
Reprogramming Megakaryocytes for Controlled Release of Platelet-like Particles Carrying a Single-Chain Thromboxane A2 Receptor-G-Protein Complex with Therapeutic Potential

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell Culture
2.3. Designing SC-TP-Gαq and SC-TP-Gαs
2.4. cDNA Synthesis and Plasmid Preparation
2.5. Expression of the Recombinant SC-TP-Gαq and SC-TP-Gαs in MK Cells
2.6. Western Blot
2.7. Flow Cytometry Analysis
3. Results
3.1. Bio-Engineering of SC-TP-Gαq and SC-TP-Gαs for Controlling of Platelet Functions
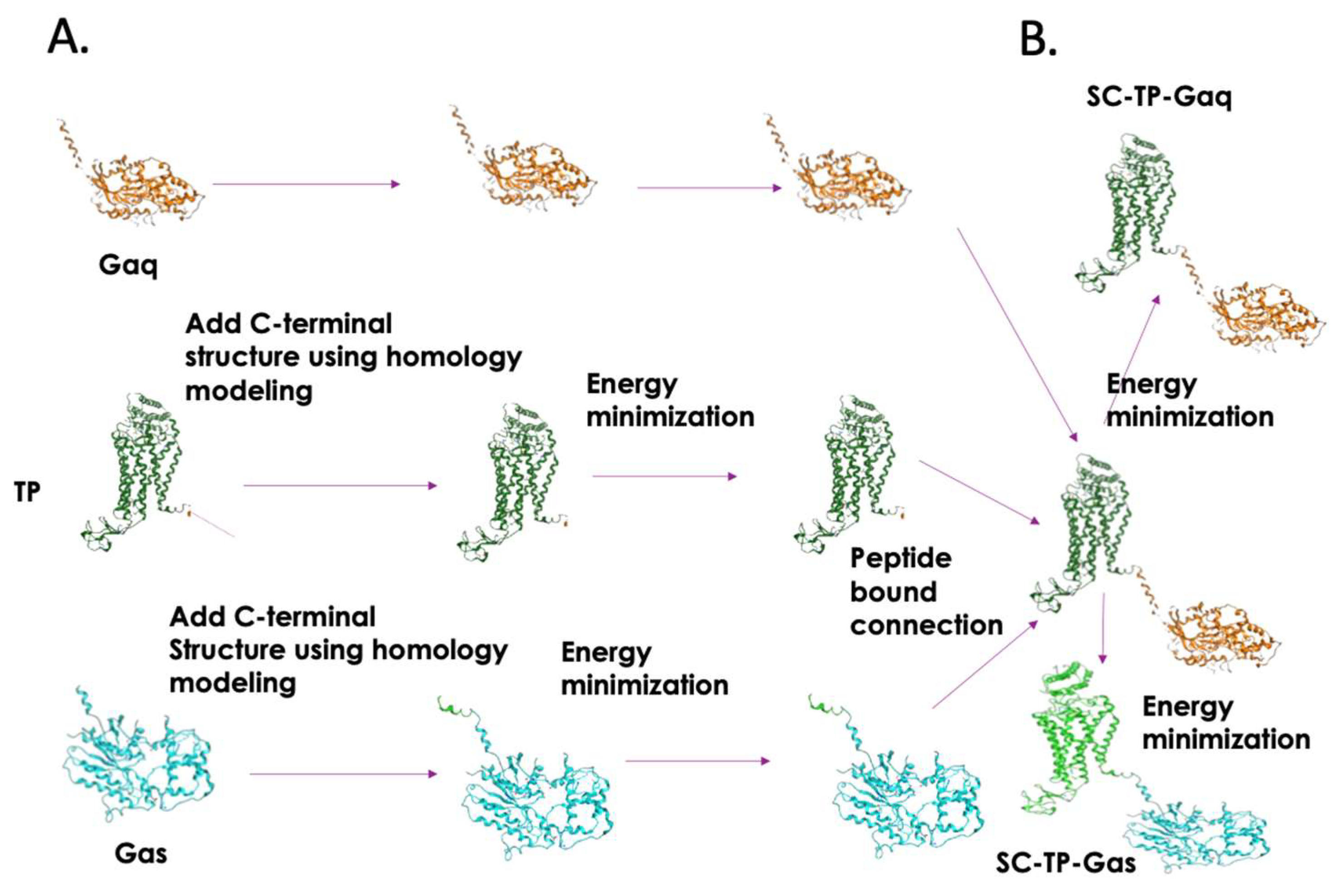
3.2. Structural Designs and Modeling of the Bio-Engineered SC-TP-Gαq and SC-TP-Gαs
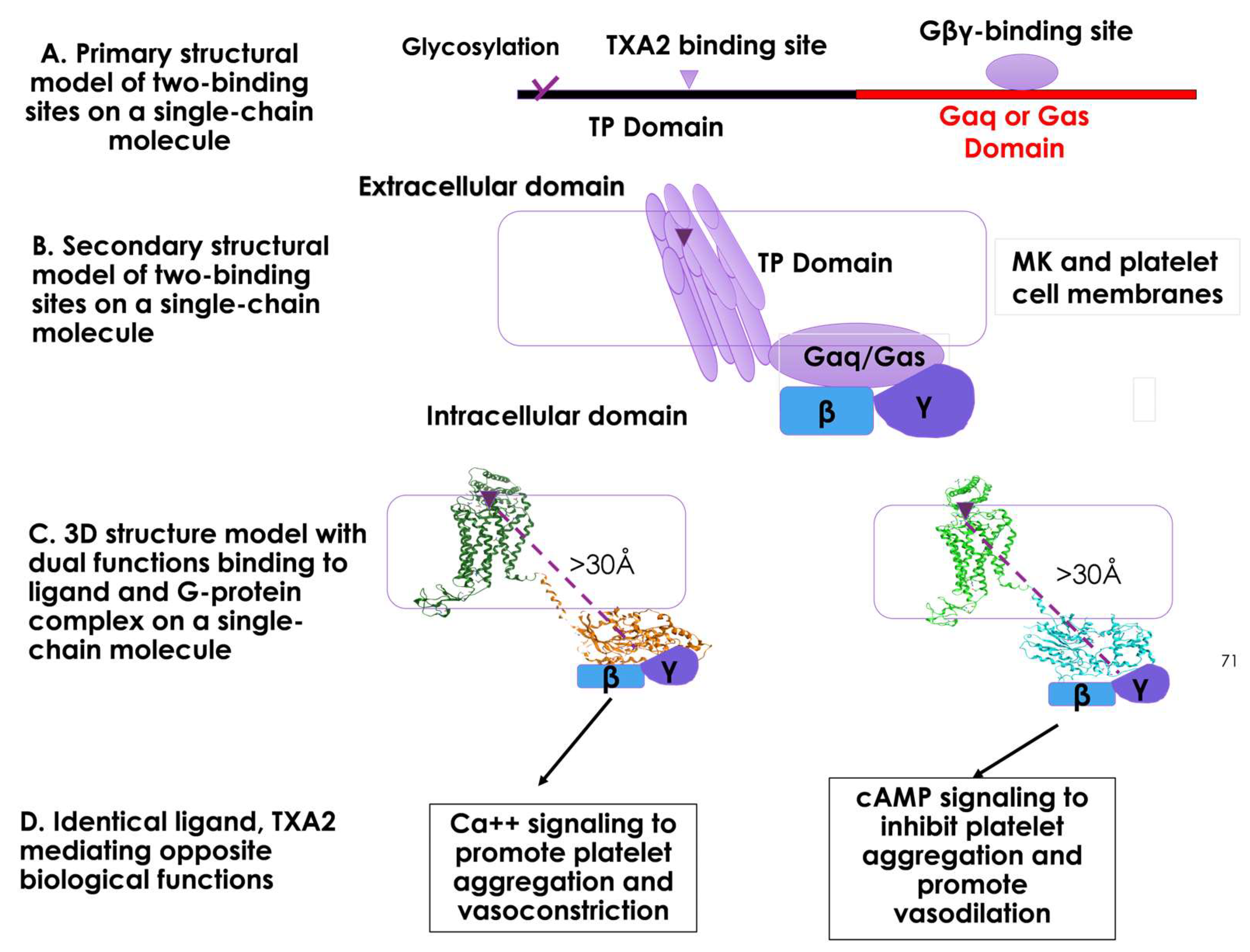
3.3. Structural and Functional Relationship of the Two Domains of TP and Gα within a Single Polypeptide Chain of SC-TP-Gαq and SC-TP-Gαs in Respect to the Cell Membrane
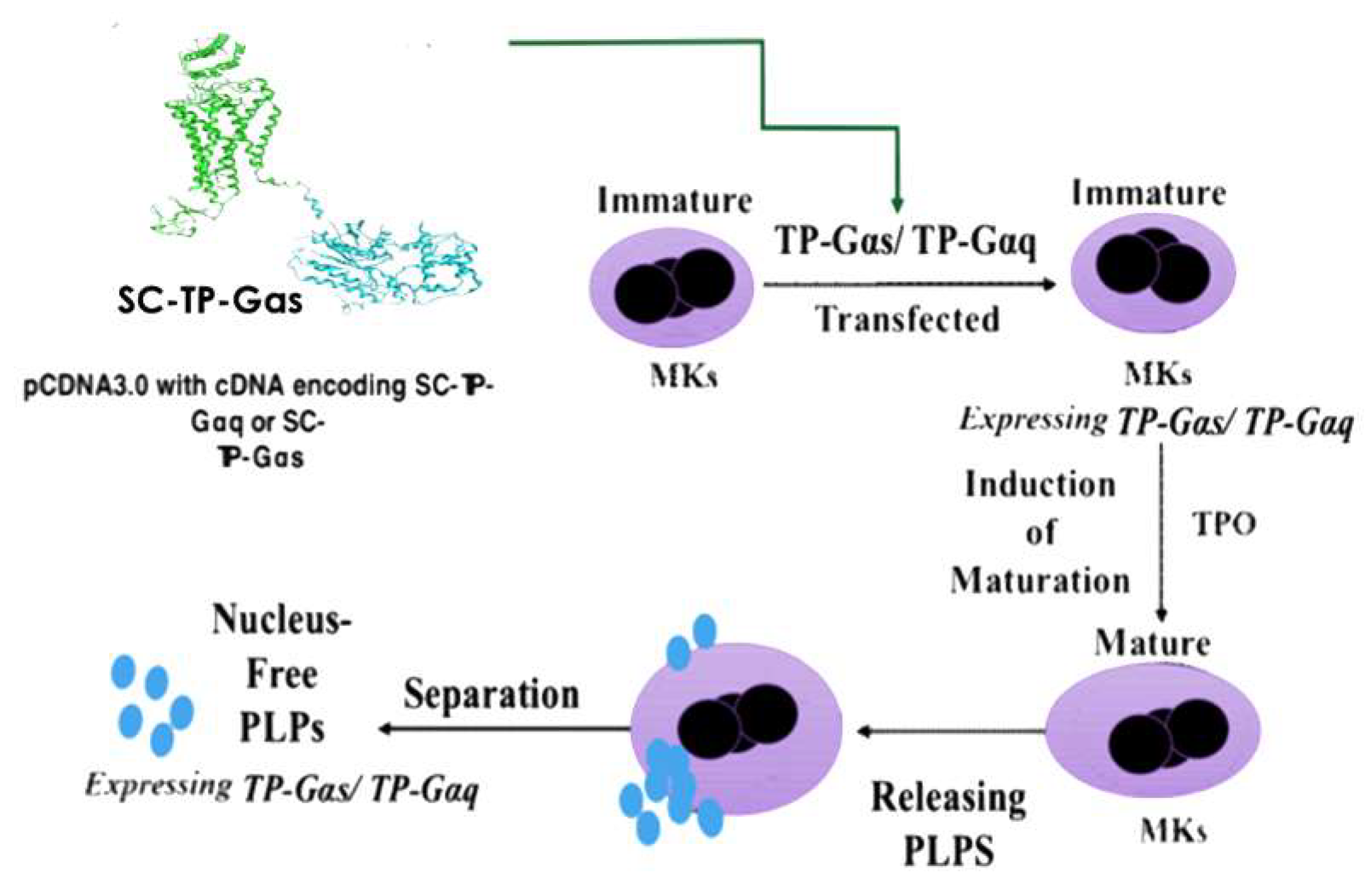
3.4. Design of Nucleus-Free PLPs to Deliver Recombinant SC-TP-Gαq or SC-TP-Gαs to Regulate Platelet Functions
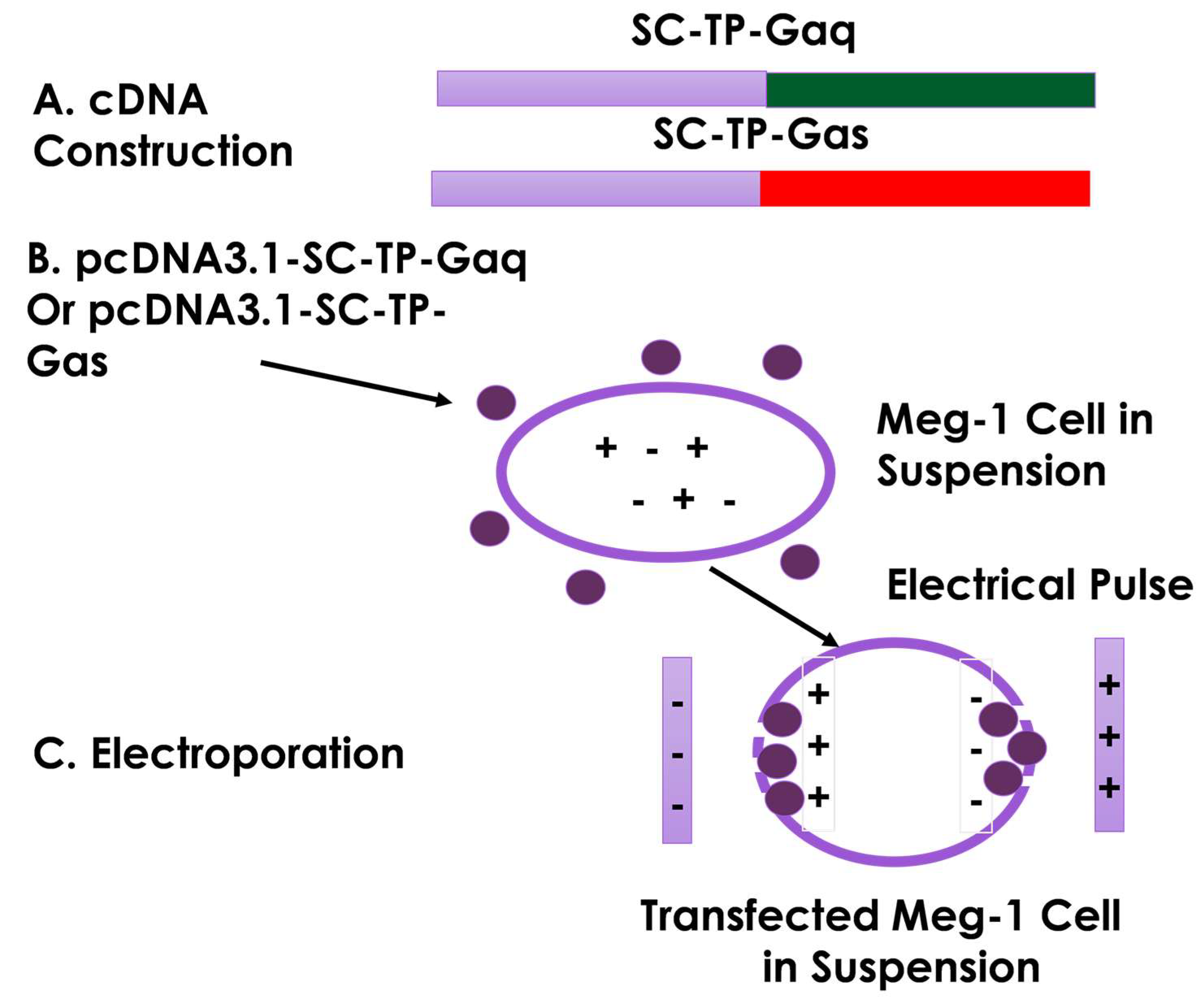
3.5. Construction of cDNA Vectors and Transfection of MK Cells Using Electroporation
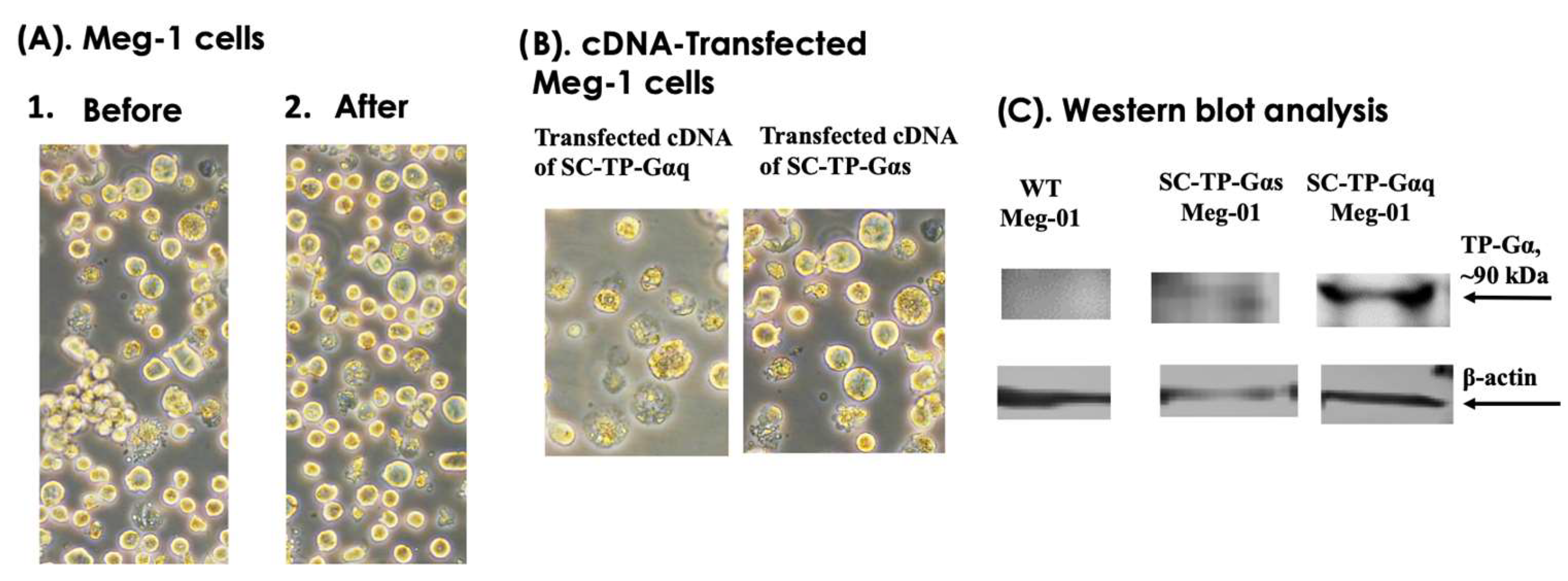
3.6. Transient Gene Expression of SC-TP-Gαq and SC-TP-Gαs on MK Cell Lines
