

Gradient Magnetic Field Accelerates Division of E. coli Nissle 1917
 ,
,
Abstract
1. Introduction
2. Materials and Methods
3. Results
3.1. Magnetic Properties and Magnetophoresis of E. coli Nissle 1917
3.2. EcN Cultivation in the Presence of a Gradient MF
3.3. Model and Predictions of Accelerating Cell Division in Gradient Magnetic Fields of a Special Spatial Configuration
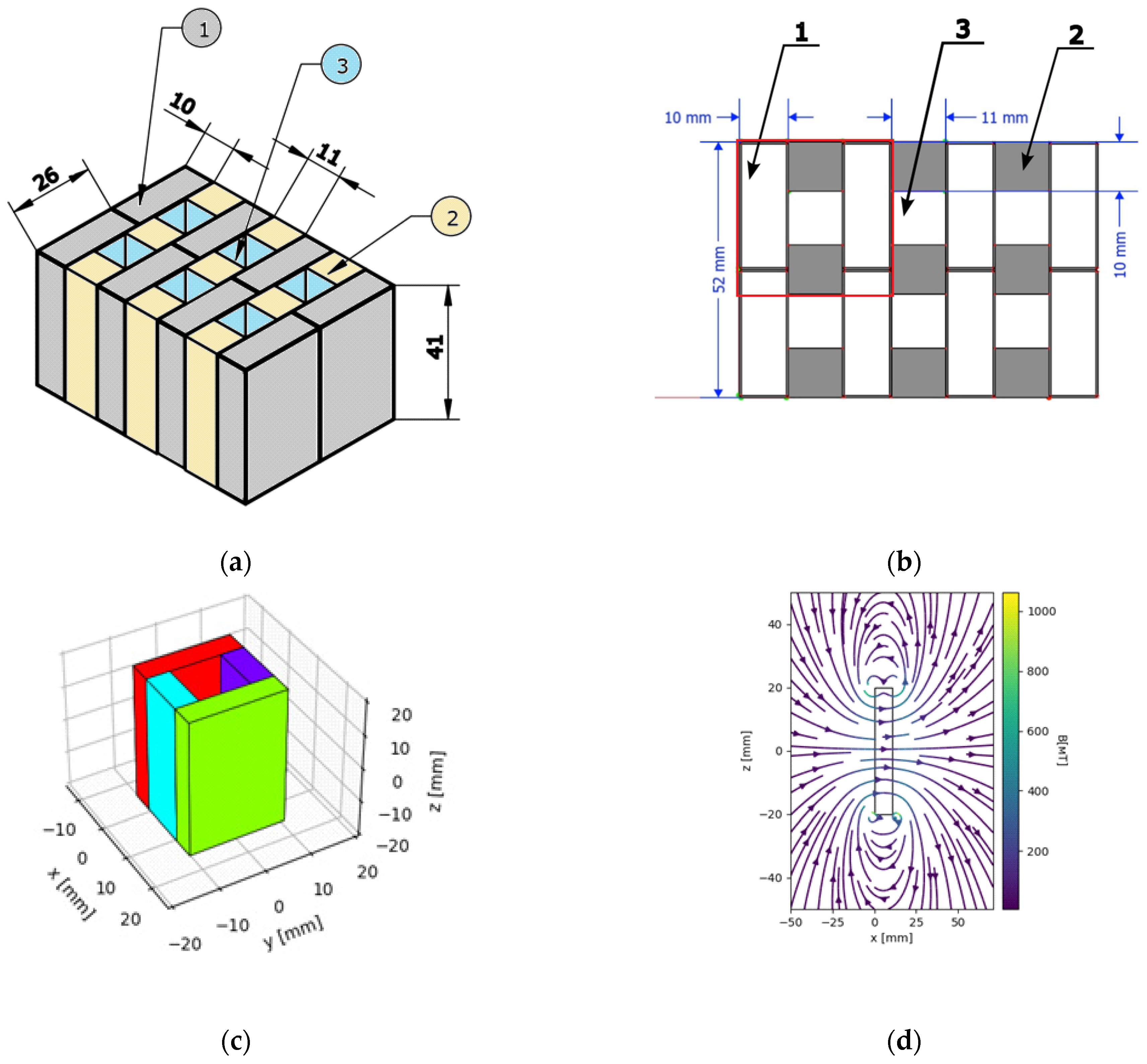
3.3.1. Mapping Magnetic Field Distribution
3.3.2. Magnetic Forces Exerted on BMN Chains
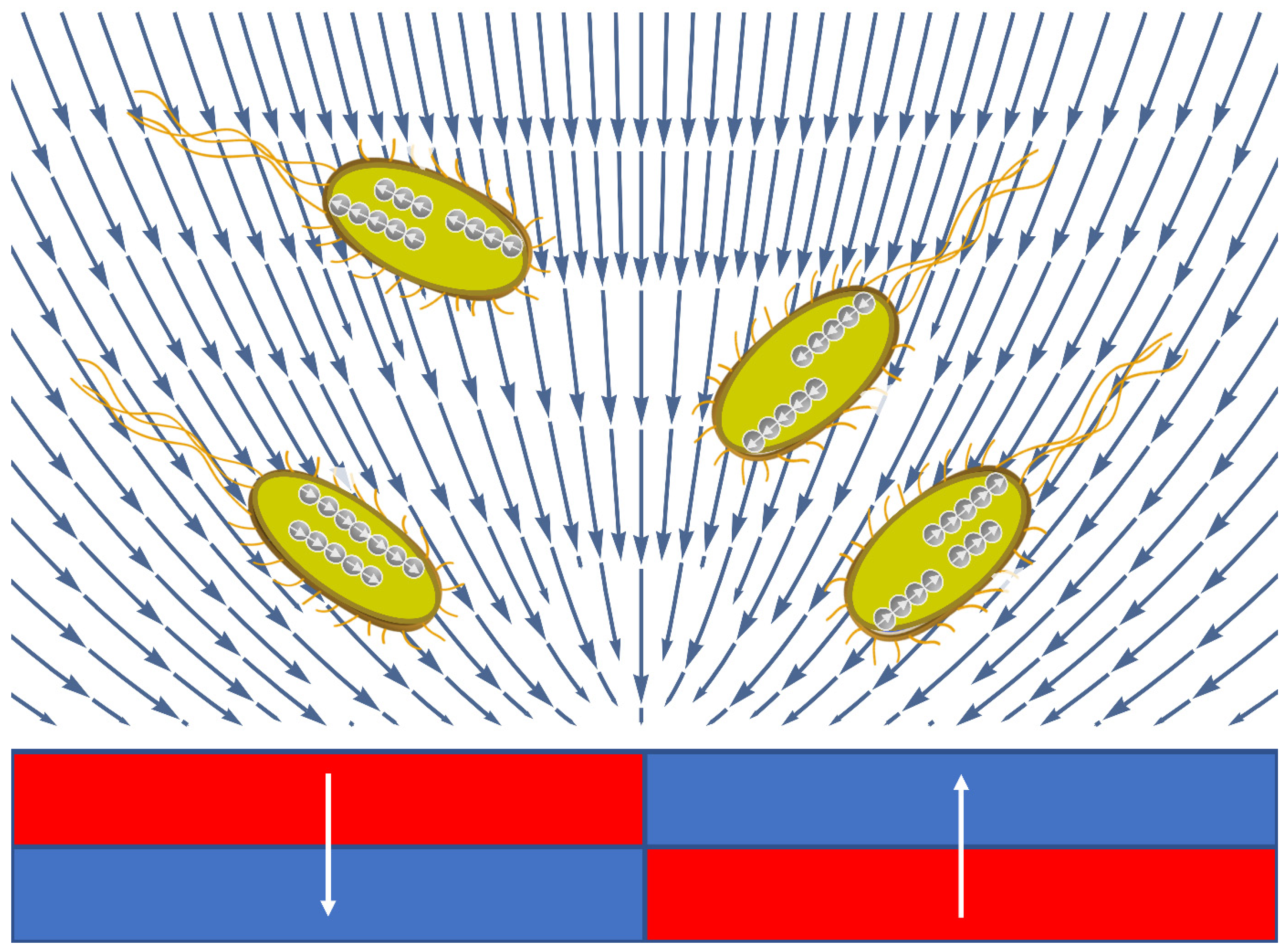
3.3.3. Mechanism of E. coli Division in an MF Requires a Special Spatial Configuration of the Field
3.3.4. Magnetic Gradient Force Changes the Timescale of Bacterial Division
3.3.5. Experimental: Static Gradient MF Accelerates Growth of E. coli Nissle 1917
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, J.D.; Levin, P.A. Metabolism. Cell Growth and the Bacterial Cell Cycle. Nat. Rev. Microbiol. 2009, 7, 822. [Google Scholar] [CrossRef] [PubMed]
- Schaechter, M.; MaalOe, O.; Kjeldgaard, N.O. Dependency on Medium and Temperature of Cell Size and Chemical Composition during Balanced Growth of Salmonella Typhimurium. J. Gen. Microbiol. 1958, 19, 592. [Google Scholar] [CrossRef] [PubMed]
- Ayala, M.R.; Syrovets, T.; Hafner, S.; Zablotskii, V.; Dejneka, A.; Simmet, T. Spatiotemporal Magnetic Fields Enhance Cytosolic Ca2+ Levels and Induce Actin Polymerization via Activation of Voltage-Gated Sodium Channels in Skeletal Muscle Cells. Biomaterials 2018, 163, 174. [Google Scholar] [CrossRef]
- Zablotskii, V.; Lunov, O.; Novotná, B.; Churpita, O.; Trošan, P.; Holáň, V.; Syková, E.; Dejneka, A.; Kubinová, Š. Down-Regulation of Adipogenesis of Mesenchymal Stem Cells by Oscillating High-Gradient Magnetic Fields and Mechanical Vibration. Appl. Phys. Lett. 2014, 105, 103702. [Google Scholar] [CrossRef]
- Zablotskii, V.; Polyakova, T.; Dejneka, A. Cells in the Non-Uniform Magnetic World: How Cells Respond to High-Gradient Magnetic Fields. BioEssays 2018, 40, 1800017. [Google Scholar] [CrossRef] [PubMed]
