J. Extracell. Vesicles 2021, 10, e12175. [Google Scholar] [CrossRef]
- Svenningsen, P.; Sabaratnam, R.; Jensen, B.L. Urinary extracellular vesicles: Origin, role as intercellular messengers and biomarkers; efficient sorting and potential treatment options. Acta Physiol. 2020, 228, e13346. [Google Scholar] [CrossRef]
- Van Rooij, E.; Olson, E.N. MicroRNAs: Powerful new regulators of heart disease and provocative therapeutic targets. J. Clin. Investig. 2007, 117, 2369–2376. [Google Scholar]
- Fochi, S.; Giuriato, G.; De Simone, T.; Gomez-Lira, M.; Tamburin, S.; Del Piccolo, L.; Schena, F.; Venturelli, M.; Romanelli, M.G. Regulation of microRNAs in satellite cell renewal, muscle function, sarcopenia and the role of exercise. Int. J. Mol. Sci. 2020, 21, 6732. [Google Scholar] [CrossRef] [PubMed]
- Qin, W.; Dallas, S.L. Exosomes and extracellular RNA in muscle and bone aging and crosstalk. Curr. Osteoporos. Rep. 2019, 17, 548–559. [Google Scholar] [CrossRef]
- Steensberg, A.; Febbraio, M.A.; Osada, T.; Schjerling, P.; Van Hall, G.; Saltin, B.; Pedersen, B.K. Interleukin-6 production in contracting human skeletal muscle is influenced by pre-exercise muscle glycogen content. J. Physiol. 2001, 537, 633–639. [Google Scholar] [CrossRef] [PubMed]
- He, C.; He, W.; Hou, J.; Chen, K.; Huang, M.; Yang, M.; Luo, X.; Li, C. Bone and muscle crosstalk in aging. Front. Cell Dev. Biol. 2020, 8, 585644. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Cui, Y.; Luan, J.; Zhou, X.; Li, H.; Han, J. Exosomes from C2C12 myoblasts enhance osteogenic differentiation of MC3T3-E1 pre-osteoblasts by delivering miR-27a-3p. Biochem. Biophys. Res. Commun. 2018, 498, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Saito, S.; Saito, W.; Saito, M.; Hashimoto, Y.; Mori, S.; Noda, K.; Namba, K.; Ishida, S. Acute zonal occult outer retinopathy in Japanese patients: Clinical features, visual function, and factors affecting visual function. PLoS ONE 2015, 10, e0125133. [Google Scholar] [CrossRef]
- Fulzele, S.; Mendhe, B.; Khayrullin, A.; Johnson, M.; Kaiser, H.; Liu, Y.; Isales, C.M.; Hamrick, M.W. Muscle-derived miR-34a increases with age in circulating extracellular vesicles and induces senescence of bone marrow stem cells. Aging 2019, 11, 1791. [Google Scholar] [CrossRef]
- Zheng, Y.; Kong, J.; Li, Q.; Wang, Y.; Li, J. Role of miRNAs in skeletal muscle aging. Clin. Interv. Aging 2018, 2407–2419. [Google Scholar] [CrossRef]
- Kukreti, H.; Amuthavalli, K. MicroRNA-34a causes ceramide accumulation and effects insulin signaling pathway by targeting ceramide kinase (CERK) in aging skeletal muscle. J. Cell. Biochem. 2020, 121, 3070–3089. [Google Scholar] [CrossRef]
- Chen, X.; Huang, Z.; Chen, D.; Yang, T.; Liu, G. Role of micro RNA-27a in myoblast differentiation. Cell Biol. Int. 2014, 38, 266–271. [Google Scholar] [CrossRef]