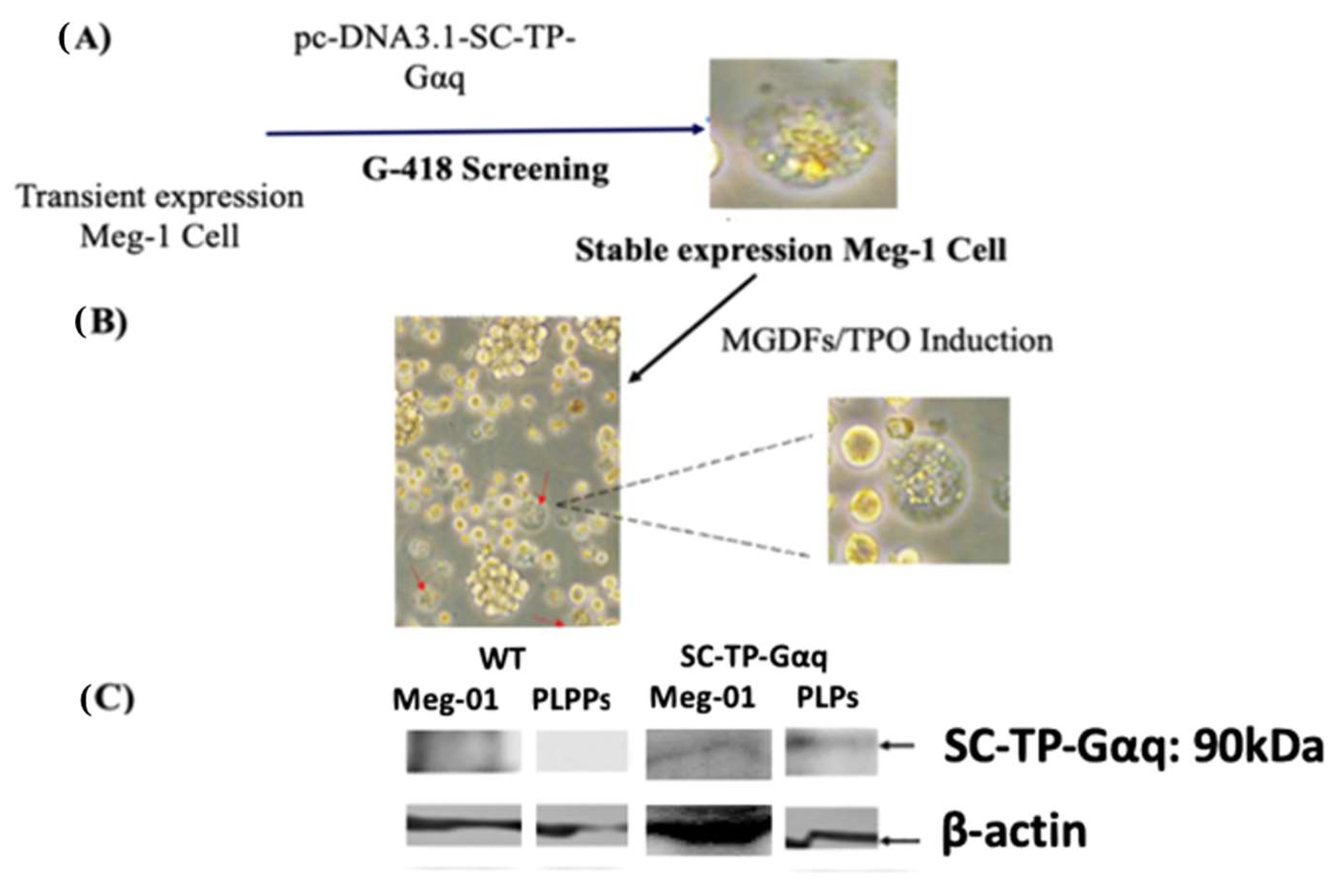
3.7. Screening of the Immature Meg-01 Cells Stably Expressing SC-TP-Gαq to Generate PLPs Carrying SC-TP-Gαq
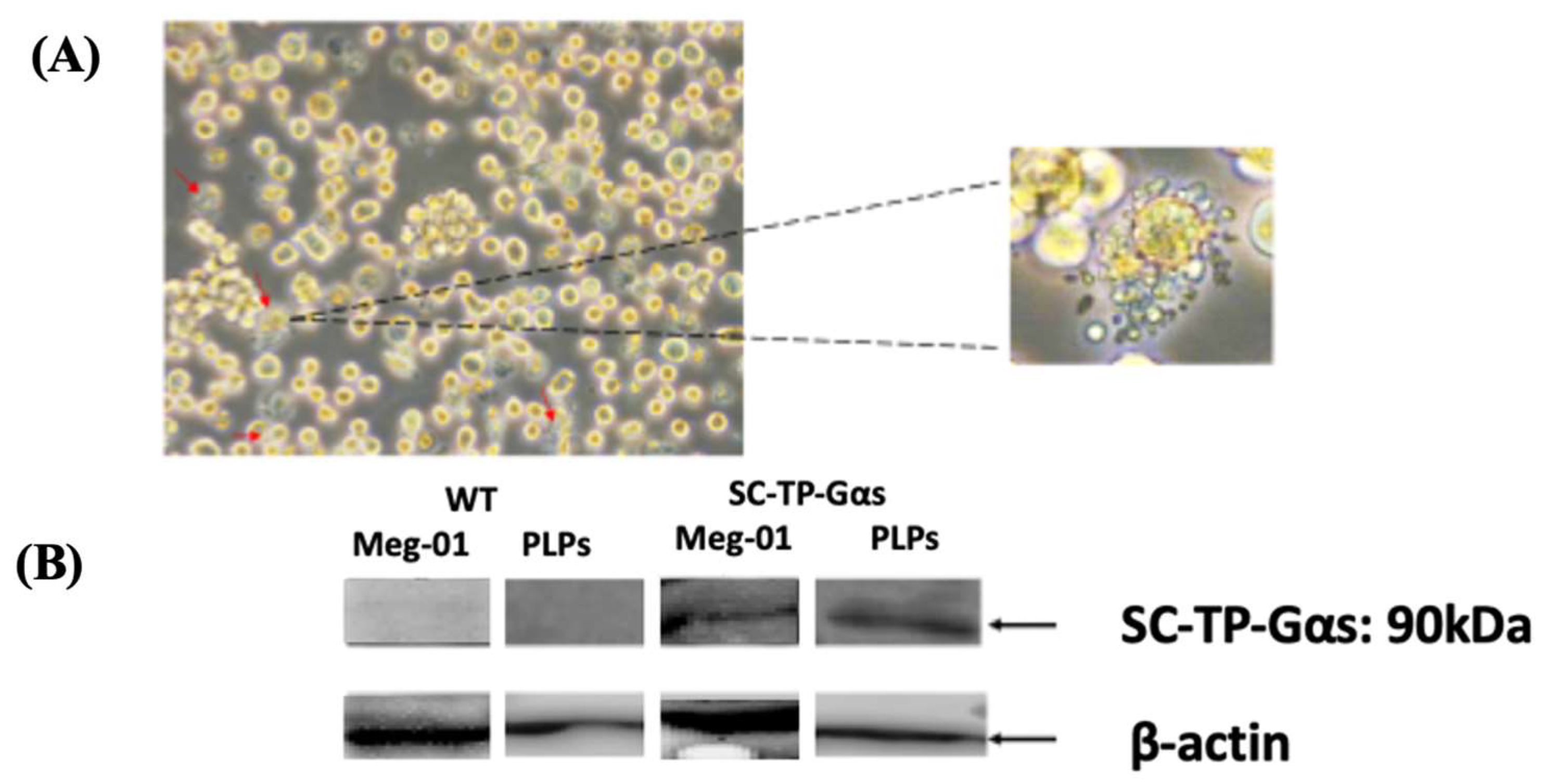
3.8. Screening of Meg-01 Cells Stably Expressing SC-TP-Gαs to Generate PLPs Carrying SC-TP-Gαs
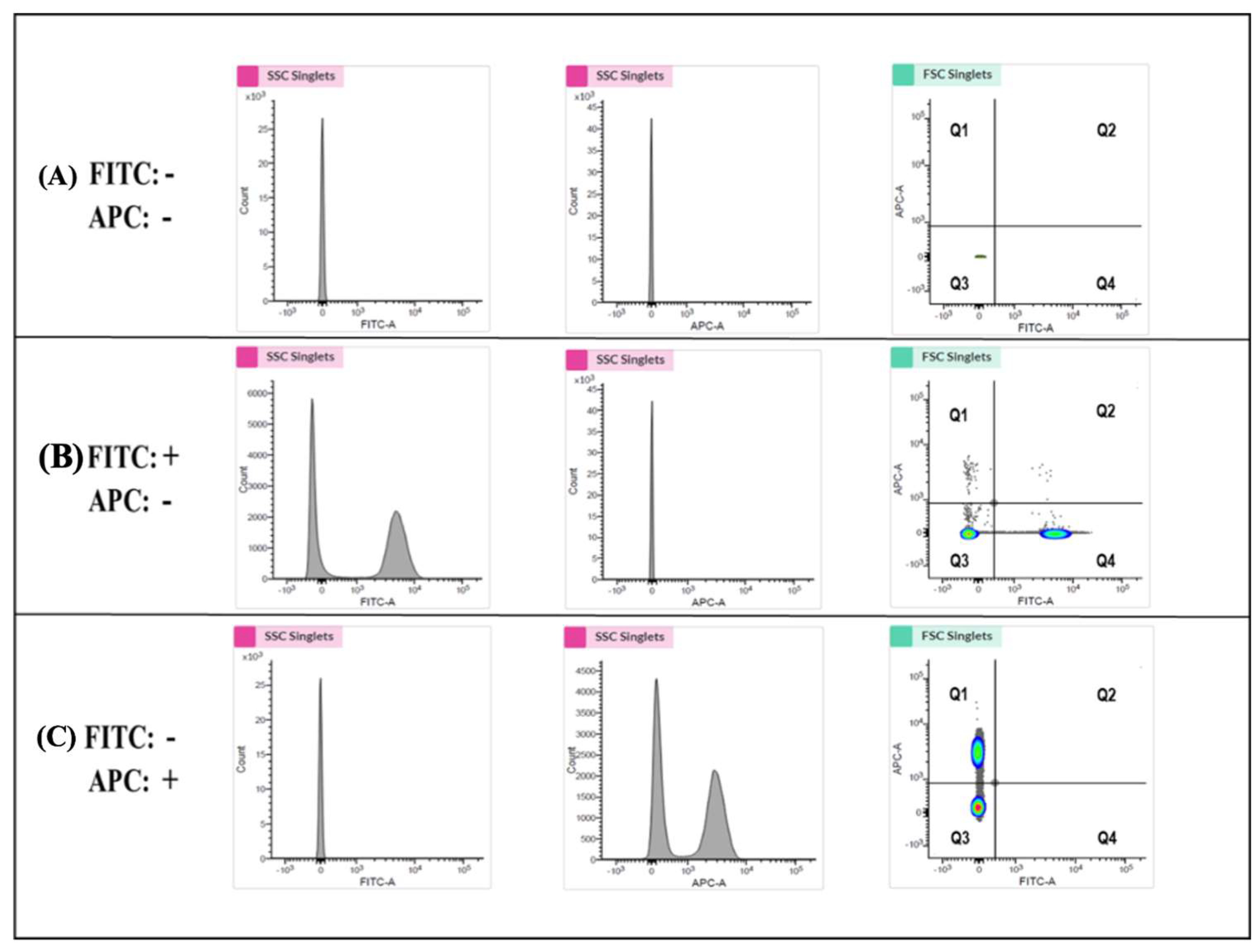
3.9. Establishing Highly Sensitive Flow Cytometry Using Double-Stained Human Platelets for Identification of the Biological Activities of PLP-SC-TP-Gαq and PLP-SC-TP-Gαs
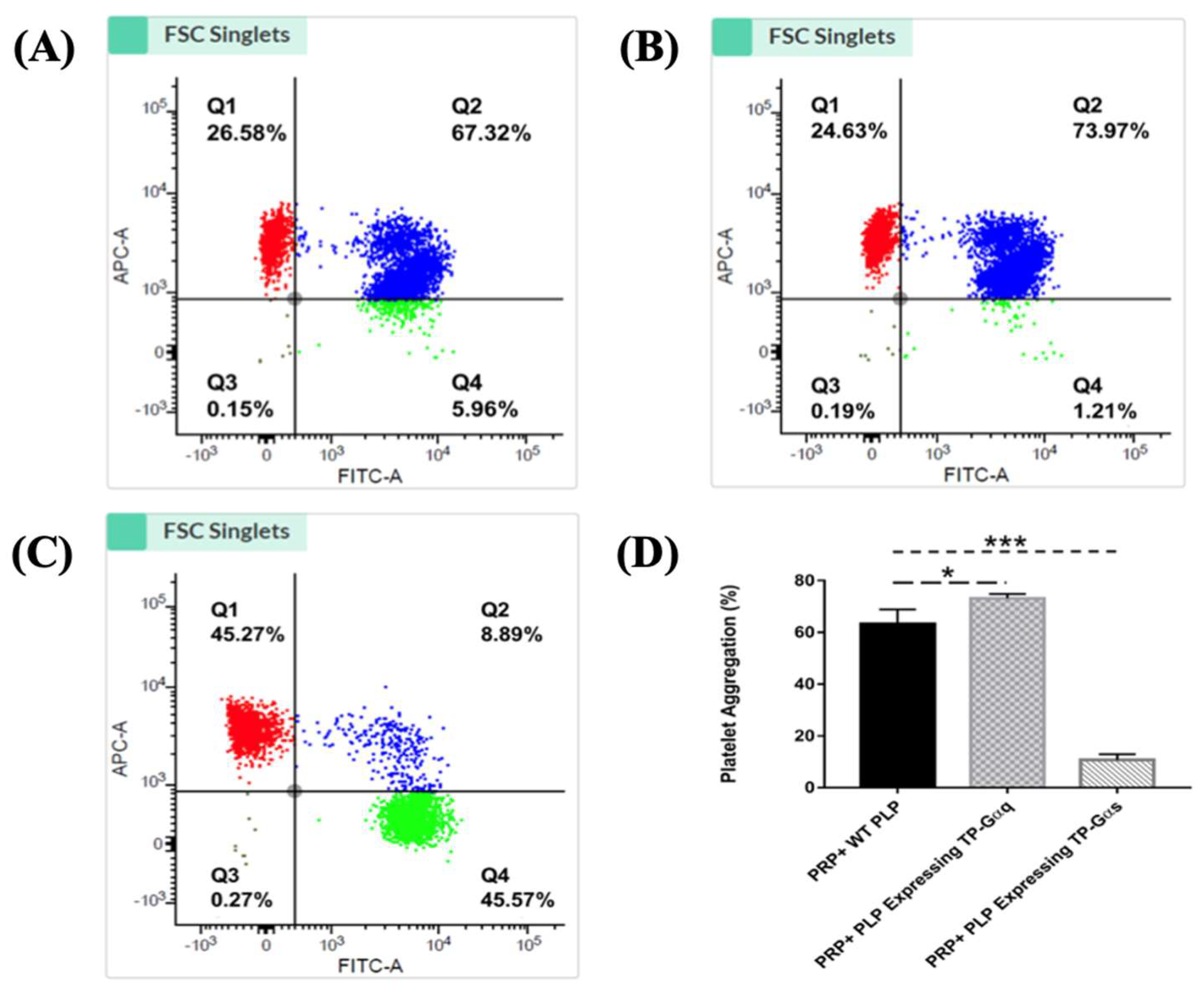
3.10. Determination of the Effects of the Nucleus-Free PLP-SC-TP-Gαq and PLP-SC-TP-Gαs on Regulating Platelet Aggregation Using the Double-Stained Flow Cytometry
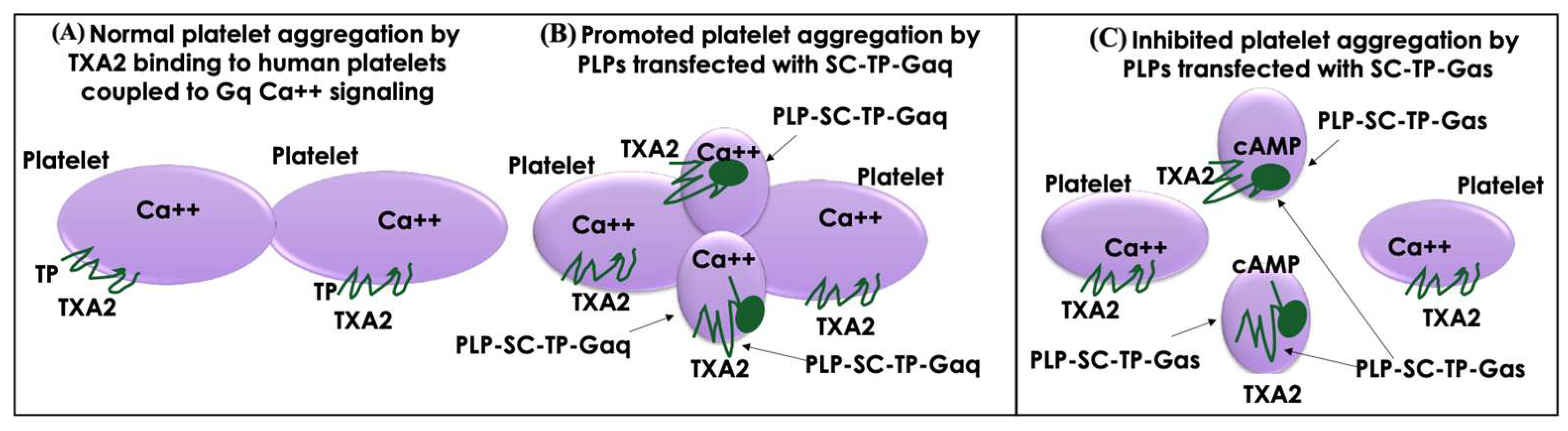
3.11. Working Model of PLP-SC-TP-Gαq and PLP-SC-TP-Gαs Regulation of Human Platelet Aggregation
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Atanasov, A.G.; Zotchev, S.B.; Dirsch, V.M.; Orhan, I.E.; Banach, M.; Rollinger, J.M.; Barreca, D.; Weckwerth, W.; Bauer, R.; Bayer, E.A.; et al. Natural products in drug discovery: Advances and opportunities. Nat. Rev. Drug Discov. 2021, 20, 200–216. [Google Scholar] [CrossRef] [PubMed]
