
The Role of Selected Adipocytokines in Ovarian Cancer and Endometrial Cancer
 , , and
, , and
Abstract
:1. Ovarian Cancer and Endometrial Cancer
1.1. Ovarian Cancer
1.2. Endometrial Cancer
2. Adipocytokines
3. The Role of Obesity in the Development of Cancer
4. Leptin
4.1. The Role of Leptin in Ovarian Cancer
4.2. The Role of Leptin in Endometrial Cancer
5. Adiponectin
5.1. The Role of Adiponectin in Ovarian Cancer
5.2. The Role of Adiponectin in Endometrial Cancer
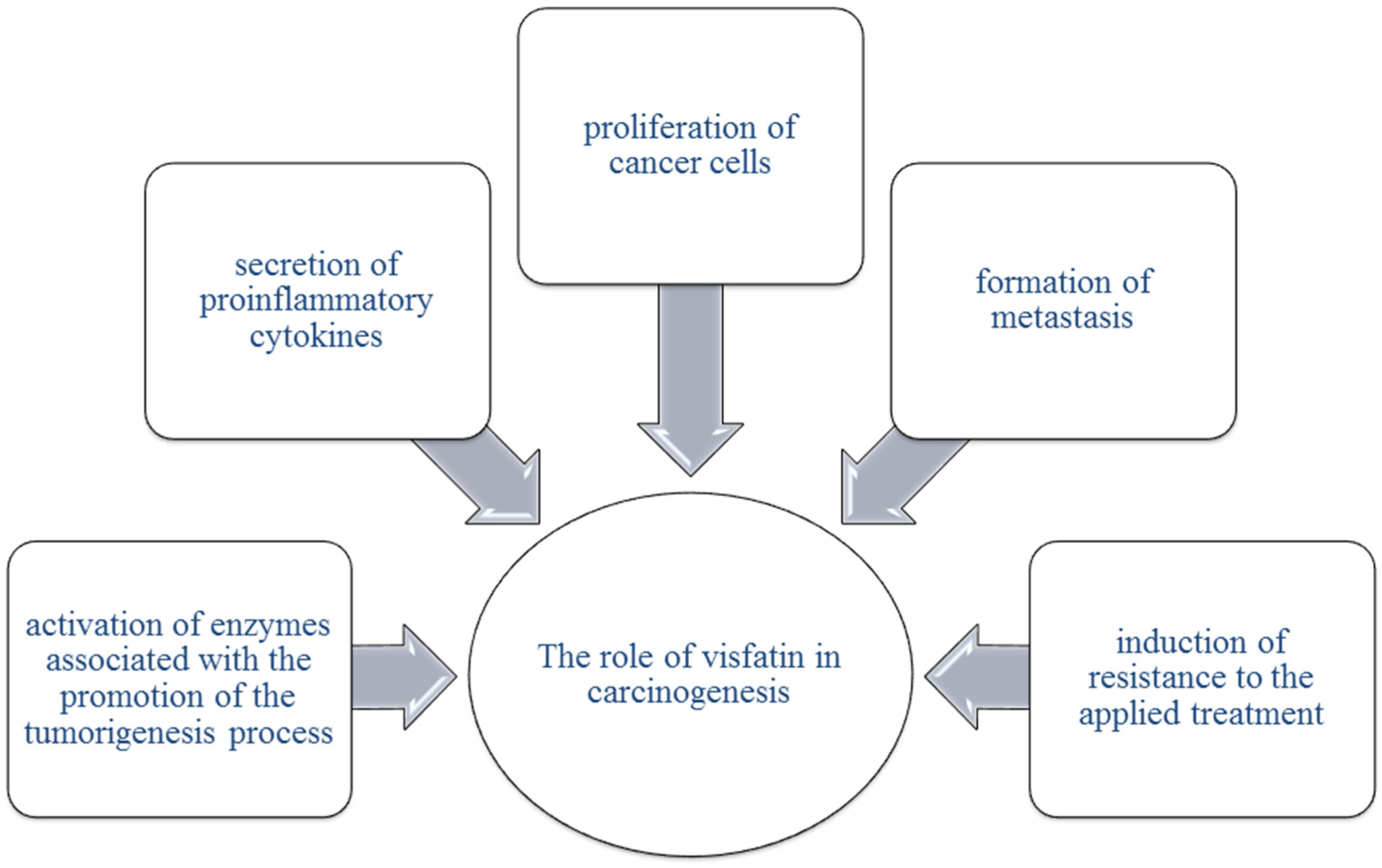
6. Visfatin
6.1. Visfatin in Ovarian Cancer
6.2. Visfatin in Endometrial Cancer
7. Resistin
7.1. Resistin in Ovarian Cancer
7.2. Resistin in Endometrial Cancer
8. Apelin
8.1. Apelin in Ovarian Cancer
8.2. Apelin in Endometrial Cancer
9. Chemerin
9.1. Chemerin in Ovarian Cancer
9.2. Chemerin in Endometrial Cancer
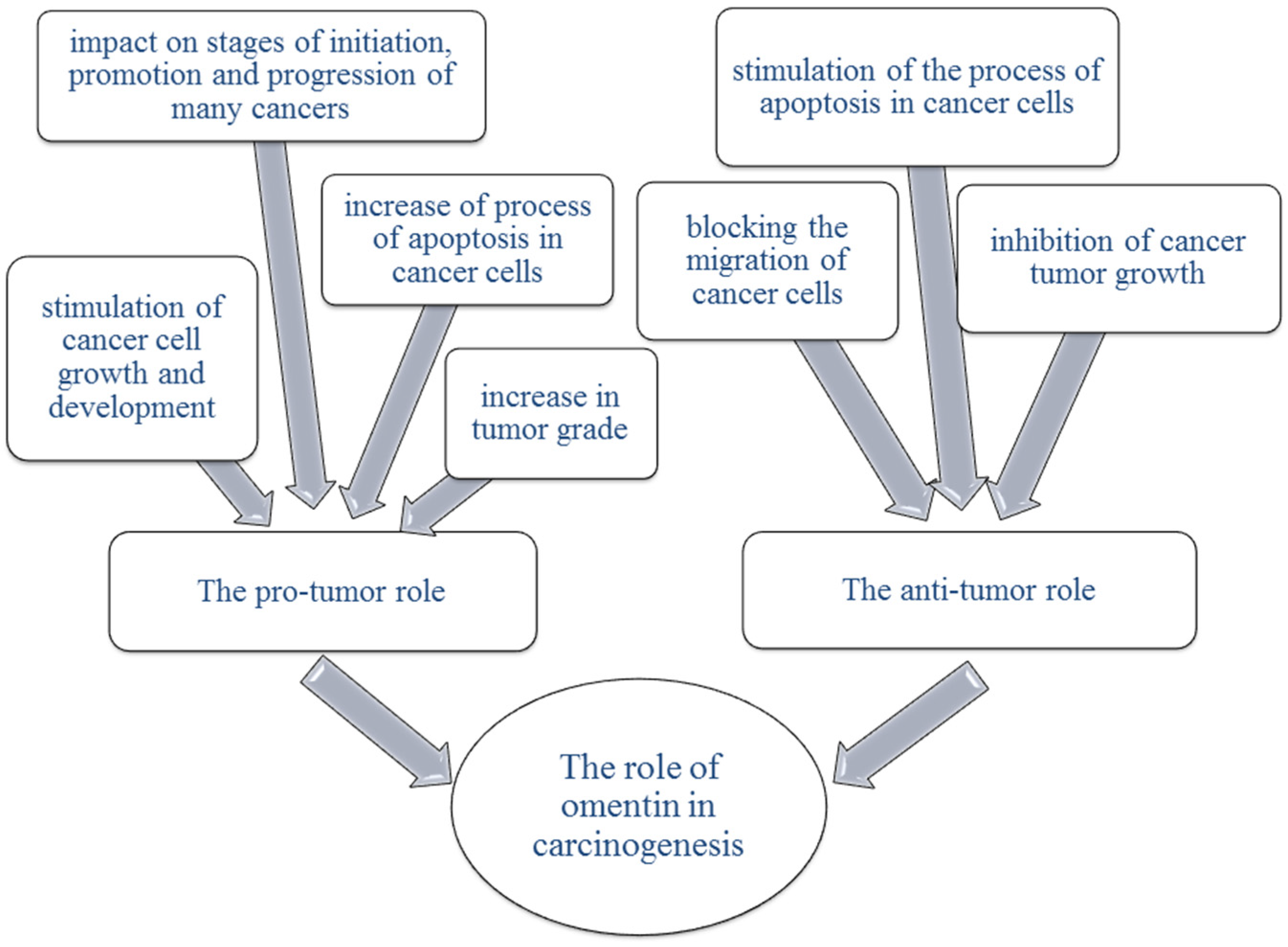
10. Omentin
10.1. Omentin in Ovarian Cancer
10.2. Omentin in Endometrial Cancer
11. Vaspin
11.1. Vaspin in Ovarian Cancer
11.2. Vaspin in Endometrial Cancer
12. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kossaï, M.; Leary, A.; Scoazec, J.-Y.; Genestie, C. Ovarian Cancer: A Heterogeneous Disease. Pathobiology 2018, 85, 41–49. [Google Scholar] [CrossRef]
- Lheureux, S.; Braunstein, M.; Oza, A.M. Epithelial Ovarian Cancer: Evolution of Management in the Era of Precision Medicine. CA Cancer J. Clin. 2019, 69, 280–304. [Google Scholar] [CrossRef] [Green Version]
- Nebgen, D.R.; Lu, K.H.; Bast, R.C. Novel Approaches to Ovarian Cancer Screening. Curr. Oncol. Rep. 2019, 21, 75. [Google Scholar] [CrossRef] [PubMed]
- Kroeger, P.T.; Drapkin, R. Pathogenesis and Heterogeneity of Ovarian Cancer. Curr. Opin. Obstet. Gynecol. 2017, 29, 26–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Natanzon, Y.; Goode, E.L.; Cunningham, J.M. Epigenetics in Ovarian Cancer. Semin. Cancer Biol. 2018, 51, 160–169. [Google Scholar] [CrossRef]
- Muñoz-Galván, S.; Carnero, A. Targeting Cancer Stem Cells to Overcome Therapy Resistance in Ovarian Cancer. Cells 2020, 9, 1402. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Xu, J.; Yu, J.; Yi, P. Shaping Immune Responses in the Tumor Microenvironment of Ovarian Cancer. Front. Immunol. 2021, 12, 692360. [Google Scholar] [CrossRef]
- Zhang, B.; Chen, F.; Xu, Q.; Han, L.; Xu, J.; Gao, L.; Sun, X.; Li, Y.; Li, Y.; Qian, M.; et al. Revisiting Ovarian Cancer Microenvironment: A Friend or a Foe? Protein Cell 2018, 9, 674–692. [Google Scholar] [CrossRef] [Green Version]
- Kim, B.; Park, Y.; Kim, B.; Ahn, H.J.; Lee, K.-A.; Chung, J.E.; Han, S.W. Diagnostic Performance of CA 125, HE4, and Risk of Ovarian Malignancy Algorithm for Ovarian Cancer. J. Clin. Lab. Anal. 2019, 33, e22624. [Google Scholar] [CrossRef] [Green Version]
- Chandra, A.; Pius, C.; Nabeel, M.; Nair, M.; Vishwanatha, J.K.; Ahmad, S.; Basha, R. Ovarian Cancer: Current Status and Strategies for Improving Therapeutic Outcomes. Cancer Med. 2019, 8, 7018–7031. [Google Scholar] [CrossRef] [Green Version]
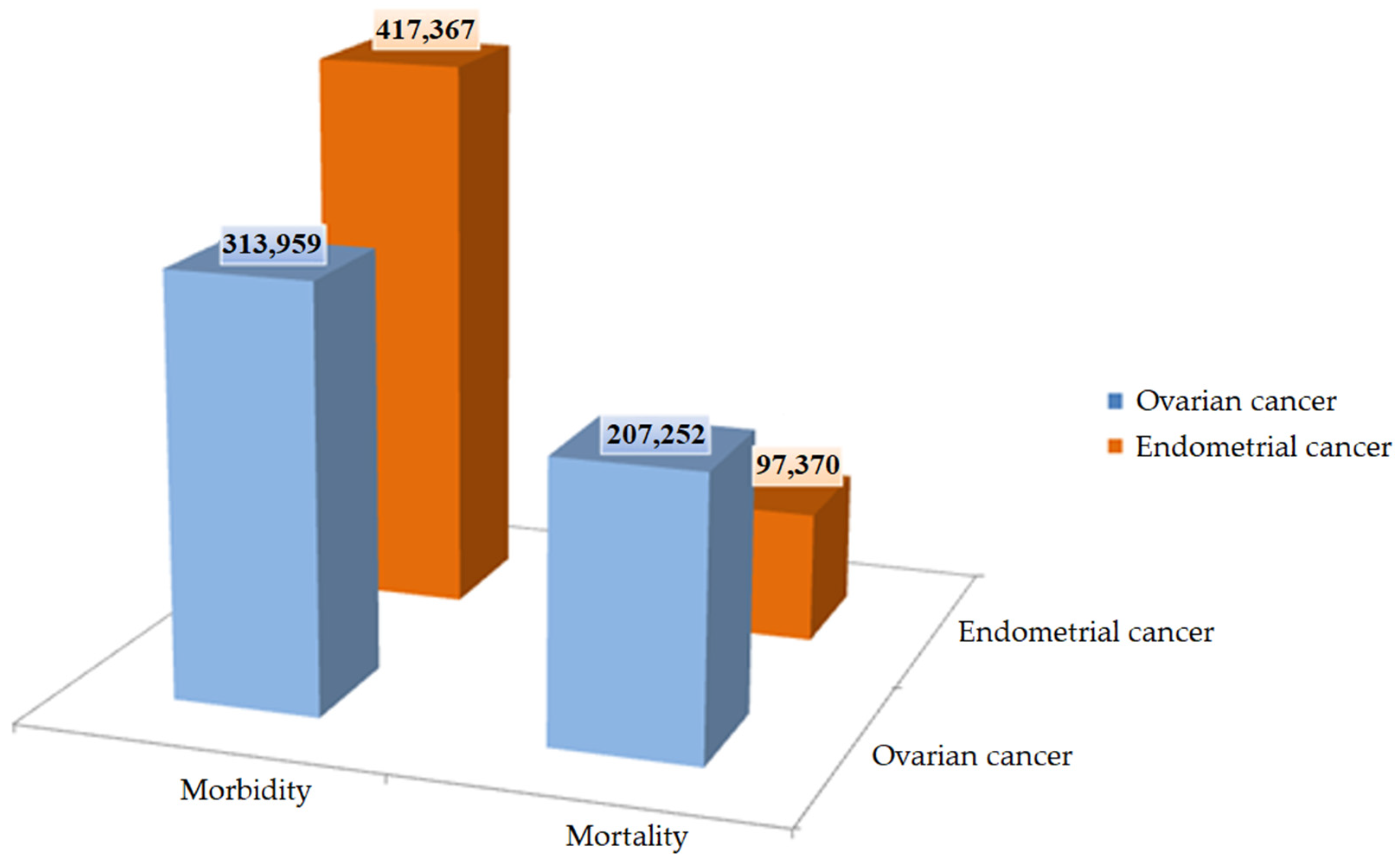
- Torre, L.A.; Trabert, B.; DeSantis, C.E.; Miller, K.D.; Samimi, G.; Runowicz, C.D.; Gaudet, M.M.; Jemal, A.; Siegel, R.L. Ovarian Cancer Statistics, 2018: Ovarian Cancer Statistics, 2018. CA Cancer J. Clin. 2018, 68, 284–296. [Google Scholar] [CrossRef] [Green Version]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Lin, Q.; Fei, H.; Xue, L.; Li, L.; Xi, Q.; Jiang, H. Bioinformatics Analysis of Potential Therapeutic Targets and Prognostic Biomarkers amid CXC Chemokines in Ovarian Carcinoma Microenvironment. J. Oncol. 2021, 2021, 8859554. [Google Scholar] [CrossRef] [PubMed]
- Cortez, A.J.; Tudrej, P.; Kujawa, K.A.; Lisowska, K.M. Advances in Ovarian Cancer Therapy. Cancer Chemother. Pharm. 2018, 81, 17–38. [Google Scholar] [CrossRef] [Green Version]
- Bogani, G.; Lopez, S.; Mantiero, M.; Ducceschi, M.; Bosio, S.; Ruisi, S.; Sarpietro, G.; Guerrisi, R.; Brusadelli, C.; Dell’Acqua, A.; et al. Immunotherapy for Platinum-Resistant Ovarian Cancer. Gynecol. Oncol. 2020, 158, 484–488. [Google Scholar] [CrossRef]
- Elias, K.M.; Guo, J.; Bast, R.C. Early Detection of Ovarian Cancer. Hematol. Oncol. Clin. N. Am. 2018, 32, 903–914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aleksandra Kujawa, K.; Lisowska, K.M. Ovarian Cancer–from Biology to Clinic. Postep. Hig. Med. Dosw. 2015, 69, 1275–1290. [Google Scholar] [CrossRef]
- Xiao, Y.; Bi, M.; Guo, H.; Li, M. Multi-Omics Approaches for Biomarker Discovery in Early Ovarian Cancer Diagnosis. eBioMedicine 2022, 79, 104001. [Google Scholar] [CrossRef]
- Zhang, R.; Siu, M.K.Y.; Ngan, H.Y.S.; Chan, K.K.L. Molecular Biomarkers for the Early Detection of Ovarian Cancer. Int. J. Mech. Sci. 2022, 23, 12041. [Google Scholar] [CrossRef]
- Lawson-Michod, K.A.; Watt, M.H.; Grieshober, L.; Green, S.E.; Karabegovic, L.; Derzon, S.; Owens, M.; McCarty, R.D.; Doherty, J.A.; Barnard, M.E. Pathways to Ovarian Cancer Diagnosis: A Qualitative Study. BMC Women’s Health 2022, 22, 430. [Google Scholar] [CrossRef]
- Kumari, S. Serum Biomarker Based Algorithms in Diagnosis of Ovarian Cancer: A Review. Ind. J. Clin. Biochem. 2018, 33, 382–386. [Google Scholar] [CrossRef]
- Singh, A.; Gupta, S.; Sachan, M. Epigenetic Biomarkers in the Management of Ovarian Cancer: Current Prospectives. Front. Cell Dev. Biol. 2019, 7, 182. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.; Gupta, S.; Badarukhiya, J.A.; Sachan, M. Detection of Aberrant Methylation of HOXA9 and HIC1 through Multiplex MethyLight Assay in Serum DNA for the Early Detection of Epithelial Ovarian Cancer. Int. J. Cancer 2020, 147, 1740–1752. [Google Scholar] [CrossRef]
- Liberto, J.M.; Chen, S.-Y.; Shih, I.-M.; Wang, T.-H.; Wang, T.-L.; Pisanic, T.R. Current and Emerging Methods for Ovarian Cancer Screening and Diagnostics: A Comprehensive Review. Cancers 2022, 14, 2885. [Google Scholar] [CrossRef]
- Braun, M.M.; Overbeek-Wager, E.A.; Grumbo, R.J. Diagnosis and Management of Endometrial Cancer. Am. Fam. Physician 2016, 93, 468–474. [Google Scholar] [PubMed]
- Barcellini, A.; Roccio, M.; Laliscia, C.; Zanellini, F.; Pettinato, D.; Valvo, F.; Mirandola, A.; Orlandi, E.; Gadducci, A. Endometrial Cancer: When Upfront Surgery Is Not an Option. Oncology 2021, 99, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Murali, R.; Soslow, R.A.; Weigelt, B. Classification of Endometrial Carcinoma: More than Two Types. Lancet Oncol. 2014, 15, e268–e278. [Google Scholar] [CrossRef] [PubMed]
- Banz-Jansen, C.; Helweg, L.P.; Kaltschmidt, B. Endometrial Cancer Stem Cells: Where Do We Stand and Where Should We Go? Int. J. Mech. Sci. 2022, 23, 3412. [Google Scholar] [CrossRef]
- McAlpine, J.N.; Gilks, C.B. Precision Medicine in Endometrial Cancer. Gynecol. Oncol. 2019, 154, 451–453. [Google Scholar] [CrossRef] [PubMed]
- Słabuszewska-Jóźwiak, A.; Lukaszuk, A.; Janicka-Kośnik, M.; Wdowiak, A.; Jakiel, G. Role of Leptin and Adiponectin in Endometrial Cancer. Int. J. Mech. Sci. 2022, 23, 5307. [Google Scholar] [CrossRef]
- Crosbie, E.J.; Kitson, S.J.; McAlpine, J.N.; Mukhopadhyay, A.; Powell, M.E.; Singh, N. Endometrial Cancer. Lancet 2022, 399, 1412–1428. [Google Scholar] [CrossRef]
- Clarke, M.A.; Long, B.J.; Del Mar Morillo, A.; Arbyn, M.; Bakkum-Gamez, J.N.; Wentzensen, N. Association of Endometrial Cancer Risk With Postmenopausal Bleeding in Women: A Systematic Review and Meta-Analysis. JAMA Intern. Med. 2018, 178, 1210. [Google Scholar] [CrossRef] [Green Version]
- Makker, V.; MacKay, H.; Ray-Coquard, I.; Levine, D.A.; Westin, S.N.; Aoki, D.; Oaknin, A. Endometrial Cancer. Nat. Rev. Dis. Prim. 2021, 7, 88. [Google Scholar] [CrossRef]
- Terzic, M.; Aimagambetova, G.; Kunz, J.; Bapayeva, G.; Aitbayeva, B.; Terzic, S.; Laganà, A.S. Molecular Basis of Endometriosis and Endometrial Cancer: Current Knowledge and Future Perspectives. Int. J. Mech. Sci. 2021, 22, 9274. [Google Scholar] [CrossRef]
- Yen, T.-T.; Wang, T.-L.; Fader, A.N.; Shih, I.-M.; Gaillard, S. Molecular Classification and Emerging Targeted Therapy in Endometrial Cancer. Int. J. Gynecol. Pathol. 2020, 39, 26–35. [Google Scholar] [CrossRef]
- Trojano, G.; Olivieri, C.; Tinelli, R.; Damiani, G.R.; Pellegrino, A.; Cicinelli, E. Conservative Treatment in Early Stage Endometrial Cancer: A Review. Acta Bio Med. Atenei Parm. 2019, 90, 405–410. [Google Scholar] [CrossRef]
- Passarello, K.; Kurian, S.; Villanueva, V. Endometrial Cancer: An Overview of Pathophysiology, Management, and Care. Semin. Oncol. Nurs. 2019, 35, 157–165. [Google Scholar] [CrossRef]
- Aoki, Y.; Kanao, H.; Wang, X.; Yunokawa, M.; Omatsu, K.; Fusegi, A.; Takeshima, N. Adjuvant Treatment of Endometrial Cancer Today. Jpn. J. Clin. Oncol. 2020, 50, 753–765. [Google Scholar] [CrossRef] [PubMed]
- Onstad, M.A.; Schmandt, R.E.; Lu, K.H. Addressing the Role of Obesity in Endometrial Cancer Risk, Prevention, and Treatment. J. Clin. Oncol. 2016, 34, 4225–4230. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.-S.; Scherer, P.E. Obesity, Diabetes, and Increased Cancer Progression. Diabetes Metab. J. 2021, 45, 799–812. [Google Scholar] [CrossRef] [PubMed]