- Wosik, J.; Chen, W.; Qin, K.; Ghobrial, R.M.; Kubiak, J.Z.; Kloc, M. Magnetic Field Changes Macrophage Phenotype. Biophys. J. 2018, 114, 2001. [Google Scholar] [CrossRef] [PubMed]
- Zablotskii, V.; Polyakova, T.; Lunov, O.; Dejneka, A. How a High-Gradient Magnetic Field Could Affect Cell Life. Sci. Rep. 2016, 6, 37407. [Google Scholar] [CrossRef] [PubMed]
- Strahl, H.; Hamoen, L.W. Membrane Potential Is Important for Bacterial Cell Division. Proc. Natl. Acad. Sci. USA 2010, 107, 12281. [Google Scholar] [CrossRef]
- Colin, A.; Micali, G.; Faure, L.; Lagomarsino, M.C.; van Teeffelen, S. Two Different Cell-Cycle Processes Determine the Timing of Cell Division in Escherichia Coli. Elife 2021, 10, e67495. [Google Scholar] [CrossRef]
- CBrangwynne, P.; Koenderink, G.H.; MacKintosh, F.C.; Weitz, D.A. Nonequilibrium Microtubule Fluctuations in a Model Cytoskeleton. Phys. Rev. Lett. 2008, 100, 118104. [Google Scholar] [CrossRef]
- Zablotskii, V.; Lunov, O.; Kubinova, S.; Polyakova, T.; Sykova, E.; Dejneka, A. Effects of High-Gradient Magnetic Fields on Living Cell Machinery. J. Phys. D Appl. Phys. 2016, 49, 493003. [Google Scholar] [CrossRef]
- KCylke, C.; Banerjee, S. Accelerating Growth and Stochastic Shape Dynamics in Rod-like Bacteria. BioRxiv 2022. [Google Scholar] [CrossRef]
- Biswas, K.; Brenner, N. Cell-Division Time Statistics from Stochastic Exponential Threshold-Crossing. BioRxiv 2022, 10.1101/20. [Google Scholar] [CrossRef]
- Zhang, L.; Hou, Y.; Li, Z.; Ji, X.; Wang, Z.; Wang, H.; Tian, X.; Yu, F.; Yang, Z.; Pi, L.; et al. 27 T Ultra-High Static Magnetic Field Changes Orientation and Morphology of Mitotic Spindles in Human Cells. Elife 2017, 6, e22911. [Google Scholar] [CrossRef] [PubMed]
- Tseng, P.; Judy, J.W.; Di Carlo, D. Magnetic Nanoparticle–Mediated Massively Parallel Mechanical Modulation of Single-Cell Behavior. Nat. Methods 2012, 9, 1113. [Google Scholar] [CrossRef] [PubMed]
- Mendelson, N.H. Bacterial Growth and Division: Genes, Structures, Forces, and Clocks. Microbiol. Rev. 1982, 46, 341–375. [Google Scholar] [CrossRef]