- Lu, R.M.; Hwang, Y.C.; Liu, I.J.; Lee, C.C.; Tsai, H.Z.; Li, H.J.; Wu, H.C. Development of therapeutic antibodies for the treatment of diseases. J. Biomed. Sci. 2020, 27, 1. [Google Scholar] [CrossRef] [PubMed]
- Kaplon, H.; Chenoweth, A.; Crescioli, S.; Reichert, J.M. Antibodies to watch in 2022. MAbs 2022, 14, 2014296. [Google Scholar] [CrossRef] [PubMed]
- Walsh, G. Biopharmaceutical benchmarks 2018. Nat. Biotechnol. 2018, 36, 1136–1145. [Google Scholar] [CrossRef] [PubMed]
- Hodgson, J. Refreshing the biologic pipeline 2020. Nat. Biotechnol. 2021, 39, 135–143. [Google Scholar] [CrossRef]
- Allen, D.; Ruan, C.H.; King, B.; Ruan, K.H. Recent advances and near future of insulin production and therapy. Future Med. Chem. 2019, 11, 1513–1517. [Google Scholar] [CrossRef]
- Shakoor, S.; Kibble, E.; El-Jawhari, J.J. Bioengineering Approaches for Delivering Growth Factors: A Focus on Bone and Cartilage Regeneration. Bioengineering 2022, 9, 223. [Google Scholar] [CrossRef]
- Su, T.; Guan, Q.; Cheng, H.; Zhu, Z.; Jiang, C.; Guo, P.; Tai, Y.; Sun, H.; Wang, M.; Wei, W.; et al. Functions of G protein-coupled receptor 56 in health and disease. Acta Physiol. 2022, 236, e13866. [Google Scholar] [CrossRef]
- Yang, M.; Zhang, C.Y. G protein-coupled receptors as potential targets for nonalcoholic fatty liver disease treatment. World J. Gastroenterol. 2021, 27, 677–691. [Google Scholar] [CrossRef]