- Ray, I.; Meira, L.B.; Michael, A.; Ellis, P.E. Adipocytokines and Disease Progression in Endometrial Cancer: A Systematic Review. Cancer Metastasis Rev. 2022, 41, 211–242. [Google Scholar] [CrossRef] [PubMed]
- Ellis, P.E.; Barron, G.A.; Bermano, G. Adipocytokines and Their Relationship to Endometrial Cancer Risk: A Systematic Review and Meta-Analysis. Gynecol. Oncol. 2020, 158, 507–516. [Google Scholar] [CrossRef]
- Shaw, E.; Farris, M.; McNeil, J.; Friedenreich, C. Obesity and Endometrial Cancer. In Obesity and Cancer; Pischon, T., Nimptsch, K., Eds.; Recent Results in Cancer Research; Springer International Publishing: Cham, Switzerland, 2016; Volume 208, pp. 107–136. ISBN 978-3-319-42540-5. [Google Scholar]
- Booth, A.; Magnuson, A.; Fouts, J.; Foster, M. Adipose Tissue, Obesity and Adipokines: Role in Cancer Promotion. Horm. Mol. Biol. Clin. Investig. 2015, 21, 57–74. [Google Scholar] [CrossRef] [PubMed]
- Cinti, S. Adipose Organ Development and Remodeling. In Comprehensive Physiology; Terjung, R., Ed.; Wiley: Hoboken, NJ, USA, 2018; pp. 1357–1431. ISBN 978-0-470-65071-4. [Google Scholar]
- Funcke, J.-B.; Scherer, P.E. Beyond Adiponectin and Leptin: Adipose Tissue-Derived Mediators of Inter-Organ Communication. J. Lipid Res. 2019, 60, 1648–1697. [Google Scholar] [CrossRef]
- Gao, S.-J.; Liu, D.-Q.; Li, D.-Y.; Sun, J.; Zhang, L.-Q.; Wu, J.-Y.; Song, F.-H.; Zhou, Y.-Q.; Mei, W. Adipocytokines: Emerging Therapeutic Targets for Pain Management. Biomed. Pharmacother. 2022, 149, 112813. [Google Scholar] [CrossRef]
- Maximus, P.S.; Al Achkar, Z.; Hamid, P.F.; Hasnain, S.S.; Peralta, C.A. Adipocytokines: Are They the Theory of Everything? Cytokine 2020, 133, 155144. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Han, X. Adipocytokines and Breast Cancer. Curr. Probl. Cancer 2018, 42, 208–214. [Google Scholar] [CrossRef] [PubMed]
- Zwick, R.K.; Guerrero-Juarez, C.F.; Horsley, V.; Plikus, M.V. Anatomical, Physiological, and Functional Diversity of Adipose Tissue. Cell Metab. 2018, 27, 68–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Rourke, R.W. Adipose Tissue and the Physiologic Underpinnings of Metabolic Disease. Surg. Obes. Relat. Dis. 2018, 14, 1755–1763. [Google Scholar] [CrossRef]
- Frigolet, M.E.; Gutiérrez-Aguilar, R. The colors of adipose tissue. Gac. Med. Mex. 2020, 156, 3932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pang, L.; Chang, X. Resistin Expression in Epithelial Ovarian Cancer Promotes the Proliferation and Migration of Ovarian Cancer Cells to Worsen Prognosis. J. Cancer 2021, 12, 6796–6804. [Google Scholar] [CrossRef]
- Wang, Y.-Y.; Hung, A.C.; Lo, S.; Yuan, S.-S.F. Adipocytokines Visfatin and Resistin in Breast Cancer: Clinical Relevance, Biological Mechanisms, and Therapeutic Potential. Cancer Lett. 2021, 498, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Petrelli, F.; Cortellini, A.; Indini, A.; Tomasello, G.; Ghidini, M.; Nigro, O.; Salati, M.; Dottorini, L.; Iaculli, A.; Varricchio, A.; et al. Association of Obesity With Survival Outcomes in Patients with Cancer: A Systematic Review and Meta-Analysis. JAMA Netw. Open 2021, 4, e213520. [Google Scholar] [CrossRef]
- Friedenreich, C.M.; Ryder-Burbidge, C.; McNeil, J. Physical Activity, Obesity and Sedentary Behavior in Cancer Etiology: Epidemiologic Evidence and Biologic Mechanisms. Mol. Oncol. 2021, 15, 790–800. [Google Scholar] [CrossRef] [PubMed]
- Lauby-Secretan, B.; Scoccianti, C.; Loomis, D.; Grosse, Y.; Bianchini, F.; Straif, K. Body Fatness and Cancer—Viewpoint of the IARC Working Group. N. Engl. J. Med. 2016, 375, 794–798. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Giffney, H.E.; Arthur, R.S.; Rohan, T.E.; Dannenberg, A.J. Cancer Risk in Normal Weight Individuals with Metabolic Obesity: A Narrative Review. Cancer Prev. Res. 2021, 14, 509–520. [Google Scholar] [CrossRef]
- Avgerinos, K.I.; Spyrou, N.; Mantzoros, C.S.; Dalamaga, M. Obesity and Cancer Risk: Emerging Biological Mechanisms and Perspectives. Metabolism 2019, 92, 121–135. [Google Scholar] [CrossRef]
- Kany, S.; Vollrath, J.T.; Relja, B. Cytokines in Inflammatory Disease. Int. J. Mech. Sci. 2019, 20, 6008. [Google Scholar] [CrossRef] [Green Version]
- Di Zazzo, E.; Polito, R.; Bartollino, S.; Nigro, E.; Porcile, C.; Bianco, A.; Daniele, A.; Moncharmont, B. Adiponectin as Link Factor between Adipose Tissue and Cancer. Int. J. Mech. Sci. 2019, 20, 839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gelsomino, L.; Naimo, G.D.; Catalano, S.; Mauro, L.; Andò, S. The Emerging Role of Adiponectin in Female Malignancies. Int. J. Mech. Sci. 2019, 20, 2127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kompella, P.; Vasquez, K.M. Obesity and Cancer: A Mechanistic Overview of Metabolic Changes in Obesity That Impact Genetic Instability. Mol. Carcinog. 2019, 58, 1531–1550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Włodarczyk, M.; Nowicka, G. Obesity, DNA Damage, and Development of Obesity-Related Diseases. Int. J. Mech. Sci. 2019, 20, 1146. [Google Scholar] [CrossRef]
- Gee, M.E.; Faraahi, Z.; McCormick, A.; Edmondson, R.J. DNA Damage Repair in Ovarian Cancer: Unlocking the Heterogeneity. J. Ovarian Res. 2018, 11, 50. [Google Scholar] [CrossRef] [Green Version]
- Cuccu, I.; D’Oria, O.; Sgamba, L.; De Angelis, E.; Golia D’Augè, T.; Turetta, C.; Di Dio, C.; Scudo, M.; Bogani, G.; Di Donato, V.; et al. Role of Genomic and Molecular Biology in the Modulation of the Treatment of Endometrial Cancer: Narrative Review and Perspectives. Healthcare 2023, 11, 571. [Google Scholar] [CrossRef]
- Roy, B.; Palaniyandi, S.S. Tissue-Specific Role and Associated Downstream Signaling Pathways of Adiponectin. Cell Biosci. 2021, 11, 77. [Google Scholar] [CrossRef]
- Qu, A.; Shah, Y.M.; Matsubara, T.; Yang, Q.; Gonzalez, F.J. PPARα-Dependent Activation of Cell Cycle Control and DNA Repair Genes in Hepatic Nonparenchymal Cells. Toxicol. Sci. 2010, 118, 404–410. [Google Scholar] [CrossRef]
- Wilk, A.; Hayat, F.; Cunningham, R.; Li, J.; Garavaglia, S.; Zamani, L.; Ferraris, D.M.; Sykora, P.; Andrews, J.; Clark, J.; et al. Extracellular NAD+ Enhances PARP-Dependent DNA Repair Capacity Independently of CD73 Activity. Sci. Rep. 2020, 10, 651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, B.; Deng, X.; Sun, Y.; Bai, L.; Xiahou, Z.; Cong, Y.; Xu, X. Nampt Is Involved in DNA Double-Strand Break Repair. Chin. J. Cancer 2012, 31, 392–398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Modzelewska, P.; Chludzińska, S.; Lewko, J.; Reszeć, J. The Influence of Leptin on the Process of Carcinogenesis. Contemp. Oncol. 2019, 23, 63–68. [Google Scholar] [CrossRef]
- Kato, S.; Abarzua-Catalan, L.; Trigo, C.; Delpiano, A.; Sanhueza, C.; García, K.; Ibañez, C.; Hormazábal, K.; Diaz, D.; Brañes, J.; et al. Leptin Stimulates Migration and Invasion and Maintains Cancer Stem-like Properties in Ovarian Cancer Cells: An Explanation for Poor Outcomes in Obese Women. Oncotarget 2015, 6, 21100–21119. [Google Scholar] [CrossRef] [Green Version]
- Crean-Tate, K.K.; Reizes, O. Leptin Regulation of Cancer Stem Cells in Breast and Gynecologic Cancer. Endocrinology 2018, 159, 3069–3080. [Google Scholar] [CrossRef]
- Kokts-Porietis, R.L.; Elmrayed, S.; Brenner, D.R.; Friedenreich, C.M. Obesity and Mortality among Endometrial Cancer Survivors: A Systematic Review and Meta-analysis. Obes. Rev. 2021, 22, e13337. [Google Scholar] [CrossRef]
- Donkers, H.; Fasmer, K.E.; Mcgrane, J.; Pijnenborg, J.M.A.; Bekkers, R.; Haldorsen, I.S.; Galaal, K. Obesity and Visceral Fat: Survival Impact in High-Grade Endometrial Cancer. Eur. J. Obstet. Gynecol. Reprod. Biol. 2021, 256, 425–432. [Google Scholar] [CrossRef]
- Thrastardottir, T.O.; Copeland, V.J.; Constantinou, C. The Association Between Nutrition, Obesity, Inflammation, and Endometrial Cancer: A Scoping Review. Curr. Nutr. Rep. 2022, 12, 98–121. [Google Scholar] [CrossRef] [PubMed]
- Assidi, M.; Yahya, F.M.; Al-Zahrani, M.H.; Elkhatib, R.; Zari, A.; Elaimi, A.; Al-Maghrabi, J.; Dallol, A.; Buhmeida, A.; Abu-Elmagd, M. Leptin Protein Expression and Promoter Methylation in Ovarian Cancer: A Strong Prognostic Value with Theranostic Promises. Int. J. Mech. Sci. 2021, 22, 12872. [Google Scholar] [CrossRef]
- Dixon, S.C.; Nagle, C.M.; Thrift, A.P.; Pharoah, P.D.; Pearce, C.L.; Zheng, W.; Painter, J.N.; AOCS Group & Australian Cancer Study (Ovarian Cancer); Chenevix-Trench, G.; Fasching, P.A.; et al. Adult Body Mass Index and Risk of Ovarian Cancer by Subtype: A Mendelian Randomization Study. Int. J. Epidemiol. 2016, 45, 884–895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, L.; Gajarawala, S.; McCrary, M.D. Endometrial Cancer and Obesity: Addressing the Awkward Silence. JAAPA 2023, 36, 28–31. [Google Scholar] [CrossRef] [PubMed]
- Kho, P.; Amant, F.; Annibali, D.; Ashton, K.; Attia, J.; Auer, P.L.; Beckmann, M.W.; Black, A.; Brinton, L.; Buchanan, D.D.; et al. Mendelian Randomization Analyses Suggest a Role for Cholesterol in the Development of Endometrial Cancer. Int. J. Cancer 2021, 148, 307–319. [Google Scholar] [CrossRef]