- Blakemore, R. Magnetotactic Bacteria. Science 1975, 190, 377. [Google Scholar] [CrossRef]
- Kobayashi, A.; Yamamoto, N.; Kirschvink, J. Studies of Inorganic Crystals in Biological Tissue: Magnetic in Human Tumor. J. Jpn. Soc. Powder Powder Metall. 1997, 44, 294. [Google Scholar] [CrossRef]
- Gorobets, O.; Gorobets, S.; Koralewski, M. Physiological Origin of Biogenic Magnetic Nanoparticles in Health and Disease: From Bacteria to Humans. Int. J. Nanomed. 2017, 12, 4371. [Google Scholar] [CrossRef]
- Kirschvink, J.L. Magnetite Biomineralization and Geomagnetic Sensitivity in Higher Animals: An Update and Recommendations for Future Study. Bioelectromagnetics 1989, 10, 239–259. [Google Scholar] [CrossRef]
- Kirschvink, J.L.; Gould, J.L. Biogenic Magnetite as a Basis for Magnetic Field Detection in Animals. Biosystems 1981, 13, 181–201. [Google Scholar] [CrossRef] [PubMed]
- Kirschvink, J. Magnetite-Based Magnetoreception. Curr. Opin. Neurobiol. 2001, 11, 462. [Google Scholar] [CrossRef] [PubMed]
- JKirschvink, L.; Douglas, S.J.; Bruce, J.M. Magnetite Biomineralization and Magnetoreception in Organisms a New Biomagnetism; Plenum Press: New York, NY, USA, 1985. [Google Scholar]
- Bauer, G.B.; Fuller, M.; Perry, A.; Dunn, J.R.; Zoeger, J. Magnetoreception and Biomineralization of Magnetite in Cetaceans; Springer: Boston, MA, USA, 1985. [Google Scholar]
- Gorobets, S.V.; Yu, G.O.; Demianenko, I.V.; Nikolaenko, R.N. Self-Organization of Magnetite Nanoparticles in Providing Saccharomyces Cerevisiae Yeasts with Magnetic Properties. J. Magn. Magn. Mater. 2013, 337, 53–57. [Google Scholar] [CrossRef]
- Gorobets, S.V.; Gorobets, O.Y.; Chyzh, Y.M.; Sivenok, D.V. Magnetic Dipole Interaction of Endogenous Magnetic Nanoparticles with Magnetoliposomes for Targeted Drug Delivery. Biophysics 2013, 58, 379. [Google Scholar] [CrossRef]
- Bazylinski, D.A.; Frankel, R.B. Magnetosome Formation in Prokaryotes. Nat. Rev. Microbiol. 2004, 2, 217. [Google Scholar] [CrossRef]