- Zhang, D.; Zhao, M.M.; Wu, J.M.; Wang, R.; Xue, G.; Xue, Y.B.; Shao, J.Q.; Zhang, Y.Y.; Dong, E.D.; Li, Z.Y.; et al. Dual-omics reveals temporal differences in acute sympathetic stress-induced cardiac inflammation following α(1) and β-adrenergic receptors activation. Acta Pharmacol. Sin. 2023, 44, 1350–1365. [Google Scholar] [CrossRef]
- Quiñones, M.; Fernø, J.; Diéguez, C.; Nogueiras, R.; Al-Massadi, O. Exciting advances in GPCR-based drugs discovery for treating metabolic disease and future perspectives. Exp. Opin. Drug Discov. 2019, 14, 421–431. [Google Scholar] [CrossRef] [PubMed]
- Lattanzi, R.; Miele, R. Special Issue “GPCR: Roles in Cell Development and Disease”. Int. J. Mol. Sci. 2023, 24, 7943. [Google Scholar] [CrossRef] [PubMed]
- Aldossary, H.S.; Alzahrani, A.A.; Nathanael, D.; Alhuthail, E.A.; Ray, C.J.; Batis, N.; Kumar, P.; Coney, A.M.; Holmes, A.P. G-Protein-Coupled Receptor (GPCR) Signaling in the Carotid Body: Roles in Hypoxia and Cardiovascular and Respiratory Disease. Int. J. Mol. Sci. 2020, 21, 6012. [Google Scholar] [CrossRef] [PubMed]
- Davenport, A.P.; Scully, C.C.G.; de Graaf, C.; Brown, A.J.H.; Maguire, J.J. Advances in therapeutic peptides targeting G protein-coupled receptors. Nat. Rev. Drug Discov. 2020, 19, 389–413. [Google Scholar] [CrossRef]