- Childs, G.V.; Odle, A.K.; MacNicol, M.C.; MacNicol, A.M. The Importance of Leptin to Reproduction. Endocrinology 2021, 162, bqaa204. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Proenca, R.; Maffei, M.; Barone, M.; Leopold, L.; Friedman, J.M. Positional Cloning of the Mouse Obese Gene and Its Human Homologue. Nature 1994, 372, 425–432. [Google Scholar] [CrossRef] [PubMed]
- Obradovic, M.; Sudar-Milovanovic, E.; Soskic, S.; Essack, M.; Arya, S.; Stewart, A.J.; Gojobori, T.; Isenovic, E.R. Leptin and Obesity: Role and Clinical Implication. Front. Endocrinol. 2021, 12, 585887. [Google Scholar] [CrossRef] [PubMed]
- Lustig, R.H.; Fennoy, I. The History of Obesity Research. Horm. Res. Paediatr. 2022, 95, 638–648. [Google Scholar] [CrossRef]
- Park, H.-K.; Ahima, R.S. Physiology of Leptin: Energy Homeostasis, Neuroendocrine Function and Metabolism. Metabolism 2015, 64, 24–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Münzberg, H.; Morrison, C.D. Structure, Production and Signaling of Leptin. Metabolism 2015, 64, 13–23. [Google Scholar] [CrossRef] [Green Version]
- Greco, M.; De Santo, M.; Comandè, A.; Belsito, E.L.; Andò, S.; Liguori, A.; Leggio, A. Leptin-Activity Modulators and Their Potential Pharmaceutical Applications. Biomolecules 2021, 11, 1045. [Google Scholar] [CrossRef]
- Casado, M.E.; Collado-Pérez, R.; Frago, L.M.; Barrios, V. Recent Advances in the Knowledge of the Mechanisms of Leptin Physiology and Actions in Neurological and Metabolic Pathologies. Int. J. Mech. Sci. 2023, 24, 1422. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Pérez, A.; Sánchez-Jiménez, F.; Vilariño-García, T.; Sánchez-Margalet, V. Role of Leptin in Inflammation and Vice Versa. Int. J. Mech. Sci. 2020, 21, 5887. [Google Scholar] [CrossRef]
- Zhang, F.; Chen, Y.; Heiman, M.; DiMarchi, R. Leptin: Structure, Function and Biology. In Vitamins & Hormones; Elsevier: Amsterdam, The Netherlands, 2005; Volume 71, pp. 345–372. ISBN 978-0-12-709871-5. [Google Scholar]
- Münzberg, H.; Singh, P.; Heymsfield, S.B.; Yu, S.; Morrison, C.D. Recent Advances in Understanding the Role of Leptin in Energy Homeostasis. F1000Research 2020, 9, 451. [Google Scholar] [CrossRef]
- Farr, O.M.; Gavrieli, A.; Mantzoros, C.S. Leptin Applications in 2015: What Have We Learned about Leptin and Obesity? Curr. Opin. Endocrinol. Diabetes Obes. 2015, 22, 353–359. [Google Scholar] [CrossRef] [Green Version]
- Chin, Y.-T.; Wang, L.-M.; Hsieh, M.-T.; Shih, Y.-J.; Nana, A.W.; Changou, C.A.; Yang, Y.-C.S.H.; Chiu, H.-C.; Fu, E.; Davis, P.J.; et al. Leptin OB3 Peptide Suppresses Leptin-Induced Signaling and Progression in Ovarian Cancer Cells. J. Biomed. Sci. 2017, 24, 51. [Google Scholar] [CrossRef] [Green Version]
- Akinyemiju, T.; Moore, J.X.; Judd, S.E.; Pisu, M.; Goodman, M.; Howard, V.J.; Long, L.; Safford, M.; Gilchrist, S.C.; Cushman, M. Pre-Diagnostic Biomarkers of Metabolic Dysregulation and Cancer Mortality. Oncotarget 2018, 9, 16099–16109. [Google Scholar] [CrossRef] [Green Version]
- Fischer, A.W.; Cannon, B.; Nedergaard, J. Leptin: Is It Thermogenic? Endocr. Rev. 2020, 41, 232–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, A.W.; Hoefig, C.S.; Abreu-Vieira, G.; de Jong, J.M.A.; Petrovic, N.; Mittag, J.; Cannon, B.; Nedergaard, J. Leptin Raises Defended Body Temperature without Activating Thermogenesis. Cell Rep. 2016, 14, 1621–1631. [Google Scholar] [CrossRef] [Green Version]
- Fischer, A.W.; Cannon, B.; Nedergaard, J. Leptin-Deficient Mice Are Not Hypothermic, They Are Anapyrexic. Mol. Metab. 2017, 6, 173. [Google Scholar] [CrossRef] [PubMed]
- Deem, J.D.; Muta, K.; Ogimoto, K.; Nelson, J.T.; Velasco, K.R.; Kaiyala, K.J.; Morton, G.J. Leptin Regulation of Core Body Temperature Involves Mechanisms Independent of the Thyroid Axis. Am. J. Physiol.-Endocrinol. Metab. 2018, 315, E552–E564. [Google Scholar] [CrossRef] [PubMed]
- Kaiyala, K.J.; Ogimoto, K.; Nelson, J.T.; Muta, K.; Morton, G.J. Physiological Role for Leptin in the Control of Thermal Conductance. Mol. Metab. 2016, 5, 892–902. [Google Scholar] [CrossRef] [PubMed]
- Kaiyala, K.J.; Ogimoto, K.; Nelson, J.T.; Schwartz, M.W.; Morton, G.J. Leptin Signaling Is Required for Adaptive Changes in Food Intake, but Not Energy Expenditure, in Response to Different Thermal Conditions. PLoS ONE 2015, 10, e0119391. [Google Scholar] [CrossRef]
- Boroń, D.; Nowakowski, R.; Grabarek, B.O.; Zmarzły, N.; Opławski, M. Expression Pattern of Leptin and Its Receptors in Endometrioid Endometrial Cancer. J. Clin. Med. 2021, 10, 2787. [Google Scholar] [CrossRef]
- Chen, Y.-C.; Chien, C.-Y.; Hsu, C.-C.; Lee, C.-H.; Chou, Y.-T.; Shiah, S.-G.; Liu, S.-Y.; Yen, C.-Y.; Hsieh, A.C.-T.; Wabitsch, M.; et al. Obesity-Associated Leptin Promotes Chemoresistance in Colorectal Cancer through YAP-Dependent AXL Upregulation. Am. J. Cancer Res. 2021, 11, 4220–4240. [Google Scholar]
- Harbuzariu, A.; Rampoldi, A.; Daley-Brown, D.S.; Candelaria, P.; Harmon, T.L.; Lipsey, C.C.; Beech, D.J.; Quarshie, A.; Ilies, G.O.; Gonzalez-Perez, R.R. Leptin-Notch Signaling Axis Is Involved in Pancreatic Cancer Progression. Oncotarget 2017, 8, 7740–7752. [Google Scholar] [CrossRef] [Green Version]
- Barone, I.; Giordano, C. Leptin and Beyond: Actors in Cancer. Biomolecules 2021, 11, 1836. [Google Scholar] [CrossRef] [PubMed]
- Ray, A.; Fornsaglio, J.; Dogan, S.; Hedau, S.; Naik, D.; De, A. Gynaecological Cancers and Leptin: A Focus on the Endometrium and Ovary. Facts Views Vis. Obgyn 2018, 10, 5–18. [Google Scholar] [PubMed]
- Gu, F.; Zhang, H.; Yao, L.; Jiang, S.; Lu, H.; Xing, X.; Zhang, C.; Jiang, P.; Zhang, R. Leptin Contributes to the Taxol Chemoresistance in Epithelial Ovarian Cancer. Oncol. Lett. 2019, 18, 561–570. [Google Scholar] [CrossRef] [Green Version]
- Jiménez-Cortegana, C.; López-Saavedra, A.; Sánchez-Jiménez, F.; Pérez-Pérez, A.; Castiñeiras, J.; Virizuela-Echaburu, J.A.; de la Cruz-Merino, L.; Sánchez-Margalet, V. Leptin, Both Bad and Good Actor in Cancer. Biomolecules 2021, 11, 913. [Google Scholar] [CrossRef] [PubMed]
- Dupont, J.; Reverchon, M.; Cloix, L.; Froment, P.; Ramé, C. Involvement of Adipokines, AMPK, PI3K and the PPAR Signaling Pathways in Ovarian Follicle Development and Cancer. Int. J. Dev. Biol. 2012, 56, 959–967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, T.-C.; Hsiao, M. Leptin and Cancer: Updated Functional Roles in Carcinogenesis, Therapeutic Niches, and Developments. Int. J. Mech. Sci. 2021, 22, 2870. [Google Scholar] [CrossRef]
- Pham, D.-V.; Park, P.-H. Tumor Metabolic Reprogramming by Adipokines as a Critical Driver of Obesity-Associated Cancer Progression. Int. J. Mech. Sci. 2021, 22, 1444. [Google Scholar] [CrossRef]
- Uddin, S.; Bu, R.; Ahmed, M.; Abubaker, J.; Al-Dayel, F.; Bavi, P.; Al-Kuraya, K.S. Overexpression of Leptin Receptor Predicts an Unfavorable Outcome in Middle Eastern Ovarian Cancer. Mol. Cancer 2009, 8, 74. [Google Scholar] [CrossRef] [Green Version]
- Matte, I.; Garde-Granger, P.; Bessette, P.; Piché, A. Serum CA125 and Ascites Leptin Level Ratio Predicts Baseline Clinical Resistance to First-Line Platinum-Based Treatment and Poor Prognosis in Patients with High Grade Serous Ovarian Cancer. Am. J. Cancer Res. 2019, 9, 160–170. [Google Scholar]
- Feng, Z.; Wen, H.; Ju, X.; Bi, R.; Chen, X.; Yang, W.; Wu, X. Expression of Hypothalamic-Pituitary-Gonadal Axis-Related Hormone Receptors in Low-Grade Serous Ovarian Cancer (LGSC). J. Ovarian Res. 2017, 10, 7. [Google Scholar] [CrossRef] [Green Version]
- Kukla, A.; Piotrowska, K.; Misiek, M.; Chudecka-Glaz, A.M. Role of Adipokines in Ovarian Cancer Epidemiology and Prognosis. Ginekol. Pol. 2022, 93, 496–500. [Google Scholar] [CrossRef]
- Zhao, S.; Kusminski, C.M.; Scherer, P.E. Adiponectin, Leptin and Cardiovascular Disorders. Circ. Res. 2021, 128, 136–149. [Google Scholar] [CrossRef]
- Peng, J.; Chen, Q.; Wu, C. The Role of Adiponectin in Cardiovascular Disease. Cardiovasc. Pathol. 2023, 64, 107514. [Google Scholar] [CrossRef]
- Dezonne, R.S.; Pereira, C.M.; de Moraes Martins, C.J.; de Abreu, V.G.; Francischetti, E.A. Adiponectin, the Adiponectin Paradox, and Alzheimer’s Disease: Is This Association Biologically Plausible? Metab. Brain Dis. 2023, 38, 109–121. [Google Scholar] [CrossRef]
- Khoramipour, K.; Chamari, K.; Hekmatikar, A.A.; Ziyaiyan, A.; Taherkhani, S.; Elguindy, N.M.; Bragazzi, N.L. Adiponectin: Structure, Physiological Functions, Role in Diseases, and Effects of Nutrition. Nutrients 2021, 13, 1180. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.M.; Doss, H.M.; Kim, K.S. Multifaceted Physiological Roles of Adiponectin in Inflammation and Diseases. Int. J. Mech. Sci. 2020, 21, 1219. [Google Scholar] [CrossRef] [Green Version]
- Hebbard, L.; Ranscht, B. Multifaceted Roles of Adiponectin in Cancer. Best Pract. Res. Clin. Endocrinol. Metab. 2014, 28, 59–69. [Google Scholar] [CrossRef] [Green Version]