- Arakaki, A.; Nakazawa, H.; Nemoto, M.; Mori, T.; Matsunaga, T. Formation of Magnetite by Bacteria and Its Application. J. R. Soc. Interface 2008, 5, 977–999. [Google Scholar] [CrossRef]
- Taoka, A.; Asada, R.; Wu, L.-F.; Fukumori, Y. Polymerization of the Actin-Like Protein MamK, Which Is Associated with Magnetosomes. J. Bacteriol. 2007, 189, 8737. [Google Scholar] [CrossRef]
- Vainshtein, M.; Suzina, N.; Kudryashova, E.; Ariskina, E. New Magnet-Sensitive Structures in Bacterial and Archaeal Cells. Biol. Cell 2002, 94, 29. [Google Scholar] [CrossRef]
- Kirschvink, J.L.; Kobayashi-Kirschvink, A.; Diaz-Ricci, J.C.; Kirschvink, S.J. Magnetite in Human Tissues: A Mechanism for the Biological Effects of Weak ELF Magnetic Fields. Bioelectromagnetics 1992, 13, 101–113. [Google Scholar] [CrossRef]
- Staniland, S.S.; Moisescu, C.; Benning, L.G. Cell Division in Magnetotactic Bacteria Splits Magnetosome Chain in Half. J. Basic Microbiol. 2010, 50, 392. [Google Scholar] [CrossRef]
- Schubbe, S.; Wurdemann, C.; Peplies, J.; Heyen, U.; Wawer, C.; Gluckner, F.O.; Schuler, D. Transcriptional Organization and Regulation of Magnetosome Operons in Magnetospirillum Gryphiswaldense. Appl. Environ. Microbiol. 2006, 72, 5757. [Google Scholar] [CrossRef] [PubMed]
- Lohße, A.; Ullrich, S.; Katzmann, E.; Borg, S.; Wanner, G.; Richter, M.; Voigt, B.; Schweder, T.; Schu, D. Functional Analysis of the Magnetosome Island in Magnetospirillum Gryphiswaldense: The MamAB Operon Is Sufficient for Magnetite Biomineralization. PLoS ONE 2011, 6, e25561. [Google Scholar] [CrossRef] [PubMed]
- Gorobets, S.V.; Medviediev, O.V.; Gorobets, O.Y.; Ivanchenko, A. Biogenic Magnetic Nanoparticles in Human Organs and Tissues. Prog. Biophys. Mol. Biol. 2018, 135, 49. [Google Scholar] [CrossRef] [PubMed]
- Gorobets, O.Y.; Gorobets, S.V.; Sorokina, L.V. Biomineralization and Synthesis of Biogenic Magnetic Nanoparticles and Magnetosensitive Inclusions in Microorganisms and Fungi. Funct. Mater. 2014, 21, 373. [Google Scholar] [CrossRef]