- Huang, Z.; Chen, X.; Yu, B.; He, J.; Chen, D. MicroRNA-27a promotes myoblast proliferation by targeting myostatin. Biochem. Biophys. Res. Commun. 2012, 423, 265–269. [Google Scholar] [CrossRef]
- Sabaratnam, R.; Wojtaszewski, J.F.; Højlund, K. Factors mediating exercise-induced organ crosstalk. Acta Physiol. 2022, 234, e13766. [Google Scholar] [CrossRef]
- Castaño, C.; Mirasierra, M.; Vallejo, M.; Novials, A.; Párrizas, M. Delivery of muscle-derived exosomal miRNAs induced by HIIT improves insulin sensitivity through down-regulation of hepatic FoxO1 in mice. Proc. Natl. Acad. Sci. USA 2020, 117, 30335–30343. [Google Scholar] [CrossRef]
- Pietrangelo, T.; Bondi, D.; Kinel, E.; Verratti, V. The bottom-up rise strength transfer in elderly after endurance and resistance training: The BURST. Front. Physiol. 2019, 9, 1944. [Google Scholar] [CrossRef]
- Judex, S.; Carlson, K.J. Is bone’s response to mechanical signals dominated by gravitational loading? Med. Sci. Sports Exerc. 2009, 41, 2037–2043. [Google Scholar] [CrossRef] [PubMed]
- Hewison, M.; Adams, J.S. Vitamin D insufficiency and skeletal development in utero. J. Bone Miner. Res. 2010, 25, 11. [Google Scholar] [CrossRef] [PubMed]
- Zymbal, V.; Baptista, F.; Letuchy, E.M.; Janz, K.F.; Levy, S.M. Mediating effect of muscle on the relationship of physical activity and bone. Med. Sci. Sports Exerc. 2019, 51, 202. [Google Scholar] [CrossRef]
- Tyrovola, J.B. The “Mechanostat” Principle and the Osteoprotegerin—OPG/RANKL/RANK System PART II. The Role of the Hypothalamic—Pituitary Axis. J. Cell. Biochem. 2017, 118, 962–966. [Google Scholar] [CrossRef] [PubMed]
- Lombardi, G.; Sanchis-Gomar, F.; Perego, S.; Sansoni, V.; Banfi, G. Implications of exercise-induced adipo-myokines in bone metabolism. Endocrine 2016, 54, 284–305. [Google Scholar] [CrossRef]
- Bouviere, J.; Fortunato, R.S.; Dupuy, C.; Werneck-de-Castro, J.P.; Carvalho, D.P.; Louzada, R.A. Exercise-stimulated ROS sensitive signaling pathways in skeletal muscle. Antioxidants 2021, 10, 537. [Google Scholar] [CrossRef]
- Louzada, R.A.; Bouviere, J.; Matta, L.P.; Werneck-de-Castro, J.P.; Dupuy, C.; Carvalho, D.P.; Fortunato, R.S. Redox signaling in widespread health benefits of exercise. Antioxid. Redox Signal. 2020, 33, 745–760. [Google Scholar] [CrossRef]
- Powers, S.K.; Radak, Z.; Ji, L.L. Exercise-induced oxidative stress: Past, present and future. J. Physiol. 2016, 594, 5081–5092. [Google Scholar] [CrossRef]
- Hargreaves, M.; Spriet, L.L. Skeletal muscle energy metabolism during exercise. Nat. Metab. 2020, 2, 817–828. [Google Scholar] [CrossRef] [PubMed]
- Brooks, G.A. The science and translation of lactate shuttle theory. Cell Metab. 2018, 27, 757–785. [Google Scholar] [CrossRef] [PubMed]