- Rizzo, M.R.; Fasano, R.; Paolisso, G. Adiponectin and Cognitive Decline. Int. J. Mech. Sci. 2020, 21, 2010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffmann, M.; Gogola, J.; Ptak, A. Adiponectin Reverses the Proliferative Effects of Estradiol and IGF-1 in Human Epithelial Ovarian Cancer Cells by Downregulating the Expression of Their Receptors. Horm. Cancer 2018, 9, 166–174. [Google Scholar] [CrossRef] [Green Version]
- Da Silva Rosa, S.C.; Liu, M.; Sweeney, G. Adiponectin Synthesis, Secretion and Extravasation from Circulation to Interstitial Space. Physiology 2021, 36, 134–149. [Google Scholar] [CrossRef] [PubMed]
- Li, H.-Y.; Hong, X.; Cao, Q.-Q.; So, K.-F. Adiponectin, Exercise and Eye Diseases. In International Review of Neurobiology; Elsevier: Amsterdam, The Netherlands, 2019; Volume 147, pp. 281–294. ISBN 978-0-12-816967-4. [Google Scholar]
- Wang, Z.V.; Scherer, P.E. Adiponectin, the Past Two Decades. J. Mol. Cell Biol. 2016, 8, 93–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shklyaev, S.S.; Melnichenko, G.A.; Volevodz, N.N.; Falaleeva, N.A.; Ivanov, S.A.; Kaprin, A.D.; Mokrysheva, N.G. Adiponectin: A Pleiotropic Hormone with Multifaceted Roles. Probl. Endokrinol. 2021, 67, 98–112. [Google Scholar] [CrossRef]
- Parida, S.; Siddharth, S.; Sharma, D. Adiponectin, Obesity, and Cancer: Clash of the Bigwigs in Health and Disease. Int. J. Mech. Sci. 2019, 20, 2519. [Google Scholar] [CrossRef] [Green Version]
- Samant, N.P.; Gupta, G.L. Adiponectin: A Potential Target for Obesity-Associated Alzheimer’s Disease. Metab. Brain Dis. 2021, 36, 1565–1572. [Google Scholar] [CrossRef]
- Kita, S.; Shimomura, I. Stimulation of Exosome Biogenesis by Adiponectin, a Circulating Factor Secreted from Adipocytes. J. Biochem. 2021, 169, 173–179. [Google Scholar] [CrossRef]
- Katira, A.; Tan, P.H. Evolving Role of Adiponectin in Cancer-Controversies and Update. Cancer Biol. Med. 2016, 13, 101–119. [Google Scholar] [CrossRef] [Green Version]
- Tumminia, A.; Vinciguerra, F.; Parisi, M.; Graziano, M.; Sciacca, L.; Baratta, R.; Frittitta, L. Adipose Tissue, Obesity and Adiponectin: Role in Endocrine Cancer Risk. Int. J. Mech. Sci. 2019, 20, 2863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Wen, K.; Han, X.; Liu, R.; Qu, Q. Adiponectin Mediates Antiproliferative and Apoptotic Responses in Endometrial Carcinoma by the AdipoRs/AMPK Pathway. Gynecol. Oncol. 2015, 137, 311–320. [Google Scholar] [CrossRef]
- Gupta, R.K.; Dholariya, S.J.; Kaushik, S.; Gupta, S.K.; Tripathi, R.; Jain, S.L. Hyperinsulinemia and Hypoadiponectinemia Are Associated with Increased Risk for Occurrence of Ovarian Cancer in Non-Diabetic Women of North Indian Population. Ind. J. Clin. Biochem. 2021, 36, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Diaz, E.S.; Karlan, B.Y.; Li, A.J. Obesity-Associated Adipokines Correlate with Survival in Epithelial Ovarian Cancer. Gynecol. Oncol. 2013, 129, 353–357. [Google Scholar] [CrossRef]
- Słomian, G.J.; Nowak, D.; Buczkowska, M.; Głogowska-Gruszka, A.; Słomian, S.P.; Roczniak, W.; Janyga, S.; Nowak, P. The Role of Adiponectin and Leptin in the Treatment of Ovarian Cancer Patients. Endokrynol. Pol. 2019, 70, 57–63. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Yu, Z.; Fang, L.; Liu, F.; Jiang, K. Expression of Adiponectin Receptor-1 and Prognosis of Epithelial Ovarian Cancer Patients. Med. Sci. Monit. 2017, 23, 1514–1521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Gao, S.; Sun, C.; Li, J.; Gao, W.; Yu, L. Clinical Significance of Serum Adiponectin and Visfatin Levels in Endometrial Cancer. Int. J. Gynecol. Obs. 2019, 145, 34–39. [Google Scholar] [CrossRef]
- Kölbl, H.; Bartl, T. Obesity in Gynecologic Oncology. Geburtshilfe Frauenheilkd. 2020, 80, 1205–1211. [Google Scholar] [CrossRef] [PubMed]
- Dashti, S.G.; English, D.R.; Simpson, J.A.; Karahalios, A.; Moreno-Betancur, M.; Biessy, C.; Rinaldi, S.; Ferrari, P.; Tjønneland, A.; Halkjær, J.; et al. Adiposity and Endometrial Cancer Risk in Postmenopausal Women: A Sequential Causal Mediation Analysis. Cancer Epidemiol. Biomark. Prev. 2021, 30, 104–113. [Google Scholar] [CrossRef] [PubMed]
- Al Abdulsalam, E.A.; Al Harithy, R.N. Visfatin and Global Histone H3K9me Levels in Colon Cancer. Ann. Med. 2021, 53, 647–652. [Google Scholar] [CrossRef]
- Dakroub, A.; Nasser, S.A.; Younis, N.; Bhagani, H.; Al-Dhaheri, Y.; Pintus, G.; Eid, A.A.; El-Yazbi, A.F.; Eid, A.H. Visfatin: A Possible Role in Cardiovasculo-Metabolic Disorders. Cells 2020, 9, 2444. [Google Scholar] [CrossRef] [PubMed]
- Ismaiel, A.; Leucuta, D.-C.; Popa, S.-L.; Dumitrascu, D.L. Serum Visfatin Levels in Nonalcoholic Fatty Liver Disease and Liver Fibrosis: Systematic Review and Meta-Analysis. J. Clin. Med. 2021, 10, 3029. [Google Scholar] [CrossRef]
- AL-Suhaimi, E.A.; Shehzad, A. Leptin, Resistin and Visfatin: The Missing Link between Endocrine Metabolic Disorders and Immunity. Eur. J. Med. Res. 2013, 18, 12. [Google Scholar] [CrossRef] [Green Version]
- Abdalla, M.M.I. Role of Visfatin in Obesity-Induced Insulin Resistance. World J. Clin. Cases 2022, 10, 10840–10851. [Google Scholar] [CrossRef]
- Bułdak, R.J.; Polaniak, R.; Kukla, M.; Zwirska-Korczala, K. Visfatin—An Enzyme, a Cytokine or an Adipokine? Biological Functions of Visfatin in Vitro. Endokrynol. Otył. Zab. Przem. Mat. 2011, 7, 16–24. [Google Scholar]
- Parimelazhagan, R.; Umapathy, D.; Sivakamasundari, I.R.; Sethupathy, S.; Ali, D.; Kunka Mohanram, R.; Namasivayan, N. Association between Tumor Prognosis Marker Visfatin and Proinflammatory Cytokines in Hypertensive Patients. BioMed Res. Int. 2021, 2021, 8568926. [Google Scholar] [CrossRef] [PubMed]
- Reverchon, M.; Ramé, C.; Bertoldo, M.; Dupont, J. Adipokines and the Female Reproductive Tract. Int. J. Endocrinol. 2014, 2014, 232454. [Google Scholar] [CrossRef] [PubMed]
- Pazgan-Simon, M.; Kukla, M.; Zuwała-Jagiełło, J.; Derra, A.; Bator, M.; Menżyk, T.; Lekstan, A.; Grzebyk, E.; Simon, K. Serum Visfatin and Vaspin Levels in Hepatocellular Carcinoma (HCC). PLoS ONE 2020, 15, e0227459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, D.; Wang, Y.; Liu, M.; Cheng, J.; Liu, Z.; Song, Y.; Du, J. Visfatin Promotes Angiogenesis of RF/6A Cells through Upregulation of VEGF/VEGFR-2 under High-glucose Conditions. Exp. Med. 2021, 21, 389. [Google Scholar] [CrossRef]
- Lv, H.; Lv, G.; Chen, C.; Zong, Q.; Jiang, G.; Ye, D.; Cui, X.; He, Y.; Xiang, W.; Han, Q.; et al. NAD+ Metabolism Maintains Inducible PD-L1 Expression to Drive Tumor Immune Evasion. Cell Metab. 2021, 33, 110–127.e5. [Google Scholar] [CrossRef]
- Lin, T.-C. The Role of Visfatin in Cancer Proliferation, Angiogenesis, Metastasis, Drug Resistance and Clinical Prognosis. Cancer Manag. Res. 2019, 11, 3481–3491. [Google Scholar] [CrossRef] [Green Version]
- Ghaneialvar, H.; Shiri, S.; Kenarkoohi, A.; Fallah Vastani, Z.; Ahmadi, A.; Khorshidi, A.; Khooz, R. Comparison of Visfatin Levels in Patients with Breast Cancer and Endometrial Cancer with Healthy Individuals: A Systematic Review and Meta-analysis. Health Sci. Rep. 2022, 5, e895. [Google Scholar] [CrossRef]
- Zhao, Q.; Li, J.-Y.; Zhang, J.; Long, Y.-X.; Li, Y.-J.; Guo, X.-D.; Wei, M.-N.; Liu, W.-J. Role of Visfatin in Promoting Proliferation and Invasion of Colorectal Cancer Cells by Downregulating SDF-1/CXCR4-Mediated MiR-140-3p Expression. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 5367–5377. [Google Scholar] [CrossRef]
- Song, C.-Y.; Chang, S.L.-Y.; Lin, C.-Y.; Tsai, C.-H.; Yang, S.-Y.; Fong, Y.-C.; Huang, Y.-W.; Wang, S.-W.; Chen, W.-C.; Tang, C.-H. Visfatin-Induced Inhibition of MiR-1264 Facilitates PDGF-C Synthesis in Chondrosarcoma Cells and Enhances Endothelial Progenitor Cell Angiogenesis. Cells 2022, 11, 3470. [Google Scholar] [CrossRef]
- Shackelford, R.E.; Bui, M.M.; Coppola, D.; Hakam, A. Over-Expression of Nicotinamide Phosphoribosyltransferase in Ovarian Cancers. Int. J. Clin. Exp. Pathol. 2010, 3, 522–527. [Google Scholar]
- Montecucco, F.; Cea, M.; Bauer, I.; Soncini, D.; Caffa, I.; Lasiglie, D.; Nahimana, A.; Uccelli, A.; Bruzzone, S.; Nencioni, A. Nicotinamide Phosphoribosyltransferase (NAMPT) Inhibitors as Therapeutics: Rationales, Controversies, Clinical Experience. Curr. Drug Targets 2013, 14, 637–643. [Google Scholar] [CrossRef]
- Bruzzone, S.; Parenti, M.D.; Grozio, A.; Ballestrero, A.; Bauer, I.; Del Rio, A.; Nencioni, A. Rejuvenating Sirtuins: The Rise of a New Family of Cancer Drug Targets. Curr. Pharm. Des. 2013, 19, 614–623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bauer, I.; Grozio, A.; Lasigliè, D.; Basile, G.; Sturla, L.; Magnone, M.; Sociali, G.; Soncini, D.; Caffa, I.; Poggi, A.; et al. The NAD+-Dependent Histone Deacetylase SIRT6 Promotes Cytokine Production and Migration in Pancreatic Cancer Cells by Regulating Ca2+ Responses. J. Biol. Chem. 2012, 287, 40924–40937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malavasi, F.; Deaglio, S.; Damle, R.; Cutrona, G.; Ferrarini, M.; Chiorazzi, N. CD38 and Chronic Lymphocytic Leukemia: A Decade Later. Blood 2011, 118, 3470–3478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, D.; Chen, N.-N.; Cao, J.-M.; Sun, W.-P.; Zhou, Y.-M.; Li, C.-Y.; Wang, X.-X. BRCA1 as a Nicotinamide Adenine Dinucleotide (NAD)-Dependent Metabolic Switch in Ovarian Cancer. Cell Cycle 2014, 13, 2564–2571. [Google Scholar] [CrossRef]