- Latendresse, G.; Elmore, C.; Deneris, A. Selective Serotonin Reuptake Inhibitors as First-Line Antidepressant Therapy for Perinatal Depression. J. Midwifery Womens Health 2017, 62, 317–328. [Google Scholar] [CrossRef]
- Sim, X.; Poncz, M.; Gadue, P.; French, D.L. Understanding platelet generation from megakaryocytes: Implications for in vitro-derived platelets. Blood 2016, 127, 1227–1233. [Google Scholar] [CrossRef]
- Nakamura, S.; Takayama, N.; Hirata, S.; Seo, H.; Endo, H.; Ochi, K.; Fujita, K.; Koike, T.; Harimoto, K.; Dohda, T.; et al. Expandable megakaryocyte cell lines enable clinically applicable generation of platelets from human induced pluripotent stem cells. Cell Stem Cell 2014, 14, 535–548. [Google Scholar] [CrossRef]
- Li, Y.; Li, Q.Y.; Ling, Q.L.; So, S.P.; Ruan, K.H. A novel single-chain enzyme complex with chain reaction properties rapidly producing thromboxane A(2) and exhibiting powerful anti-bleeding functions. J. Cell Mol. Med. 2019, 23, 8343–8354. [Google Scholar] [CrossRef]
- Ruan, K.H.; Cervantes, V.; So, S.P. Engineering of a novel hybrid enzyme: An anti-inflammatory drug target with triple catalytic activities directly converting arachidonic acid into the inflammatory prostaglandin E2. Protein Eng. Des. Sel. 2009, 22, 733–740. [Google Scholar] [CrossRef]