- Summer, R.; Shaghaghi, H.; Schriner, D.; Roque, W.; Sales, D.; Cuevas-Mora, K.; Desai, V.; Bhushan, A.; Ramirez, M.I.; Romero, F. Activation of the mTORC1/PGC-1 axis promotes mitochondrial biogenesis and induces cellular senescence in the lung epithelium. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2019, 316, L1049–L1060. [Google Scholar] [CrossRef] [PubMed]
- Koh, J.-H.; Pataky, M.W.; Dasari, S.; Klaus, K.A.; Vuckovic, I.; Ruegsegger, G.N.; Kumar, A.P.; Robinson, M.M.; Nair, K.S. Enhancement of anaerobic glycolysis–a role of PGC-1α4 in resistance exercise. Nat. Commun. 2022, 13, 2324. [Google Scholar] [CrossRef]
- Akimoto, T.; Ribar, T.J.; Williams, R.S.; Yan, Z. Skeletal muscle adaptation in response to voluntary running in Ca2+/calmodulin-dependent protein kinase IV-deficient mice. Am. J. Physiol.-Cell Physiol. 2004, 287, C1311–C1319. [Google Scholar] [CrossRef]
- Akimoto, T.; Pohnert, S.C.; Li, P.; Zhang, M.; Gumbs, C.; Rosenberg, P.B.; Williams, R.S.; Yan, Z. Exercise stimulates Pgc-1α transcription in skeletal muscle through activation of the p38 MAPK pathway. J. Biol. Chem. 2005, 280, 19587–19593. [Google Scholar] [CrossRef]
- Jin, X.J.; Chen, X.J.; Zhang, Z.F.; Hu, W.S.; Ou, R.Y.; Li, S.; Xue, J.S.; Chen, L.L.; Hu, Y.; Zhu, H. Long noncoding RNA SNHG12 promotes the progression of cervical cancer via modulating miR-125b/STAT3 axis. J. Cell. Physiol. 2019, 234, 6624–6632. [Google Scholar] [CrossRef]
- Ruas, J.L.; White, J.P.; Rao, R.R.; Kleiner, S.; Brannan, K.T.; Harrison, B.C.; Greene, N.P.; Wu, J.; Estall, J.L.; Irving, B.A. A PGC-1α isoform induced by resistance training regulates skeletal muscle hypertrophy. Cell 2012, 151, 1319–1331. [Google Scholar] [CrossRef]
- Halling, J.F.; Ringholm, S.; Olesen, J.; Prats, C.; Pilegaard, H. Exercise training protects against aging-induced mitochondrial fragmentation in mouse skeletal muscle in a PGC-1α dependent manner. Exp. Gerontol. 2017, 96, 1–6. [Google Scholar] [CrossRef]
- Ding, Y.; Yang, H.; Wang, Y.; Chen, J.; Ji, Z.; Sun, H. Sirtuin 3 is required for osteogenic differentiation through maintenance of PGC-1ɑ-SOD2-mediated regulation of mitochondrial function. Int. J. Biol. Sci. 2017, 13, 254. [Google Scholar] [CrossRef]
- Jing, E.; Emanuelli, B.; Hirschey, M.D.; Boucher, J.; Lee, K.Y.; Lombard, D.; Verdin, E.M.; Kahn, C.R. Sirtuin-3 (Sirt3) regulates skeletal muscle metabolism and insulin signaling via altered mitochondrial oxidation and reactive oxygen species production. Proc. Natl. Acad. Sci. USA 2011, 108, 14608–14613. [Google Scholar] [CrossRef] [PubMed]
- Widiastuti, I.A.E.; Arsyad, A.; Idris, I.; Patellongi, I.; Kadriyan, H.; Buanayuda, G.W.; Sari, D.P.; Rosyidi, R.M. Exercise adaptations and TGF-β1 levels in recreational cyclists. Ann. Med. Surg. 2021, 70, 102872. [Google Scholar] [CrossRef] [PubMed]