- Gorobets, O.Y.; Gorobets, S.V.; Gorobets, Y.I. Biogenic Magnetic Nanoparticles. Biomineralization in Prokaryotes and Eukaryotes. In Dekker Encyclopedia of Nanoscience and Nanotechnology, 3rd ed.; CRC Press: New York, NY, USA, 2014; pp. 300–308. [Google Scholar]
- Medviediev, O.; Gorobets, O.Y.; Gorobets, S.V.; Yadrykhins’Ky, V.S. The Prediction of Biogenic Magnetic Nanoparticles Biomineralization in Human Tissues and Organs. J. Phys. Conf. Ser. 2017, 903, 012002. [Google Scholar] [CrossRef]
- Van de Walle, A.; Sangnier, A.P.; Abou-Hassan, A.; Curcio, A.; Hémadi, M.; Menguy, N.; Lalatonne, Y.; Luciani, N.; Wilhelm, C. Biosynthesis of Magnetic Nanoparticles from Nano-Degradation Products Revealed in Human Stem Cells. Proc. Natl. Acad. Sci. USA 2019, 116, 4044. [Google Scholar] [CrossRef]
- Schüler, D. Magnetoreception and Magnetosomes in Bacteria; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 2007. [Google Scholar]
- Elkady, A.S.; Iskakova, L.; Zubarev, A. On the Self-Assembly of Net-like Nanostructures in Ferrofluids. Phys. A Stat. Mech. Appl. 2015, 428, 257–265. [Google Scholar] [CrossRef]
- Gorobets, S.V.; Legenkii, Y.A.; Melnichuk, I.A. Break-up of Ni Microparticle Clusters in a Magnetic Field. J. Magn. Magn. Mater. 2000, 222, 159. [Google Scholar] [CrossRef]
- Baryakhtar, F.G.; Gorobets, Y.I.; Kosachevski, L.Y.; Ilchishyn, O.V.; Hizhenkov, P.K. Hexagonal Lattice of Cylindrical Magnetic Domains in Thin Films of a Ferrofluid. Magn. Gidrodin. 1981, 3, 120–123. [Google Scholar]
- Gorobets, S.; Gorobets, O.; Magerman, A.; Gorobets, Y.; Sharay, I. Biogenic Magnetic Nanoparticles in Plants. arXiv 2019, arXiv:1901.07212. [Google Scholar]
- Zhou, Y.; Han, Y. Engineered Bacteria as Drug Delivery Vehicles: Principles and Prospects. Eng. Microbiol. 2022, 2, 100034. [Google Scholar] [CrossRef]
- Akolpoglu, M.B.; Alapan, Y.; Dogan, N.O.; Baltaci, S.F.; Yasa, O.; Tural, G.A.; Sitti, M. Magnetically Steerable Bacterial Microrobots Moving in 3D Biological Matrices for Stimuli-Responsive Cargo Delivery. Sci. Adv. 2022, 8, eabo6163. [Google Scholar] [CrossRef]