- Franco, J.; Piacente, F.; Walter, M.; Fratta, S.; Ghanem, M.; Benzi, A.; Caffa, I.; Kurkin, A.V.; Altieri, A.; Herr, P.; et al. Structure-Based Identification and Biological Characterization of New NAPRT Inhibitors. Pharmaceuticals 2022, 15, 855. [Google Scholar] [CrossRef] [PubMed]
- Pramono, A.A.; Rather, G.M.; Herman, H.; Lestari, K.; Bertino, J.R. NAD- and NADPH-Contributing Enzymes as Therapeutic Targets in Cancer: An Overview. Biomolecules 2020, 10, 358. [Google Scholar] [CrossRef] [Green Version]
- Wei, Y.; Xiang, H.; Zhang, W. Review of Various NAMPT Inhibitors for the Treatment of Cancer. Front. Pharmacol. 2022, 13, 970553. [Google Scholar] [CrossRef]
- Hasmann, M.; Schemainda, I. FK866, a Highly Specific Noncompetitive Inhibitor of Nicotinamide Phosphoribosyltransferase, Represents a Novel Mechanism for Induction of Tumor Cell Apoptosis. Cancer Res. 2003, 63, 7436–7442. [Google Scholar]
- Tolstikov, V.; Nikolayev, A.; Dong, S.; Zhao, G.; Kuo, M.-S. Metabolomics Analysis of Metabolic Effects of Nicotinamide Phosphoribosyltransferase (NAMPT) Inhibition on Human Cancer Cells. PLoS ONE 2014, 9, e114019. [Google Scholar] [CrossRef]
- Sociali, G.; Raffaghello, L.; Magnone, M.; Zamporlini, F.; Emionite, L.; Sturla, L.; Bianchi, G.; Vigliarolo, T.; Nahimana, A.; Nencioni, A.; et al. Antitumor Effect of Combined NAMPT and CD73 Inhibition in an Ovarian Cancer Model. Oncotarget 2016, 7, 2968–2984. [Google Scholar] [CrossRef] [Green Version]
- Kudo, K.; Nomura, M.; Sakamoto, Y.; Ito, S.; Morita, M.; Kawai, M.; Yamashita, Y.; Ito, K.; Yamada, H.; Shima, H.; et al. Divergent Metabolic Responses Dictate Vulnerability to NAMPT Inhibition in Ovarian Cancer. FEBS Lett. 2020, 594, 1379–1388. [Google Scholar] [CrossRef]
- Nacarelli, T.; Fukumoto, T.; Zundell, J.A.; Fatkhutdinov, N.; Jean, S.; Cadungog, M.G.; Borowsky, M.E.; Zhang, R. NAMPT Inhibition Suppresses Cancer Stem-like Cells Associated with Therapy-Induced Senescence in Ovarian Cancer. Cancer Res. 2020, 80, 890–900. [Google Scholar] [CrossRef] [Green Version]
- Ding, S.; Madu, C.O.; Lu, Y. The Impact of Hormonal Imbalances Associated with Obesity on the Incidence of Endometrial Cancer in Postmenopausal Women. J. Cancer 2020, 11, 5456–5465. [Google Scholar] [CrossRef]
- Wang, Y.; Gao, C.; Zhang, Y.; Gao, J.; Teng, F.; Tian, W.; Yang, W.; Yan, Y.; Xue, F. Visfatin Stimulates Endometrial Cancer Cell Proliferation via Activation of PI3K/Akt and MAPK/ERK1/2 Signalling Pathways. Gynecol. Oncol. 2016, 143, 168–178. [Google Scholar] [CrossRef]
- Cymbaluk-Płoska, A.; Chudecka-Głaz, A.; Pius-Sadowska, E.; Sompolska-Rzechuła, A.; Machaliński, B.; Menkiszak, J. Circulating Serum Level of Visfatin in Patients with Endometrial Cancer. BioMed Res. Int. 2018, 2018, 8576179. [Google Scholar] [CrossRef] [Green Version]
- Luhn, P.; Dallal, C.M.; Weiss, J.M.; Black, A.; Huang, W.-Y.; Lacey, J.V.; Hayes, R.B.; Stanczyk, F.Z.; Wentzensen, N.; Brinton, L.A. Circulating Adipokine Levels and Endometrial Cancer Risk in the Prostate, Lung, Colorectal, and Ovarian Cancer Screening Trial. Cancer Epidemiol. Biomark. Prev. 2013, 22, 1304–1312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ilhan, T.T.; Kebapcilar, A.; Yilmaz, S.A.; Ilhan, T.; Kerimoglu, O.S.; Pekin, A.T.; Akyurek, F.; Unlu, A.; Celik, C. Relations of Serum Visfatin and Resistin Levels with Endometrial Cancer and Factors Associated with Its Prognosis. Asian Pac. J. Cancer Prev. 2015, 16, 4503–4508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michalczyk, K.; Niklas, N.; Rychlicka, M.; Cymbaluk-Płoska, A. The Influence of Biologically Active Substances Secreted by the Adipose Tissue on Endometrial Cancer. Diagnostics 2021, 11, 494. [Google Scholar] [CrossRef] [PubMed]
- Rachwalik, M.; Hurkacz, M.; Sienkiewicz-Oleszkiewicz, B.; Jasiński, M. Role of Resistin in Cardiovascular Diseases: Implications for Prevention and Treatment. Adv. Clin. Exp. Med. 2021, 30, 865–874. [Google Scholar] [CrossRef]
- Acquarone, E.; Monacelli, F.; Borghi, R.; Nencioni, A.; Odetti, P. Resistin: A Reappraisal. Mech. Ageing Dev. 2019, 178, 46–63. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Yang, Q.; Cai, D.; Guo, H.; Fang, J.; Cui, H.; Gou, L.; Deng, J.; Wang, Z.; Zuo, Z. Resistin, a Novel Host Defense Peptide of Innate Immunity. Front. Immunol. 2021, 12, 699807. [Google Scholar] [CrossRef] [PubMed]
- Sudan, S.K.; Deshmukh, S.K.; Poosarla, T.; Holliday, N.P.; Dyess, D.L.; Singh, A.P.; Singh, S. Resistin: An Inflammatory Cytokine with Multi-Faceted Roles in Cancer. Biochim. Biophys. Acta (BBA)-Rev. Cancer 2020, 1874, 188419. [Google Scholar] [CrossRef]
- Zieba, D.A.; Biernat, W.; Barć, J. Roles of Leptin and Resistin in Metabolism, Reproduction, and Leptin Resistance. Domest. Anim. Endocrinol. 2020, 73, 106472. [Google Scholar] [CrossRef]
- Codoñer-Franch, P.; Alonso-Iglesias, E. Resistin: Insulin Resistance to Malignancy. Clin. Chim. Acta 2015, 438, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Macchi, C.; Greco, M.F.; Botta, M.; Sperandeo, P.; Dongiovanni, P.; Valenti, L.; Cicero, A.F.G.; Borghi, C.; Lupo, M.G.; Romeo, S.; et al. Leptin, Resistin, and Proprotein Convertase Subtilisin/Kexin Type 9. Am. J. Pathol. 2020, 190, 2226–2236. [Google Scholar] [CrossRef]
- Estienne, A.; Bongrani, A.; Reverchon, M.; Ramé, C.; Ducluzeau, P.-H.; Froment, P.; Dupont, J. Involvement of Novel Adipokines, Chemerin, Visfatin, Resistin and Apelin in Reproductive Functions in Normal and Pathological Conditions in Humans and Animal Models. Int. J. Mech. Sci. 2019, 20, 4431. [Google Scholar] [CrossRef] [Green Version]
- Tripathi, D.; Kant, S.; Pandey, S.; Ehtesham, N.Z. Resistin in Metabolism, Inflammation, and Disease. FEBS J. 2020, 287, 3141–3149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parafiniuk, K.; Skiba, W.; Pawłowska, A.; Suszczyk, D.; Maciejczyk, A.; Wertel, I. The Role of the Adipokine Resistin in the Pathogenesis and Progression of Epithelial Ovarian Cancer. Biomedicines 2022, 10, 920. [Google Scholar] [CrossRef]
- Deb, A.; Deshmukh, B.; Ramteke, P.; Bhati, F.K.; Bhat, M.K. Resistin: A Journey from Metabolism to Cancer. Transl. Oncol. 2021, 14, 101178. [Google Scholar] [CrossRef] [PubMed]
- Qiu, L.; Zhang, G.-F.; Yu, L.; Wang, H.-Y.; Jia, X.-J.; Wang, T.-J. Novel Oncogenic and Chemoresistance-Inducing Functions of Resistin in Ovarian Cancer Cells Require MiRNAs-Mediated Induction of Epithelial-to-Mesenchymal Transition. Sci. Rep. 2018, 8, 12522. [Google Scholar] [CrossRef] [PubMed]
- Linkov, F.; Goughnour, S.L.; Edwards, R.P.; Lokshin, A.; Ramanathan, R.C.; Hamad, G.G.; McCloskey, C.; Bovbjerg, D.H. Endometrial Cancer Associated Biomarkers in Bariatric Surgery Candidates: Exploration of Racial Differences. Surg. Obes. Relat. Dis. 2017, 13, 862–868. [Google Scholar] [CrossRef]
- Tsai, H.-C.; Cheng, S.-P.; Han, C.-K.; Huang, Y.-L.; Wang, S.-W.; Lee, J.-J.; Lai, C.-T.; Fong, Y.-C.; Tang, C.-H. Resistin Enhances Angiogenesis in Osteosarcoma via the MAPK Signaling Pathway. Aging 2019, 11, 9767–9777. [Google Scholar] [CrossRef] [PubMed]
- Nergiz Avcıoğlu, S.; Yüksel, H. Adipocyte Related Peptides—Galanin and Resistin in Endometrioid Type Endometrium Cancer. Ginekol. Pol. 2022, 93, 941–947. [Google Scholar] [CrossRef]
- Ozgor, B.Y.; Iyibozkurt, C.; Bastu, E.; Berkman, S.; Yalcın, O.; Cakmakoglu, B.; Saygılı, H. Investigation of Resistin 420 and 62 Gene Polymorphism in Patients with Endometrial Cancer. Taiwan. J. Obstet. Gynecol. 2019, 58, 164–167. [Google Scholar] [CrossRef]
- Mughal, A.; O’Rourke, S.T. Vascular Effects of Apelin: Mechanisms and Therapeutic Potential. Pharmacol. Ther. 2018, 190, 139–147. [Google Scholar] [CrossRef]
- Chapman, F.A.; Nyimanu, D.; Maguire, J.J.; Davenport, A.P.; Newby, D.E.; Dhaun, N. The Therapeutic Potential of Apelin in Kidney Disease. Nat. Rev. Nephrol. 2021, 17, 840–853. [Google Scholar] [CrossRef]
- Girault-Sotias, P.-E.; Gerbier, R.; Flahault, A.; de Mota, N.; Llorens-Cortes, C. Apelin and Vasopressin: The Yin and Yang of Water Balance. Front. Endocrinol. 2021, 12, 735515. [Google Scholar] [CrossRef]
- Hu, G.; Wang, Z.; Zhang, R.; Sun, W.; Chen, X. The Role of Apelin/Apelin Receptor in Energy Metabolism and Water Homeostasis: A Comprehensive Narrative Review. Front. Physiol. 2021, 12, 632886. [Google Scholar] [CrossRef]
- Dravecká, I.; Figurová, J.; Lazúrová, I. Is Apelin a New Biomarker in Patients with Polycystic Ovary Syndrome? Physiol. Res. 2021, 70, S635–S641. [Google Scholar] [CrossRef] [PubMed]
- Dawid, M.; Mlyczyńska, E.; Jurek, M.; Respekta, N.; Pich, K.; Kurowska, P.; Gieras, W.; Milewicz, T.; Kotula-Balak, M.; Rak, A. Apelin, APJ, and ELABELA: Role in Placental Function, Pregnancy, and Foetal Development—An Overview. Cells 2021, 11, 99. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Cheng, H.; Adhikari, B.K.; Wang, S.; Yang, N.; Liu, W.; Sun, J.; Wang, Y. The Role of Apelin–APJ System in Diabetes and Obesity. Front. Endocrinol. 2022, 13, 820002. [Google Scholar] [CrossRef] [PubMed]