- Iaquinta, M.R.; Lanzillotti, C.; Mazziotta, C.; Bononi, I.; Frontini, F.; Mazzoni, E.; Oton-Gonzalez, L.; Rotondo, J.C.; Torreggiani, E.; Tognon, M. The role of microRNAs in the osteogenic and chondrogenic differentiation of mesenchymal stem cells and bone pathologies. Theranostics 2021, 11, 6573. [Google Scholar] [CrossRef] [PubMed]
- Marques-Carvalho, A.; Kim, H.-N.; Almeida, M. The role of reactive oxygen species in bone cell physiology and pathophysiology. Bone Rep. 2023, 101664. [Google Scholar] [CrossRef]
- Mens, M.M.; Ghanbari, M. Cell cycle regulation of stem cells by microRNAs. Stem Cell Rev. Rep. 2018, 14, 309–322. [Google Scholar] [CrossRef]
- Zhang, D.; Ran, J.; Li, J.; Yu, C.; Cui, Z.; Amevor, F.K.; Wang, Y.; Jiang, X.; Qiu, M.; Du, H. miR-21-5p regulates the proliferation and differentiation of skeletal muscle satellite cells by targeting KLF3 in chicken. Genes 2021, 12, 814. [Google Scholar] [CrossRef]
- Peng, Y.; Xu, M.; Dou, M.; Xin’E, S.; Yang, G.; Li, X. MicroRNA-129-5p inhibits C2C12 myogenesis and represses slow fiber gene expression in vitro. Am. J. Physiol.-Cell Physiol. 2021, 320, C1031–C1041. [Google Scholar] [CrossRef]
- Carrer, M.; Liu, N.; Grueter, C.E.; Williams, A.H.; Frisard, M.I.; Hulver, M.W.; Bassel-Duby, R.; Olson, E.N. Control of mitochondrial metabolism and systemic energy homeostasis by microRNAs 378 and 378. Proc. Natl. Acad. Sci. USA 2012, 109, 15330–15335. [Google Scholar] [CrossRef]
- Dreher, S.I.; Höckele, S.; Huypens, P.; Irmler, M.; Hoffmann, C.; Jeske, T.; Hastreiter, M.; Moller, A.; Birkenfeld, A.L.; Häring, H.-U. TGF-β Induction of miR-143/145 Is Associated to Exercise Response by Influencing Differentiation and Insulin Signaling Molecules in Human Skeletal Muscle. Cells 2021, 10, 3443. [Google Scholar] [CrossRef]
- Wang, R.; Zhang, H.; Ding, W.; Fan, Z.; Ji, B.; Ding, C.; Ji, F.; Tang, H. miR-143 promotes angiogenesis and osteoblast differentiation by targeting HDAC7. Cell Death Dis. 2020, 11, 179. [Google Scholar] [CrossRef]
- Ehsan, M.; Khan, R.; Wakefield, D.; Qureshi, A.; Murray, L.; ZuWallack, R.; Leidy, N.K. A longitudinal study evaluating the effect of exacerbations on physical activity in patients with chronic obstructive pulmonary disease. Ann. Am. Thorac. Soc. 2013, 10, 559–564. [Google Scholar] [CrossRef]
- Barbosa, C.C.L.; da Costa, J.C.; Romanzini, C.L.P.; Batista, M.B.; Blasquez-Shigaki, G.; Fernandes, R.A.; Martinho, D.V.; Oliveira, T.; Ribeiro, L.P.; Coelho-e-Silva, M.J. Interrelationship between muscle fitness in childhood and bone mineral density in adulthood: Mediation analysis of muscle fitness in adulthood. BMC Public Health 2023, 23, 1–10. [Google Scholar]