- Aubry, M.; Wang, W.-A.; Guyodo, Y.; Delacou, E.; Guigner, J.-M.; Espeli, O.; Lebreton, A.; Guyot, F.; Gueroui, Z. Engineering E. Coli for Magnetic Control and the Spatial Localization of Functions. ACS Synth. Biol. 2020, 9, 3030. [Google Scholar] [CrossRef] [PubMed]
- Stritzker, J.; Weibel, S.; Hill, P.; Oelschlaeger, T.; Goebel, W.; Szalay, A. Tumor-Specific Colonization, Tissue Distribution, and Gene Induction by Probiotic Escherichia Coli Nissle 1917 in Live Mice. Int. J. Med. Microbiol. 2007, 297, 151. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, Y.; Xia, L.; Zhang, X.; Ding, X.; Yan, F.; Wu, F. Escherichia Coli Nissle 1917 Targets and Restrains Mouse B16 Melanoma and 4T1 Breast Tumors through Expression of Azurin Protein. Appl. Environ. Microbiol. 2012, 78, 7603. [Google Scholar] [CrossRef] [PubMed]
- Westphal, K.; Leschner, S.; Jablonska, J.; Loessner, H.; Weiss, S. Containment of Tumor-Colonizing Bacteria by Host Neutrophils. Cancer Res. 2008, 68, 2952. [Google Scholar] [CrossRef]
- Sturm, A.; Rilling, K.; Baumgart, D.C.; Gargas, K.; Abou-Ghazale, T.; Raupach, B.; Eckert, J.; Schumann, R.R.; Enders, C.; Sonnenborn, U.; et al. Escherichia Coli Nissle 1917 Distinctively Modulates T-Cell Cycling and Expansion via Toll-Like Receptor 2 Signaling. Infect. Immun. 2005, 73, 1452. [Google Scholar] [CrossRef]
- Arribas, B.; Rodríguez-Cabezas, M.; Camuesco, D.; Comalada, M.; Bailón, E.; Utrilla, P.; Nieto, A.; Concha, A.; Zarzuelo, A.; Gálvez, J. A Probiotic Strain of Escherichia Coli, Nissle 1917, given Orally Exerts Local and Systemic Anti-Inflammatory Effects in Lipopolysaccharide-Induced Sepsis in Mice. Br. J. Pharmacol. 2009, 157, 1024. [Google Scholar] [CrossRef]
- Zhang, Y.; Ji, W.; He, L.; Chen, Y.; Ding, X.; Sun, Y.; Hu, S.; Yang, H.; Huang, W.; Zhang, Y.; et al. Coli Nissle 1917-Derived Minicells for Targeted Delivery of Chemotherapeutic Drug to Hypoxic Regions for Cancer Therapy. Theranostics 2018, 8, 1690. [Google Scholar] [CrossRef]
- Gwyddion: Scanning Probe Microscopy Data Visualisation and Analysis. Available online: https://sourceforge.net/projects/gwyddion/ (accessed on 30 October 2022).