- Li, Q.-Y.; Li, Y.; Inoue, A.; Lu, R.; Xu, A.; Ruan, K.-H. Reversing thromboxane A2 receptor activity from calcium to cAMP signaling by shifting Gαq to Gαs covalently linked to the receptor. Biochem. Eng. J. 2022, 184, 108465. [Google Scholar] [CrossRef]
- Sadana, R.; Dessauer, C.W. Physiological roles for G protein-regulated adenylyl cyclase isoforms: Insights from knockout and overexpression studies. Neurosignals 2009, 17, 5–22. [Google Scholar] [CrossRef] [PubMed]
- De Cuyper, I.M.; Meinders, M.; van de Vijver, E.; de Korte, D.; Porcelijn, L.; de Haas, M.; Eble, J.A.; Seeger, K.; Rutella, S.; Pagliara, D.; et al. A novel flow cytometry-based platelet aggregation assay. Blood 2013, 121, e70–e80. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.; Chen, S.; Yuan, X.; Han, S.; Zhang, H.; Xia, W.; Xu, Y.; Zhao, Q.; Wu, B. Structural basis for ligand recognition of the human thromboxane A(2) receptor. Nat. Chem. Biol. 2019, 15, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Lyon, A.M.; Dutta, S.; Boguth, C.A.; Skiniotis, G.; Tesmer, J.J. Full-length Gα(q)-phospholipase C-β3 structure reveals interfaces of the C-terminal coiled-coil domain. Nat. Struct. Mol. Biol. 2013, 20, 355–362. [Google Scholar] [CrossRef]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 2007, 131, 861–872. [Google Scholar] [CrossRef]