- Wahba, A.S.; Ibrahim, M.E.; Abo-elmatty, D.M.; Mehanna, E.T. Association of the Adipokines Chemerin, Apelin, Vaspin and Omentin and Their Functional Genetic Variants with Rheumatoid Arthritis. J. Pers. Med. 2021, 11, 976. [Google Scholar] [CrossRef]
- Liet, B.; Nys, N.; Siegfried, G. Elabela/Toddler: New Peptide with a Promising Future in Cancer Diagnostic and Therapy. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2021, 1868, 119065. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Yi, X.; Lu, C.; Wang, Y.; Xiao, Q.; Zhang, L.; Pang, Y.; Guan, X. Study Progression of Apelin/APJ Signaling and Apela in Different Types of Cancer. Front. Oncol. 2021, 11, 658253. [Google Scholar] [CrossRef]
- Uribesalgo, I.; Hoffmann, D.; Zhang, Y.; Kavirayani, A.; Lazovic, J.; Berta, J.; Novatchkova, M.; Pai, T.; Wimmer, R.A.; László, V.; et al. Apelin Inhibition Prevents Resistance and Metastasis Associated with Anti-angiogenic Therapy. EMBO Mol. Med. 2019, 11, e9266. [Google Scholar] [CrossRef]
- Masoumi, J.; Jafarzadeh, A.; Khorramdelazad, H.; Abbasloui, M.; Abdolalizadeh, J.; Jamali, N. Role of Apelin/APJ Axis in Cancer Development and Progression. Adv. Med. Sci. 2020, 65, 202–213. [Google Scholar] [CrossRef]
- Kurowska, P.; Barbe, A.; Różycka, M.; Chmielińska, J.; Dupont, J.; Rak, A. Apelin in Reproductive Physiology and Pathology of Different Species: A Critical Review. Int. J. Endocrinol. 2018, 2018, 9170480. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Li, Z.; Zhao, Q.; Chen, L. Roles of Apelin/APJ System in Cancer: Biomarker, Predictor, and Emerging Therapeutic Target. J. Cell. Physiol. 2022, 237, 3734–3751. [Google Scholar] [CrossRef]
- Yang, Y.; Lv, S.-Y.; Ye, W.; Zhang, L. Apelin/APJ System and Cancer. Clin. Chim. Acta 2016, 457, 112–116. [Google Scholar] [CrossRef]
- Hu, D.; Cui, Z.; Peng, W.; Wang, X.; Chen, Y.; Wu, X. Apelin Is Associated with Clinicopathological Parameters and Prognosis in Breast Cancer Patients. Arch. Gynecol. Obs. 2022, 306, 1185–1195. [Google Scholar] [CrossRef]
- Yang, L.; Li, Y.-L.; Li, X.-Q.; Zhang, Z. High Apelin Level Indicates a Poor Prognostic Factor in Muscle-Invasive Bladder Cancer. Dis. Mrk. 2019, 2019, 4586405. [Google Scholar] [CrossRef] [Green Version]
- Jaiprasart, P.; Dogra, S.; Neelakantan, D.; Devapatla, B.; Woo, S. Identification of Signature Genes Associated with Therapeutic Resistance to Anti-VEGF Therapy. Oncotarget 2020, 11, 99–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polak, K.; Czyzyk, A.; Simoncini, T.; Meczekalski, B. New Markers of Insulin Resistance in Polycystic Ovary Syndrome. J. Endocrinol. Investig. 2017, 40, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Ozkan, Z.S.; Cilgin, H.; Simsek, M.; Cobanoglu, B.; Ilhan, N. Investigation of Apelin Expression in Endometriosis. J. Reprod. Infertil. 2013, 14, 50–55. [Google Scholar] [PubMed]
- Neelakantan, D.; Dogra, S.; Devapatla, B.; Jaiprasart, P.; Mukashyaka, M.C.; Janknecht, R.; Dwivedi, S.K.D.; Bhattacharya, R.; Husain, S.; Ding, K.; et al. Multifunctional APJ Pathway Promotes Ovarian Cancer Progression and Metastasis. Mol. Cancer Res. 2019, 17, 1378–1390. [Google Scholar] [CrossRef] [Green Version]
- Altinkaya, S.O.; Nergiz, S.; Küçük, M.; Yüksel, H. Apelin Levels Are Higher in Obese Patients with Endometrial Cancer: Apelin Levels in Endometrial Cancer. J. Obs. Gynaecol. Res. 2015, 41, 294–300. [Google Scholar] [CrossRef]
- Chen, P.; Guo, Y.; Jia, L.; Wan, J.; He, T.; Fang, C.; Li, T. Interaction Between Functionally Activate Endometrial Microbiota and Host Gene Regulation in Endometrial Cancer. Front. Cell Dev. Biol. 2021, 9, 727286. [Google Scholar] [CrossRef] [PubMed]
- Nagpal, S.; Patel, S.; Jacobe, H.; DiSepio, D.; Ghosn, C.; Malhotra, M.; Teng, M.; Duvic, M.; Chandraratna, R.A.S. Tazarotene-Induced Gene 2 (TIG2), a Novel Retinoid-Responsive Gene in Skin. J. Investig. Dermatol. 1997, 109, 91–95. [Google Scholar] [CrossRef] [Green Version]
- Shin, W.J.; Pachynski, R.K. Chemerin Modulation of Tumor Growth: Potential Clinical Applications in Cancer. Discov. Med. 2018, 26, 31–37. [Google Scholar]
- Zabel, B.A.; Kwitniewski, M.; Banas, M.; Zabieglo, K.; Murzyn, K.; Cichy, J. Chemerin Regulation and Role in Host Defense. Am. J. Clin. Exp. Immunol. 2014, 3, 1–19. [Google Scholar]
- Ben Dhaou, C.; Mandi, K.; Frye, M.; Acheampong, A.; Radi, A.; De Becker, B.; Antoine, M.; Baeyens, N.; Wittamer, V.; Parmentier, M. Chemerin Regulates Normal Angiogenesis and Hypoxia-Driven Neovascularization. Angiogenesis 2022, 25, 159–179. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, N.; Naruse, K.; Kobayashi, Y.; Miyabe, M.; Saiki, T.; Enomoto, A.; Takahashi, M.; Matsubara, T. Chemerin Promotes Angiogenesis in Vivo. Physiol. Rep. 2018, 6, e13962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Y.; Liu, L. Role of Chemerin/ChemR23 Axis as an Emerging Therapeutic Perspective on Obesity-Related Vascular Dysfunction. J. Transl. Med. 2022, 20, 141. [Google Scholar] [CrossRef]
- Goralski, K.B.; McCarthy, T.C.; Hanniman, E.A.; Zabel, B.A.; Butcher, E.C.; Parlee, S.D.; Muruganandan, S.; Sinal, C.J. Chemerin, a Novel Adipokine That Regulates Adipogenesis and Adipocyte Metabolism. J. Biol. Chem. 2007, 282, 28175–28188. [Google Scholar] [CrossRef] [Green Version]
- Han, L.; Zhang, Y.; Wan, S.; Wei, Q.; Shang, W.; Huang, G.; Fang, P.; Min, W. Loss of Chemerin Triggers Bone Remodeling in Vivo and in Vitro. Mol. Metab. 2021, 53, 101322. [Google Scholar] [CrossRef] [PubMed]
- Issa, M.E.; Muruganandan, S.; Ernst, M.C.; Parlee, S.D.; Zabel, B.A.; Butcher, E.C.; Sinal, C.J.; Goralski, K.B. Chemokine-like Receptor 1 Regulates Skeletal Muscle Cell Myogenesis. Am. J. Physiol.-Cell Physiol. 2012, 302, C1621–C1631. [Google Scholar] [CrossRef] [Green Version]
- Karampela, I.; Christodoulatos, G.S.; Vallianou, N.; Tsilingiris, D.; Chrysanthopoulou, E.; Skyllas, G.; Antonakos, G.; Marinou, I.; Vogiatzakis, E.; Armaganidis, A.; et al. Circulating Chemerin and Its Kinetics May Be a Useful Diagnostic and Prognostic Biomarker in Critically Ill Patients with Sepsis: A Prospective Study. Biomolecules 2022, 12, 301. [Google Scholar] [CrossRef] [PubMed]
- Banas, M.; Zabieglo, K.; Kasetty, G.; Kapinska-Mrowiecka, M.; Borowczyk, J.; Drukala, J.; Murzyn, K.; Zabel, B.A.; Butcher, E.C.; Schroeder, J.M.; et al. Chemerin Is an Antimicrobial Agent in Human Epidermis. PLoS ONE 2013, 8, e58709. [Google Scholar] [CrossRef]
- Zegar, A.; Godlewska, U.; Kozłowska-Chmielewska, D.; Majewski, P.; Zabel, B.A.; Cichy, J. Chemerin-Derived Peptide Val66-Pro85 Is Effective in Limiting Methicillin-Resistant S. Aureus Skin Infection. Front. Microbiol. 2021, 12, 742610. [Google Scholar] [CrossRef] [PubMed]
- Ferland, D.J.; Garver, H.; Contreras, G.A.; Fink, G.D.; Watts, S.W. Chemerin Contributes to in Vivo Adipogenesis in a Location-Specific Manner. PLoS ONE 2020, 15, e0229251. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Leung, L.L.; Morser, J. Chemerin Forms: Their Generation and Activity. Biomedicines 2022, 10, 2018. [Google Scholar] [CrossRef]
- İnci, S.; Aksan, G.; Doğan, P. Chemerin as an Independent Predictor of Cardiovascular Event Risk. Ther. Adv. Endocrinol. 2016, 7, 57–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goralski, K.B.; Jackson, A.E.; McKeown, B.T.; Sinal, C.J. More Than an Adipokine: The Complex Roles of Chemerin Signaling in Cancer. Int. J. Mech. Sci. 2019, 20, 4778. [Google Scholar] [CrossRef] [Green Version]
- Banas, M.; Zegar, A.; Kwitniewski, M.; Zabieglo, K.; Marczynska, J.; Kapinska-Mrowiecka, M.; LaJevic, M.; Zabel, B.A.; Cichy, J. The Expression and Regulation of Chemerin in the Epidermis. PLoS ONE 2015, 10, e0117830. [Google Scholar] [CrossRef]
- Mattern, A.; Zellmann, T.; Beck-Sickinger, A.G. Processing, Signaling, and Physiological Function of Chemerin: Processing, Signaling, and Physiological Function of Chemerin. IUBMB Life 2014, 66, 19–26. [Google Scholar] [CrossRef]
- Treeck, O.; Buechler, C.; Ortmann, O. Chemerin and Cancer. Int. J. Mech. Sci. 2019, 20, 3750. [Google Scholar] [CrossRef] [Green Version]
- Helfer, G.; Wu, Q.-F. Chemerin: A Multifaceted Adipokine Involved in Metabolic Disorders. J. Endocrinol. 2018, 238, R79–R94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stojek, M. The Role of Chemerin in Human Disease. Postep. Hig. Med. Dosw. 2017, 71, 110–117. [Google Scholar] [CrossRef]
- Luo, X.; Gong, Y.; Cai, L.; Zhang, L.; Dong, X. Chemerin Regulates Autophagy to Participate in Polycystic Ovary Syndrome. J. Int. Med. Res. 2021, 49, 3000605211058376. [Google Scholar] [CrossRef] [PubMed]
- Fischer, T.F.; Beck-Sickinger, A.G. Chemerin—Exploring a Versatile Adipokine. Biol. Chem. 2022, 403, 625–642. [Google Scholar] [CrossRef] [PubMed]