- NT_MDT, MFM General Concept. Available online: https://www.ntmdt-si.com/resources/spm-theory/theoretical-background-of-spm/2-scanning-force-microscopy--(sfm)/27-magnetic-force-microscopy-quantitative-results-treatment/271-mfm-general-concept (accessed on 30 October 2022).
- Stover, N.A.; Cavalcanti, A.R.O. Using NCBI BLAST. Curr. Protoc. Essent. Lab. Tech. 2014, 8, 11.1.1–11.1.35. [Google Scholar] [CrossRef]
- Hubert, A.; Schäfer, R. Magnetic Domains: The Analysis of Magnetic Microstructures; Springer: New York, NY, USA, 1998. [Google Scholar]
- Zablotskii, V.; Pastor, J.M.; Larumbe, S.; Pe, J.I.; Recarte, V.; Go, C.; Ha, U.; Schu, W.; Zborowski, M. High-Field Gradient Permanent Micromagnets for Targeted Drug Delivery with Magnetic Nanoparticles; American Institute of Physics: College Park, MD, USA, 2010; Volume 1311, pp. 152–157. [Google Scholar]
- Samofalov, V.N.; Belozorov, D.P.; Ravlik, A.G. Strong Stray Fields in Systems of Giant Magnetic Anisotropy Magnets. Physics-Uspekhi 2013, 56, 269. [Google Scholar] [CrossRef]
- Nadkarni, R.; Barkley, S.; Fradin, C. A Comparison of Methods to Measure the Magnetic Moment of Magnetotactic Bacteria through Analysis of Their Trajectories in External Magnetic Fields. PLoS ONE 2013, 8, e82064. [Google Scholar] [CrossRef] [PubMed]
- Callaway, E. How Bacteria Break a Magnet. Nature 2011. [Google Scholar] [CrossRef]
- Katzmann, E.; Müller, F.D.; Lang, C.; Messerer, M.; Winklhofer, M.; Plitzko, J.M.; Schüler, D. Magnetosome Chains Are Recruited to Cellular Division Sites and Split by Asymmetric Septation. Mol. Microbiol. 2011, 82, 1316. [Google Scholar] [CrossRef] [PubMed]
- Sabass, B.; Koch, M.D.; Liu, G.; Stone, H.A.; Shaevitz, J.W. Force Generation by Groups of Migrating Bacteria. Proc. Natl. Acad. Sci. USA 2017, 114, 7266. [Google Scholar] [CrossRef] [PubMed]
- Dewachter, L.; Verstraeten, N.; Fauvart, M.; Michiels, J. An Integrative View of Cell Cycle Control in Escherichia Coli. FEMS Microbiol. Rev. 2018, 42, 116. [Google Scholar] [CrossRef] [PubMed]
- Michelsen, O.; de Mattos, M.J.T.; Jensen, P.R.; Hansen, F.G. Precise Determinations of C and D Periods by Flow Cytometry in Escherichia Coli K-12 and B/R. Microbiology 2003, 149, 1001. [Google Scholar] [CrossRef] [PubMed]
- Zwietering, M.H.; Jongenburger, I.; Rombouts, F.M.; van ’t Riet, K. Modeling of the Bacterial Growth Curve. Appl. Environ. Microbiol. 1990, 56, 1875. [Google Scholar] [CrossRef]
- You, Z.; Pearce, D.J.G.; Sengupta, A.; Giomi, L. Geometry and Mechanics of Microdomains in Growing Bacterial Colonies. Phys. Rev. X 2018, 8, 031065. [Google Scholar] [CrossRef]
- Panlilio, M.; Grilli, J.; Tallarico, G.; Iuliani, I.; Sclavi, B.; Cicuta, P.; Lagomarsino, M.C. Threshold Accumulation of a Constitutive Protein Explains E. Coli Cell-Division Behavior in Nutrient Upshifts. Proc. Natl. Acad. Sci. USA 2021, 118, e2016391118. [Google Scholar] [CrossRef]
- Milo, R.; Phillips, R. Bionumbers. Available online: http://bionumbers.hms.harvard.edu/ (accessed on 30 October 2022).