- Feng, Q.; Shabrani, N.; Thon, J.N.; Huo, H.; Thiel, A.; Machlus, K.R.; Kim, K.; Brooks, J.; Li, F.; Luo, C.; et al. Scalable generation of universal platelets from human induced pluripotent stem cells. Stem Cell Rep. 2014, 3, 817–831. [Google Scholar] [CrossRef]
- Ditadi, A.; Sturgeon, C.M.; Keller, G. A view of human haematopoietic development from the Petri dish. Nat. Rev. Mol. Cell Biol. 2017, 18, 56–67. [Google Scholar] [CrossRef]
- Nandi, S.; Sproul, E.P.; Nellenbach, K.; Erb, M.; Gaffney, L.; Freytes, D.O.; Brown, A.C. Platelet-like particles dynamically stiffen fibrin matrices and improve wound healing outcomes. Biomater. Sci. 2019, 7, 669–682. [Google Scholar] [CrossRef]
- Chee, E.; Mihalko, E.; Nellenbach, K.; Sollinger, J.; Huang, K.; Hon, M.; Pandit, S.; Cheng, K.; Brown, A. Wound-triggered shape change microgels for the development of enhanced biomimetic function platelet-like particles. J. Biomed. Mater. Res. A 2023. [Google Scholar] [CrossRef]