- Valenti, M.T.; Braggio, M.; Minoia, A.; Dorelli, G.; Bertacco, J.; Bertoldo, F.; Cominacini, M.; De Simone, T.; Romanelli, M.G.; Bhandary, L. Effects of a 4400 km ultra-cycling non-competitive race and related training on body composition and circulating progenitors differentiation. J. Transl. Med. 2022, 20, 397. [Google Scholar] [CrossRef] [PubMed]
- Myers, M.J.; Shepherd, D.L.; Durr, A.J.; Stanton, D.S.; Mohamed, J.S.; Hollander, J.M.; Alway, S.E. The role of SIRT1 in skeletal muscle function and repair of older mice. J. Cachexia Sarcopenia Muscle 2019, 10, 929–949. [Google Scholar] [CrossRef]
- Houtkooper, R.H.; Pirinen, E.; Auwerx, J. Sirtuins as regulators of metabolism and healthspan. Nat. Rev. Mol. Cell Biol. 2012, 13, 225–238. [Google Scholar] [CrossRef]
- Shen, K.; Liu, X.; Qin, H.; Chai, Y.; Wang, L.; Yu, B. HA-g-CS Implant and Moderate-intensity Exercise Stimulate Subchondral Bone Remodeling and Promote Repair of Osteochondral Defects in Mice. Int. J. Med. Sci. 2021, 18, 3808. [Google Scholar] [CrossRef] [PubMed]
- Fry, C.S.; Lee, J.D.; Mula, J.; Kirby, T.J.; Jackson, J.R.; Liu, F.; Yang, L.; Mendias, C.L.; Dupont-Versteegden, E.E.; McCarthy, J.J. Inducible depletion of satellite cells in adult, sedentary mice impairs muscle regenerative capacity without affecting sarcopenia. Nat. Med. 2015, 21, 76–80. [Google Scholar] [CrossRef] [PubMed]
- Englund, D.A.; Murach, K.A.; Dungan, C.M.; Figueiredo, V.C.; Vechetti Jr, I.J.; Dupont-Versteegden, E.E.; McCarthy, J.J.; Peterson, C.A. Depletion of resident muscle stem cells negatively impacts running volume, physical function, and muscle fiber hypertrophy in response to lifelong physical activity. Am. J. Physiol.-Cell Physiol. 2020, 318, C1178–C1188. [Google Scholar] [CrossRef]
- Shochat, C.; Wang, Z.; Mo, C.; Nelson, S.; Donaka, R.; Huang, J.; Karasik, D.; Brotto, M. Deletion of SREBF1, a functional bone-muscle pleiotropic gene, alters bone density and lipid signaling in zebrafish. Endocrinology 2021, 162, bqaa189. [Google Scholar] [CrossRef]
- DePasquale, C.; Franklin, K.; Jia, Z.; Jhaveri, K.; Buderman, F.E. The effects of exploratory behavior on physical activity in a common animal model of human disease, zebrafish (Danio rerio). Front. Behav. Neurosci. 2022, 16, 1020837. [Google Scholar] [CrossRef]
- Carter, S.; Solomon, T.P. In vitro experimental models for examining the skeletal muscle cell biology of exercise: The possibilities, challenges and future developments. Pflügers Arch.-Eur. J. Physiol. 2019, 471, 413–429. [Google Scholar]
- Hoppeler, H. Molecular networks in skeletal muscle plasticity. J. Exp. Biol. 2016, 219, 205–213. [Google Scholar] [CrossRef]
- Nikolić, N.; Skaret Bakke, S.; Tranheim Kase, E.; Rudberg, I.; Flo Halle, I.; Rustan, A.C.; Thoresen, G.H.; Aas, V. Electrical pulse stimulation of cultured human skeletal muscle cells as an in vitro model of exercise. PLoS ONE 2012, 7, e33203. [Google Scholar] [CrossRef] [PubMed]