- Wallden, M.; Fange, D.; Lundius, E.G.; Baltekin, Ö.; Elf, J. The Synchronization of Replication and Division Cycles in Individual E. Coli Cells. Cell 2016, 166, 729. [Google Scholar] [CrossRef] [PubMed]
- Kuchel, P.W.; Chapman, B.E.; Bubb, W.A.; Hansen, P.E.; Durrant, C.J.; Hertzberg, M.P. Magnetic susceptibility: Solutions, emulsions, and cells. Concepts Magn. Reson. Educ. J. 2003, 18, 56–71. [Google Scholar] [CrossRef]
- Biedermann, A.R.; Pettke, T.; Koch, C.B.; Hirt, A.M. Magnetic anisotropy in clinopyroxene and orthopyroxene single crystals. Am. Mineral. 1968, 53, 406–415. [Google Scholar] [CrossRef]
- Liu, P.Y.; Chin, L.K.; Ser, W.; Ayi, T.C.; Yap, P.H.; Bourouina, T.; Leprince-Wang, Y. Real-time Measurement of Single Bacterium’s Refractive Index Using Optofluidic Immersion Refractometry. Procedia Eng. 2014, 87, 356–359. [Google Scholar] [CrossRef]
- Semsey, S.; Andersson, A.M.C.; Krishna, S.; Jensen, M.H.; Massé, E.; Sneppen, K. Genetic regulation of fluxes: Iron homeostasis of Escherichia coli. Nucleic. Acids. Res. 2006, 34, 4960–4967. [Google Scholar] [CrossRef] [PubMed]
- Fasnacht, M.; Polacek, N. Oxidative Stress in Bacteria and the Central Dogma of Molecular Biology. Front. Mol. Biosci. 2021, 8, 671037. [Google Scholar] [CrossRef] [PubMed]
- Nies, D.H. Resistance to cadmium, cobalt, zinc, and nickel in microbes. Plasmid 1992, 27, 17–28. [Google Scholar] [CrossRef] [PubMed]
- Hill, P.J.; Stritzker, J.; Scadeng, M.; Geissinger, U.; Haddad, D.; Basse-Lüsebrink, T.C.; Gbureck, U.; Jakob, P.; Szalay, A.A. Magnetic resonance imaging of tumors colonized with bacterial ferritin-expressing Escherichia coli. PLoS ONE 2011, 6, e25409. [Google Scholar] [CrossRef]
- García-Prieto, A.; Alonso, J.; Muñoz, D.; Marcano, L.; de Cerio, A.A.D.; Fernández de Luis, R.; Orue, I.; Mathon, O.; Muela, A.; Fdez-Gubieda, M.L. On the Mineral Core of Ferritin-like Proteins: Structural and Magnetic Characterization. Nanoscale 2016, 8, 1088. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| The Strain of the Microorganism | E-Number (І, %) | ||||
|---|---|---|---|---|---|
| Proteins of Magnetospirillum gryphiswaldense MSR-1 | |||||
| MamA | MamB | MamM | MamO | MamE | |
| Escherichia coli Nissle 1917 | 0.001 23.86% | 30.54% | 29.89% | 29.70% | 40.38% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gorobets, S.; Gorobets, O.; Sharai, I.; Polyakova, T.; Zablotskii, V. Gradient Magnetic Field Accelerates Division of E. coli Nissle 1917. Cells 2023, 12, 315. https://doi.org/10.3390/cells12020315
Gorobets S, Gorobets O, Sharai I, Polyakova T, Zablotskii V. Gradient Magnetic Field Accelerates Division of E. coli Nissle 1917. Cells. 2023; 12(2):315. https://doi.org/10.3390/cells12020315
Chicago/Turabian StyleGorobets, Svitlana, Oksana Gorobets, Iryna Sharai, Tatyana Polyakova, and Vitalii Zablotskii. 2023. "Gradient Magnetic Field Accelerates Division of E. coli Nissle 1917" Cells 12, no. 2: 315. https://doi.org/10.3390/cells12020315
APA StyleGorobets, S., Gorobets, O., Sharai, I., Polyakova, T., & Zablotskii, V. (2023). Gradient Magnetic Field Accelerates Division of E. coli Nissle 1917. Cells, 12(2), 315. https://doi.org/10.3390/cells12020315