- Gollomp, K.; Lambert, M.P.; Poncz, M. Current status of blood ‘pharming’: Megakaryoctye transfusions as a source of platelets. Curr. Opin. Hematol. 2017, 24, 565–571. [Google Scholar] [CrossRef]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, R.; Li, Y.; Xu, A.; King, B.; Ruan, K.-H. Reprogramming Megakaryocytes for Controlled Release of Platelet-like Particles Carrying a Single-Chain Thromboxane A2 Receptor-G-Protein Complex with Therapeutic Potential. Cells 2023, 12, 2775. https://doi.org/10.3390/cells12242775
Lu R, Li Y, Xu A, King B, Ruan K-H. Reprogramming Megakaryocytes for Controlled Release of Platelet-like Particles Carrying a Single-Chain Thromboxane A2 Receptor-G-Protein Complex with Therapeutic Potential. Cells. 2023; 12(24):2775. https://doi.org/10.3390/cells12242775
Chicago/Turabian StyleLu, Renzhong, Yan Li, Anna Xu, Bridgette King, and Ke-He Ruan. 2023. "Reprogramming Megakaryocytes for Controlled Release of Platelet-like Particles Carrying a Single-Chain Thromboxane A2 Receptor-G-Protein Complex with Therapeutic Potential" Cells 12, no. 24: 2775. https://doi.org/10.3390/cells12242775
APA StyleLu, R., Li, Y., Xu, A., King, B., & Ruan, K. -H. (2023). Reprogramming Megakaryocytes for Controlled Release of Platelet-like Particles Carrying a Single-Chain Thromboxane A2 Receptor-G-Protein Complex with Therapeutic Potential. Cells, 12(24), 2775. https://doi.org/10.3390/cells12242775