- Nedachi, T.; Fujita, H.; Kanzaki, M. Contractile C2C12 myotube model for studying exercise-inducible responses in skeletal muscle. Am. J. Physiol.-Endocrinol. Metab. 2008, 295, E1191–E1204. [Google Scholar] [CrossRef] [PubMed]
- Dalle Carbonare, L.; Mottes, M.; Cheri, S.; Deiana, M.; Zamboni, F.; Gabbiani, D.; Schena, F.; Salvagno, G.; Lippi, G.; Valenti, M.T. Increased gene expression of RUNX2 and SOX9 in mesenchymal circulating progenitors is associated with autophagy during physical activity. Oxidative Med. Cell. Longev. 2019, 2019, 8426259. [Google Scholar] [CrossRef] [PubMed]
- Turnbull, G.; Clarke, J.; Picard, F.; Riches, P.; Jia, L.; Han, F.; Li, B.; Shu, W. 3D bioactive composite scaffolds for bone tissue engineering. Bioact. Mater. 2018, 3, 278–314. [Google Scholar] [CrossRef]
- Phelps, J.; Leonard, C.; Shah, S.; Krawetz, R.; Hart, D.A.; Duncan, N.A.; Sen, A. Production of mesenchymal progenitor cell-derived extracellular vesicles in suspension bioreactors for use in articular cartilage repair. Stem Cells Transl. Med. 2022, 11, 73–87. [Google Scholar] [CrossRef] [PubMed]
- Ronaldson-Bouchard, K.; Teles, D.; Yeager, K.; Tavakol, D.N.; Zhao, Y.; Chramiec, A.; Tagore, S.; Summers, M.; Stylianos, S.; Tamargo, M. A multi-organ chip with matured tissue niches linked by vascular flow. Nat. Biomed. Eng. 2022, 6, 351–371. [Google Scholar] [CrossRef]



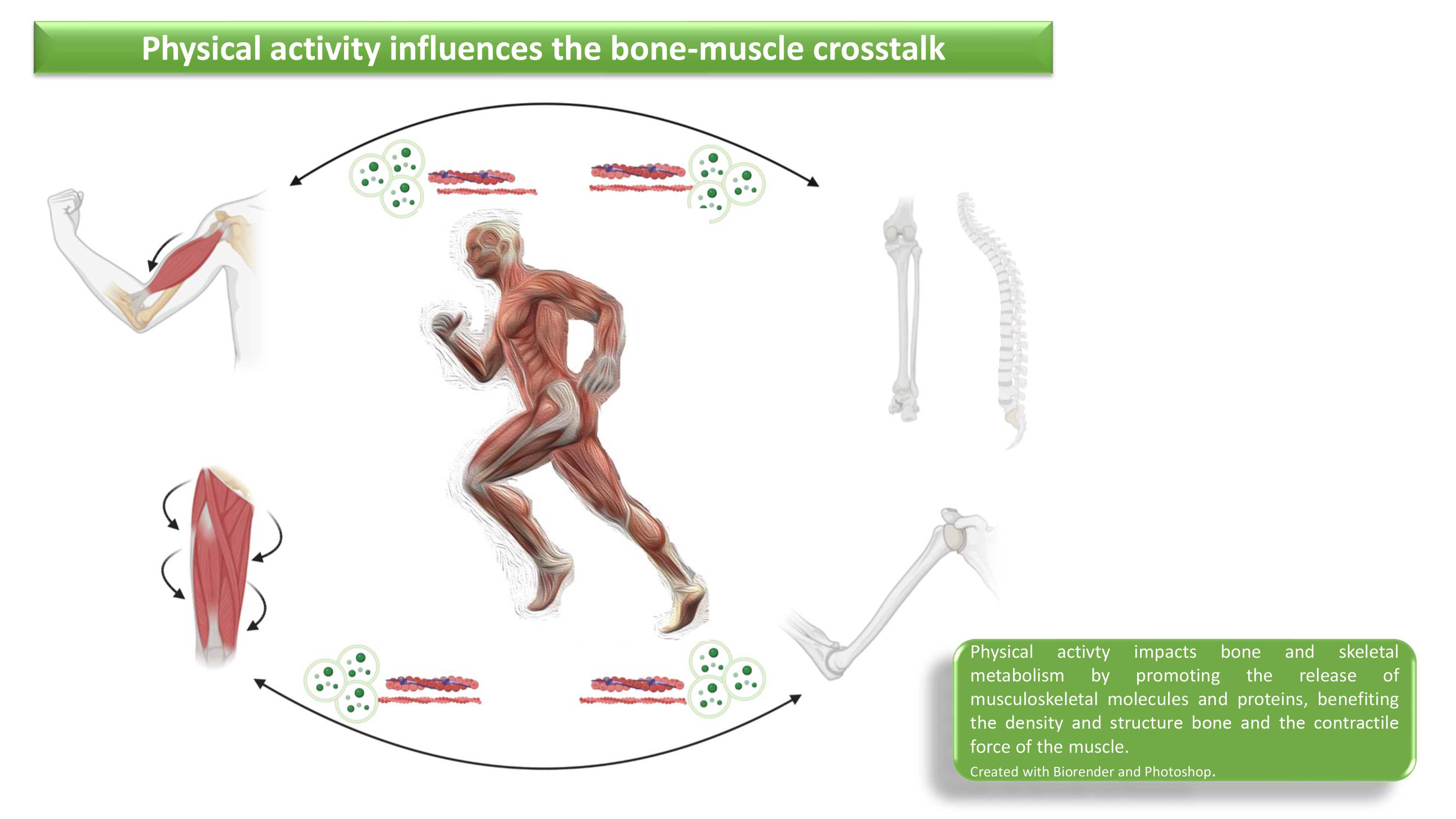{kind=link}
{kind=link}
{kind=link}
| MicroRNAs | Biological Functions | References |
|---|---|---|
| • miR-21 | Implicated in promoting bone formation targeting Spry1 in MSCs and involved in the proliferation and differentiation processes of muscle satellite cells by targeting KLF3. | [85,86] |
| • miR-21/miR129/miR-378 | Involved in osteogenic differentiation after physical activity and play a role in myogenesis. | [49,87,88] |
| • miR-143 | Upregulated during physical activity with a reduction in IGFBP-5 and is also involved in osteogenic differentiation. | [29,89,90] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dalle Carbonare, L.; Minoia, A.; Zouari, S.; Piritore, F.C.; Vareschi, A.; Romanelli, M.G.; Valenti, M.T. Crosstalk between Bone and Muscles during Physical Activity. Cells 2023, 12, 2088. https://doi.org/10.3390/cells12162088
Dalle Carbonare L, Minoia A, Zouari S, Piritore FC, Vareschi A, Romanelli MG, Valenti MT. Crosstalk between Bone and Muscles during Physical Activity. Cells. 2023; 12(16):2088. https://doi.org/10.3390/cells12162088
Chicago/Turabian StyleDalle Carbonare, Luca, Arianna Minoia, Sharazed Zouari, Francesca Cristiana Piritore, Anna Vareschi, Maria Grazia Romanelli, and Maria Teresa Valenti. 2023. "Crosstalk between Bone and Muscles during Physical Activity" Cells 12, no. 16: 2088. https://doi.org/10.3390/cells12162088