- Buechler, C.; Feder, S.; Haberl, E.; Aslanidis, C. Chemerin Isoforms and Activity in Obesity. Int. J. Mech. Sci. 2019, 20, 1128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, M.; Yang, Y.; Huang, C.; Ge, L.; Xue, L.; Xiao, Z.; Xiao, T.; Zhao, H.; Ren, P.; Zhang, J.V. Chemerin: A Functional Adipokine in Reproductive Health and Diseases. Biomedicines 2022, 10, 1910. [Google Scholar] [CrossRef]
- Kennedy, A.J.; Davenport, A.P. International Union of Basic and Clinical Pharmacology CIII: Chemerin Receptors CMKLR1 (Chemerin1) and GPR1 (Chemerin2) Nomenclature, Pharmacology, and Function. Pharm. Rev. 2018, 70, 174–196. [Google Scholar] [CrossRef] [Green Version]
- Mariani, F.; Roncucci, L. Chemerin/ChemR23 Axis in Inflammation Onset and Resolution. Inflamm. Res. 2015, 64, 85–95. [Google Scholar] [CrossRef]
- Reverchon, M.; Ramé, C.; Dupont, J. La Chémérine: Une Adipocytokine pro-Inflammatoire Impliquée Dans La Fonction de Reproduction? Med. Sci. 2015, 31, 493–498. [Google Scholar] [CrossRef]
- De Henau, O.; Degroot, G.-N.; Imbault, V.; Robert, V.; De Poorter, C.; Mcheik, S.; Galés, C.; Parmentier, M.; Springael, J.-Y. Signaling Properties of Chemerin Receptors CMKLR1, GPR1 and CCRL2. PLoS ONE 2016, 11, e0164179. [Google Scholar] [CrossRef]
- Treeck, O.; Buechler, C. Chemerin Signaling in Cancer. Cancers 2020, 12, 3085. [Google Scholar] [CrossRef]
- El-Sagheer, G.; Gayyed, M.; Ahmad, A.; Abd El-Fattah, A.; Mohamed, M. Expression of Chemerin Correlates with a Poor Prognosis in Female Breast Cancer Patients. Breast Cancer Targets Ther. 2018, 10, 169–176. [Google Scholar] [CrossRef] [Green Version]
- Alkady, M.M.; Abdel-Messeih, P.L.; Nosseir, N.M. Assessment of Serum Levels of the Adipocytokine Chemerin in Colorectal Cancer Patients. J. Med. Biochem. 2018, 37, 313–319. [Google Scholar] [CrossRef]
- Yue, G.; An, Q.; Xu, X.; Jin, Z.; Ding, J.; Hu, Y.; Du, Q.; Xu, J.; Xie, R. The Role of Chemerin in Human Diseases. Cytokine 2023, 162, 156089. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Wu, W.K.K.; Liu, X.; To, K.-F.; Chen, G.G.; Yu, J.; Ng, E.K.W. Increased Serum Chemerin Level Promotes Cellular Invasiveness in Gastric Cancer: A Clinical and Experimental Study. Peptides 2014, 51, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Bozaoglu, K.; Curran, J.E.; Stocker, C.J.; Zaibi, M.S.; Segal, D.; Konstantopoulos, N.; Morrison, S.; Carless, M.; Dyer, T.D.; Cole, S.A.; et al. Chemerin, a Novel Adipokine in the Regulation of Angiogenesis. J. Clin. Endocrinol. Metab. 2010, 95, 2476–2485. [Google Scholar] [CrossRef] [Green Version]
- Gao, C.; Shi, J.; Zhang, J.; Li, Y.; Zhang, Y. Chemerin Promotes Proliferation and Migration of Ovarian Cancer Cells by Upregulating Expression of PD-L1. J. Zhejiang Univ. Sci. B 2022, 23, 164–170. [Google Scholar] [CrossRef]
- Wittamer, V.; Franssen, J.-D.; Vulcano, M.; Mirjolet, J.-F.; Le Poul, E.; Migeotte, I.; Brézillon, S.; Tyldesley, R.; Blanpain, C.; Detheux, M.; et al. Specific Recruitment of Antigen-Presenting Cells by Chemerin, a Novel Processed Ligand from Human Inflammatory Fluids. J. Exp. Med. 2003, 198, 977–985. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.; Rak, A.; Ptak, A. Bisphenol A and Its Derivatives Decrease Expression of Chemerin, Which Reverses Its Stimulatory Action in Ovarian Cancer Cells. Toxicol. Lett. 2018, 291, 61–69. [Google Scholar] [CrossRef]
- Schmitt, M.; Gallistl, J.; Schüler-Toprak, S.; Fritsch, J.; Buechler, C.; Ortmann, O.; Treeck, O. Anti-Tumoral Effect of Chemerin on Ovarian Cancer Cell Lines Mediated by Activation of Interferon Alpha Response. Cancers 2022, 14, 4108. [Google Scholar] [CrossRef]
- Dobrzyn, K.; Kiezun, M.; Zaobidna, E.; Kisielewska, K.; Rytelewska, E.; Gudelska, M.; Kopij, G.; Bors, K.; Szymanska, K.; Kaminska, B.; et al. The In Vitro Effect of Prostaglandin E2 and F2α on the Chemerin System in the Porcine Endometrium during Gestation. Int. J. Mech. Sci. 2020, 21, 5213. [Google Scholar] [CrossRef]
- Chen, P.; Jia, R.; Liu, Y.; Cao, M.; Zhou, L.; Zhao, Z. Progress of Adipokines in the Female Reproductive System: A Focus on Polycystic Ovary Syndrome. Front. Endocrinol. 2022, 13, 881684. [Google Scholar] [CrossRef]
- Guzel, E.C.; Celik, C.; Abali, R.; Kucukyalcin, V.; Celik, E.; Guzel, M.; Yilmaz, M. Omentin and Chemerin and Their Association with Obesity in Women with Polycystic Ovary Syndrome. Gynecol. Endocrinol. 2014, 30, 419–422. [Google Scholar] [CrossRef]
- Szydło, B.; Kiczmer, P.; Świętochowska, E.; Ostrowska, Z. Role of Omentin and Chemerin in Metabolic Syndrome and Tumor Diseases. Postep. Hig. Med. Dosw. 2016, 70, 844–849. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, T.; Watanabe-Kominato, K.; Takahashi, Y.; Kojima, M.; Watanabe, R. Adipose Tissue-Derived Omentin-1 Function and Regulation. In Comprehensive Physiology; Terjung, R., Ed.; Wiley: Hoboken, NJ, USA, 2017; pp. 765–781. ISBN 978-0-470-65071-4. [Google Scholar]
- Zhou, Y.; Zhang, B.; Hao, C.; Huang, X.; Li, X.; Huang, Y.; Luo, Z. Omentin-A Novel Adipokine in Respiratory Diseases. Int. J. Mech. Sci. 2017, 19, 73. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Zhao, X.; Chen, M. Autocrine Action of Adipokine Omentin-1 in the SW480 Colon Cancer Cell Line. Oncol. Lett. 2020, 19, 892–898. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; He, W.; Wang, W.; Zhou, D. Altered Circulating Levels of Adipokine Omentin-1 in Patients with Prostate Cancer. OncoTargets Ther. 2019, 12, 3313–3319. [Google Scholar] [CrossRef] [Green Version]
- Escoté, X.; Gómez-Zorita, S.; López-Yoldi, M.; Milton-Laskibar, I.; Fernández-Quintela, A.; Martínez, J.; Moreno-Aliaga, M.; Portillo, M. Role of Omentin, Vaspin, Cardiotrophin-1, TWEAK and NOV/CCN3 in Obesity and Diabetes Development. Int. J. Mech. Sci. 2017, 18, 1770. [Google Scholar] [CrossRef] [Green Version]
- Waluga, M.; Kukla, M.; Kotulski, R.; Zorniak, M.; Boryczka, G.; Kajor, M.; Ciupinska-Kajor, M.; Lekstan, A.; Olczyk, P.; Waluga, E. Omentin, Vaspin and Irisin in Chronic Liver Diseases. J. Physiol. Pharmacol. 2019, 70, 277–285. [Google Scholar] [CrossRef]
- Parida, S.; Siddharth, S.; Sharma, D. Role of Omentin in Obesity Paradox in Lung Cancer. Cancers 2021, 13, 275. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Huang, D.; Liu, X.; Wang, Y.; Liu, J.; Liu, F.; Yu, B. Omentin-1 Effects on Mesenchymal Stem Cells: Proliferation, Apoptosis, and Angiogenesis in Vitro. Stem Cell Res. 2017, 8, 224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Xue, J.; Hong, P. Relationships between Serum Omentin-1 Concentration, Body Composition and Physical Activity Levels in Older Women. Medicine 2021, 100, e25020. [Google Scholar] [CrossRef]
- He, H.; Wu, S.; Hao, J.; Wang, L.; Ai, K.; Zhu, X.; Xu, R.; Zhao, X.; Wang, Y.; Zhong, Z. Serum Omentin-1 Level in Patients with Benign Prostatic Hyperplasia. BMC Urol. 2020, 20, 52. [Google Scholar] [CrossRef] [PubMed]
- Khadem Ansari, M.H.; Gholamnejad, M.; Meghrazi, K.; Khalkhali, H.R. Association of Circulating Omentin-1 Level with Lung Cancer in Smokers. Med. J. Islam. Repub. Iran 2018, 779–783. [Google Scholar] [CrossRef] [PubMed]
- Tahmasebpour, N.; Hosseinpour Feizi, M.A.; Ziamajidi, N.; Pouladi, N.; Montazeri, V.; Farhadian, M.; Abbasalipourkabir, R. Association of Omentin-1 with Oxidative Stress and Clinical Significances in Patients with Breast Cancer. Adv. Pharm. Bull. 2019, 10, 106–113. [Google Scholar] [CrossRef]
- Abas, A.-S.M.; Sherif, M.H.; Elmoneam Farag, S.A. Diagnostic and Prognostic Role of Serum Omentin and NGAL Levels in Egyptian Breast Cancer Patients. Int. J. Breast Cancer 2022, 2022, 5971981. [Google Scholar] [CrossRef]
- Feng, Z.; Sun, H.; Liu, P.; Shi, W.; Han, W.; Ma, L. Analysis of the Expression of Plasma Omentin-1 Level in Colorectal Cancer and Its Correlation with Prognosis. Transl. Cancer Res. TCR 2020, 9, 6479–6486. [Google Scholar] [CrossRef]
- Au-Yeung, C.-L.; Yeung, T.-L.; Achreja, A.; Zhao, H.; Yip, K.-P.; Kwan, S.-Y.; Onstad, M.; Sheng, J.; Zhu, Y.; Baluya, D.L.; et al. ITLN1 Modulates Invasive Potential and Metabolic Reprogramming of Ovarian Cancer Cells in Omental Microenvironment. Nat. Commun. 2020, 11, 3546. [Google Scholar] [CrossRef]
- Paval, D.R.; Di Virgilio, T.G.; Skipworth, R.J.E.; Gallagher, I.J. The Emerging Role of Intelectin-1 in Cancer. Front. Oncol. 2022, 12, 767859. [Google Scholar] [CrossRef]
- Cymbaluk-Płoska, A.; Chudecka-Głaz, A.; Jagodzińska, A.; Pius-Sadowska, E.; Sompolska-Rzechuła, A.; Machaliński, B.; Menkiszak, J. Evaluation of Biologically Active Substances Promoting the Development of or Protecting against Endometrial Cancer. OncoTargets Ther. 2018, 11, 1363–1372. [Google Scholar] [CrossRef] [Green Version]
- Soliman, P.T.; Zhang, Q.; Broaddus, R.R.; Westin, S.N.; Iglesias, D.; Munsell, M.F.; Schmandt, R.; Yates, M.; Ramondetta, L.; Lu, K.H. Prospective Evaluation of the Molecular Effects of Metformin on the Endometrium in Women with Newly Diagnosed Endometrial Cancer: A Window of Opportunity Study. Gynecol. Oncol. 2016, 143, 466–471. [Google Scholar] [CrossRef] [Green Version]
- Yates, M.S.; Coletta, A.M.; Zhang, Q.; Schmandt, R.E.; Medepalli, M.; Nebgen, D.; Soletsky, B.; Milbourne, A.; Levy, E.; Fellman, B.; et al. Prospective Randomized Biomarker Study of Metformin and Lifestyle Intervention for Prevention in Obese Women at Increased Risk for Endometrial Cancer. Cancer Prev. Res. 2018, 11, 477–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kastl, S.P.; Katsaros, K.M.; Krychtiuk, K.A.; Jägersberger, G.; Kaun, C.; Huber, K.; Wojta, J.; Speidl, W.S. The Adipokine Vaspin Is Associated with Decreased Coronary In-Stent Restenosis in Vivo and Inhibits Migration of Human Coronary Smooth Muscle Cells in Vitro. PLoS ONE 2020, 15, e0232483. [Google Scholar] [CrossRef]
- Liu, P.; Li, G.; Wu, J.; Zhou, X.; Wang, L.; Han, W.; Lv, Y.; Sun, C. Vaspin Promotes 3T3-L1 Preadipocyte Differentiation. Exp. Biol. Med. 2015, 240, 1520–1527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurowska, P.; Mlyczyńska, E.; Dawid, M.; Dupont, J.; Rak, A. Role of Vaspin in Porcine Ovary: Effect on Signaling Pathways and Steroid Synthesis via GRP78 Receptor and Protein Kinase A. Biol. Reprod. 2020, 102, 1290–1305. [Google Scholar] [CrossRef] [PubMed]
- Suliga, E.; Kozieł, D.; Cieśla, E.; Rębak, D.; Wawszczak, M.; Adamus-Białek, W.; Naszydłowska, E.; Piechowska, A.; Głuszek, S. Associations between Vaspin Rs2236242 Gene Polymorphism, Walking Time and the Risk of Metabolic Syndrome. Balk. J. Med. Genet. 2019, 22, 41–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurowska, P.; Mlyczyńska, E.; Dawid, M.; Jurek, M.; Klimczyk, D.; Dupont, J.; Rak, A. Review: Vaspin (SERPINA12) Expression and Function in Endocrine Cells. Cells 2021, 10, 1710. [Google Scholar] [CrossRef]
- Dimova, R.; Tankova, T. The Role of Vaspin in the Development of Metabolic and Glucose Tolerance Disorders and Atherosclerosis. BioMed Res. Int. 2015, 2015, 823481. [Google Scholar] [CrossRef]
- Li, Q.; Chen, R.; Moriya, J.; Yamakawa, J.; Sumino, H.; Kanda, T.; Takahashi, T. A Novel Adipocytokine, Visceral Adipose Tissue-Derived Serine Protease Inhibitor (Vaspin), and Obesity. J. Int. Med. Res. 2008, 36, 625–629. [Google Scholar] [CrossRef]
- Feng, R.; Li, Y.; Wang, C.; Luo, C.; Liu, L.; Chuo, F.; Li, Q.; Sun, C. Higher Vaspin Levels in Subjects with Obesity and Type 2 Diabetes Mellitus: A Meta-Analysis. Diabetes Res. Clin. Pract. 2014, 106, 88–94. [Google Scholar] [CrossRef]
- Yang, H.; Huang, Y.; Gai, C.; Chai, G.; Lee, S. Serum Vaspin Levels Are Positively Associated with Diabetic Retinopathy in Patients with Type 2 Diabetes Mellitus. J. Diabetes Investig. 2021, 12, 566–573. [Google Scholar] [CrossRef]
- Kan, J.-Y.; Lee, Y.-C.; Lin, Y.-D.; Ho, W.-Y.; Moi, S.-H. Effect of Baseline Characteristics and Tumor Burden on Vaspin Expression and Progressive Disease in Operable Colorectal Cancer. Diagnostics 2020, 10, 801. [Google Scholar] [CrossRef]
- Cao, X.; Chen, X.; Yang, K.; Wang, Y.; Liang, M.; Fei, Y.; Tang, J. Vaspin Accelerates the Proliferation, Invasion and Metastasis of Triple-Negative Breast Cancer through MiR -33a-5p/ABHD2. Cancer Med. 2023, 12, 4530–4542. [Google Scholar] [CrossRef]
- Sun, Y.; Zhu, S.; Wu, Z.; Huang, Y.; Liu, C.; Tang, S.; Wei, L. Elevated Serum Visfatin Levels Are Associated with Poor Prognosis of Hepatocellular Carcinoma. Oncotarget 2017, 8, 23427–23435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, T.-J.; Han, L.-H.; Cong, R.-S.; Liang, J. Caspase Family Proteases and Apoptosis. Acta Biochim. Biophys. Sin. 2005, 37, 719–727. [Google Scholar] [CrossRef] [Green Version]
- Van Opdenbosch, N.; Lamkanfi, M. Caspases in Cell Death, Inflammation, and Disease. Immunity 2019, 50, 1352–1364. [Google Scholar] [CrossRef] [PubMed]
- Kesavardhana, S.; Malireddi, R.K.S.; Kanneganti, T.-D. Caspases in Cell Death, Inflammation, and Pyroptosis. Annu. Rev. Immunol. 2020, 38, 567–595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Magaña, A.; Blanco, F.J. Human PCNA Structure, Function and Interactions. Biomolecules 2020, 10, 570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boehm, E.M.; Gildenberg, M.S.; Washington, M.T. The Many Roles of PCNA in Eukaryotic DNA Replication. In The Enzymes; Elsevier: Amsterdam, The Netherlands, 2016; Volume 39, pp. 231–254. ISBN 978-0-12-804735-4. [Google Scholar]
- Erdogan, S.; Sezer, S.; Baser, E.; Gun-Eryilmaz, O.; Gungor, T.; Uysal, S.; Yilmaz, F.M. Evaluating Vaspin and Adiponectin in Postmenopausal Women with Endometrial Cancer. Endocr.-Relat. Cancer 2013, 20, 669–675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozłowski, M.; Pietrzyk, D.; Rychlicka, M.; Piotrowska, K.; Nowak, K.; Kwiatkowski, S.; Cymbaluk-Płoska, A. Evaluation of Tissue Expression of Vaspin and Serum Vaspin Concentration as a Prognostic and Risk Factor in Endometrial Cancer. Cells 2022, 11, 3196. [Google Scholar] [CrossRef] [PubMed]
- Naimo, G.D.; Gelsomino, L.; Catalano, S.; Mauro, L.; Andò, S. Interfering Role of ERα on Adiponectin Action in Breast Cancer. Front. Endocrinol. 2020, 11, 66. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.G.; Kim, E.O.; Jeong, B.R.; Min, Y.J.; Park, J.W.; Kim, E.S.; Namgoong, I.S.; Kim, Y.I.; Lee, B.J. Visfatin Stimulates Proliferation of MCF-7 Human Breast Cancer Cells. Mol. Cells 2010, 30, 341–345. [Google Scholar] [CrossRef]
- Wang, N.; Wang, Q.; Feng, Y.; Shang, W.; Cai, M. Overexpression of Chemerin Was Associated with Tumor Angiogenesis and Poor Clinical Outcome in Squamous Cell Carcinoma of the Oral Tongue. Clin. Oral Investig. 2014, 18, 997–1004. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Other Kinds of Cancer | Ovarian Cancer | Endometrial Cancer | ||||
|---|---|---|---|---|---|---|
| Leptin |  | increase in tumor mass, facilitated metastatic ability, effect the process of angiogenesis, prevention of apoptosis of tumor cells [101,107,108,109,110] | | stimulation of cancer cell proliferation, inhibition of apoptosis, promotion of malignant cancer phenotypes, reduced survival rate [106,111,112] | | metastatic capacity, proliferation of cancer cells, induction of chemoresistance [30,41,105] |
| Adiponectin |  | inhibition of cell cycle, blocking angiogenesis, induction of apoptosis [118,119,130,131,295] | | inhibition of angiogenesis, growth and proliferation, invasion, migration, formation of tumor metastases [62,108,114,127,133,134] | | increase in cancer risk and stage [114,131,137] |
| Visfatin | | increase in risk of cancer development, shortened survival time, proliferation of cancer cells, metastasis formation [151,152,296] | | promotion of process of carcinogenesis, stimulation of process of angiogenesis, promotion of metastases formation [155,156,157,161,162] | | proliferative, proangiogenic effects, facilitated ability to form metastases [41,169,171] |
| Resistin | | facilitation of cancer growth and development, increase in cancer stage, metastatic ability, increase in aggressiveness [53,54,94,162] | | ability to promote and progress cancer, metastatic formation, angiogenesis promotion, induction of chemoresistance [53,178,184,185] | | facilitation of growth and development of cancer, ability to metastasize, increase in stage of progression, decrease in survival rate [41,173,178,187,188] |
| Apelin | | advanced stage of cancer, presence of metastases, induction of angiogenesis, poor prognosis [200,204,205,206] | | survivability, proliferation of cancer cells, metastasis formation, shortened survival time [203,208,211] | | increase in cancer risk in women with obesity [205,212,213] |
| Chemerin |  | migration of immune system cells toward the tumor, intracellular signaling, facilitated angiogenesis [215,218,229,245,246,247,248,297] | | effect on cancer cell adhesion, metastatic capacity [229,249,250] | no data available | |
| Omentin |  | stimulation of cancer growth and development, metastasis formation, inhibition of proliferation, induction of apoptosis [268,269,270] | | proliferation of tumor cells, metastasis formation [268,271,272] | | increase in stage of cancer [273,274,275] |
| Vaspin | | increase in degree of malignancy of cancer, decrease in survival time, formation of metastasis [44,287] | | stimulation of process of angiogenesis, limiting the process of apoptosis [203,278,280] | | increase in risk of developing cancer, increase in stage of cancer [171,174,293,294] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stępień, S.; Olczyk, P.; Gola, J.; Komosińska-Vassev, K.; Mielczarek-Palacz, A. The Role of Selected Adipocytokines in Ovarian Cancer and Endometrial Cancer. Cells 2023, 12, 1118. https://doi.org/10.3390/cells12081118
Stępień S, Olczyk P, Gola J, Komosińska-Vassev K, Mielczarek-Palacz A. The Role of Selected Adipocytokines in Ovarian Cancer and Endometrial Cancer. Cells. 2023; 12(8):1118. https://doi.org/10.3390/cells12081118
Chicago/Turabian StyleStępień, Sebastian, Paweł Olczyk, Joanna Gola, Katarzyna Komosińska-Vassev, and Aleksandra Mielczarek-Palacz. 2023. "The Role of Selected Adipocytokines in Ovarian Cancer and Endometrial Cancer" Cells 12, no. 8: 1118. https://doi.org/10.3390/cells12081118